
Abstract
This paper describes two new species of lizards of the infraorder Anguimorpha from the Aptian-Albian of Mongolia. One of these, Paradorsetisaurus postumus gen. et sp. nov., is the youngest in the family Dorsetisauridae, while the other, Xenostius futilus gen. et sp. nov. is the earliest fossil find within the family Xenosauridae s. str. The paper also includes remarks on the importance of the fossil finds from Mongolia for the reconstruction of lizard diversity in Central Asia up to the beginning of the late Cretaceous.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
The Khoobur (Hobur, Höövör, Khovboor, Guchin Us) terrestrial vertebrate locality is located in Central Mongolia (Guchin basin, Övörkhangai aimag). It is composed of Aptian-Albian fluvial and fluvial-lacustrine sandstones and shales. Tetrapod fossils from Khoobur (mammals, dinosaurs, pterosaurs, crocodiles, turtles, lizards) in the collections of the A.A. Borissiak Paleontological Institute (PIN) mainly come from screen-washed fossil-bearing layers.
Fieldwork was carried out by several missions of the Soviet-Mongolian Expedition, mostly between 1969 and 1972. Most of the fossils within the collection are lizards.
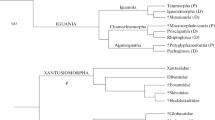
Lizards from Khoobur are represented by numerous (hundreds of specimens) isolated skeletal, cranial, mandibular elements, which are difficult to identify. However, already the first and largely preliminary identifications (Alifanov 1993, 2000a, 2000b) showed a large number of lizard families (Fig. 1), making Khoobur the richest assemblage between the Middle Jurassic and late Early Cretaceous. The assemblage also has key importance for the elucidation of the details of early diversification within the studied group.

Family composition (compiled from several sources) and faunistic structure of the lizard assemblage from the Khoobur locality (Mongolia, Övörkhangai aimag; early Cretaceous, Upper Aptian–Lower Albian, Hühteeg Horizon).
Some lizards from Khoobur have been studied in more detail. Within Gekkonidae s.l. (Gekkota), we have described the earliest representative, Hoburogekko suchanovi (Alifanov, 1989, Daza et al., 2012). The family Hodzhakuliidae Alifanov, 1993 (Scincomorpha) is represented by Hodzhakulia magna Nessov, 1985, previously described from the Late Albian of Uzbekistan, as well as by new species: Carnoscincus eublepharus, Bagaluus primigenius and Platynotoides altidentatus (Alifanov, 2016). We have also published descriptions of material attributed to the Temuijiniidae family (Temuijiniidae gen. indet., Alifanov, 2018) belonging to the microorder Iguanomorpha (Iguania).
This article continues our research on the diversity of lizards from Khoobur. This time it is focused on fossils from the infraorder Anguimorpha, in particular the families Dorsetisauridae and Xenosauridae. The systematic description is preceded by a summary of the history of views regarding their classification, phylogeny, and geographic distribution. In the closing section of the article we discuss details of the formation of lizard diversity in Central Asia during the Jurassic and Early Cretaceous.
CURRENT ISSUES IN THE RESEARCH OF DORSETISAURIDAE AND XENOSAURIDAE
The family Dorsetisauridae was established by P. Hoffstetter (1967) for Dorsetisaurus purbeckensis that he described from cranial and mandibular fragments from the Purbeck Formation (Tithonian–Neocomian) of England. The species has osteodermal sculpture on skull roof bones in the form of mid-sized scutes, is characterized by pineal opening located within the parietal, and the small number of teeth with a high and lancet-shaped apical part (Figs. 2a–2c). Later, representatives of the family were found in the Late Jurassic of the United States (Prothero and Estes, 1980) and Portugal (Seiffert, 1973, Broschinski, 2000), as well as in the Early Cretaceous of Mongolia (Alifanov, 1993, 2000a, 2000b).
Originally, Dorsetisauridae were either placed in the Anguoidea (Hoffstetter, 1967), or between the modern Xenosauridae s.l. and the Anguidae (Estes, 1983). The type species was seen as a basal anguimorph (Evans et al., 2006) or as a basal platynotan (Conrad, 2008). However, its affiliation with the Anguimorpha has not been proven so far because of fragmentary material available, which makes any hypotheses regarding its relationships tentative. It is as a tentative suggestion that we previously thought possible a close phylogenetic link between dorsetisaurids and Xenosauridae s.l. (Alifanov, 2000a, 2012).
The discovery of Kuwajimalla kagaensis Evans et Manabe, 2008 from the Early Cretaceous of Japan (Ishikawa) sheds new light on Dorsetisauridae. This species, described from fragments of the skull and of the mandible with teeth having a lancet-shaped apex was placed, in the original description, within the cladistic taxon Borioteiioidea Nydam et al., 2007. This taxon aggregates what we previously had described (Alifanov, 2000a, 2012) as the Late Cretaceous microorders Polyglyphanodontia (Agamognatha) and Macrocephalosauria (Chameleomorpha). It is worth pointing out that none of their representatives have been found in Central Asia. Besides that, K. kagaensis does not have subpleurodont mandibular teeth which are typical for Polyglyphanodontia and Macrocephalosauria. All of this makes this original reconstruction of phylogenetic affinity doubtful. It is also difficult to agree with the interpretation, given in the original description, of serrate-dentate edges of tooth apices as multiple cusps. On the other hand, lancet-shaped teeth, eupleurodonty (transitional condition between subpleurodonty and hyperpleurodonty) and the presence of sockets for tooth bases (Figs. 2d, 2e) admit an interpretation of affinity of K. kagaensis with Dorsetisauridae.

Lizards of the family Dorsetisauridae (Anguimorpha): (a–c) Dorsetisaurus purbeckensis Hoffstetter, 1967 (Hoffstetter, 1967, text-figs. 10A, 11C): (a, b) right maxilla articulated with the prefrontal and lacrymal in labial (a) and ventral view (b); BMNH R8248, (c) reconstructed left ramus of the dentary in labial view; UK, Tithonian–lower Neocomian; (d, e) Kuwajimalla kagaensis Evans et Manabe, 2008 (from Evans and Manabe, 2008, text-figs. 4B, 5A), right dentary (d) in labial view (SBEI 1538) and dentary teeth (e) in lingual view (SBEI 557); Japan, Barremian–Aptian; (f) Chometokadmon fitzingeri Costa, 1864; holotype MPN 539 (from Evans et al., 2006, text-fig. 2B), reconstructed skull in dorsal view; Italy, early Cretaceous; (g) Paradorsetisaurus postumus gen. et sp. nov.; holotype PIN 3334/517, right maxilla in labial view; Mongolia, Övörkhangai aimag, Khoobur locality; early Cretaceous, Upper Aptian–Lower Albian, Hühteeg Horizon. Scale 5 mm for (a–c), (f), and (g); 1 mm for (d) and (e).
In discussing Dorsetisauridae, it is interesting to look at the data on Chometokadmon fitzingeri Costa, 1864 (Fig. 2f). The structure of the frontal and the parietal in Ch. fitzingeri is identical to Dorsetisaurus purbeckensis Hoffstetter, 1967. In addition, C. fitzingeri has free osteoderms in the head region, similar to Xenosauridae s.l. (Bever et al., 2005). However, S. Evans et al. (2006) noted the generalization of Ch. fitzingeri, relating it, besides D. purbeckensis, to Parviraptor Evans, 1994 from the Late Jurassic and Neocomian of the United States and Great Britain. It is remarkable that this latter taxon, known from isolated skeletal fragments, has been interpreted quite differently, e.g., as related to snakes (Caldwell et al., 2015). We consider Parviraptor as representative of the Mosasauria, which are characterized by sharp teeth as well as by a zygosphenic-zyganthral vertebral articulation.
Judging from the details of skull structure, the mandible, and the tooth system, the embryos from the Early Cretaceous of Thailand found in hard shell (unusual for lizards) can also be attributed to the Dorsetisauridae, rather than the Platynota (varanoid lizards are not known in the Early Cretaceous), as was suggested in the original description (Fernandez et al., 2015). Our attribution takes into account the skull reconstruction in the cited paper, which shows the maxillary participating in the suborbital fenestra border (not found in varanoids), the structure of frontals (wide, paired and sculptured), the presence of a rostral process of the coronoid, a deep notch of the posterior margin of the generally short dentaries and flattened apices of the tooth crown.
On the other hand, the attribution of Changetisaurus estesi Nessov, 1992 from the Middle Jurassic of Kyrgyzstan to dorsetisaurids is questionable because of a well-developed angular process and rectangular osteoderms. This condition is typical both for the fossil Paramacellodidae (Cordyloidea, Scincomorpha), e.g., Sharovisaurus karatauensis Hecht et Hecht, 1984 from the Middle Jurassic of Kazakhstan, and for Parmeosaurus scutatus Gao et Hou, 2000 from the late Cretaceous of Mongolia.
Unlike the dorsetisaurids, Xenosauridae s.l., represented by the modern genera Xenosaurus (Mexico) and Shinisaurus (China), undoubtedly belong to the Anguimorpha. They were historically seen as related to the Anguidae within the microorder Diploglossa, the sister taxon to the Platynota (Camp, 1923; McDowell and Bogert, 1954). Cladistic analysis places xenosaurids among the basal Anguimorpha (Gauthier et al., 2012) or Platynota (Conrad, 2008). The platynotan hypothesis concerns Shinisaurus and related fossil forms. We consider it to be in contradiction with the key morphological features of the taxa being compared.
It should be pointed out that Shinisaurus, similarly to all the other Diploglossa (except Anniella: Alifanov, 2000a, 2012) is characterized by the diploglossopalatine condition (the absence of a connection between the lateral processes of ectopterygoid and palatine bones which doesn’t form the margin of the suborbital fenestra). This type can be contrasted with another type of the platynotopalatine condition, where maxillaries are isolated from the suborbital fenestrae by a contact of the lateral process of palatine and the ectopterygoid. Our data shows that the two conditions can not be reduced to one, as they form the different conditions of two different branches of semogenetic tree and one cannot be derived from the other directly, without intermediate stages (Alifanov, 2012, 2016).
Considering the issues and data just discussed, we should draw attention to tooth structure in Shinisaurus and Xenosaurus. In the former, the central cusp has indistinct symmetric shoulders (Conrad, 2004, text-figs. 17A, 17C), whereas in the latter the cusp has a rostral arms that makes the tooth look asymmetrical. If Xenosauridae s.l. are related to the Dorsetisauridae, then we have to deal with something that has not been addressed in previous work, including our own (Figs. 3a, 3b). Namely, the two modern forms must then stand in different relationship to the fossil family. In particular, Xenosaurus appears morphologically isolated within the group of a comparison. Based on this fact, it is possible to talk about two modern xenosaurid forms in different families (Fig. 3c), a suggestion previously made more than once (Bever et al., 2005; Conrad, 2008).

Proposed phylogenetic relationships of the families Dorsetisauridae and Xenosauridae (Anguimorpha) according to different authors: (a) S. Evans et al., 2006, (b) V.R. Alifanov, 2012, (c) this study.
A group sometimes synonymized with Xenosauridae s.l. are the late Cretaceous Carusiidae Borsuk-Białynicka, 1987 or its type genus (Gao and Norell, 1998, 2000; Conrad, 2008). Such attribution is contradicted, however, by the structure of the dentaries. Carusiids have scincomorph dentaries with the surangular process either incipient or absent, the angular process wide at the base and often lengthened, the coronoid process overlapping the lateral surface of the labial and/or the dorsal process of the coronoid. Scincomorph morphology of the dentaries was discussed in the first description of the group (Borsuk-Białynicka, 1985, 1987). Recent research places it either basally within the Scincoidea (Gauthier et al., 2012) or as one of the families of the superfamily Ardeosauroidea (Alifanov, 2012, 2016). The members of the type family of this last taxon are usually seen, in cladistic analyses, as related to the Gekkota (e.g., Conrad, 2008; Gauthier et al., 2012).
In North America, Xenosauridae s.str. are represented by the fossil genera Exostinus Cope, 1873 (late Cretaceous–early Oligocene, includes two species, possibly not monophyletic, see Bhullar, 2011) and Restes Gauthier, 1982 (late Paleocene). The oldest fossils here are Cenomanian in age (Nydam, 2013). In Asia, the family is represented by Oxia karakalpakensis Nessov, 1985 (Fig. 4a) from the late Albian of Uzbekistan (Gao and Nessov, 1998). The species is described from a dentary carrying blunted teeth and having a posterior region typical for the anguimorphs: the coronoid process is well-developed; the surangular process and the angular process do not reach occipitally beyond the center of the coronoid.

Lizards of the family Xenosauridae (Anguimorpha): (a) Oxia karakalpakensis Nessov, 1985; holotype CMGE 28/12000 (from Gao and Nessov, 1998, fig. 3, in partim), left dentary in lingual view; Uzbekistan, Upper Aptian; (b) “Xenosauridae” gen. indet. (?Ardeosauridae fam. indet.); PIN 3334/515 (Alifanov, 2000, text-fig. 28d), posterior region of the left dentary in labial view; (c) Xenostius futilus gen. et sp. nov. (Xenosauridae, Anguimorpha), holotype PIN 3334/522, left dentary in lingual view; Mongolia, Övörkhangai aimag, Khoobur locality; early Cretaceous, Upper Aptian–Lower Albian, Hühteeg Horizon. Scale 3 mm for (a), 1 mm for (b) and (c).
The discovery of Oxia karakalpakensis Nessov, 1985 is evidence of an early origin (earlier than late Cretaceous) of xenosaurids. The hypothesis of Central Asian origin must be supported with older fossils. We previously considered PIN 3334/515 from Khoobur (Fig. 4b) as a possible candidate (Alifanov, 2000a). However, a reexamination of the specimen, which is a fragment of the dentary with teeth, showed that the posterior region, which, as noted previously, is diagnostic, is missing but resembles the dentary structure in the Anguimorpha. It likely belongs to a scincomorph. However, xenosaurids are found in the Khoobur assemblage, as shown by specimens in this article.
The origin of the Shinisauridae the distribution of which are now confined to a small area in Southern China and Northern Vietnam, is enigmatic. Its wider distribution in the past is attested by fossil finds in the Eocene of North America (Conrad, 2006) and the Miocene of Europe (Klembara, 2008). Apparently, the group originated in one of the three areas. We believe that its most likely origin is Cretaceous Europe, from which it subsequently migrated into Southwestern Asia in post-Mesozoic times either through North America and Central Asia or through Asia Minor and the Indian subcontinent. The latter may have been originally zoogeographically related with Europe rather than with the southern continents: Kalandadze and Rautian (1992; 1997).
SYSTEMATIC PALEONTOLOGY
-
Order LACERTILIA
-
Infraorder ANGUIMORPHA
-
Microorder DIPLOGLOSSA
-
Superfamily Xenosauroidea Cope, 1868
Diagnosis. Diploglossian lizards characterized by the presence of free scutelike osteoderms in the head region and their absence in the trunk region.
Composition. Xenosauridae Cope, 1868; early Cretaceous of Uzbekistan and Mongolia, late Cretaceous to Modern, North America; Dorsetisauridae Hoffstetter, 1967; late Jurassic of North America, Tithonian to early Neocomian of Europe, early Cretaceous of Central Asia; Shinisauridae Ahl, 1930; Eocene of North America, early Miocene of Europe, Modern S. China and N. Vietnam.
Remarks. As noted above, anguimorphs of the microorder Diploglossa, here taken to circumscribe the superfamilies Xenosauroidea and Anguoidea, are characterized by a diploglossopalatine type of the palate fenestration. Superfamilies are distinguished according to the degree to which osteoderms are developed. In Xenosauridae these are concentrated in the head region, whereas in Anguoidea they extend over the whole body forming armour. The circumscription as well as the phylogenetic affinities of the superfamily Anguoidea is in need of a revision, just as the Xenosauridae s. str. The revision should include the fossil Glyptosauridae, as well as the living Anguidae (this latter group may have to be split into two to three families). Based on palatal morphology, the family Anniellidae forms part of the Platynota, not of the Diploglossa, as is usually assumed. Within the platynotan, the Anniellidae share some cranial characters with the Lanthanotidae to which it is likely related (Alifanov, 2012).
-
Family Dorsetisauridae Hoffstetter, 1967
Type genus. Dorsetisaurus Hoffstetter, 1967; late Jurassic of North America, late Jurassic to early Cretaceous of Europe.
Diagnosis. Nares openings elongate. Frontals paired. Lateral processes of the postfrontals are bifurcated for articulation with the medial process of the postorbitals. Osteodermal sculpture of skull roof bones is of the medium-sized scutes type. Tooth attachment eupleurodont or subpleurodont. Tooth crown lancet-shaped.
Composition. In addition to the type genus, genera: Chometokadmon Costa, 1864; early Cretaceous of Italy; Kuwajimalla Evans et Manabe, 2008; early Cretaceous of Japan; Paradorsetisaurus gen. nov.; early Cretaceous of Mongolia.
Remarks. Family diagnosis takes into account the new composition. The phylogenetic affinities of dorsetisaurid genera are shown in Fig. 5. The diagram shows the undoubted close affinity between Dorsetisaurus Hoffstetter, 1967 and Paradorsetisaurus gen. nov., based on considerable similarities in tooth structure; as well as the basal position of Chometokadmon Costa, 1864 based on weak expansion of the crown apex and a larger tooth count.

Proposed phylogenetic relationships of the family Dorsetisauridae
-
Genus Paradorsetisaurus Alifanov, gen. nov.
Etymology. From the Greek para (near) and Dorsetisaurus.
Type species. Paradorsetisaurus postumus sp. nov.
Diagnosis. Premaxillary and jugal processes of the maxillary relatively long, the jugal process with a sharp tip. The medial branch of the premaxillary process carries a tubercle. The largest teeth are located in the middle of the toothrow. Their tips are flattened, somewhat curved inwards, turned at a small angle to the axis of the jaw and protrude by half their height beyond the maxilla. Tooth tip edge long and smooth. Upper tooth number—15.
Species composition. Type species.
Comparison. The new species is distinguished from other genera of the same family by the development of a dorsal tubercle on the medial branch of the premaxillary process and by a smaller tooth count. Compared with the type genus it is unique in a relatively long premaxillary and jugal processes of the maxillaries and in the arrangement of tooth crowns at an angle to the jaw axis. Further, the new genus is distinguished from Kuwajimalla Evans et Manabe, 2008 in having smooth cutting edges and from Chometokadmon Costa, 1864, in having taller teeth and expanded tooth tips.
-
Paradorsetisaurus postumus Alifanov, sp. nov.
-
Plate 14, figs. 1–4
Dorsetisaurus [sp.]: Alifanov, 1993, p. 9; 2000b, p. 382.
Etymology. From the Latin postumus (born last).

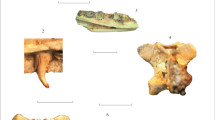
Explanation of Plate 14
Figs. 1–4.Paradorsetisaurus postumus gen. et sp. nov. (Dorsetisauridae), holotype PIN 3334/517, right maxilla: (1) labial view, (2) lingual view, (3) ventral view; (4) teeth in labial view, enlarged. Abbreviation: (Tpmx) dorsorostral tubercle.
Fig. 5. Lacertilia (?Dorsetisauridae) fam. indet., PIN 3334/519, fragment of a frontal in dorsal view.
Fig. 6 .Xenostius futilus gen. et sp. nov. (Xenosauridae s.str.), holotype PIN 3334/522, left dentary in lingual view.
Mongolia, Övörkhangai aimag, Khoobur locality; Lower Cretaceous, Upper Aptian–Lower Albian, Hühteeg Horizon.
Holotype. PIN 3334/517, right maxilla; Mongolia, Övörkhangai aimag, Khoobur Locality; early Cretaceous, Upper Aptian to Lower Albian, Hühteeg Horizon.
Description (Fig. 2g). The dorsal process of the maxillary passes gradually into the occipital process. The occipital process makes up half of the bone’s length. Premaxillary process wide at the base, its length more than a quarter of the total length of the maxilla. According to our reconstruction of the broken-off lateral part of the rostral process, the nares were enlarged. The base of the dorsal process makes up a third of the total length. The outer surface of the maxilla carries labially numerous foramina, no less than seven in the lower row. The supradental shelf wide, especially in the third quarter of the bone, at the level of teeth 8–11. The alveolar foramen is located on the dorsal surface of the supradental shelf below tooth 10. The tooth edge of the maxilla festoon-like. Teeth are set in shallow sockets, tooth implantation subpleurodont. At the base of the teeth a small resorption foramen is usually formed. There is also some cement deposition. Teeth project by half their height beyond the dental parapet.
Holotype size in mm. Length of the maxilla (reconstructed) 14.5 (15.5), height 4.3, height of the highest maxillary tooth 2.5.
Remarks. An interesting specimen, PIN 3334/519 (Pl. 14, fig. 5), is found in the collection of fossils from the Khoobur locality. It is an unpaired frontal belonging to a relatively large form. The bone is wide, with a sculptured dorsal surface and a series of foramina for the passage of nutrient vessels, arranged in two rows on either side of an imaginary middle line. Subolfactory processes are low and wide in the occipital part, thin and high in the rostral part of the bone. The facets for the attachment of the prefrontals are deep. Their posterior margin is located on the level of the middle of the orbit. Generally, the form represented by the specimen PIN 3334/519 is similar in appearance to Dorsetisaurus but is distinct in being unpaired and lacking transverse sulci on the dorsal surface. Quite possible this specimen belongs to the described species, though until further and more complete material becomes available, it cannot be identified more precisely than just Lacertilia fam. indet.
Material. Holotype.
-
Family Xenosauridae Cope, 1868
Type genus. Xenosaurus Cope, 1886, modern Mexico.
Diagnosis. The bones of the skull roof are usually covered in granular or small-scute ornamentation. Frontals always unpaired and narrow. Squamosals flattened occipitally. Teeth blunt, with a rostral shoulder.
Composition. In addition to the type genus, genera Exostinus Cope, 1873, late Cretaceous to early Oligocene of North America; Restes Gauthier, 1982, late Paleocene of North America; Oxia Nessov, 1985, early Cretaceous of Uzbekistan; Xenostius gen. nov., early Cretaceous of Central Mongolia.
-
Genus Xenostius Alifanov, gen. nov.
Etymology. Anagram of Exostinus.
Type species. Xenostius futilus sp. nov.
Diagnosis. Dentaries long, slim, straightened and weakly expanded occipitally. Posterior margin does not develop a deep notch. The flank of the subdental ridge narrow along its whole length. Teeth relatively large, straight and with an slightly expanded base. The part of the teeth projecting beyond the dorsal edge of the dentary makes up a third to a half of its total length. The largest teeth form in the middle of the tooth row. There is a small rostral shoulder on the tooth tips. Total tooth count 20.
Species composition. Type species.
Comparison. The new genus is unique within the family, judging by the small size and the gracility of the dentary. It is additionally distinguished from Oxia Nessov, 1985 in the weakly expanded flank of the subdental ridge, the presence of a distinct rostral shoulder near the tooth tip, as well as by a large part of the teeth extending beyond the dental parapet.
-
Xenostius futilus Alifanov, sp. nov.
-
Plate 14, fig. 6
Etymology. From the Latin futilus (fragile).
Holotype. PIN 3334/522, left dentary, Mongolia, Övörkhangai aimag, Khoobur locality; early Cretaceous, Upper Aptian–Lower Albian, Hühteeg Horizon.
Description (Fig. 4c). Dentary shallow and almost straight, its labial side notably rounded. The rostral part of the Meckelian canal is oriented ventromedially, which is usually typical for anguimorphs. The expanded base of the larger teeth does not have a transitional zone towards the tip. All of the teeth are generally oriented vertically, sometimes slightly deviating forward or back. In the Khoobur specimen, we have not been able to find the facet separating the central cusp of the tooth from its rostral shoulder.
Holotype size in mm. Length of the dentary: 7.4, height of its posterior margin: 1.2 (reconstructed height 1.5), height of the highest dentary tooth: 1.1.
Material. In addition to the holotype, PIN 3334/521; left dentary with teeth, type locality.
THE DIVERSITY OF THE LIZARDS OF CENTRAL ASIA IN THE JURASSIC AND THE EARLY CRETACEOUS
The lizard families of the Khoobur assemblage can be split into two groups (Fig. 1). Part of the first group was widely distributed not only in the early Cretaceous of Central Asia, but also beyond it in the Jurassic of Europe, North America (Paramacellodidae and Dorsetisauridae) and Africa (Paramacellodidae). The other group is found in the Jurassic (Ardeosauridae) and early Cretaceous of Europe (Globauridae). All representatives of this group likely were distributed within the Jurassic Laurasia or Afrolaurasia, e.g., Laurasia connected with Africa.
The second group within the assemblage is endemic to early Cretaceous Central Asia. The all families composing this group (Eoxantidae, Gekkonidae s.l., Hodzhakuliidae, Temujiniidae, and Xenosauridae) first appeared in early Cretaceous of Central Asia and their appearance is tied with the territorial isolation of this paleocontinent which lasted during the final Jurassic and most of the early Cretaceous. The isolation of Central Asia, as part of the breakup of Afrolaurasia, was suggested by N.N. Kalandadze and A.S. Rautian (1992, 1997).
Outside of Mongolia, the Khoobur assemblage can certainly be compared in age with two lizards of the family Hodzhakuliidae: Pachygenys thlastesa Gao et Cheng, 1999 from the Doushan Formation (found to be Aptian-Albian in age by S. Lucas (2006)) in China (Shandong); and another species of the same genus, P. adashii Ikeda et al., 2015 from the lower part of the Sasayama Formation (Aptian–Albian boundary) in Japan (Hyogo).
The makeup of the only post-Khoobur (late Albian) assemblage from the lower part of the Hodzhakul Formation in Uzbekistan is remarkable, with four groups identified so far (Gao and Nessov, 1998). Our identifications suggest that, in addition to the already mentioned Xenosauridae, the Hodzhakul assemblage also includes members of the families Hodzhakuliidae, Ardeosauridae gen. indet. and Temujiniidae gen. indet. This family composition, even taking into account incomplete material, indicates lower lizard diversity in post-Khoobur times. Based on the material from the late Cretaceous of Mongolia, the only families to go extinct among those found in Khoobur are Hodzhakuliidae and Dorsetisauridae. The family Xenosauridae s. str. also disappears from Central Asia but appears in the late Cretaceous of another continent, North America.
The diversity of lizards at the Aptian–Albian boundary stands in contrast with the low diversity of lizards in earlier times. For instance, only one family is found in the Jurassic deposits of Central Asia, the Paramacellodidae (Changetisaurus estesi Nessov, 1992, middle Jurassic of Kazakhstan; Sharovisaurus karatauensis Hecht et Hecht, 1984, late Jurassic of Kazakhstan; Mimobecklesisaurus gansuensis Li, 1991, late Jurassic of China). The only Neocomian lizard fossil found in Central Asia comes from the Hodzhiabat Formation of Kyrghyzstan (Lacertilia gen. indet.: Averianov and Fedorov, 2004; Ardeosauridae: Alifanov, 2012).
Among the pre-Khoobur lizards fossils of Mongolia we should note Norellius nyctisaurops Conrad et Daza, 2015 (Ardeosauridae, our identification) from the Undurukhinsk Formation (Oosh Formation), exposed in Central Mongolia, and Bavarisauridae gen. indet. (Alifanov, 2000a, 2012) from the Tsagantsab Formation in Western Mongolia. Both stratigraphic units occur within the Tsagantsab Group, for which absolute dates are known. These fall within the range of 141–119 Ma (Shuvalov, 2000), which is closer to the Hauterivian–Barremian, rather than the entire Neocomian (Shuvalov, 2000) or the Barremian–Aptian (Lucas, 2006).
Outside of Mongolia, lizards of Barremian–Aptian age are known from the Tetori Group in Japan: Kaganaias hakusanensis Evans et al., 2006 (Dolichosauridae), Kuwajimalla kagaensis Evans et Manabe, 2008 (Dorsetisauridae; our identification) from the Kuwajima Formation (Ishikawa Prefecture) and Sakurasaurus shokawensis Evans et Manabe, 1999 (Ardeosauridae; our identification) from the Okurodani Formation (Gifu Prefecture). Fossil finds of similar age have been reported from the Ilek Formation in the Kemerovo District, Russia (Paramacellodidae indet., Xenosauridae indet.: Averianov, Fayngerts, 2001) and the Murtoy Formation in Transbaikalia, Russia (Paramacellodidae indet.: Averianov and Skutchas, 1999). There are numerous lizard fossils in the deposits of the Jehol Group (Barremian–Aptian) of Northeastern China (Jehololacerta formosa Ji et Ren, 1999; Dalinghosaurus longidigitus Ji, 1998; Liaoningolacerta brevirostra Ji, 2005; Liushusaurus acanthocaudata Evans et Wang, 2010; Yabeinosaurus tenuis Endo et Shikama, 1942). All of these, most likely, belong to scincomorphs of the family Ardeosauridae which, in our view, was the dominant group in Central Asia throughout the whole early Cretaceous.
Compared to this low diversity of early Cretaceous lizards in Central Asia, all the more notable is the Aptian-Albian diversity maximum established using the material from the Khoobur locality. Increased fossil diversity is not only reflective of taphonomic conditions favorable for the accumulation of fossil remains but also of environmental conditions (climate warming, increased insect diversity and biomass). This was the first of the two Cretaceous diversity peaks in Central Asia since their appearance in the fossil record. The second, much higher peak is registered in the Campanian (Alifanov, 2000a).
It should be emphasized that the Jurassic to early Cretaceous time interval forms a distinct historical stage in lizard faunas. Its notable feature is the predominance of the scincogekkonomorphs, especially the Scincomorpha (Paramacellodidae in the Jurassic, Ardeosauridae in the early Cretaceous in general, Hodzhakuliidae in the Upper Aptian and Albian). This composition of dominant taxa is in stark contrast with other such stages: the late Cretaceous with the predominance of members of the infraorder Chamaeleomorpha (Macrocephalosauria, and Priscagamia) and the Paleogene, when the microorder Pachyglossa (Changjiangosauridae, Uromastycidae, and Agamidae) reached its maximum diversity.
In summary, several phases can be reconstructed within the Jurassic to early Cretaceous stage in the development of lizard faunas of Central Asia. These are the second half of the Jurassic (the dominance of Paramacellodidae), Neocomian to early Aptian (the dominance of Ardeosauridae and the maintenance of archaic diversity), late Aptian to early Albian (maximum diversity achived with the appearance of endemic groups), and late Albian (reducing compared to the previous phase).
REFERENCES
Alifanov, V.R., The oldest gecko (Lacertilia, Gekkonidae) from the early Cretaceous of Mongolia. Paleontol. J., 1989, no 1, pp. 124–126.
Alifanov, V.R., Some peculiarities of the Cretaceous and Palaeogene lizard faunas of the Mongolian People’s Republic, Kaupia, 1993, no. 3, pp. 9–13.
Alifanov, V.R., Makrotsefalozavry i rannie etapy evolyutsii yashcherits Mongolii (Macrocephalosaurs and the Early Stages of the Evolution of the Lizards of Mongolia), Tr. Paleontol. Inst. Ross. Akad. Nauk, vol. 272, Moscow, GEOS, 2000a.
Alifanov, V.R., The fossil record of Cretaceous lizards of Mongolia, The Age of Dinosaurs in Russia and Mongolia, Benton, M., Shishkin, M.A., Unwin, D.M., and Kurochkin, E.N., Eds., Cambridge: Univ. Press, 2000b, pp. 368–389.
Alifanov, V.R., Order Lacertilia, Iskopaemye pozvonochnye Rossii i sopredel’nyh stran. Iskopaemye reptilii i ptitsy. Chast’ 2 (Fossil Vertebrates of Russia and Neighbouring Countries: Fossil Reptiles and Birds. Part 2), Kurochkin, E.N. and Lopatin, A.V., Eds., Moscow: GEOS, 2012, pp. 7–136.
Alifanov, V.R., Lizards of the family Hodzhakuliidae (Scincomorpha) from early Cretaceous of Mongolia, Paleontol. J., 2016, vol. 50, no 5, pp. 504–513.
Alifanov, V.R., Lizards of the family Temujiniidae (Iguanomorpha): fossil finds from Aptian–Albian of Mongolia, classification and geographic origin. Paleontol. J., 2018, vol. 52, no 6, pp. 653–663.
Averianov, A.O. and Fayngerts, A.V., Lizards of the early Cretaceous of Western Siberia, Voprosy gerpetologii: Mater. I s’’yezda Gerpetol. ob-va im. A.M. Nikol’skogo (Issues in Herpetology: Proc. 1st Session of the Nikolsky Herpetology Society), Pushchino–Moscow, 2001, pp. 6–8.
Averianov, A.O. and Fedorov, P.V., Early Cretaceous lizard from the Hodzhiabad Formation of Fergana (Kyrgyzstan). Vestn. SpbGU, Ser. 7, 2004, no. 1, pp. 81–82.
Averianov, A.O. and Skutchas, P.P., Paramacellodid lizard (Squamata, Scincomorpha) from the early Cretaceous of Transbaikalia, Russ. J. Herpetol., 1999, vol. 6, no. 2, pp. 115–117.
Bever, G.S., Bell, C.J., and Maisano, J.A., The ossified braincase and cephalic osteoderms of Shinisaurus crocodilurus (Squamata, Shinisauridae), Palaeontol. Electron., 2005, vol. 8, no. 1, pp. 1–36.
Bhullar, B.-A. S., The power and utility of morphological characters in systematics: a fully resolved phylogeny of Xenosaurus and its fossil relatives (Squamata: Anguimorpha), Bull. Mus. Comp. Zool., 2011, vol. 160, no. 3, pp. 65–181.
Borsuk-Białynicka, M., Carolinidae, a new family of xenosaurid-like lizards from the Upper Cretaceous of Mongolia, Acta Palaeontol. Pol., 1985, vol. 30, nos. 3–4, pp. 151–176.
Borsuk-Białynicka, M., Carussia, a new name for the late Cretaceous lizards from the Upper Cretaceous of Mongolia, Acta Palaeontol. Pol., 1987, vol. 32, nos. 1–2, p. 153.
Broschinski, A., The lizards from the Guimarota mine, Guimarota: A Jurassic Ecosystem, Martin, T. and Krebs, B., Eds., Munich: Dr. F. Pfeil Verlag, 2000, pp. 59–68.
Caldwell, M.W., Randall, L., Nydam, R.L., Palci, A., and Apesteguía, S., The oldest known snakes from the Middle Jurassic–Lower Cretaceous provide insights on snake evolution, Nature Communicat., 2015, no. 5976, pp. 1–11.
Camp, C.L., Classification of the lizards, Bull. Am. Mus. Nat. Hist., 1923, vol. 48, pp. 289–481.
Conrad, J.L., Skull, mandible, and hyoid of Shinisaurus crocodilurus Ahl (Squamata, Anguimorpha), Zool. J. Linn. Soc., 2004, vol. 141, no. 3, pp. 399–434.
Conrad, J.L., An Eocene shinisaurid (Reptilia, Squamata) from Wyoming, USA, J. Vertebr. Paleontol., 2006, vol. 26, no. 1, pp. 113–126.
Conrad, J.L., Phylogeny and systematics of Squamata (Reptilia) based on morphology, Bull. Am. Mus. Nat. Hist., 2008, vol. 310, no. 1, pp. 1–182.
Daza, J.D., Alifanov, V.R., and Bauer, A.M., A redescription and phylogenetic reinterpretation of lizard Hoburogekko suchanovi Alifanov 1989 (Squamata: Gekkota), J. Vertebr. Paleontol., 2012, vol. 32, no. 6, pp. 1303–1312.
Estes, R., Sauria terrestria, Amphisbaenia, Handbuch der Paläoherpetologie, vol. 10A, Stuttgart: G. Fischer Verlag, 1983.
Evans, S.E. and Manabe, M., An early herbivorous lizard from the Lower Cretaceous of Japan, Palaeontology, 2008, vol. 51, part 2, pp. 487–498.
Evans, S.E., Raia, P., and Barbera, C., The Lower Cretaceous lizard genus Chometokadmon from Italy, Cret. Res., 2006, vol. 27, no. 5, pp. 673–683.
Fedorov, P.V. and Nessov, L.A., A lizard from the middle–late Jurassic boundary of the Northwestern Fergana, Vestn. SPbGU, Ser. 7, 1992, vol. 3, no. 21, pp. 9–14.
Fernandez, V., Buffetaut, E., Suteethorn, V., Rage, J.-C., Tafforeau, P., and Kundrát, M., Evidence of egg diversity in Squamate evolution from Cretaceous anguimorph embryos, PLoS One, 2015, vol. 10, no. 7, pp. 1–20.
Gao, K. and Nessov, L.A., Early Cretaceous squamates from the Kyzylkum Desert, Uzbekistan, N. Jb. Geol. Paläontol. Abh., 1998, vol. 207, pp. 289–309.
Gao, K. and Norell, M.A., Taxonomic revision of Carusia (Reptilia: Squamata) from the late Cretaceous of the Gobi Desert and phylogenetic relationships of anguimorphan lizards, Am. Mus. Novit., 1998, no. 3230.
Gao, K. and Norell, M.A., Taxonomic composition and systematics of late Cretaceous lizard assemblages from Ukhaa Tolgod and adjacent localities, Mongolian Gobi Desert, Bull. Am. Mus. Nat. Hist., 2000, vol. 249, pp. 1–118.
Gauthier, J.A., Kearney, M., Maisano, J.A., Rieppel, O., and Behlke, A.D.B., Assembling the squamate tree of life: Perspectives from the phenotype and the fossil record, Bull. Peabody Mus. Nat. Hist., 2012, vol. 53, no. 1, pp. 3–308.
Hoffstetter, R., Coup d’oeil sur les sauriens (=lacertiliens) des couches de Purbeck (Jurassique supérieur d’Angleterre), Problèmes Actuels de Paléontologie (Evolution des Vertébrés). Coll. Intern. CNRS, 1967, vol. 163, pp. 349–371.
Kalandadze, N.N. and Rautian, A.S., The system of mammals and historical zoogeography, Filogenetika mlekopitaiushchih (Mammal Phylogenetics), Rossolimo, O.L., Ed., Tr. Zool. Mus. Mosk. Gos. Univ., vol. 29, Moscow: Mosk. Gos. Univ., 1992, pp. 44–152.
Kalandadze, N.N. and Rautian, A.S., Historical zoogeography of terrestrial tetrapods and new method of global palaeogeographical reconstructions, Evolution of the Biosphere, Rozanov, A.Yu. and Vickers-Rich, P., Eds., Rec. Queen Victoria Mus., Art Gallery, no. 104, Launceston: C. Tassel, 1997, pp. 95–98.
Klembara, J., A new anguimorph lizard from the lower Miocene of north-west Bohemia, Czech Republic, Palaeontology, 2008, vol. 51, part 1, pp. 81–94.
Lucas, S.G., The Psittacosaurus biochron, Early Cretaceous of Asia, Cret. Res., 2006, vol. 27, no. 2, pp. 189–198.
McDowell, S.B.J. and Bogert, C.M., The systematic position of Lanthanotus and the affinities of the anguinomorphan lizards, Bull. Am. Mus. Nat. Hist., 1954, vol. 105, no. 1, pp. 1–105.
Nydam, R.L., Squamates from the Jurassic and Cretaceous of North America, Palaeobiol. Palaeoenv., 2013, vol. 93, no. 4, pp. 535–565.
Prothero, D. and Estes, R., Late Jurassic lizards from Como Bluff, Wyoming, and their palaeobiogeographic significance, Nature, 1980, vol. 286, no. 5772, pp. 484–486.
Seiffert, J., Upper Jurassic lizards from Central Portugal, Mem. Serv. Geol. Port., 1973, vol. 22, pp. 1–85.
Shuvalov, V.F., The stratigraphy and palaeobiogeography of Mongolia, The Age of Dinosaurs in Russia and Mongolia, Benton, M. J., Shishkin, M.A., Unwin, D.M., and Kurochkin, E.N., Eds., Cambridge: Univ. Press, 2000, pp. 256–278.
ACKNOWLEDGMENTS
This work was supported by the Russian Foundation for Basic Research, project no. 16-05-00408.
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by D. Ponomarenko
Rights and permissions
About this article

Cite this article
Alifanov, V.R. Lizards of the Families Dorsetisauridae and Xenosauridae (Anguimorpha) from the Aptian–Albian of Mongolia. Paleontol. J. 53, 183–193 (2019). https://doi.org/10.1134/S0031030119020023
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030119020023