
Abstract
Squamates from the Mesozoic of North America have been collected since the end of the nineteenth century. To date, the fossils are known to occur in the Late Jurassic, Aptian-Albian, Cenomanian, Turonian, Santonian, Campanian, and Maastrichtian. Most of the records are from the Western Interior in the arid regions associated with the Rocky Mountains. Geographically, these records extend from central Alberta, Canada, south to northern Mexico. The earliest squamates are primitive forms of scincoideans and anguimorphans from the Late Jurassic–Early Cretaceous. At the beginning of the Late Cretaceous, the squamate fauna in North America changes dramatically to include a much greater diversity of taxa with a mix of lineages now extinct (e.g. Polyglyphanodontini, Chamopsiidae, Contogeniidae) and lineages still alive today (e.g. Anguidae, Xantusiidae, Platynota, Serpentes). The greatest diversity appears to be during the late Campanian, but diversity appears correlated with number of localities sampled and the late Campanian is the best sampled horizon in the Mesozoic of North America. The apparent sudden change in the North American squamate fauna is coincident with similar changes to other vertebrates (mammals, dinosaurs) and the opening of a land bridge with Asia. The lack of taxonomic and systematic study of the squamates from the Early Cretaceous of Asia makes comparison difficult, but it is likely that introduction of Asian taxa into North America was responsible, at least in part, for the relatively rapid change in the North American fauna. The hypotheses of an additional invasion from Asia during the Turonian is not supported, but the hypothesis of a second opening with Asia during the Santonian is weakly supported by the appearance of an iguanian in North America. Additional iguanians from the Campanian may have evolved in situ or may have entered North America from Asia as late as the mid-Campanian. Many of the most conspicuous lizards of the Late Cretaceous (Polyglyphanodontini, Chamopsiidae, paramacellodid-cordylid grade scincoideans) went extinct at the terminal Cretaceous extinction event, while most of the anguimorphans and snakes appear little affected. Amphisbaenians do not appear in North America until after the early Paleocene.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The earliest report of terrestrial fossil squamatesFootnote 1 from the Mesozoic of North America is a short description of jaws of the lizards Chamops segnis and Iguanavus teres (later synonymised with Chamops segnis; Estes 1983a) and a vertebra of the snake Coniophis precedens from the Late Cretaceous of Wyoming (Marsh 1892). There are a couple of aspects that make this initial report fascinating. Firstly, the specimens described are isolated vertebrae and jaws (which are typically incomplete or worn). With only a few notable exceptions, the squamate record from the Mesozoic of North America is comprised of taxa based on isolated elements; mostly incomplete mandibles and maxillaries of lizards and isolated vertebrae of snakes. The second interesting aspect of this initial report is the region from which the fossils were recovered. The Lance Formation of Wyoming is one in a series of many rock units from the arid Western Interior of North America that preserves terrestrial vertebrates spanning the Upper Jurassic through Upper Cretaceous. This vast region extends along the Rocky Mountains from Canada into northern Mexico and is the source of the overwhelming majority of fossil lizards and snakes from the Mesozoic of North America. These two aspects of this first publication are the iterative themes of nearly all subsequent reports on terrestrial lizards and snakes from the Mesozoic of North America: a record of mostly fragmentary and isolated remains of lizards and snakes from a longitudinally narrow and latitudinally expansive region of the continent.
The next substantial contributions to the terrestrial squamate record from the Mesozoic (all Late Cretaceous) of North America were by C. W. Gilmore. His monograph on North American fossil lizards (Gilmore 1928) identified 11 additional lizard species from the Late Cretaceous (subsequently revised to six lizards; e.g. Estes 1983a; Sullivan 1991; Gao and Fox 1996), which include the first report of anguids, xenosaurids, and platynotans from the Mesozoic of North America. The next contribution was Gilmore’s monograph (1938) on fossil snakes of North America in which Marsh’s Coniophis precedens remained the only known snake from the Mesozoic. Gilmore (1942a, 1943) later described the first, and currently most complete, articulated lizard skeleton from the Mesozoic of North America, the large-bodied Polyglyphanodon sternbergi from the Maastrichtian of Utah. For the most part, all these discoveries of squamate fossils had been made as serendipitous discoveries during investigations into macrofossil occurrences, particularly those of dinosaurs.
With the introduction in the 1950s and 1960s of the use of underwater screen-wash concentration techniques specifically for the recovery of mammal teeth, the record of squamates from the Mesozoic of North America began to increase substantially (see reviews in Cifelli 1996; Estes 1964; Kielan-Jaworoswka et al. 2004). In cases of exceptional preservation (e.g. Evans and Chure 1998a, b, 1999; Prothero and Estes 1980; Reynoso 1998; Reynoso and Callison 2000) hand-quarrying continues to be a preferred technique. Most of the screen-wash-based records are from the Campanian–Maastrichtian ages of the latter half of the Late Cretaceous (Armstrong-Ziegler 1978, 1980; Denton and O'Neill 1995; Estes 1964, 1969a, b; Estes et al. 1969; Fox 1975; Gao and Fox 1991, 1996; Longrich et al. 2012a; McCord 1998; Nydam 1999, 2013; Nydam et al., 2007, 2010; Nydam and Fitzpatrick 2009; Nydam and Voci 2007; Peng et al. 2001; Rowe et al. 1992; Sullivan 1981). Other contributions include taxa from the Cenomanian–Santonian ages of the first half of the Late Cretaceous (Cifelli and Nydam 1995; Nydam 1999, 2000, 2002; Nydam and Cifelli 2002a, b; Nydam and Fitzpatrick 2009), and the Early Cretaceous (Nydam and Cifelli 2002a; Reynoso 1995, 1996, 1998; Winkler et al. 1990). There are currently no records of squamates from North America prior to the Late Jurassic.
The result to date of these published reports is a substantial fossil record of squamates from the Late Jurassic–Late Cretaceous of North America. Previous reviews of this record (most notably those of Estes 1983b; Evans 1995, 2003) have examined and summarised the North American fauna within the broader context of the evolution of the global squamate fauna. Within such a framework, any questions regarding regionally specific occurrence and distributional patterns of squamates within North America were necessarily reduced to presence/absence and first/last occurrences on the continent. However, the record of squamates from the Late Jurassic–Late Cretaceous of North America is one of the richest in the world, with a nearly continuous temporal record and extensive latitudinal distribution of taxa. The goal of this paper is to review and summarise the geographic and temporal distributions of squamates from the Late Jurassic through Late Cretaceous of North America and to identify patterns and/or changes in these distributions.
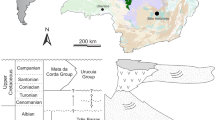
There are some constraints on this record. Firstly, nearly all the fossil squamates come from localities from the Western Interior of North America (Fig. 1), which, during the Late Cretaceous, were along the western shore of the mid-continental seaway. Other regions of North America lack appropriately aged terrestrial sediments, are too heavily vegetated, or remain uninvestigated. Secondly, for the sake of brevity and to allow for higher taxonomic comparisons, the taxonomic levels used here are primarily at the family level (e.g. Anguidae, Contogeniidae, Chamopsiidae, etc.). However, because of the fragmentary nature of many of the specimens, there is insufficient information for family-level referrals and broader taxonomic categories (e.g. Serpentes, Platynota, Scincoidea) or non-taxonomic categories (e.g. of paramacellodid–cordylid grade) are used instead. For those groups that apparently survived the terminal Cretaceous extinction event (see also Longrich et al. 2012a), I note their presence in the Paleocene, but the Cenozoic record of squamates from North America will not be summarised here.

Squamate-producing localities from the Upper Jurassic through Paleocene of North America. Upper Jurassic Morrison Formation (X): 1 Como Bluff; 2 Dinosaur National Monument; 3 Fruita Paleontological Area. Aptian-Albian (crosses): 4 Antlers Formation; 5 Trinity Group; 6 Tlayúa Formation; 7 Cloverly Formation. Lower Cenomanian (open circle): 8 Cedar Mountain Formation. Upper Cenomanian (circle with X): 9 Dakota Formation and lower portion of Iron Springs Formation. Turonian (diamond): 10 Smoky Hollow Member of Straight Cliffs Formation. Santonian (trapezoid): 11 John Henry Member of Straight Cliffs Formation and upper portion of Iron Springs Formation; 12 Milk River Formation. Lower-mid Campanian (south pointing triangles): 13 Wahweap Formation. Upper Campanian (north pointing triangles): 14 Foremost and Oldman formations; 15 Wapiti Formation; 16, 17 Dinosaur Park Formation (16 Dinosaur Provincial Park locality; 17 Irvine locality); 18 Two Medicine Formation; 19 Judith River Formation; 20 Mesaverde Formation; 21 Kaiparowits Formation; 22 Fruitland Formation; 23 Marshalltown Formation; 24 Bladen Formation; 25 “El Gallo” Formation; 26 Fort Crittenden Formation;. 27 Aguja Formation; 28 Cerro del Pueblo Formation. Maastrichtian (squares): 29 Scollard Formation; 30, 31 Frenchman Formation; 32, 33 Hell Creek Formation (32 Fort Peck Reservoir area; 33 Ekalaka area); 34 Fox Hills Formation; 35 Lance Formation; 36 lower portion of North Horn Formation; 37 Laramie Formation; 38 Kirtland Formation. Paleocene (stars): 39 Tullock Formation 40, 41 Tongue River Formation (40 Polecat Bench; 41 Ekalaka); 42 upper portion of North Horn Formation; 43 Denver Formation; 44 Goler Formation; 45 Nacimiento Formation. See Table 1 and text for specific references. Canadian province abbreviations: AB Alberta; SK Saskatchewan. US state abbreviations: AZ Arizona; CO Colorado; MT Montana; NC North Carolina; NJ New Jersey; NM New Mexico; OK Oklahoma; SD South Dakota; TX Texas; UT Utah; WY Wyoming. Mexico state abbreviations: MX-BC Baja California; MX-CO Coahuila; MX-PB Puebla
Abbreviations for institutions: SMNH, Saskatchewan Museum of Natural History, Regina, Saskatchewan, Canada; UALVP, University of Alberta Laboratory of Vertebrate Paleontology, Edmonton, Alberta, Canada; UCMP, University of California Museum of Paleontology, Berkeley, California, U.S.A.; USNM, United States National Museum, Washington, D.C., U.S.A.
Localities
The localities from the Jurassic and Cretaceous of North America that have produced squamate fossils are mostly within the Western Interior along a narrow north–south line extending from central Canada to central Mexico adjacent to the Rocky Mountains (Table 1; Fig. 1). The primary reason for this distribution is the arid climate allowing for the exposure of the vast amounts of Mesozoic-aged sedimentary rocks in this part of the continent. Another factor particularly important to the distribution of fossil squamates from the Late Cretaceous is that this narrow strip of land was, at that time, a lowland region of accumulating sediments between palaeomountain systems to the west and a seaway to the east (e.g. Eberth 2002; Eberth and Hamblin 1993; Eberth et al. 2004; Lehman 1997, 2001; Titus et al. 2005). During the Late Cretaceous, this seaway separated the North American continent into two sub-continents; Laramidia to the west and Appalachia to the east (terminology sensu Gates et al. 2010 and references therein). There are only a few examples of squamate-producing localities that are not along this line (e.g. “El Gallo”, Bladen, and Marshalltown formations during the upper Campanian). I expect that additional localities will continue to be discovered as more investigations focus on regions outside of the Western Interior.
Another important similarity between these faunas is the depositional environment. Most of the localities are freshwater fluvial sediments in the form of overbank splay or flood deposits from meandering river systems (see citations below). These localities are mostly characterised by mudstones with interbedded channel sandstones, and the vertebrate fossils include freshwater aquatic inhabitants of the stream systems and numerous terrestrial animals presumably from the immediate area around the fluvial systems. Nearly all of the vertebrate fossils from these localities, including the vast majority of fossil lizards and snakes, occur as isolated elements aggregated in lag deposits in mudstones. As noted above, the adoption of screen-wash-based sampling techniques allowed for the collection of thousands of specimens which can be assumed to have lived in relatively similar environments (lowlands along stream/river systems) throughout the Western Interior.
Because the North American Land Mammal Ages (e.g. Aquilan, Judithian, Lancian) of the Mesozoic are defined only for the latter half of the Late Cretaceous, I have chosen to use the invertebrate-based ages (e.g. Cenomanian, Turonian, Coniacian, etc.) instead. This maintains consistency of terminology throughout the temporal sequence and is effectively more portable when making comparisons to faunas outside of North America. North American Land Mammal Ages (NALMAs) are given as parenthetical equivalents as appropriate.
Review of squamate-producing localities
The known squamate-producing horizons are summarised below. Relevant citations regarding squamates are given in Table 1 and locality plots are given in Fig. 1. Additional citations are given in the review of taxonomic occurrences below.
Late Jurassic
Morrison Formation
The earliest temporal occurrence of squamates from North America is from the Morrison Formation of Colorado, Utah, and Wyoming, USA (Callison 1987; Carrano and Velez-Juarbe 2006; Evans 1996; Evans and Chure 1998a, b, 1999; Prothero and Estes 1980). Unlike later squamate faunas, the lizards from the Morrison Formation co-occur with sphenodontians (Callison 1987; Carrano and Velez-Juarbe 2006; Chure et al. 1998; Fraser and Wu 1998; Rasmussen and Callison 1981). The typical sedimentology containing squamate remains is mudstones associated with fluvial deposits (Callison 1987; Carrano and Velez-Juarbe 2006).
Early Cretaceous
Aptian–Albian
Unlike Europe and Asia (e.g. Evans and Manabe 1999; Evans et al. 2000, 2005; Evans and Wang 2005), which have produced a modest record of squamates from the pre-Aptian Cretaceous, there is a gap in the record of squamates from North America between the Late Jurassic and late Early Cretaceous. The next oldest North American fossil squamates are Aptian–Albian from terrestrial/fresh water localities along the coasts of the encroaching northern (Montana, Wyoming) and southern (Texas, Oklahoma) seaways that eventually connected to form the mid-continental seaway during the Late Cretaceous. The Cloverly Formation (Montana, Wyoming) microvertebrate localities are from fluvial deposits (Forster 1984; Ostrom 1970), the Antlers Formation (Oklahoma) is a flood plain (Brinkman et al. 1998, and references therein), and the Trinity Group of Texas is a series of near-shore terrestrial facies (Winkler et al. 1990). Additionally. there is a near-shore marine locality (sensu Kashiyama et al. 2004) in Puebla, Mexico (Fig. 1) which has produced some nearly complete lizards and sphenodontians (Reynoso 1997, 1998, 2000; Reynoso and Callison 2000).
Recent investigations of the microvertebrate fauna of the Lakota Formation (Barremian) of South Dakota have resulted in the recovery of some possible lizard jaw specimens, but these have not yet been published (R.L. Cifelli. personal communication).
Late Cretaceous
Cenomanian
Squamate fossils have been recovered from both the base of the Cenomanian in the Mussentuchit Member of the Cedar Mountain Formation and the mid-Cenomanian from the Dakota Formation, both in Utah. The age of the Mussentuchit Local Fauna has been variably reported to be “Early” or “Lower” Cretaceous (e.g. Burge 1996; Cifelli 1993; Cifelli and Nydam 1995; Currie 1998; Gardner 1994; Garrison et al. 2007) or as being “Medial Cretaceous” or “Albian-Cenomanian boundary” (Cifelli 1999, 2004; Cifelli and de Muizon 1997; Cifelli and Madsen 1998, 1999; Cifelli et al. 1999c; Eaton and Cifelli 2001; Gardner 1999; Gardner and Cifelli 1999; Kirkland et al. 1999; Nydam 1999, 2000; Nydam and Cifelli 2002b). A 40Ar/39Ar test of an undisturbed ash layer within primary microvertebrate locality gave a date of 98.39 ± 0.07 Ma (Cifelli et al. 1997). This date places the fauna near the base of the Cenomanian (100.5 Ma; sensu Ogg and Hinnov 2012), but certainly well within the early part of the Late Cretaceous. The primary sedimentological facies for the squamate-producing localities is fluvial-based mudstones (Cifelli et al. 1999c; Garrison et al. 2007).
The age of the Dakota Formation in southern Utah is typically cited as middle Cenomanian (Eaton 1991, 1995; Eaton et al. 1999a). Recently reported radiometric dates for the middle portion of the Dakota Formation (contains vertebrate localities; Eaton 1995) place the age at 95.97 ± 0.22 Ma (Dyman et al. 2002a) or 96.06 ± 0.30 Ma (Dyman et al. 2002b). Based on the latest geologic time scale for the Cretaceous (Ogg and Hinnov 2012), both these dates place the age of the middle member of the Dakota Formation in the middle Cenomanian.
Cenomanian-age lizards have also been recovered from the base of the Iron Springs Formation in southern Utah. This is a very thick unit that extends into the Santonian, making it the lateral equivalent of several distinct nearby Upper Cretaceous formations (Dakota, Tropic Shale, Straight Cliffs) (Eaton 1999). Indeed, there is a very thick sequence of Upper Cretaceous strata in southern Utah that includes many fluvial-based terrestrial units that are known to contain squamate fossils. For a more comprehensive review of the geology of the Late Cretaceous of southern Utah, I refer readers to Eaton (1991) and Titus et al. (2005), as well as Titus et al. (2013).
Turonian
The Smoky Hollow Member of the Straight Cliffs Formation in southern Utah has produced a diversity of squamate fossils. This unit is also important in that it has produced the only known Turonian mammals from North America (Cifelli 1990a; Eaton 1991, 1995). A radiometric date from a level very close to the productive horizon of this unit gives the age as 91.9 ± 0.3 Ma (Titus et al. 2013), or about middle Turonian. Since this is the only known horizon from the Turonian of North America to produce squamate fossils, all references henceforth will simply refer to “Turonian” as the age.
Santonian
The John Henry Member of the Straight Cliffs Formation and the uppermost portion of the Iron Springs Formation (both in southern Utah) have produced a small number of squamate fossils. The age of the microvertebrate producing horizons of both of these units has been determined to be Santonian (Eaton 1999, 2006; Eaton and Cifelli 1988; Eaton et al. 1999a, b). Mammals of the Iron Springs Formation appear to be very similar to those of the Milk River Formation (Eaton 2006) and radiometric dates of 84.5–83.5 Ma for the Milk River Formation (Payenberg et al. 2002)—initially thought to place the lower portion in the Santonian and the upper portion in the early Campanian—can now be considered to be completely within the Santonian, based on the most recent geologic time scale for the Cretaceous (Ogg and Hinnov 2012). The squamates from the Milk River Formation have been recovered from near the top of the unit (Gao and Fox 1996) placing them at the upper Santonian, based on these current chronostratigraphic interpretations.
Early-mid Campanian
The Wahweap Formation of southern Utah is generally considered to be lower–mid-Campanian (approximately equal to the Aquilan NALMA), particularly the fossil-producing horizons from the lower 2/3 of the unit (Cifelli 1990b; Eaton and Cifelli 1988). The Wahweap has previously been correlated with the Milk River Formation of southeastern Alberta based on similarities in the mammal fauna (Cifelli 1990b; Cifelli and Madsen 1986; Eaton 2002; Eaton and Cifelli 1988; Eaton et al. 1999a) as well as some aspects of the selachian fauna (Kirkland et al. 2013). A radiometric date for the Wahweap (Jinnah et al. 2009) supports a lower–mid-Campanian age, but the suggestion by Jinnah et al. (2009) that the unit is correlative to Judithian faunas (∼upper Campanian) is not supported by the faunal similarities between the Wahweap and Milk River formations (see citations above), in addition to the lack of similarity between the Wahweap and well-accepted Judithian faunas of the late Campanian.
The similarity in the vertebrate (particularly mammal) faunas between the Milk River Formation (upper Santonian) and the Wahweap Formation (lower–mid-Campanian) is not considered problematic since they are likely to be approximately stratigraphically equivalent and the index taxa likely spanned the upper Santonian–early Campanian. However, the problem does require additional study, and additional data (e.g. additional radiometric dates, additional fossil specimens, and continued reinterpretation of the age of the Santonian–Campanian boundary) may change these relationships again. The squamates of the Wahweap Formation are too poorly known (Nydam 2013) to aid in the resolution of this problem. However, the local stratigraphic relationships are clear: in southern Utah, the Wahweap Formation underlies the Kaiparowits Formation (the base of which is dated at 76.1 Ma, or the base of the upper Campanian; Roberts et al. 2005) and in southern Alberta, the Milk River Formation underlies the Judith River Group (the mid–upper Campanian Foremost and Oldman formations and the upper Campanian Dinosaur Park Formation; Eberth and Hamblin 1993).
Late Campanian
This horizon (approximately equal to the Judithian NALMA) is the most productive in North America in terms of fossil squamates. The rock units from which squamates have been recovered include the Wapiti (west central Alberta), Dinosaur Park (southern AlbertaFootnote 2), Oldman (southern Alberta), Foremost (southern Alberta), Judith River (Montana), Mesaverde (central Wyoming), Kaiparowits (Utah), Fort Crittenden (southern Arizona), Fruitland (northwestern New Mexico), “El Gallo” (informal; Baja California), Aguja (southern Texas), Cerro del Pueblo (Coahuila), and Marshalltown (central New Jersey) formations. These localities also represent the greatest paracontemporaneous geographical distribution of squamates (central Alberta to northern Mexico), which has been reviewed in more detail in Nydam et al. (2013).
Most of these units have been evaluated to be representative of the Judithian NALMA through comparisons of the mammalian fauna (Kielan-Jaworoswka et al. 2004). Radiometric dates have been obtained for some of these units. The Kaiparowits Formation has been estimated to be between 76.1 and 74 Ma (Roberts et al. 2005). The Dinosaur Park Formation has been dated at 75.3 Ma (Eberth 2005). The top of the Oldman Formation has been dated at 76.5 Ma (Eberth and Hamblin 1993). The fossil-producing horizons of the “El Gallo” Formation have been dated at 73.59–72.45 Ma (Renne et al. 1991). The Two Medicine Formation has been dated at approximately 74 Ma at the top of the unit and 80 Ma at the bottom (Rogers et al. 1993). The Fruitland Formation is constrained by a radiometric date of 75.56 Ma near the base of the unit and 74.11 Ma at the top of the unit (Fassett and Steiner 1997). The Judith River Formation in Montana appears to be among the oldest of the Judithian units with a radiometric age estimate of 78 Ma (Goodwin and Deino 1989), but it is not clear how the sampling locality for the ash compares stratigraphically to the horizon producing squamates. The Cerro del Pueblo Formation has no radiometric dates, but has been determined to be no older than 72.5 Ma based on magnetostratigraphic estimates (Eberth et al. 2004). This makes this unit one of the youngest of the “Judithian” faunas.
Maastrichtian
This is the horizon (approximately equal to the Lancian NALMA) from which the first Mesozoic squamate fossils from North American were recovered (Marsh 1892). The known squamate-producing rock units include the Scollard (southern Alberta), Frenchman (Saskatchewan), Hell Creek (central-eastern Montana), Lance (eastern Wyoming), North Horn (central Utah), Laramie (east central Colorado), and Kirtland (northwest New Mexico) formations. Most of these localities have been interpreted to be terrestrial deposits ranging from fluvial to swampy (Carpenter 1979; Condon 2000; Estes 1964; Fassett and Hinds 1971; Fastovsky 1987; Hunt and Lucas 1992; Olsen 1995). I am currently not aware of any reports of squamates from the Maastrichtian-aged sedimentary rocks from the far north (i.e. North Slope of Alaska).
Cenozoic
Paleocene
The Paleocene is reviewed here because some of the squamate lineages established in the Cretaceous survived the terminal Cretaceous extinction event and have been recovered from the early to mid-Paleocene. The relevant rock units are the Polecat Bench (Montana), Tongue River (Montana), Fort Union (Montana and Wyoming), Denver (Colorado), and Goler (California) formations. The effect of the terminal Cretaceous extinction event on North American lizards has been explored by Longrich et al. (2012a) and will only be briefly summarised here.
Squamates from the Late Jurassic–Late Cretaceous
What follows is a review of the current record of squamates from North America by major taxonomic groups. For the most part, I follow the taxonomic conclusions of the most recent publication for each record. In some cases, the most current taxonomic referrals are considered problematic and briefly discussed, but it is beyond the scope of this review to revise the taxonomy in all of these cases.
“Basal squamates” (Fig. 2)
Basal squamates include a series of taxa from the Jurassic and Early Cretaceous that are consistently recovered in phylogenetic analysis as stem squamates, but which have uncertain interrelationships (Evans and Chure 1999; Evans et al. 2002; Reynoso 1998). There are two putative basal squamates from North America: Schilleria utahensis from the Late Jurassic Morrison Formation of Utah (Evans and Chure 1999) and Huehuecuetzpalli mixtecus from the Lower Cretaceous (Albian) Tepexi de Rodrígues locality of the Tlayúa Formation in Puebla, Mexico (Reynoso 1998). Evans and Chure (1999) reported that a phylogenetic analysis places Schilleria at the base of the squamate tree, but did not include their dataset or resulting trees. They do note that the specimen is incomplete and difficult to code for many of its characters thus leaving open the possibility that this taxon may be found to occupy a position within the squamate crown upon recovery and analysis of more complete material. The phylogenetic analysis of Huehuecuetzpalli mixtecus presented by Reynoso (1998) placed this taxon at the base of the squamate tree with relatively high support, a position contrary to the findings of Conrad (Conrad 2008), but supported by Gauthier et al. (2012). Huehuecuetzpalli mixtecus co-occurs with a putative stem scincomorphan (see below) and sphenodontians (Reynoso 1997, 2000; Reynoso and Callison 2000). In North America, the combination of primitive lizards and sphenodontians is typical only of lepidosauromorphan faunas from the Late Jurassic, but in the Cretaceous it is only known for the Tlayúa Formation. Based on the geology and fauna, the Tepexi de Rodrígues locality has been interpreted as near-shore marine deposit (Kashiyama et al. 2004) containing rare tetrapods from a localised, archipelago-based refugium that appears to have preserved a relictual fauna similar to that of the Late Jurassic of North America (Evans 2003; Reynoso and Callison 2000). It is also likely that differences in depositional environment (marine vs. fluvial), geographical separation, latitude, and possible non-trivial differences in age from the poorly constrained “Aptian–Albian” localities to the north may also contribute to the differences between these faunas.

Temporal and geographic distribution of “basal” squamates
Iguania (Fig. 3)
Iguanians in the Old World are known from as early as the Jurassic of India (Evans et al. 2002), but the earliest reports of North American fossils referable to Iguania are putative iguanids from the Late Cretaceous. The oldest of these is Cnephasaurus locustivorus and several unidentified jaw fragments from the upper Santonian Milk River Formation of southern Alberta, Canada (Gao and Fox 1996). The ventrally concave arc formed by the maxillary tooth row of Cnephasaurus locustivorus was noted by Gao and Fox (1996) to be similar to the iguanid Crotaphytus. A similar differential crown height along the maxillary tooth row is also present in the teiids Tupinambis and Callopistes, and the morphology in Cnephasaurus most closely matches the dental asymmetry of the scincoidean Dimekodontosaurus madseni from the early Cenomanian (Nydam 2002). Longrich et al. (2012a) placed Cnephasaurus in their “Polyglyphanodontidae”. While they do not provide justification for this referral, it is tentatively supported by the presence of a well-developed, medially directed crista transversalis enclosing a posteriorly directed shallow excavation that is also present on the maxillae of Chamops, Leptochamops, and Bicuspidon (personal observation). Support for referral to Iguanidae is insufficient and I tentatively refer Cnephasaurus to Chamopsiidae (below).
The indeterminate jaw fragments described by Gao and Fox (1996: figs. 5D–F) from the Milk River Formation are more convincingly iguanid in possessing a dental morphology similar to that of the iguanid Anchaurosaurus gilmorei from the mid-Campanian of Mongolia (Gao and Hou 1995). The specimens are otherwise too incomplete for further evaluation.
Several additional unnamed jaw fragments from the Frenchman Formation (Maastrichtian) of southern Saskatchewan have also been referred to Iguanidae (Gao and Fox 1996). Two of these specimens, SMNH P1927.885 and SMNH P1927.998 (Gao and Fox 1996: figs. 5H, I), are indistinguishable from specimens of the chamopsiid Leptochamops denticulatus that Gao and Fox (1996) report from the same locality. The crown morphology of the teeth of the other specimens does appear to be iguanian in having tall, narrow tooth shafts, and very well developed accessory cusps on the crown. With the exception of possible L. denticulatus specimens, and pending recovery of better material or presentation of evidentiary support for alternative interpretation, the referral of the specimens from the Frenchman Formation to Iguanidae is tentatively retained (contra Longrich et al. 2012a).
Longrich et al. (2012a) named a new putative iguanid, Pariguana lancensis, from the Lance Formation (Maastrichtian) of Wyoming based on a partial mandible. This taxon differs from those described by Gao and Fox (1996) in having closely spaced teeth with crowns that are nearly chisel-like in medial view with only diminutive accessory cusps that flank the tooth crown. The justification for referral of this taxon to Iguanidae is the restriction of the Meckelian fossa by the ventral margin of the dentary (Longrich et al. 2012a: supplementary information). The same morphology of the dentary, as well as a generally similar (though not identical) dentition, is also found in the contemporaneous contogeniid scincoidean Contogenys sloani (Estes 1969b; Nydam and Fitzpatrick 2009). The identification of Pariguana lancensis as an iguanid is considered tentative, but if confirmed would indicate that, by the terminal Cretaceous, there were at least two iguanians in the northern latitudes North America. None of these these putative iguanians appear to have survived the terminal Cretaceous extinction (Longrich et al. 2012a) as they are absent from known Paleocene squamate faunas.
In a recent abstract, DeMar et al. (2012) reported an articulated iguanian from the upper Campanian Two Medicine Formation of Montana. Initial phylogenetic analysis indicates that this specimen is closely related to corytophanids and polychrotids. If confirmed, this specimen will be an important addition to the North American fossil squamate record. Addition skeletons of lizards are also known to occur at the Two Medicine Formation locality (DeMar, personal communication).
The only known Paleocene record is of a possible iguanian is Swainiguanoides milleri from the Fort Union Formation in Wyoming (Sullivan 1982). Evidence supporting the referral of Swainiguanoides to Iguanidae was considered weak by Estes (1983a), but Smith (2009) thought it convincing. If substantiated as an iguanian, Swainiguanoides milleri indicates that iguanians straddled the K/T boundary in North America though no specific or generic taxa are known to have survived the terminal Cretaceous extinction (Longrich 2012a). The earliest indisputable iguanids in North America are from the early Eocene (e.g. Smith 2009).

Temporal and geographic distribution of Iguania
Borioteiioidea
Nydam et al. (2007) erected the Borioteiioidea as a clade of teiid-like scincomorphans from the Late Cretaceous of North America and Asia that formed a sister taxon of Teiidae. A similar composition was recovered by the phylogenetic analyses—including only the large-bodied taxa—of Conrad (2008), using Polyglyphandontidae Gilmore, 1942a for the combination of Asian and North American forms. Alternatively, Nydam et al. (2007) used Polyglyphanodontinae Estes, 1983b for the clade containing the North American and Asian large-bodied taxa (see the “Teiidae” of Gao and Norell 2000) and erected the Polyglyphanodontini for a North American radiation of Polyglyphanodontinae distinguished by the shared possession of transversely oriented teeth. Additionally, Chamopsiidae was recognised as an additional radiation of borioteiioid lizards in North America (Nydam et al. 2010). Wiens et al. (2010), using a total evidence analysis (fossils and molecular data for living squamates), found the polyglyphanodontines Polyglyphanodon, Macrocephalosaurus (= Gilmoreteius; Langer 1998), and Sineoamphisbaena within crown Teiidae. Longrich et al. (2012a) combined Chamopsiidae with Polyglyphanodon and Peneteius for their Polyglyphanodontidae. A massive phylogenetic analysis by Gauthier et al. (2012) recovered “Polyglyphanodontia” (five Asian taxa plus Polyglyphanodon sternbergi from North America; no other North American taxa were included in the analysis) as stem scleroglossans and not scincomorphans. This finding is consistent with two of the trees recovered by Lee (2005): figs. 2, 7), but contrary to Lee’s strict consensus tree (fig. 9) and also contrary to the variable findings of scincomorphan affinities for polgyphanodontines in analyses using smaller datasets (Kearney 2003a; Nydam et al. 2007). The characterisation by Gauthier et al. (2012: 27) that Nydam et al. (2010) describe polyglyphanotontians as “less abundant” in North America than Asia is incorrect because Nydam et al. (2010:1090) recognised that Chamopsiidae and Polyglyphanodontinae together comprise Borioteiioidea (approximately equivalent to Polyglyphanodontia of Gauthier et al. 2012) with nearly a dozen North American taxa in the group. I follow the systematic conclusions of Gauthier et al. (2012), though it is likely that the systematic position of the borioteiioid lizards will continue to shift as additional specimens, particularly those from Asia are better understood. Regardless of the taxonomic model or their systematic position within or outside of Scincoidea, it is clear that polyglyphanodontine and chamopsiid lizards are an important component of the North American fauna during the Cretaceous.
Polyglyphanodontini (Fig. 4)
The Polyglyphanodontini (sensu Nydam et al. 2007) from North America include only those taxa that share transversely oriented, interdigitating teeth in the upper and lower jaws; Bicuspidon, Dicothodon, Paraglyphanodon, Peneteius, and Polyglyphanodon. The oldest known representatives are Bicuspidon numerosus and Dicothodon moorensis from the Cedar Mountain Formation (lower Cenomanian) of central Utah (Nydam 1999; Nydam and Cifelli 2002b). In North America, the only other known occurrence of Bicuspidon is a new species from the Dakota Formation (middle Cenomanian) of southern Utah (Nydam 2013). Dicothodon has a more extensive geographical and temporal record including Dicothodon sp. from the Iron Springs Formation (middle Cenomanian) of southern Utah (Nydam 2013), D. cifellii in the Smoky Hollow Member of the Straight Cliffs Formation (Turonian) of southern Utah (Nydam et al. 2007), and D. bajaensis from the “El Gallo” Formation (upper Campanian) of Baja California (Estes 1983a; Nydam 1999; Nydam et al. 2007). More complete skeletal material for D. bajaensis has been discovered (Montellano et al. 2005), but not yet described. McCord and Gillette (2005: 100) and McCord et al. (2001) referred an isolated large and heavily eroded vertebra from the Fort Crittenden Formation to Polyglyphanodontinae (sensu Gilmore 1940). The specimen has not been figured and the description indicates that only the centrum remains, so the record is noted here as highly tentative. If confirmed, this specimen would currently represent the only record of a squamate from the Mesozoic of Arizona, the others having been rejected by McCord et al. (2001). Also from the late Campanian are the earliest records of the mammal-like Peneteius with Peneteius sp. from the Cerro del Pueblo Formation in Coahuila (Martinez 2010) and Aguja Formation of southern Texas (Nydam et al. 2007) and Peneteius saueri from the Kaiparowits Formation of southern Utah (McCord 1998; Nydam et al. 2007). The youngest records of Peneteius are the Maastrichtian P. aquilonius from the Hell Creek Formation of Montana (Bryant 1989; Estes 1969a) and Naashoibito Member of the Kirtland Formation in New Mexico (Williamson and Weil 2004). Also from the Maastrichtian is the large-bodied Polyglyphanodon sternbergi from the North Horn Formation of central Utah which co-occurs with the more diminutive polyglyphanodontine taxa Paraglyphanodon gazeni and Paraglyphanodon utahensis (Gilmore 1940, 1942a, 1943).
All these taxa appear to have incorporated a significant degree of oral food processing through use of relatively precise interdigitation of the upper and lower tooth rows (Gilmore 1942a; Nydam and Cifelli 2002b, 2005; Nydam et al. 2000, 2007). While the interrelationships within Polyglyphanodontini are not yet clear, the teeth of Dicothodon are a logical morphological intermediate between (1) the simple conical teeth with a transverse apical ridge found in Bicuspidon and the complex, multicuspate, bulbous teeth of Peneteius saueri (the basal constriction of the teeth is secondarily lost in P. aquilonius), and (2) Bicuspidon and the expanded chisel-like teeth of Polyglyphanodon (Nydam and Cifelli 2005; Nydam et al. 2007). As pointed out by Nydam et al. (2013), pre-Maastrichtian representatives of Polyglyphanodontini are restricted to more southern latitudes of North America. The absence of Polyglyphanodontini from the well-sampled Milk River, Oldman, Judith River, Dinosaur Park, Frenchman, or Lance formations is unlikely to be a sampling error since the mammal-like multicuspid morphology of the teeth of many of these lizards are very distinct and most of the investigations of the microvertebrate assemblages in these units have been led by mammal specialists looking for and recovering numerous mammal specimens with similarly distinctive multicuspid teeth.

Temporal and geographic distribution of Polyglyphanodontini (i.e. transversely-toothed borioteiioids; sensu Nydam et al. 2007)
Chamopsiidae (Fig. 5)
These lizards are only known from the Late Cretaceous and are characterised by heterodont dentition in which the teeth of the posterior portion of the tooth row are commonly trifid, and in possessing a massive U-shaped mandibular symphysis of dentary (Nydam et al. 2010). Currently there are 11 known genera and 14 species of chamopsiid lizards described from the Upper Cretaceous units in North America. Each of these counts would increase by one if the “polyglyphanodontian” Obamadon gracilis described by Longrich et al. (2012a) is included. Longrich et al. (2012a: supplementary information) excluded Obamadon from Chamopsiidae based on the lack of a heavily developed mandibular symphysis, but conceded the possibility that it may be a “basal chamopsiid”. Based on small size of the specimen and the evidence of active tooth replacement, I agree with the supposition of Longrich et al. (2012a) that the specimen is likely a juvenile form of a chamopsiid. The detailed photographs provided by Longrich et al. (2012a) of the type of Obamadon gracilis are very similar to many known specimens of Leptochamops, which similarly has the weakest development of the U-shaped mandibular symphysis (personal observation of USNM and UALVP specimens) or Tripennaculus (based on dentition, the condition of the mandibular symphysis is unknown; Nydam and Voci, 2007). Longrich et al. (2012a: fig. 6) included all of the chamopsiid taxa within their “Polyglyphanodontidae”, but differentiated chamopsiids and other “polyglyphanodontians” in their supplementary information and included Peneteius aquilonius within Chamopsiidae (Longrich et al. 2012a: 21396, supplementary information) without explanation.
The earliest known chamopsiid is Harmondontosaurus emeryensis form the Cedar Mountain Formation (lower Cenomanian) of central Utah (Nydam 2002; Nydam et al. 2010). The next occurrence of a chamopsiid is Chamops sp. from the Smoky Hollow Member of the Straight Cliffs Formation (Turonian) of southern Utah (Nydam 2013). No additional chamopsiids have been recovered as yet from any of the other extensive Cretaceous-aged sediments of southern Utah below the upper Campanian Kaiparowits Formation (Nydam 2013).
Upper Santonian specimens from the Milk River Formation of Alberta include Chamops sp., Sphenosiagon simplex, and possibly Glyptogenys ornata (Gao and Fox 1991, 1996). I also tentatively transfer Cnephasaurus locustivorus from the Milk River Formation (Gao and Fox 1996) from Iguanidae to Chamopsiidae (see above). From the mid-Campanian Peng et al. (2001) reported Leptochamops sp. from the Foremost and Oldman formations and Socognathus unicuspis from the Oldman Formation. From the late Campanian, Nydam and Voci (2007) identified Meniscognathus molybrochoros, Tripennaculus eatoni, Chamops cf. C. segnis, and Leptochamops sp. in the Kaiparowits Formation of southern Utah. Armstrong-Ziegler (1978, 1980) and Sullivan (1981) identified Chamops segnis from the Fruitland Formation of New Mexico (Gao and Fox 1996: 25–26 suggest that Chamops sp. or Chamopsiidae indet. are more appropriate for these specimens). The only confirmed chamopsiid described and figured from the upper Campanian Mesaverde Formation of Wyoming is a dentary of Leptochamops thrinax (DeMar and Breithaupt 2006). They also report in a faunal list the presence of L. denticulatus, cf. Chamops, and Meniscognathus sp. (DeMar and Breithaupt 2006: 37), but provide no other information upon which to confirm these identifications. From the upper Campanian Judith River Formation of Montana are several specimens referable to Chamops sp. (Gao and Fox 1996; Sahni 1972), but newly recovered specimens in the collections of the UCMP indicate the presence of possibly 3–5 chamopsiid taxa (Nydam, work in progress). Upper Campanian specimens from the Dinosaur Park Formation include Socognathus unicuspis, Sphenosiagon simplex, Glyptogenys ornata, Gerontoseps irvinensis, and Leptochamops thrinax (Gao and Fox 1991; Gao and Fox 1996). In their fig. 39 Gao and Fox (1996) also listed Chamops as occurring at this horizon, which must refer to the nearly complete mandible from a crocodilian fecal pellet reported by Waldman (1970). Due to the eroded tooth crowns and lack of a complete mandibular symphysis, this specimen is better referred to cf. Chamopsiidae genus indet. Socognathus unicuspis has also been recovered from the upper Campanian Wapiti Formation of west central Alberta (Nydam et al. 2010).
There are fewer chamopsiid taxa known to occur in localities from the Maastrichtian. Leptochamops sp. has been reported from the North Horn Formation of central Utah (Cifelli et al. 1999b). Taxa from the Lance Formation of Wyoming include Chamops segnis, Leptochamops denticulatus, Meniscognathus altmani, and Haptosphenus placodon (Estes 1964; Marsh 1892). Taxa from the Hell Creek Formation include Chamops segnis, Leptochamops denticulatus, Meniscognathus altmani, and Haptosphenus placodon (Estes et al. 1969) with the possible addition of Socognathus unicuspis (Gao and Fox 1996). Taxa from the Frenchman Formation of southern Saskatchewan include Chamops segnis, Leptochamops denticulatus, Haptosphenus placodon, and Meniscognathus altmani. Taxa from the Scollard Formation of central Alberta include Chamops segnis, Haptosphenus placodon, and Stypodontosaurus melletes.
Unlike their polyglyphanodontine counterparts, the distribution of chamopsiids trends to more northern latitudes of North America. This is most notable during the late Campanian for which there is the greatest number of comparable localities (Nydam et al. 2013). No demonstrable chamopsiids are known from south of the New Mexico and Utah occurrences (see Nydam et al. 2013 regarding the putative report of Chamops from the Aguja Formation). An important caveat here is the limited investigations of Upper Cretaceous sediments in more southern latitudes, but while the extensive collections of the Aguja Formation and more modest collections of the Cerro del Pueblo Formation has resulted in the recovery of a numerous lizard taxa, including Peneteius, there is no convincing evidence of chamopsiids. Even if a chamopsiid is recovered from these units, it is still clear that the greatest diversity of the group occurs in southern Canada and Montana/Wyoming with decreasing diversity in successively more southern localities.

Temporal and geographic distribution of Chamopsiidae
Scincoidea
Scincoidean lizards are among the most common from the Jurassic and Cretaceous of North America, but have also been among the most difficult to resolve taxonomically. Many have historically been placed within modern families (e.g. Estes 1964, 1983b; Gilmore 1928, 1942a) or later reinterpreted to be part of extinct families composed only of fossil forms (Estes 1983a; Nydam and Fitzpatrick 2009; Nydam et al. 2007). The lack of resolution is primarily due to the fragmentary nature of the specimens, which are mostly broken jaws and isolated osteoderms. As more complete remains are recovered, there will hopefully be an improvement in our understanding of the systematics of these lizards.
Paramacellodidae and specimens of paramacellodid-cordylid grade dentitions (Fig. 6)
The genus Paramacellodus is an early scincoidean most commonly found in the Late Jurassic and earliest Cretaceous of North America and Europe and is considered one of the lizards that was part of a pan Laurasian fauna during that time (Estes 1983b; Evans 1995, 1998, 2003). Specimens referable to Paramacellodus have been identified from the Morrison Formation (Upper Jurassic) as Paramacellodus in Colorado (Evans 1996), Paramacellodus sp. cf. P. oweni in Utah (Evans and Chure 1998a, b, 1999), and Paramacellodus sp. in Wyoming (Prothero and Estes 1980) and from the Lower Cretaceous Cloverly Formation (Aptian–Albian) as cf. Paramacellodus keebleri in Montana (Nydam and Cifelli 2002a). An additional taxon tentatively referred to the Paramacellodidae is Atokasaurus metarsiodon from the Antlers Formation (Aptian–Albian) of Oklahoma (Nydam and Cifelli 2002a).
No paramacellodids have been described from the Late Cretaceous of North America, but there are several jaw and partial jaw specimens from multiple Upper Cretaceous horizons that possess dental morphology very similar to that described and illustrated for Paramacellodus and other paramacellodid lizards (Evans and Searle 2002; Kosma 2004; Nydam 2013; Richter 1994) and many cordylids (Kosma 2004). While useful for distinguishing these fossil forms from scincoideans with more distinct or unique dental characteristics (e.g. polyglyphanodontids, chamopsiid, contogeniids), these dental features are insufficient to independently distinguish taxa among most of the specimens within this grade. As such, Nydam (2013) combined the named taxa and unnamed morphotypes together within the informal “scincomorphans of paramacellodid–cordylid grade”. The grouping of indeterminate scincoideans with a paramacellodid–cordylid grade dentition is not meant to imply any form of monophyletic association and the association is probably paraphyletic. This grouping was used by Nydam (2013) to organise, present, and demonstrate the morphological diversity of the lizard specimens from the Late Cretaceous of southern Utah otherwise too incomplete for systematic treatment. In many cases, such fragmentary specimens are left out of faunal assessments and remain undescribed in many collections (Nydam, personal observation). Some of these specimens were sufficiently complete to differentiate them from known genera and species of lizards, but cannot be placed in higher order taxa.
The oldest specimens referable to the paramacellodid–cordylid grade are several isolated and unnamed specimens described by Winkler et al. (1990) from the Lower Cretaceous (Aptian–Albian) Trinity Group of northern Texas. This includes a specimen they originally referred to Anguidae, but was moved to Scincomorpha by Gao and Fox (1996: 74). Also included are the unnamed scincomorphan specimens from the Cloverly and Antlers formations described by Nydam and Cifelli (2002a). From the lower Cenomanian Cedar Mountain Formation of Utah is the jaw specimen referred to Pseudosaurillus sp. (Nydam 2002), though the generic referral should be treated as tentative. From the middle Cenomanian Dakota Formation of Utah, there is a new genus and species and the three unnamed morphotypes “Dakota A, B, C” (Nydam 2013). Four morphotypes, “SHMbrSCFm A–D”, are described from the Turonian Smoky Hollow Member of the Straight Cliffs Formation (Nydam 2013). Another new genus and species has been identified from the John Henry Member (Santonian) of the Straight Cliffs Formation of Utah (Nydam 2013). I also refer the putative scincid Aocnodromeus corrugatus (Gao and Fox 1996) from the Milk River Formation (upper Santonian) of Alberta as well as the unnamed “?Cordylidae” from the same unit (Gao and Fox 1996) to this grade based on my observations of these specimens. The only specimen from the lower Campanian that is referable to this grade is the unnamed morphotype “Iron Springs A” from the uppermost Iron Springs Formation, Utah (Nydam 2013). From the upper Campanian, there are four morphotypes, “Kaiparowits A–D”, from the Kaiparowits Formation, Utah (Nydam 2013). Additional specimens that were tentatively referred to “?Cordylidae” by Gao and Fox (1996) recovered from the Dinosaur Park Formation (upper Campanian) of Alberta, the Frenchman Formation (Maastrichtian) of Saskatchewan, and the Hell Creek Formation (Maastrichtian) of Montana all fit this grade as well.

Temporal and geographic distribution of Paramacellodus (“P”) and scincoideans of paramacellodid–cordylid grade dentition
Contogeniidae (Fig. 7)
Contogeniid lizards (sensu Nydam and Fitzpatick 2009) are xantusiid-like lizards with distinctive dentition characterised by chisel-like tooth crowns with apical grooves along the mesial and distal aspects of the tooth tips. These lizards are excluded from Xantusiidae based on the retention of an open Meckelian fossa of the dentary and retention of an independent splenial. The earliest contogeniid taxon is a newly identified species of Utahgenys (Nydam 2013; previously identified as Utahgenys sp. of Nydam and Fitzpatrick 2009) from the Dakota Formation (middle Cenomanian) of southern Utah. Utahgenys evansi is known from the Smoky Hollow Member of the Straight Cliffs Formation (Turonian), also of southern Utah (Nydam and Fitzpatrick 2009). The largest of the known contogeniids is Palaeoscincosaurus pharkidodon from the upper Campanian Kaiparowits Formation of southern Utah (Nydam and Fitzpatrick 2009). Estes (1969b) described Contogenys sloani (see also Bryant 1989; Nydam and Fitzpatrick 2009) from the Hell Creek Formation (Maastrichtian) of Montana. Contogeniids survived the terminal Cretaceous extinction and two taxa are known from the Paleocene: C. ekalakaensis from the Tongue River Formation of Montana (Estes 1969b; Nydam and Fitzpatrick 2009) and P. middletoni from the Denver Formation in Colorado (Sullivan and Lucas 1996).
Sullivan (1982) referred a partial maxilla from the Fort Union Formation of the Paleocene of Montana to “C. sloani” based on the presence of a posterolateral expansion of the element. This cannot be considered to be a sufficient anatomical distinction because both contogeniids and xantusiids share this morphology (Nydam and Fitzpatrick 2009; Nydam et al. 2013; Nydam, personal observation). Based on the similarity in size with a dentary of the xantusiid Palaeoxantusia fera (see below) from the same locality (Sullivan 1982: figs. 4, 5), it is possible that the maxilla belongs instead to P. fera, but confirmation of either possibility requires more complete material.

Temporal and geographic distribution of Contogeniidae
Xantusiidae (Fig. 8)
Xantusiids, or night lizards, make their potentially earliest appearance in North America in the form of Catactegenys solaster from the late Campanian Aguja Formation of southern Texas (Nydam et al. 2013). The evidence supporting referral of Catactegenys to Xantusiidae is the presence of a fused spleniodentary as opposed to the independent splenial and dentary of Contongeniidae (Nydam et al. 2013). The phylogenetic analyses of Nydam et al. (2013) recovered Catactegenys as a xantusiid, but the results are not robust. If Catactegenys is confirmed as a xantusiid, it would indicate that the family appeared in North America during the Late Cretaceous and, like the apparently closely related Contogeniids, survived the terminal Cretaceous extinction. Palaeoxantusia fera has been reported from the Paleocene Fort Union Formation of Montana (Estes 1983a; Sullivan 1982, 1991). Xantusiids currently have a fragmented distribution in the southwest United States, Mexico, Central America, and Cuba (Crother et al. 1986).
Gauthier et al. (2012) argued that the enigmatic Tepexisaurus tepexii, originally interpreted to be a basal scincoidean (Reynoso and Callison 2000), may actually be a stem xantusiid (although the closed, but unfused Meckel’s canal is comparable to the condition in Contogeniidae; sensu Nydam and Fitzpatrick 2009). If this interpretation is accurate, then the origins of lineage leading to Xantusiidae extends back to the Albian of Mexico.

Temporal and geographic distribution of Xantusiidae. ? = tentative record
Indeterminate scincoidean fossils (Fig. 9)
Fossil forms that can be allied with Scincoidea, but not placed with confidence within any higher taxon within Scincoidea (referral to the family Scincidae, can be inferred from isolated jaw fragments, but the lack of diagnostic characters prevents confident assignment to the family) nor do they fit the paramacellodid-cordylid grade of tooth morphology. These taxa include Saurillodon from the Upper Jurassic Morrison Formation (Evans 1996; Evans and Chure 1999) and the unusual “basal scincoid” Tepexisaurus tepexii from the Tlayúa Formation (Albian) (possbily a stem xantusiid; Gauthier et al. 2012, see above). From the Aptian–Albian of Oklahoma (Antlers Formation) and Wyoming (Cloverly Formation) is Ptilotodon wilsoni (Nydam and Cifelli 2002a; Oreska et al. 2013). This taxon was referred to Teiidae in both cases, but the specimens illustrated provide no features specifically diagnostic of that family (see also Nydam et al. 2007). Oreska et al. (2013) also included in their faunal list for the Cloverly Formation in Wyoming two morphotypic specimens, “Teiidae A” and “Teiidae B”. As with Ptilotodon wilsoni, these specimens lack features specifically diagnostic of Teiidae. From the early Cenomanian are Dimekodontosaurus madseni, the possible “scincid” Bothriogenys mysterion, and several unnamed “scincomorphan” jaw fragments from the Cedar Mountain Formation (Nydam 2002). From the middle Cenomanian are a new genus and species of a possible scincid and the unnamed morphotype “Dakota D”, from the Dakota Formation (Nydam 2013). From the Turonian are unnamed morphotypes “SHMbrSCFm E–H”, from the Smoky Hollow Member of the Straight Cliffs Formation (Nydam 2013). From the Santonian is the unnamed morphotype “JHMbrSCFmA”, from the John Henry Formation of the Straight Cliffs Formation (Nydam 2013). From the Santonian is the indeterminate scincoidean Penemabuya antecessor, Milk River Formation (Gao and Fox 1996). From the low-mid Campanian is the unnamed morphotype “Wahweap A”, from the Wahweap Formation, Utah (Nydam 2013). From the upper Campanian are the unnamed morphotypes “Kaiparowits E–G”, from the Kaiparowits Formation (Nydam 2013), Kleskunsaurus grandprairiensis from the Wapiti Formation of Alberta (Nydam et al. 2010), and Apsgnathus triptodon and a ?scincid from the Aguja Formation of southern Texas (Nydam et al. 2013).
Osteoderms referable to Scincoidea have also been recovered from many lizard-producing localities and almost certainly belong to taxa identified based on jaw elements, but specific referrals are impossible in the absence of clear association. The only specimens attributed to a named taxon are those referred to Paramacellodus from the Morrison Formation in Utah (Evans and Chure 1999). Indeterminate scincoidean osteoderms are also reported from the Early Cretaceous of Oklahoma, Montana, and Wyoming (Nydam and Cifelli 2002a), the early Cenomanian of central Utah (Nydam 2002), the middle Cenomanian, Turonian, Santonian, early Campanian, and late Campanian of southern Utah (Nydam 2013), the late Campanian of southern Texas (Nydam et al. 2013; Rowe et al. 1992), and the late Campanian of southern Alberta (Peng et al. 2001). There is little known taxonomic value of osteoderms in scincoideans though some attempts have been made to characterise these elements for some European taxa (Broschinski 2000; Broschinski and Sigogneau-Russell 1996; Richter 1994), but these specimens are skeletons with associated osteoderms. It is not clear if the absence of scincoidean osteoderms in localities other than those noted above is a consequence of exclusion due to lack of taxonomic clarity or if they are actually absent from the collections of these faunas.

Temporal and geographic distribution of scincoideans of indeterminate taxonomic referral
Anguimorpha
The fossil record for anguimorphan lizards is similar to that of scincoidean lizards in extending from the Late Jurassic into the Cenozoic. Unlike the scincoideans, there is a generally agreed better resolution of the systematics and taxonomy for anguimorphan lizards from the Mesozoic of North America. Even so, these taxa are represented by similarly fragmentary specimens and more complete materials may support alternative referrals.
Dorsetisauridae (Fig. 10)
Dorsetisaurus is a conspicuous anguimorphan lizard from the Late Jurassic and Early Cretaceous of Europe (Broschinski 2000; Broschinski and Sigogneau-Russell 1996; Estes 1983a; Evans 2003). The North American distribution of this taxon is limited to jaws of Dorsetisaurus sp. from the Morrison Formation (Late Jurassic) of Colorado, Utah, and Wyoming (Callison 1987; Evans and Chure 1998a, 1999; Prothero and Estes 1980). Eaton et al. (1999a) included “cf. Dorsetisaurus” in their faunal list from the Smoky Hollow Member of the Straight Cliffs Formation (Turonian) of southern Utah, but Nydam (2013) transferred the specimen to a new taxon (see below).

Temporal and geographic distribution of the anguimorphan genus Dorsetisaurus
Xenosauridae (Fig. 11)
Xenosaurids from the Cretaceous of North America have a long reported history, but confident referral of specimens to the family is tentative at best. This is because most of the diagnostic characters for Xenosauridae (sensu Estes 1983a) are for skull features that cannot be evaluated from isolated elements and jaws. The one dental character cited by Estes (1983b) is that tooth succession is variably direct or interdental, which is of limited use for diagnosing isolated jaws. Estes et al. (1988) gave 12 synapomorphies for Xenosauridae, but of these only four pertain to specific cranial elements, two for osteoderms, and none apply to jaws/teeth. Conrad (2008) identified a cranial feature (exposure of ectopterygoid relative to maxilla) and incipient cusps on the teeth as synapomorphies of Xenosauridae. Conrad et al. (2010) did not define Xenosauridae, but instead grouped Xenosaurus with Carusia and anguids together in an unnamed clade. Gauthier et al. (2012) recovered Xenosauridae in their analysis, but do not identify the synapomorphies. For most of the specimens below, referral to Xenosauridae (sensu Estes et al. 1988) remains tentative, but more likely than referral to any other known taxon.
The earliest specimens referred to Xenosauridae are jaws of a new genus and species from the middle Cenomanian Dakota Formation of southern Utah (Nydam 2013). Additional fragmentary jaws from the Smoky Hollow Member of the Straight Cliffs Formation of southern Utah are referred to the same new genus from the Dakota Formation (Nydam 2013). This includes a specimen originally identified by Eaton et al. (1999a) as Dorsetisaurus. Gao and Fox (1996) described unnamed xenosaurid specimens from the Milk River Formation (upper Santonian) of southern Alberta. The next records are from the upper Campanian with specimens referred to ?Exostinus sp. recovered from the Kaiparowits Formation of southern Utah (Nydam 2013) and the Dinosaur Park Formation of southern Alberta (Gao and Fox 1996). Sahni (1972) referred some specimens from the upper Campanian Judith River Formation of Montana to Exostinus lancensis and Exostinus sp. Gao and Fox (1996: 66) rejected Sahni’s generic and specific identifications, but did conclude that one of the specimens from the Judith River Formation may be a new genus of xenosaurid. Osteoderms identified as cf. Xenosauridae are described from the upper Campanian Aguja Formation of southern Texas (Nydam et al. 2013), but no referable cranial elements are known from the unit. A xenosaur not referable to Exostinus or Restes is present in the Marshalltown Formation (upper Campanian) of New Jersey (R. Denton, personal communication; not included in Fig. 11).
Maastrichtian records of xenosaurids are all for Exostinus lancensis with occurrences in the Lance Formation of Wyoming (Gao and Fox 1996; Gilmore 1928) and the Hell Creek Formation of Montana (Estes et al. 1969; Gao and Fox 1996). There are several referred cranial elements for Exostinus lancensis (Gao and Fox 1996) and referral of this taxon to Xenosauridae is relatively well supported. Estes (1976) referred a specimen of a maxilla recovered from the Fort Union Formation of Montana to Exostinus lancensis. This same specimen was later cited as simply Exostinus by Longrich et al. (2012a: supplementary information). Sullivan (1982) also referred several jaw fragments from the Swain Quarry in the Fort Union Formation (Paleocene) of Montana to Exostinus cf. E. lancensis. In his comprehensive analysis of morphology Bhullar (2010, 2011) found that E. lancensis is closely related to Xenosaurus, but he also considers E. lancensis a problematic taxon, possibly chimaeric and in need of additional study. An additional xenosaurid, Restes rugosus, has also been recovered from the Fort Union Formation (Gauthier 1982; Gilmore 1942b).

Temporal and geographic distribution of Xenosauridae. ? = tentative records
Anguidae (Fig. 12)
The earliest lizards referable to Anguidae from North America all belong to the genus Odaxosaurus. Estes (1983b) referred Odaxosaurus to Glyptosaurinae, but noted that the only diagnostic features supporting this referral was tooth crown morphology and osteoderm shape. Gao and Fox (1996) provided a comprehensive review of the studies on Odaxosaurus and agreed with Estes that the fossil taxon is glyptosaurine-like, but not convincingly within Glyptosaurinae. The earliest occurrence of Odaxosaurus is Odaxosaurus sp. from the Smoky Hollow Member of the Straight Cliffs Formation (Turonian) of southern Utah (Nydam 2013). Indeterminate anguid specimens from the Milk River Formation (upper Santonian) of southern Alberta (Gao and Fox 1996) appear to be Odaxosaurus, but are missing the diagnostic tooth crowns. Upper Campanian records include Odaxosaurus priscus from the Dinosaur Park Formation (Gao and Fox 1996); O. priscus and a new species of Odaxosaurus from the Kaiparowits Formation of southern Utah (Nydam 2013); Peltosaurus piger from the Judith River Formation (Sahni 1972), which was transferred to Odaxosaurus sp. by Gao and Fox (1996), though they removed many specimens from the referral; Peltosaurus piger from the Mesaverde Formation of Wyoming (DeMar and Breithaupt 2006) which is almost certainly Odaxosaurus piger and not O. priscus (DeMar, personal Communication); O. piger from the Aguja Formation of southern Texas (Nydam et al. 2013); and an unnamed new species of Odaxosaurus from the Cerro del Pueblo Formation in Coahuila (Martinez 2010). Another potential record for Odaxosaurus from the late Campanian includes a jaw fragment identified by Armstrong-Ziegler (1980) as cf. Gerrhonotus from the Fruitland Formation (see Gao and Fox 1996), but Estes (1983b) was convinced the specimen is better referred to cf. Gerrhonotus. The possible presence of a glyptosaurine lizard from the upper Campanian Marshalltown Formation of New Jersey (Denton et al. 1991) is now considered to be an annielline (R. Denton, personal communication, work in progress). An indeterminate squamate from the upper Campanian Bladen Formation of North Carolina (Crane 2011) is a possible glyptosaurine anguid currently under study by Cynthia Crane as part of a larger analysis of the North Carolina Cretaceous fauna (Crane, personal communication).
Odaxosaurus piger is the most common anguid from the Maastrichtian of North America. It has been recovered in the Lance Formation of Wyoming (Estes 1964; Gilmore 1928), Hell Creek Formation of Montana (Bryant 1989; Estes et al. 1969), Laramie Formation of Colorado (Carpenter 1979), Scollard Formation of central Alberta (Gao and Fox 1996), and Frenchman Formation of southern Saskatchewan (Gao and Fox 1996). Cf. ‘Gerrhonotus’ (quotes sensu Longrich et al. 2012a) was also identified from the Lance Formation by Estes (1964, 1983b).
The Paleocene anguid taxon Proxestops jepseni is considered to be a morphological intermediate between the smaller Odaxosaurus from the Cretaceous and the larger Xestops from the Eocene (Gauthier 1982). However, the differential diagnosis distinguishing these two taxa is based solely on difference in osteodermal morphology, elements that are only tentatively referred to Odaxosaurus (see below). The dentition of Proxestops is nearly identical to that of Odaxosaurus, but is also considered a feature diagnostic of glyptosaurine anguids (Gauthier 1982) or primitive to glyptosaurine anguids, but lost in more recent forms (Smith 2009); the latter interpretation agrees with the phylogenetic results of Conrad et al. (2010). Proxestops jepseni has been recovered from the Fort Union Formation of Montana (Estes 1983a; Gilmore 1928, 1942b) and Wyoming (Meszoely 1970), the Tongue River Formation of Montana (Estes 1976, 1983b), the Tullock Formation of Montana (Bryant 1989; Estes 1983a; Sloan and Van Valen 1965) and the North Horn Formation (upper portion) of central Utah (Cifelli et al. 1999b; Gilmore 1942b). Another specimen referable to cf. Gerrhonotus was recovered from the Tongue River Formation (Estes 1976). Sullivan (1981) reported the presence of several anguids from the Nacimiento Formation of New Mexico including Odaxosaurus piger. He later transferred one of the specimens (partial frontal) to a new taxon, Parodaxosaurus sanjuanensis (Sullivan 1986), but still notes the presence of O. piger in the Paleocene of New Mexico as well as the Tongue River Formation of Wyoming (Sullivan 1991; Sullivan and Lucas 1986). However, the specimens used to vouch for the New Mexico record are an opisthotic, a splenial, and a collection of osteoderms with ornamentation varying from vermiculate to tubercular. None of these elements is anatomically comparable to the type material for O. piger, and the presence of the taxon in the Paleocene is dubious considering there are three other named anguid taxa and three unnamed morphotypes (sensu Sullivan and Lucas 1986).
Many of the records of Odaxosaurus have included the referral of thin, quadrangular/trapezoidal, imbricating osteoderms (Bryant 1989; Estes 1964, 1983b; Estes et al. 1969; Gilmore 1928; Meszoely 1970) and rare cranial elements (Estes 1964; Meszoely 1970). Both Gao and Fox (1996) and Nydam (2013) have cautioned that, because the type specimen of Odaxosaurus is only a dentary, referral of the osteoderms and cranial material should be treated as tentative. Gao and Fox (1996) further argued that their maxilla of O. piger preserving an irregular and deeply pitted fused osteoderm was further support of the refutation by Armstrong-Zeigler (1980) of referral of the osteoderms to Odaxosaurus. However, it is not unusual for modern anguids such as Elgaria and Gerrhonotus to have similar osteoderms low on their maxilla, whereas the rest of the osteoderms of the skull and body are plate-like and more subtly ornamented (Nydam, personal observation). While I still caution that the referrals of skull elements and osteoderms to Odaxosaurus should be treated as tentative, it is also likely that the referrals are correct, but should not be used independent of jaw and tooth morphology for taxonomic or systematic comparisons. Bryant (1989: 46–47) added an interesting observation to this problem when she noted that, in some of the localities in the Tullock Formation (Paleocene), osteoderms referable to Odaxosaurus, and presumably distinct from those of Proxestops (sensu Gauthier 1982) were more common than in localities of the Hell Creek Formation from which the jaws of Odaxosaurus were recovered. This may simply be a sampling error since each of these fossil anguids likely had several hundred osteoderms and only about a dozen cranial elements useful for comparison; it is much more likely that osteoderms will be recovered.

Temporal and geographic distribution of Anguidae
Platynota (Fig. 13)
As indicated by Estes (1983b), this group is often equated with Varanoidea (see Nydam 2000), and this convention also works here for the purposes of reviewing the terrestrial forms from North America. Varanoidea/Platynota typically include Varanidae, Helodermatidae, and Necrosauridae (sensu Estes 1983a; see also Conrad et al. 2010: 40). Also variably included are snakes and mosasaurs, but the arguments back and forth regarding the systematic and taxonomic positions of these groups are well beyond the scope of this paper and I refer the reader to a sample of recent contributions (Caldwell 2007; Caldwell and Lee 1997; Conrad et al. 2010; Konishi and Caldwell 2007; Palci and Caldwell 2007; Rieppel and Head 2004; Rieppel et al. 2003; Tchernov et al. 2000; Wilson et al. 2010; Zaher and Scanferla 2012). The taxonomic status of terrestrial platynotans has also been variable. In the most recent phylogenetic studies, Conrad (2008) recovered a Varanoidea composed (at higher taxonomic ranks) of Monstersauria (sensu Norell and Gao 1997), Helodermatidae, and Varaniformes (= Varanidae + Lanthonotinae). In their 2010 analysis, Conrad et al. restricted their attention to anguimorphans and recovered a Varaniformes composed of a clade of ‘necrosaurs’ (unnamed because they do not use “Necrosauridae”), Lanthonotinae, and Varaninae. However, helodermatids in the form of monstersaurs were recovered outside of their Varaniformes and in a clade with anguids and carusioids (Conrad et al. 2010: fig. 13). Gauthier et al. (2012) recovered a Varanoidea exclusive of mosasaurs and snakes. While they do not identify clades within Varanoidea, they do indicate that they were unable to recover Monstersauria. Using a combined evidence analysis Yi and Norell (2013) recovered a monophyletic Monstersauria as part of a larger polytomous grouping of numerous anguimorpan taxa. For the purposes of this review, all taxa and/or specimens with features that have been used refer them any level of Varanoidea/Platynota exclusive of mosasaurs (not treated here) and snakes (below) will be included under the umbrella of Platynota. Indeed, many specimens referred to named taxa are, in my opinion, too fragmentary to justify generic or specific referral and should be considered Platynota indet. pending the recovery of more diagnostic materials (sensu Nydam 2013; Nydam et al. 2013). I will use the original higher taxonomic referrals in quotations pending a revision of these taxa (B. Barr, work in progress).
The earliest known platynotan is the helodermatid-like (i.e. “monstersaur”) Primaderma nessovi from the Cedar Mountain Formation (lower Cenomanian) of central Utah (Cifelli and Nydam 1995; Nydam 2000). From the Dakota Formation (middle Cenomanian) of southern Utah is an unnamed morphotype based on a small jaw fragment (Nydam 2013). Two additional unnamed morphotypes have been identified by Nydam (2013) from the Smoky Hollow (Turonian) and John Henry (Santonian) members of the Straight Cliffs Formation. An additional Santonian record are specimens identified as the “necrosaurs” ?Parasaniwa and “genus and species new” from the Milk River Formation of southern Alberta (Gao and Fox 1996).
Platynotans reported from the Oldman Formation (lower-mid Campanian) of southern Alberta include cf. Paraderma bogerti, the “varanid” cf. Palaeosaniwa canadensis, and an indeterminate helodermatid based on isolated osteoderms (Peng et al. 2001). The “helodermatid” osteoderms are identical to osteoderms reported as aff. Xenosauridae from the Aguja Formation (Nydam et al. 2013) as well as osteoderms recently recovered from the Judith River Formation (Nydam, work in progress).
Several records of platynotans are known from the late Campanian. From the Cerro del Pueblo Formation of Coahuila are several isolated specimens referred to the “necrosaurid” Parasaniwa cf. P. wyomingensis, osteoderms referred to cf. Parasaniwa, a partial humerus referred to the “varanid” Palaeosaniwa sp., and vertebrae referred to the “helodermatid” Paraderma cf. P. bogerti (Martinez 2010). From the Aguja Formation of southern Texas are specimens of cf. Parasaniwa wyomingensis and indeterminate platynotan vertebrae (Nydam et al. 2013). From the Kaiparowits Formation of southern Utah Nydam (2013) has identified a new species of Parasaniwa and three unnamed, but distinct morphotypes based on jaw fragments. The “necrosaurid” Parasaniwa wyomingensis was listed in the fauna from the Mesaverde Formation of Wyoming (DeMar and Breithaupt 2006). From the Judith River Formation of Montana Sahni (1972) reported the presence of the “parasaniwids” (“necrosaurid”; sensu Estes 1983a) Parasaniwa wyomingensis and Paraderma bogerti (a taxon more frequently allied with helodermatids, e.g. Pregill et al. 1986, or monstersaurs, e.g. Norell and Gao 1997). Neither of the figured specimens appears to be sufficiently complete to justify generic or specific identifications, but both are clearly varanoid. Platynotans from the Dinosaur Park Formation of southern Alberta include the “necrosaurids” Parasaniwa n. sp., cf. P. wyomingensis, the “helodermatids” Labrodioctes montanensis, Palaeosaniwa canadensis and an unnamed form (Gao and Fox 1996).
Platynotans from the Maastrichtian are currently only known from more northerly localities. Taxa recovered from the Lance Formation of Wyoming include Parasaniwa wyomingensis (Estes 1964; Gilmore 1928), Paraderma bogerti (Estes 1964), and Palaeosaniwa canadensis (Estes 1964). From the Hell Creek Formation of Montana, the reported platynotans are Parasaniwa wyomingensis, Paraderma bogerti, and Palaeosaniwa canadensis (Bryant 1989; Estes et al. 1969). From the Scollard Formation of central Alberta, Gao and Fox (1996) reported the presence of Paraderma bogerti, and Parasaniwa wyomingensis. Only Paraderma bogerti and Parasaniwa wyomingensis are reported from the Frenchman Formation of southern Saskatchewan by Gao and Fox (1996), but the skull of a new platynotan taxon has been also recently been recovered (Barr 2012, work in progress). Longrich (2012a) erected Cemeterius monstrosus based on a fragmentary portion of dentary of a very large varanoid from the Hell Creek Formation.
No platynotans from the terminal Cretaceous have been recovered from the Paleocene (Longrich et al. 2012a). Platynotans from the late Paleocene include Provaranosaurus acutus (Gilmore 1942b), vertebrae tentatively referred to cf Saniwa (Sullivan 1982), and a jaw fragments referred to an unidentified varanoid taxon (Sullivan 1991) all from the Fort Union Formation. Cf Provaranosaurus has also been noted for the Tongue River Formation of Montana (Estes 1976).

Temporal and geographic distribution of Platynota
Indeterminate anguimorphan specimens (Fig. 14)
Specimens that are mostly likely referable to Anguimorpha, but otherwise taxonomically and systematically indeterminate have been recovered from several horizons throughout the Cretaceous. A jaw fragment preserving a single, Dorsetisaurus-like tooth was reported from the Antlers Formation (Aptian–Albian) of Oklahoma (Nydam and Cifelli 2002a). A jaw fragment preserving two teeth with anguid-type tooth attachment was reported from the Mussentuchit Member of the Cedar Mountain Formation (early Cenomanian) of Utah (Nydam 2002). Anguimorphan osteoderms are known from the Dakota Formation (mid-Cenomanian) of Utah (Nydam 2013), but may belong to either Cnodontosaurus suchockii or the indeterminate platynotan. Specimens referable to the genus Colpodontosaurus—formally considered an enigmatic “necrosaurid” (Estes 1983a), but here considered an indeterminate anguimorphanFootnote 3—have been recovered from the John Henry Member of the Straight Cliffs Formation (Santonian) of Utah (Nydam 2013). Specimens referred to Colpodontosaurus cracens have been reported from throughout the Oldman Formation (early–mid-Campanian) of Alberta (Peng et al. 2001) as well as the Maastrichtian Lance (Estes 1964), Hell Creek (Bryant 1989; Estes et al. 1969), and Scollard (Gao and Fox 1996) formations. An anguimorphan osteoderm not comparable to the “Odaxosaurus”-grade common in other Upper Cretaceous localities has been recovered from the Wahweap Formation (early–mid-Campanian) of Utah (Nydam 2013).

Temporal and geographic distribution of indeterminate anguimorphan specimens
Serpentes (Fig. 15)
The presence of snakes in the Mesozoic of North America is currently restricted to the Upper Cretaceous rocks. Unlike the fossil record of lizards, the snakes are almost exclusively known from vertebrae with only a few referred cranial specimens known. Nearly all of these specimens have been referred to Coniophis with all pre-Maastrichtian species referrals viewed as tentative (see Gardner and Cifelli 1999 for a brief review of the taxonomic problems). The earliest evidence of a snake in North America is vertebrae referred to Coniophis sp. from the Cedar Mountain Formation (early Cenomanian) of central Utah (Gardner and Cifelli 1999). Additional snake vertebrae (also tentatively referred to Coniophis sp.) are described by Nydam (2013) from southern Utah’s Dakota Formation (middle Cenomanian), Smoky Hollow (Turonian) and John Henry (Santonian) members of the Straight Cliffs Formation, Wahweap Formation (lower Campanian), and the Kaiparowits Formation (upper Campanian). Additional upper Santonian records of snakes include Coniophis cf. precedens and an unnamed genus and species “Aniliidae” from the Milk River Formation of southern Alberta (Fox 1975). Additional upper Campanian records include Coniphis sp. from the Mesaverde Formation of Wyoming (DeMar and Breithaupt 2006), C. cosgriffi (generic referral questioned by Longrich et al. 2012b: supplemental information) from the Fruitland Formation of New Mexico (Armstrong-Ziegler 1978), an indeterminate snake (jaw fragments) from the Aguja Formation of southern Texas (Nydam et al. 2013), and several vertebrae (some in articulation) of Coniophis sp. from the Cerro del Pueblo Formation of Coahuila (Martinez 2010). The specimens from the Aguja Formation include a dentary similar in size and depth to those described for Dinilysia and Najash (Zaher et al. 2009; Zaher and Scanferla 2012), but the specimen is too incomplete for more conclusive comparisons (Nydam et al. 2013).
Maastrichtian snakes show a modest increase in diversity. Vertebrae of Coniophis precedens have been recovered from the Lance Formation of Wyoming (Estes 1964; Marsh 1892). Additional vertebrae of Coniophis precedens and a partial vertebra referred to Boidae indet. (Bryant 1989; Estes et al. 1969) have been recovered from the Hell Creek Formation of Montana. Bryant (1989: 49) further noted that the boid vertebra showed indications of an arthritic-like pathology. Longrich et al. (2012a) erected Cerberophis robustus based on a boid vertebra from the Hell Creek Formation. Longrich et al. (2012b) also referred several isolated jaw specimens from the Hell Creek Formation to Coniophis precedens and rediagnosed the taxon. As with the cranial and osteodermal elements referred to Odaxosaurus piger (above) and cranial elements referred to Chamops segnis (see arguments in Nydam et al. 2010), the referral of these jaws to Coniophis precedens is considered tentative pending the recovery of jaws in association/articulation with vertebrae comparable to the holotype specimen.
Assuming all of the generic referrals are correct, Coniophis survived the terminal Cretaceous extinction event in North America (Longrich et al. 2012a) and is present in the Paleocene. Estes (1976) reported several vertebrae from the Tongue River Formation of Montana that he assigns to Coniophis sp. (cited as Coniophis precedens by Longrich et al. 2012b). Longrich et al. (2012b: supplementary information) also suggest that the vertebrae from the Tongue River Formation referred by Estes (1976) to Dunnophis should be transferred to Coniophis. Vertebrae from Wyoming (McGrew 1959) and North Dakota (Smith 2013) suggest the possibility that Coniophis survived into the Eocene of North America, but Smith (2013: footnote on 158) noted that the work of Longrich et al. (2012b) calls into question the referral of the Eocene specimens to Coniophis. Smith (2013) provided a detailed review of the fossil record and publication history of Coniophis and I refer the reader there for additional information.
The distribution of snakes during the Late Cretaceous is interesting in that the range during the late Santonian–early Campanian extends from southern Alberta (Milk River Formation) south to southern Utah (John Henry Member of Straight Cliffs Formation, Wahweap Formation). However, during the late Campanian, snakes appear to be absent from the well-sampled vertebrate faunas of southern Alberta and Montana with the most northerly occurrence in the Mesaverde Formation of Wyoming. By the Maastricthian, snakes are again as far north as Montana (Hell Creek Formation). It is possible that, even with the substantial collection made in southern Alberta and Montana, this is still a sampling bias, but a similar pattern is seen in some fish taxa (D. Brinkman, personal communication).

Temporal and geographic distribution of Serpentes. The asterisk indicates the only record (Aguja Formation) that does not yet include vertebrae
Amphisbaenia (Fig. 16)
The earliest known amphisbaenians in North America are from the middle–late Paleocene of Montana (Estes 1976), Wyoming (Estes 1965, 1975; Gilmore 1942b), and New Mexico (Sullivan 1985). Amphisbaenians are currently lacking a presence in the Mesozoic of North America. See Estes (1983a), Kearny (2003b), Kearny and Stuart (2004), Hembree (2007), and Müller et al. (2011), and references therein, for a review of fossil forms and recent work on the systematic status of the group.

Temporal and geographic distribution of Amphisbaenia
Discussion
Summary of temporal distributions
As of this writing, the fossil record of squamates from the Jurassic and Cretaceous of North America (at least in the Western Interior) appears to support the earlier observations of Estes (1983a), Evans (1998, 2003), and Nydam (2002) of an early fauna (Late Jurassic–Early Cretaceous) derived from a common Laurasian stock that gives way in the Late Cretaceous to a fauna distinctly North American in its composition and diversity. The exception to this scenario may be Xantusiidae, which, if Tepexisaurus is a stem xantusiid, arose in the Early Cretaceous and continued on into the Cenozoic. For the Late Cretaceous, from early Cenomanian through the Maastrichtian, this pattern is well supported by the early appearance and apparently continuous presence of Chamopsiidae, Polyglyphanodontini, forms similar or pertaining to Xenosauridae, Platynota, and Serpentes; all groups currently unknown from the Early Cretaceous. Contogeniids first appear in the middle Cenomanian and persist through the Paleocene and anguids—particularly the ubiquitous Odaxosaurus—appear in the Turonian and are consistent faunal components through the end of the Cretaceous. Xantusiidae (minus Tepexisaurus) and Iguanidae are late-comers to the North American fauna with earliest tentative records from the late Campanian. There are surely several scincoidean groups that also fit one or the other of these patterns pending better understanding of their interrelationships, a task confounded by lack of sufficiently distinguishing characteristics of the partial jaws and unassociated osteoderms that form the bulk of these specimens. As such, it is only possible to conclude that scincoideans, particularly those with paramacellodid–cordylid grade dentitions, extend from the Early Cretaceous (or the Late Jurassic if they are indeed related to paramacellodids) through the Late Cretaceous.
At the generic level, it appears that Contogeniidae (i.e. Contogenys), Serpentes (i.e. Coniophis) and Xenosauridae (i.e. Exostinus) continued into the Paleocene (Longrich et al. 2012a). Longrich et al. (2012a) also include the anguids ‘Gerrhonotus’ and Odaxosaurus among the taxa that survive the K/T. However, specimens referred to ‘Gerrhonotus’ were actually identified as cf. Gerrhonotus (Estes 1983a) and may not be the same taxon. Additionally, all post-Mesozoic references to Odaxosaurus (or its taxonomic equivalents) have been convincingly transferred to other taxa or deemed unsupportable (sensu Gao and Fox 1996). Even so, at the family level, the taxa Anguidae, Platynota, and possibly Iguanidae appear to have also survived into the Paleocene although represented by taxa unknown in the Cretaceous.
Based on the known fossil record, the pre-Cenozoic squamate fauna of North America reached its greatest diversity during the late Campanian (approximately equivalent to the Judithian NALMA). The same pattern applies to the North American herbivorous dinosaur (Lehman 1997, 2001) and mammal (Kielan-Jaworoswka et al. 2004) faunas. An obvious bias to this peak diversity is the greater number and wider distribution of fossiliferous localities from the late Campanian (Fig. 17). The result is the sampling of more, with at least some locally endemic, taxa. Unlike the pattern seen in the herbivorous dinosaurs (Lehman 2001), the squamate faunal composition did not undergo any dramatic changes once it was established at the beginning of the Late Cretaceous.

Comparison of the taxonomic richness of the North American squamate fauna with the number of squamate-producing localities. Upper graph the current estimate of taxa/morphotypes for major taxonomic divisions of Squamata for each major time interval sampled. Lower graph the number of localities for each major time interval sampled. Note the tight correlation for all ages except Late Jurassic and Turonian
Sampling and diversity
The diversity of squamates during the Mesozoic in North America peaks twice prior to the Cretaceous-Cenozoic extinction event. The first peak is during the Cenomanian and the second during the late Campanian. As shown in Fig. 17, these peaks are coincident with the relatively high number of localities sampled at these time periods. The Cenomanian peak is representative of a fauna restricted to Utah while the late Campanian peak is associated with diversity throughout North America. Likewise, the apparent dip in squamate diversity during the Coniacian–Santonian is coincident with a low number of squamate-producing localities and does not necessarily represent a drop in diversity of the North American fauna. The tight correlation between numbers of localities and diversity of squamates suggests that, if the number of localities were more equitable, then the diversity of squamates would also be more consistent.
Exceptions to this correlation of number of localities to diversity of squamates occur during the Late Jurassic and the Turonian. For the Late Jurassic, there are several localities, but relatively few taxa suggesting a lower diversity of squamates from which to sample. This appears to be a widespread phenomenon considering the relatively low diversity of similar taxa throughout the Late Jurassic–Early Cretaceous of Laurasia (Evans 1993, 1995, 1998, 2003). During the Turonian, there are very few localities (all in Utah), but the diversity of squamates is relatively high. This suggests that the fauna, at least locally, was very diverse at this time interval. The taxa in this fauna include those characteristic of both earlier (Cenomanian) and later (Campanian–Maastrichtian) faunas indicating that some of the major taxonomic themes had already been established.
Development of the North American mesozoic squamate fauna
The current evidence indicates that nearly all of the major groups of squamates of the Late Cretaceous were established by the middle Cenomanian (Fig. 18). The exceptions are xantusiids (again assuming inclusion of Tepexisaurus) which appear in the Albian (approximately late Campanian if Tepexisaurus is not a stem xantusiid), xenosaurids and anguids which first appear in the Turonian, and, iguanids which first appear in the mid-Campanian. The Turonian appearance of xenosaurids and anguids (specifically Odaxosaurus) could be either a sampling bias that failed to recover them from the Cenomanian, or it is possible that these taxa evolved in situ in North America from the established anguimorphans currently of indeterminate taxonomy. Considering the close morphological similarities between contogeniids and xantusiids, it is likely that the latter evolved in North America from the established contogeniid stock (Nydam et al. 2013). The scenario for iguanians is more complicated. Prior to the Late Santonian, there is no evidence of iguanians in North America. The earliest substantive records of iguanians (mostly acrodontans) are from the Jurassic of Gondwana (Evans et al. 2002)—a report of a Triassic acrodontan from India (Datta and Ray 2006) is likely a Cenozoic record (Hutchison et al. 2012)—with additional records of purported iguanians from the Aptian–Albian of Mongolia (Gao and Nessov 1998). Conrad and Norell (2007) claim that no iguanians are present in Asia prior to the Late Cretaceous, but that, by the Campanian, there was likely a substantial diversity of iguanians in Mongolia (Conrad and Norell, 2007). A likely scenario is that iguanians were present in Asia earlier in the Late Cretaceous and entered North America during the Santonian, but appear to be the only squamate group to enter North America at this time. It is likely that the land bridge between Asia and North America was still present by the Santonian (Nockleberg et al. 2000). An earlier (pre-Cenomanian) arrival of iguanids may have occurred during the major influx of squamate taxa (discussed below) and has gone unrecorded in the fossil record, possibly due to a lack of pre-Campanian localities in the northern latitudes. The presence of iguanians in North America during the Campanian and Maastrichtian are likely a continuation of taxa from the Santonian. A limited exchange of dinosaur taxa between Asia and North America during the mid-Campanian appears to be isolated to these large-bodied taxa (Sereno 1999 and references therein). The pre-Late Cretaceous exchange of taxa between Asia and North America allowed for a freer exchange of taxa.

Distribution of squamate taxa in North America during the Late Jurassic–Paleocene. Each black box represents an occurrence of a species or form representing a major division of squamates. Boxes or bars with gray margins indicate stratigraphic uncertainty and placement is a best estimate based on published accounts
Nydam (2002) hypothesised that the apparently simultaneous appearance at the base of the Late Cretaceous of many squamate lineages characteristic of the North American squamate fauna up until the terminal Cretaceous extinction was the result of the opening of a passage from Asia to North America at the end of the Early Cretaceous, and possibly again in the Turonian and Santonian–early Campanian. Immigration of squamates from Asia to North America during the Early Cretaceous remains the most likely scenario for the relatively sudden change in the North American squamate fauna, though current evidence does not support exchanges, except iguanians, during the Turonian and Santonian–early Campanian. Taxonomic support for an Asian–North American exchange includes the shared presence of “polyglyphanodontids” (sensu Estes 1983a; “macrocephalosaurids” of Alifanov 2000 now referred to as gilmoreteiids; Langer 1998) and chamopsiids (“mongolochamopines” of Alifanov 2000; “teiids” of Gao and Norell 2000) as well as varanid-like and heldoermatid-like platynotans (Alifanov 2000; Borsuk-Białynicka 1984; Gao and Norell 2000). Taxonomic groups that were not exchanged include acrodontans, iguanids (at least not until later), and gekkotans known from the Late Cretaceous of Asia (all sensu taxonomic review by Gao and Norell 2000) and contogeniids and possibly paramacellodid–cordylid grade scincomorphans of North America (Nydam 2013; Nydam and Fitzpatrick 2009). Among the shared higher taxonomic groups, there are currently no taxa at the generic or species level present in both the Asian and North American faunas. In contrast, even though the fossil record for squamates from the Late Jurassic of North America is limited, it shares Paramacellodus, Dorsetisaurus, and Parviraptor with faunas known from the Late Jurassic–Early Cretaceous in Europe (Estes 1983b; Evans 1995, 2003) and Paramacellodus and Dorsetisaurus from the Early Cretaceous of Mongolia (Alifanov 2000). Additionally, the presence of the polyglyphanodontine Bicuspidon in the Late Cretaceous of Europe (Folie and Codrea 2005; Makádi 2006) further complicates interpretations of biogeographical patterns.
The lack of shared genera and/or species between Asia and North America during the Cretaceous is possibly due to a couple of compounding problems. The first of these is the limited amount of information regarding the Early Cretaceous squamate faunas of Asia: with the exception of two taxa based on skeletons (Evans and Wang 2007; Evans and Wang 2010), the pre-Late Cretaceous Asian lizard fauna has yet to be systematically described and illustrated. Second is the limited information of the squamate fauna from pre-Cenomanian North America. There are few Upper Jurassic–Lower Cretaceous localities from which squamates have been recovered, and most of the Lower Cretaceous remains unknown. If shared taxa are to be recovered from North America, it would mostly likely be from the Lower Cretaceous of northern latitudes, a region currently devoid of such localities.
Another consideration in regards to shared taxa between Asia and North America is the nature of the continental connection. Prior to the end of the Jurassic, Laurasia was a large, continuous landmass covering a broad range of latitudes. The connection between Asia and North America at the end of the Early Cretaceous (before 108 Ma; sensu Zanno and Makovicky 2011) was likely a rapidly developed, narrow land bridge at a relatively high (between 60° and 80° N) latitude and persisted well into the Late Cretaceous assuming an increasingly polar (> 80° N latitude) position by the Santonian (Nockleberg et al. 2000). This high latitude in association with a very active, rapidly changing tectonic environment (Nockleberg et al. 2000) almost certainly restricted the utility of the bridge for small-bodied, ectothermic tetrapods during the post-Cenomanian Cretaceous.
Another possible region of influence on the North American squamate fauna during the Cretaceous is South America. At least by the Maastrichtian, the herbivorous dinosaur fauna in southern North America was dramatically changed by the reappearance of sauropod dinosaurs from South America (Lehman 2001), but South America does not appear to have had an influence on the squamate fauna. While still not well understood, the Mesozoic squamates of South America appear to be distinct in diversity and composition. In a reverse of the North American pattern, the South American fauna appears to have been dominated by snakes and possessed few lizards (Albino 1996; Benson et al. 2013). One group long thought to represent a connection between the two continents was the “teiids” from the North American fauna (Estes 1983a, b). These taxa are now recognised as members of Borioteiioidea (chamopsiids and polyglyphanodontines) which are either distantly related to Teiioidea (sensu Nydam et al. 2007) or stem scleroglossans (sensu Gauthier et al. 2012). Estes (1983b), Nydam et al. (2007), and Gauthier et al. (2012) all recognised the close relationship of the polyglyphanodontines to Asian counterparts further weakening the hypothesis of a North American–South American exchange during the Cretaceous. The similarity in size and dimensions of the snake jaw from the Aguja Formation of southern Texas (Nydam et al. 2013) with that of the jaws of large madtsoiid snakes (e.g. Dinilsyia and Najash) from the Late Cretaceous of South America could indicate a South American influence in North America during the late Campanian. Additional evidence of the possible influence of South American madtsoiids in North America is in a published abstract (Head and Holroyd 2008) in which the snake Helgras prisciformis (Paleocene, New Mexico) is transferred to the Madtsoiidae. The possible presence of snakes from South America in North America is in contrast with an apparent lack of exchange of lizards between the two continents but is comparable to the limited exchange of other vertebrates, possibly via an archipelago (Estes and Baéz 1985). Our understanding of the diversity of the South American squamate fauna during the Mesozoic is growing rapidly (e.g. Albino 2000; Apesteguía and Zaher 2006; Candeiro 2007; Candeiro et al. 2008) and the discovery of additional evidence for exchange between North and South America is possible.
Conclusions
The record of squamates from the later Mesozoic of North America continues to grow and gaps continue to be filled, but it is clear that more specimens are needed to better understand the changes in the fauna. While all horizons should continue to be investigated, two in particular are imperative. The first is the Early Cretaceous prior to, but also including, the Aptian–Albian. One of the reasons there appears to be a rapid radiation of new taxa at the beginning of the is Late Cretaceous is due to the limited understanding of the lizard fauna prior to this time. Understanding the squamate fauna from the Early Cretaceous will better establish if any of the taxonomic groups that currently seem to appear in the Late Cretaceous might actually have evolved in North America preceding the opening of the dispersal route from Asia. Well-exposed fossiliferous rocks from the Early Cretaceous of North America are rare, but the recovery of mammals and/or sphenodontians from the Arundel Clay of Maryland (Cifelli et al. 1999a; Rose et al. 2001), the Kelvin Formation of Utah (Prothero 1983), the Kootenai Formation of Montana (Gaylord et al. 1981), and the Yellow Cat Member of the Cedar Mountain Formation (Kirkland et al. 1997) suggest that there is a potential for the recovery of squamates in all these units. The Lakota Formation of South Dakota and the Bissett Formation of southern Texas are other potential Lower Cretaceous rock units currently under investigation by Dr. R.L. Cifelli of the Oklahoma Museum of Natural History. The second horizon in great need of further investigation is the Paleocene of North America. The squamate fauna from the Paleocene represents a transitional fauna between the diverse but very distinct faunas of the Late Cretaceous and Eocene. This horizon is better understood than the Early Cretaceous, but still needs additional attention. Further collection from such areas as the Polecat Bench region of Montana and the North Horn Mountain and Dragon Valley of Utah, as well as rock units from which lizards are only now being found such as the Goler Formation of California (Nydam and Lofgren 2008, work in progress), are needed to better understand which taxa survived the terminal Cretaceous extinction and which taxa may have contributed to the new diversity in squamates of the Eocene.
Notes
The rich and diverse record of marine squamates will not be considered here, and all mention of “squamates” is intended to imply terrestrial lizards, snakes, and amphisbaenians.
I agree with Conrad et al. (2010) that the holotype specimen alone is insufficient for phylogeneticevaluation, but I disagree that it is insufficient for species-level diagnosis because it is easily distinguishable from other squamate dentaries.
References
Albino AM (1996) The South American fossil Squamata (Reptilia: Lepidosauria). Münch Geowiss Abh (A)(30):185–202
Albino AM (2000) New record of snakes from the Cretaceous of Patagonia. Geodiversitas 22(2):247–253
Alifanov VR (2000) The fossil record of Cretaceous lizards from Mongolia. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (eds) The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, pp 368–389
Apesteguía S, Zaher H (2006) A Cretaceous terrestrial snake with robust hindlimbs and a sacrum. Nature 440:1037–1040
Armstrong-Ziegler JG (1978) An aniliid snake and associated vertebrates from the Campanian of New Mexico. J Paleont 52(2):480–483
Armstrong-Ziegler JG (1980) Amphibia and Reptilia from the Campanian of New Mexico. Fieldiana Geo (New Series, 4):1–39
Barr BN (2012) Systematics of platynotan lizards. PhD thesis, University of Alberta, Edmonton
Benson RBJ, Mannion PD, Butler RJ, Upchurch P, Goswami A, Evans SE (2013) Cretaceous tetrapod fossil record sampling and faunal turnover: Implications for biogeography and the rise of modern clades. Palaeogeogr Palaeoclimatol Palaeoecol 372:88–107
Bhullar BAS (2010) Cranial osteology of Exostinus serratus (Squamata: Anguimorpha), fossil sister taxon to the enigmatic clade Xenosaurus. Zool J Linn Soc 159:921–953
Bhullar BAS (2011) The power and utility of morphological characters in systematics: A fully resolved phylogeny of Xenosaurus and its fossil relatives (Squamata: Anguimorpha). Bull Mus Comp Zoo 160(3):65–181
Borsuk-Białynicka M (1984) Anguimorphan and related lizards from the Late Cretaceous of the Gobi Desert, Mongolia. Palaeont Polon 1984(46):5–106
Brinkman DL, Cifelli RL, Czaplewski NJ (1998) First occurence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian-Albian) of Oklahoma. Okla Geo Notes 146:1–27
Broschinski A (2000) The lizards from the Guimarota mine. In: Martin T, Krebs B (eds) Guimarota, A Jurassic Ecosystem. Verlag Dr. Friedrich Pfeil, München, pp 59–68
Broschinski A, Sigogneau-Russell D (1996) Remarkable lizard remains from the Lower Cretaceous of Anoual (Morocco). Annal Paléont 82(3):147–175
Bryant LJ (1989) Non-dinosaurian lower vertebrates across the Cretaceous-Tertiary boundary in Northeastern Montana. Geol Sci 134:107
Burge DL (1996) New dinosaur discoveries in the Lower Cretaceous of southesatern Utah. Fossils of Arizona 4:85–105
Caldwell MW (2007) Ontogeny, anatomy and attachment of the dentition in mosasaurs (Mosasauridae: Squamata). Zool J Linn Soc 149(4):687–700
Caldwell MW, Lee MSY (1997) A snake with legs from the marine Cretaceous of the Middle East. Nature 386:705–709
Callison G (1987) Fruita: a place for wee fossils. In: Averett WR (ed) Paleontology and geology of the dinosaur triangle: Guidebook for 1987 field trip. Museum of Western Colorado, Grand Junction, pp. 91–96
Candeiro CRA (2007) Paleogeographic distribution of the terrestrial squamate reptiles from the Cretaceous of Brazil. Biosci J 23(1):65–74
Candeiro CRA, Santos AR, Bergqvist LP, Ribeiro LCB, Apestiguía S (2008) The Late Cretaceous fauna and flora of the Uberaba area (Minas Gerais State, Brazil). J So Am Earth Sci 25:203–216
Carpenter K (1979) Vertebrate fauna of the Laramie Formation (Maestrichtian), Weld County, Colorado. Rocky Mount Geol 17(1):37–48
Carrano MT, Velez-Juarbe J (2006) Paleoecology of the Quarry 9 vertebrate assemblage from Como Bluff, Wyoming (Morrison Formation, Late Jurassic). Palaeogeogr Palaeoclimatol Palaeoecol 237(2–4):147–159
Chure DJ, Carpenter K, Liwin R, Hasiotis S, Evanoff E (1998) Appendix. The fauna and flora of the Morrison Formation. Mod Geol 23:507–537
Cifelli RL (1990a) Cretaceous mammals of southern Utah. III. Therian Mammals from the Turonian (Early Late Cretaceous). J Vertebr Paleontol 10(3):332–345
Cifelli RL (1990b) Cretaceous mammals of southern Utah. IV. Eutherian Mammals from the Wahweap (Aquilan) and Kaiparowits (Judithian) Formations. J Vertebr Paleontol 10(3):346–360
Cifelli RL (1993) Early Cretaceous mammal from North America and the evolution of marsupial dental characters. Proc Nat Acad Sci USA 90:9413–9416
Cifelli RL (1996) Techniques for recovery and preparation of microvertebrate fossils. Okla Geol Surv Spec Publ 96–4:1–36
Cifelli RL (1999) Therian teeth of unusual design from the mid-Cretaceous (Albian-Cenomanian) Cedar Mountain Formation of Utah. J Mamm Evol 6(3):247–270
Cifelli RL (2004) Marsupial mammals from the Albian-Cenomanian (Early-Late Cretaceous) boundary, Utah. Bull Am Mus Nat Hist 285:62–79
Cifelli RL, de Muizon C (1997) Dentition and jaw of Kokopellia juddi, a primitive marsupial or near-marsupial from the medial Cretaceous of Utah. J Mamm Evol 4(4):241–258
Cifelli RL, Madsen SK (1986) An Upper Cretaceous symmetrodont (Mammalia) from southern Utah. J Vertebr Paleontol 6(3):258–263
Cifelli RL, Madsen SK (1998) Triconodont mammals from the medial Cretaceous of Utah. J Vertebr Paleontol 18(2):403–411
Cifelli RL, Madsen SK (1999) Spalacotheriid symmetrodonts (Mammalia) from the medial Cretaceous (upper Albian or lower Cenomanian) Mussentuchit local fauna, Cedar Mountain Formation, Utah, USA. Geodiversitas 21(2):167–214
Cifelli RL, Nydam RL (1995) Primitive, helodermatid-like platynotan from the Early Cretaceous of Utah. Herpetologica 51(3):286–291
Cifelli RL, Kirkland JI, Weil A, Deino AR, Kowallis BJ (1997) High-precision 40Ar/39Ar geochronology and the advent of North America's Late Cretaceous terrestrial fauna. Proc Natl Acad Sci USA 94:11163–11167
Cifelli RL, Lipka TR, Schaff CR, Rowe TB (1999a) First Early Cretaceous mammal from the eastern seaboard of the United States. J Vertebr Paleontol 19(2):199–203
Cifelli RL, Nydam RL, Eaton JG, Gardner JD and Kirkland JI (1999b) Vertebrate faunas of the North Horn Formation (Upper Cretaceous-Lower Paleocene), Emery and Sanpete counties, Utah. In: Gillette DD (ed.), Vertebrate Paleontology in Utah. Utah Geol Surv Misc Publ 99–1:pp. 377–388
Cifelli RL, Nydam RL, Gardner JD, Weil A, Eaton JG, Kirkland JI and Madsen SK (1999c) Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County: the Mussentuchit local fauna In: Gillette DD (ed.), Vertebrate Paleontology in Utah. Utah Geol Surv Misc Publ 99–1:219–242
Condon SM (2000) Stratigraphic framework of Lower and Upper Cretaceous rocks in central and eastern Montana. United States Geological Survey Digital Data Series DDS-57, Version 1.0:1–12
Conrad JL (2008) Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bull Am Mus Nat Hist 310(310):1–182
Conrad JL, Norell MA (2007) A complete Late Cretaceous iguanian (Squamata, Reptilia) from the Gobi and identification of a new iguanian clade. Am Mus Nov 3584:1–47
Conrad JL, Ast JC, Montanari S, Norell MA (2010) A combined evidence phylogenetic analysis of Anguimorpha (Reptilia: Squamata). Cladistics 27(3):230–277
Crane C (2011) Vertebrate paleontology and taphonomy of the Late Cretaceous (Campanian) Bladen Formation, Bladen County, North Carolina. Masters thesis, East Carolina University
Crother BI, Miyamoto MM, Presch W (1986) Phylogeny and Biogeography of the lizard family Xantusiidae. Syst Zool 35(1):37–45
Currie BS (1998) Upper Jurassic-Lower Cretaceous Morrison and Cedar Mountain formations, NE Utah-NW Colorado: Relationships between nonmarine deposition and early cordilleran foreland-basin development. J Sediment Res 68(4):632–652
Datta PM, Ray S (2006) Earliest lizard from the late triassic (Carnian) of India. J Vertebr Paleontol 26(4):795–800
DeMar DGJ and Breithaupt BH (2006) The nonmammalian vertebrate microfossil assemblages of the Mesaverde Formation (Upper Cretaceous, Campanian) of the Wind River and Bighorn Basins, Wyoming. In: Lucas SG and Sullivan RM (eds.), Late Cretaceous vertebrates from the Western Interior. N Mex Mus Nat Hist Sci Bull 35:33–53
DeMar DGJ, Varricchio DD, Head JJ, Moore JR, Wilson GP (2012) A nearly complete fossil iguanian from the Upper Cretaceous (Campanian) Two Medicine Formation of western Montana. J Vertebr Paleontol 32(Suppl 1):86
Denton RK, O'Neill RC (1995) Prototeius stageri, gen. et sp. nov., a new teiid lizard from the Upper Cretaceous Marshalltown Formation of New Jersey, with a preliminary phylogenetic revision of the Teiidae. J Vertebr Paleontol 15(2):235–253
Denton RK, O'Neill RC, Grandstaff BS, Parris DC (1991) Earliest record of an advanced glyptosaurine lizard from the Late Cretaceous (Campanian) of New Jersey. J Vertebr Paleontol 11(3, supplement):25A
Dyman TS, Cobban WA, Davis LE and Eves RL (2002a) Upper Cretaceous marine and brackish water strata at Grand Staircase - Esclante National Monument, Utah. In: Lund WR (ed) Field guide to geologic excursions in southwestern Utah and adjacent areas of Arizona and Nevada. US Geological Survey, Washington DC, pp 171–198
Dyman TS, Cobban WA, Titus A, Obradovich JD (2002b) New biostratigraphic and radiometric ages for Albian - Turonian Dakota Formation and tropic shale at Grand Staircase - Escalante National Monument and Iron Springs Formation near Cedar City, Parowan, and Gunlock in SW Utah. Geol Soc Am Abs Prog 34(4):13
Eaton JG (1991) Biostratigraphic framework for the Upper Cretaceous rocks of the Kaiparowits Plateau, southern Utah. In: Nations JD and Eaton JG (eds.), Stratigraphy, depositional environments, and sedimentary tectonics of the western margin, Cretaceous Western Interior Seaway, Geol Soc Am Spec Pap 260:47–63
Eaton JG (1995) Cenomanian and Turonian (early Late Cretaceous) multituberculate mammals from southwestern Utah. J Vertebr Paleontol 15(4):761–784
Eaton JG (1999) Vertebrate paleontology of the Iron Springs Formation, Upper Cretaceous, southwestern Utah. In: Gillette DD (ed.), Vertebrate Paleontology in Utah. Utah Geol Surv Misc Publ 99–1:339–343
Eaton JG (2002) Multituberculate mammals from the Wahweap (Campanian, Aquilan) and Kaiparowits (Campanian, Judithian) formations, within and near Grand Staircase-Escalante National Monument. Utah Geol Surv Misc Publ 02–04
Eaton JG (2006) Santonian (Late Cretaceous) Mammals from the John Henry Member of the Straight Cliffs Formation, Grand Staircase-Escalante National Monument, Utah. J Vertebr Paleontol 26(2):446–460
Eaton JG, Cifelli RL (1988) Preliminary report on Late Cretaceous mammals of the Kaiparowits Plateau, southern Utah. Contrib Geol 26(2):45–55
Eaton JG, Cifelli RL (2001) Multituberculate mammals from the Early-Late Cretaceous boundary, Cedar Mountain Formation, Utah. Acta Palaeontol Pol 46(4):453–518
Eaton JG, Cifelli RL, Hutchison JH, Kirkland JI and Parrish JM (1999a) Cretaceous vertebrate faunas from the Kaiparowits Plateau, south-central Utah. In: Gillette DD (ed) Vertebrate Paleontology in Utah. Utah Geol Surv Misc Publ 99–1:345–353
Eaton JG, Diem S, Archibald JD, Schierup C and Munk H (1999b) Vertebrate paleontology of the Upper Cretaceous rocks of the Markagunt Plateau, southwestern Utah. In: Gillette DD (ed) Vertebrate Paleontology in Utah. Utah Geol Surv Misc Publ 99–1:323–333
Eberth DA (2002) Review and comparison of Belly River Group and Edmonton Group stratigraphy and stratigraphic architecture in the southern Alberta plains. CSPG Diamon Jubilee Convention, Program and Abstracts 117. Abstracts of Technical Talks, Posters & Core Displays including Extended Abstracts. PDF file 227S0125. Compact Disc
Eberth DA (2005) The Geology. In: Currie PJ, Koppelhus EB (eds) Dinosaur Provincial Park: A spectacular ancient ecosystem revealed. Indiana University Press, Bloomington, pp 54–82
Eberth DA and Hamblin AP (1993) Tectonic, stratigraphic, and sedmentologic significance of a regional discontinuity in the upper Judith River Group (Belly River wedge) of southern Alberta, Saskatchewan, and northern Montana. Can J Earth Sci(30):174–200
Eberth DA, Jesús CRD-d, Lerbekmo JF, Brinkman DB, Rosa RAR-d, Sampson SD (2004) Cerro del Pueblo Fm (Difunta Group, Upper Cretaceous), Parras Basin, southern Cohuila, Mexico: reference sections, age, and correlation. Revist Mex Ciencia Geol 21(3):335–352
Estes R (1964) Fossil vertebrates from the Late Cretaceous Lance Formation, eastern Wyoming. Univ Calif Publ Geol Sci 49:1–169
Estes R (1965) Notes on some Paleocene lizards. Copeia 1965(1):104–106
Estes R (1969a) Relationships of two lizards (Sauria, Teiidae). Breviora 317:1–8
Estes R (1969b) A scincoid lizard from the Cretaceous and Paleocene of Montana. Breviora 1969(331):1–9
Estes R (1975) Lower vertebrates from the Fort Union Formation, Late Paleocene, Big Horn Basin, Wyoming. Herpetologica 31(4):365–385
Estes R (1976) Middle Paleocene lower vertebrates from the Tongue River Formation, southeastern Montana. J Paleont 50(3):500–520
Estes R (1983a) Sauria Terrestria, Amphisbaenia. Handbuch der Paläoherpetologie, Teil 10A. Gustav Fischer Verlag, Stuttgart
Estes R (1983b) The fossil record and early distribution of lizards. In: Rodin AGJ, Kiyata K (eds) Advances in herpetology and evolutionary biology: Essays in honor of Ernest E. Williams, Museum of Comparative Zoology. Harvard University, Cambridge, pp 365–398
Estes R, Baéz A (1985) Herpetofaunas of North and South America during the Late Cretaceous and Cenozoic: Evidence for interchange? In: Stehli FG, Webb SD (eds) The Great American Biotic Interchange. Plenum Press, New York, pp 139–197
Estes R, Berberian P, Meszoely CAM (1969) Lower Vertebrates from the Late Cretaceous Hell Creek Formation, McCone County, Montana. Brevoria 337:1–33
Estes R, de Quieroz K, Gauthier J (1988) Phylogenetic relationships within Squamata. In: Estes R, Pregill G (eds) Phylogenetic relationships of the lizard families: Essays commemorating Charles L. Camp. Stanford University Press, Stanford, pp 119–281
Evans SE (1993) Jurassic lizard assemblages. Rev Paléobiol Spéc 7:55–65
Evans SE (1995) Lizards: evolution, early radiation, and biogeography. In: Sun A-L, Wang Y (eds) Sixth symposium on mesozoic terrestrial ecosystems and biota, short papers. China Ocean Press, Beijing, pp 51–55
Evans SE (1996) Parviraptor (Squamata: Anguimorpha) and other lizards from the Morrison Formation at Fruita, Colorado. In: Morales M (ed.), The Continental Jurassic. Mus N Ariz Bull 60: 243–248
Evans SE (1998) Lepidosaurian faunas from the Early Cretaceous: a clade in transition. Bull N Mex Mus Nat Hist Sci 14:193–198
Evans SE (2003) At the feet of dinosaurs: the early history and radiation of lizards. Biol Rev 28:513–551
Evans SE, Chure DJ (1998a) Morrison lizards: structure, relationships and biogeography. Mod Geol 23:35–48
Evans SE, Chure DJ (1998b) Paramacellodid lizard skulls from the Jurassic Morrison Formation at Dinosaur National Monument, Utah. J Vertebr Paleontol 18(1):99–114
Evans SE and Chure DJ (1999) Upper Jurassic lizards from the Morrison Formation of Dinosaur National Monument, Utah. In: Gillette DD (ed.), Vertebrate Paleontology in Utah. Utah Geol Surv Misc Publ 99–1:151–159
Evans SE, Manabe M (1999) Early Cretaceous lizards from the Okurodani Formation of Japan. Geobios 32(6):889–899
Evans SE, Searle B (2002) Lepidosaurian reptiles from the Purbeck Limestone Group of Dorset, southern England. Spec Pap Palaeontol 68(68):145–159
Evans SE, Wang Y (2005) The Early Cretaceous lizard Dalinghosaurus from China. Acta Palaeontol Pol 50(4):725–742
Evans SE, Wang Y (2007) A juvenile lizard specimen with well-preserved skin impressions from the Upper Jurassic/Lower Cretaceous of Daohugou, Inner Mongolia, China. Naturwissenschaften 94(6):431–439
Evans SE, Wang Y (2010) A new lizard (Reptilia: Squamata) with exquisite preservation of soft tissue from the Lower Cretaceous of Inner Mongolia, China. J Syst Palaeontol 8(1):81–95
Evans SE, Prasad GVR, Manhas BK (2002) Fossil lizards from the Jurassic Kota Formation of India. J Vertebr Paleontol 22(2):299–312
Evans SE, Ruiz AL, Rey JE (2000) A lizard from the Early Cretaceous (Berriasian–Valangian) of Montsec, Catalonia, Spain. N Jb Geol Paläont, Abh 215(1):1–15
Evans SE, Wang Y, Li C (2005) The Early Cretaceous lizard genus Yabeinosaurus from China: Resolving an enigma. J Sys Palaeontol 3(4):319–335
Fassett JE, Hinds JS (1971) Geology and fuel resources of the Fruitland Formation and Kirtland shale of the San Juan Basin, New Mexico and Colorado. US Geol Surv Prof Pap 676:1–76
Fassett JE, Steiner MB (1997) Precise age of C33N-C32R magnetic-polarity reversal, San Juan Basin, New Mexico and Colorado. N Mex Geol Soc 48:239–247
Fastovsky DE (1987) Paleoenvironments of vertebrate-bearing strata during the Cretaceous-Paleogene transition, eastern Montana and Western North Dakota. Palaios 2:282–295
Folie A, Codrea V (2005) New lissamphibians and squamates from the Maastrichtian of Haţeg Basin, Romania. Acta Palaenotol Pol 50:57–71
Forster CA (1984) The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana. Mosasaur 2:151–163
Fox RC (1975) Fossil snakes from the Upper Milk River Formation (Upper Cretaceous), Alberta. Can J Earth Sci 12:1557–1563
Fraser NC, Wu X-C (1998) Sphenodontians from the Brushy Basin Member of the Morrison Formation in Dinosaur National Monument. Mod Geol 23:17–34
Gao K, Fox RC (1991) New teiid lizards from the Upper Cretaceous Oldman Formation (Judithian) of southwestern Alberta, Canada, with a review of the Cretaceous record of teiids. Ann Carnegie Mus 60(2):145–162
Gao K, Fox RC (1996) Taxonomy and evolution of Late Cretaceous lizards (Reptilia: Squamata) from western Canada. Bull Carnegie Mus Nat Hist 33:1–107
Gao K, Hou L (1995) Iguanians from the Upper Cretaceous Djadochta Formation, Gobi Desert, China. J Vertebr Paleontol 15:57–78
Gao K, Norell MA (2000) Taxonomic composition and systematics of Late Cretaceous lizard assemblages from Ukhaa Tolgod and adjacent localities, Mongolian Gobi Desert. Bull Am Mus Nat Hist 249:1–118
Gao K, Nessov LA (1998) Early Cretaceous squamates from the Kyzylkum Desert, Uzbekistan. N Jb Geol Paläont, Abh 207:289–309
Gardner JD (1994) Amphibians from the Lower Cretaceous (Albian) Cedar Mountain Formation, Emery County, Utah. Abstracts of the 54th Annual Meeting, Society of Vertebrate Paleontology. J Vertebr Paleontol 14 (Supplement to no. 3):26A
Gardner JD (1999) New albanerpetonid amphibians from the Albian to Coniacian of Utah, USA—bridging the gap. J Vertebr Paleontol 19(4):632–638
Gardner JD, Cifelli RL (1999) A primitive snake from the Cretaceous of Utah. Spec Pap Palaeontol 60:87–100
Garrison JR Jr, Brinkman D, Nichols DJ, Layer P, Burge D, Thayn D (2007) A multidisciplinary study of the Lower Cretaceous Cedar Mountain Formation, Mussentuchit Wash, Utah: a determination of the paleoenvironment and paleoecology of the Eolambia caroljonesa dinosaur quarry. Cretac Res 28(3):461–494
Gates TA, Sampson SD, Zanno LE, Roberts EM, Eaton JG, Nydam RL, Hutchison JH, Smith JA, Loewen MA, Getty MA (2010) Biogeography of terrestrial and freshwater vertebrates from the Late Cretaceous (Campanian) Western Interior of North America. Palaeogeogr Palaeoclimatol Palaeoecol 291:371–387
Gauthier JA (1982) Fossil xenosaurid and anguid lizards from the early Eocene Wasatch Formation, southeast Wyoming, and a revision of the Anguiodea. Cont Geol 21(1):7–54
Gauthier JA, Kearney M, Maisano JA, Rieppel O, Behlke ADB (2012) Assembling the squamate tree of life: Perspectives from the phenotype and the fossil record. Bull Peabody Mus Nat Hist 53(1):3–308
Gaylord ST, Hopson JA, Parks P (1981) A redescription of the Toxolophosaurus cloudi Olson, a Lower Cretaceous herbivorous sphenodontid reptile. J Paleont 55(3):586–597
Gilmore CW (1928) Fossil lizards of North America. Mem Natl Acad Sci 22:1–201
Gilmore CW (1938) Fossil snakes of North America. Geol Soc Am Spec Pap 9:1–96
Gilmore CW (1940) New fossil lizards from the Upper Cretaceous of Utah. Smithsonian Misc Coll 99(16):1–3
Gilmore CW (1942a) Osteology of Polyglyphanodon, an Upper Cretaceous lizard from Utah. Proc US Nat Mus 92(3148):229–265
Gilmore CW (1942b) Paleocene faunas of the Polecat Bench Formation, Park County, Wyoming, Part II. lizards. Proc Am Philos Soc 85(2):159–167
Gilmore CW (1943) Osteology of Upper Cretaceous lizards from Utah, with a description of a new species. Proc US Natl Mus 93(3163):209–214
Goodwin MB, Deino AL (1989) The first radiometric ages from the Judith River Formation (Upper Cretaceous), Hill County, Montana. Can J Earth Sci 26(7):1384–1391
Head J, Holroyd P (2008) Assembly and biogeography of North American Paleogene snake faunas based on an expanded fossil record. J Vertebr Paleontol 28(3, supplement):90A
Hembree DI (2007) Phylogenetic revision of the Rhineuridae (Reptilia: Squamata: Amphisbaenia) from the Eocene to Miocene of North America. Univ Kan Paleontol Contribn Ser 15:1–20
Hutchinson MN, Skinner A, Lee MSY (2012) Tikiguania and the antiquity of squamate reptiles (lizards and snakes). Biol Lett 8(4):665–669
Hunt AP, Lucas SG (1992) Stratigraphy, paleontology, and age of the Fruitland and Kirtland formations (Upper Cretaceous), San Juan Basin, New Mexico. New Mexico Geological Society 43:217–239
Jinnah ZA, Roberts EM, Deino AL, Larsen JS, Link PK, Fanning CM (2009) New 40Ar-39Ar and detrital zircon U-Pb ages for the Upper Cretaceous Wahweap and Kaiparowits formations on the Kaiparowits Plateau, Utah: implications for regional correlation, provenance, and biostratigraphy. Cretac Res 30(2):287–299
Kashiyama Y, Fastovsky DE, Rutherford S, King J, Montellano M (2004) Genesis of a locality of exceptional fossil preservation: paleoenvironments of Tepexi de Rodrı́guez (mid-Cretaceous, Puebla, Mexico). Cretac Res 25(1):153–177
Kearney M (2003a) The phylogenetic position of Sineoamphisbaena hexatabularis reexamined. J Vertebr Paleontol 23(2):394–403
Kearney M (2003b) Systematics of the Amphisbaenia (Lepidosauria:Squamata) based on morphological evidence from recent and fossil forms. Herpetol Monog 17:1–74
Kearney M, Stuart BL (2004) Repeated evolution of limblessness and digging heads in worm lizards revealed by DNA from old bones. Proc R Soc Lond B 2004(271):1677–1683
Kielan-Jaworoswka Z, Cifelli RL, Luo Z-X (2004) Mammals from the Age of Dinosaurs. Columbia University Press, New York
Kirkland JI, Britt B, Burge DL, Carpenter K, Cifelli R, Decourten F, Eaton J, Hasiotis S, Lawton T (1997) Lower to middle Cretaceous dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution and biogeography. BYU Geol Stud 42(II):69–103
Kirkland JI, Cifelli RL, Britt BB, Burge DL, DeCourten FL, Eaton JG, Parrish JM (1999) Distribution of vertebrate faunas in the Cedar Mountain Formation, east-central Utah.In: Gillette DD (ed.), Vertebrate Paleontology in Utah. Utah Geol Surv Misc Pub 99–1:201–217
Kirkland JI, Eaton JG, Brinkman DB (2013) Elasmobranchs from the Upper Cretaceous freshwater facies in southern Utah. In: Titus AL and Loewen MA (eds.) At the top of the grand staircase: The Late Cretaceous of Southern Utah. Indiana University Press, Bloomington, pp 153–194
Konishi T, Caldwell MW (2007) New specimens of Platecarpus planifrons (Cope, 1874) (Squamata: Mosasauridae) and a revised taxonomy of the genus. J Vertebr Paleontol 27(1):59–72
Kosma R (2004) The dentitions of recent and fossil scincomorphan lizards (Lacertilia, Squamata)—Systematics, functional morphology, palecology. PhD thesis, Universität Hannover, Hannover
Langer MC (1998) Gilmoreteiidae new family and Gilmoreteius new genus (Squamata, Scincomorpha): replacement names for Macrocephalosauridae Sulimski, 1975 and Macrocephalosaurus Gilmore, 1943. Comun Mus Cien Tech PUCRS Sér Zool 11:13–18
Lee MSY (2005) Squamate phylogeny, taxon sampling, and data congruence. Organ Divers Evol 5(1):25–45
Lehman TM (1997) Late Campanian dinosaur biogeography in the Western Interior of North America. Dinofest Internat Proc 1997:223–240
Lehman TM (2001) Late Cretaceous dinosaur provinciality. In: Tanke DH, Carpenter K (eds) Mesozoic vertebrate life. Indiana University Press, Bloomington, pp 310–328
Longrich NR, Bhullar BAS, Gauthier JA (2012a) Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary. Proc Natl Acad Sci USA 109(52):21396–21401
Longrich NR, Bhullar BAS, Gauthier JA (2012b) A transitional snake from the Late Cretaceous period of North America. Nature 488:205–208
Makádi L (2006) Bicuspidon aff. hatzegiensis (Squamata: Scincomorpha: Teiidae) from the Upper Cretaceous Csehbányda Formation (Hungary, Bakony Mts). Acta Geol Hung 49:373–385
Marsh OC (1892) Notice of new reptiles from the Laramie Formation. Am J Sci 43:449–453
Martinez MCA (2010) Fossil vertebrates form the Cerro del Pueblo Formation, Coahuila, Mexico, and the distribution of Late Campanian (Cretaceous) terrestrial vertebrate faunas. Masters thesis, Southern Methodist, Dallas
McCord RD (1998) A new genus and species of Cretaceous polyglyphanodontine lizard (Squamata, Teiidae) from the Kaiparowits Plateau, Utah. In: Tomida Y, Flynn LJ, Jacobs LL (eds.), Advances in Vertebrate Paleontology and Geochronology, National Science Museum, Tokyo, Nat Mus Monog 14:281–292
McCord RD, Gillette DD (2005) Cretaceous vertebrates of Arizona. Mesa Southwest Mus Bull 11:94–103
McCord RD, McCullough G, McCullough BJ, Hodnet JP, Scarborough R (2001) Ongoing research in the Late Cretaceous Fort Crittenden Formation, southeastern Arizona: the non-dinosaurian vertebrate fauna. Mesa Southwest Mus Bull 5:65–71
McGrew PO (1959) The Geology and Paleontology of the Elk Mountain and Tabernacle Butte area, Wyoming. Bull Am Mus Nat Hist 117(3):129–184
Meszoely CAM (1970) North American fossil anguid lizards. Bull Mus Comp Zool 139(2):87–150
Montellano M, Wilson GP, Alvarez-Reyes G, Hernández-Rivera R, Quintero E, Aranda-Mante CA (2005) New material of Polyglyphanodon bajaensis from El Rosario Baja California, Mexico. J Vertebr Paleontol 25 (Supplement):93A
Müller J, Hipsley CA, Head JJ, Kardjilov N, Hilger A, Wuttke M, Reisz RR (2011) Eocene lizard from Germany reveals amphisbaenian origins. Nature 473(7347):364–367
Nockleberg WJ, Parfenov LM, Monger JWH, Norton IO, Khanchuk AI, Stone DB, Scotese CR, Scholl DW, Fujita K (2000) Phanerozoic tectonic evolution of the Circum-North Pacific. US Geol Surv Prof Pap 1626:1–122
Norell MA, Gao K (1997) Braincase and phylogenetic relationships of Estesia mongoliensis from the Late Cretaceous of the Gobi Desert and the recognition of a new clade of lizards. Am Mus Nov 3211:1–25
Nydam RL (1999) Polyglyphanodontinae (Squamata: Teiidae) from the medial and Late Cretaceous: new records from Utah, U.S.A. and Baja California del Norte, Mexico. In: Gillette DD (ed.), Vertebrate Paleontology in Utah. Utah Geol Surv Misc Pub 99–1:303–317
Nydam RL (2000) A new taxon of helodermatid-like lizard from the Albian-Cenomanian of Utah. J Vertebr Paleontol 20(2):285–294
Nydam RL (2002) Lizards of the Mussentuchit Local Fauna (Albian-Cenomanian) and comments on the evolution of the Cretaceous lizard fauna of North America. J Vertebr Paleontol 22(3):645–660
Nydam RL (2013) Lizards and snakes from the Cenomanian through Campanian of Southern Utah: Filling the gap in the fossil record of Squamata from the Late Cretaceous of the Western Interior of North America. In: Titus AL, Loewen MA (eds) At the top of the grand staircase: The Late Cretaceous of southern Utah. Indiana University Press, Bloomington, pp 370–423
Nydam RL, Cifelli RL (2002a) Lizards from the Lower Cretaceous (Aptian-Albian) Antlers and Cloverly formations. J Vertebr Paleontol 22(2):286–298
Nydam RL, Cifelli RL (2002b) A new teiid lizard from the Cedar Mountain Formation (Albian-Cenomanian boundary) of Utah. J Vertebr Paleontol 22(2):276–285
Nydam RL, Cifelli RL (2005) New data on the dentition of the scincomorphan lizard Polyglyphanodon sternbergi. Acta Palaeontol Pol 50(1):73–78
Nydam RL, Fitzpatrick B (2009) The occurrence of Contogenys-like lizards in the Late Cretaceous and Early Tertiary of the Western Interior or the U.S.A. J Vertebr Paleontol 29(3):677–701
Nydam RL, Lofgren DL (2008) Preliminary report on lizards from the Goler Formation (Paleocene) of California. J Vertebr Paleontol 28(3, supplement):121A
Nydam RL, Voci GE (2007) Teiid-like scincomorphan lizards from the Late Cretaceous (Campanian) of Southern Utah. J Herpetol 41(2):215–223
Nydam RL, Gauthier JA, Chiment JJ (2000) The mammal-like teeth of the Late Cretaceous lizard Peneteius aquilonius Estes 1969 (Squamata, Teiidae). J Vertebr Paleontol 20:628–631
Nydam RL, Eaton JG, Sankey J (2007) New taxa of transversely-toothed lizards (Squamata: Scincomorpha) and new information on the evolutionary history of "teiids". J Paleont 81(3):538–549
Nydam RL, Caldwell MW, Fanti F (2010) Borioteiioidean lizard skulls from Kleskun Hill (Wapiti Formation; Upper Campanian), West-central Alberta, Canada. J Vertebr Paleontol 30(4):1–10
Nydam RL, Rowe TB, Cifelli RL (2013) Lizards and snakes of the Terlingua Local Fauna (late Campanian), Aguja Formation, Texas with comments on the distribution of paracontemporaneous squamates throughout the Western Interior of North America. J Vertebr Paleontol 33(5):1081–1099
Ogg JG, Hinnov L (2012) Cretaceous. In: Gradstein FM, Ogg JG, Schmitz M, Ogg G (eds) The Geologic Time Scale 2012, vol 2. Elsevier, Amsterdam, pp 793–853
Olsen T (1995) Fluvial and fluvio-lacustrine facies and depositional environments of the Maastrichtian to Paleocene North Horn Formation, Price Canyon, Utah. Mount Geol 32(2):27–44
Oreska MPJ CMT, Dzikiewicz KM (2013) Vertebrate paleontology of the Cloverly Formation (Lower Cretaceous), I: faunal composition, biogeographic relationships, and sampling. J Vertebr Paleontol 33(2):264–292
Ostrom JH (1970) Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana. Bull Peabody Mus Nat Hist 35(35):1–234
Palci A, Caldwell MW (2007) Vestigial forelimbs and axial elongation in a 95 million-year-old non-snake squamate. J Vertebr Paleontol 27(1):1–7
Payenberg THD, Braman DR, Davis DW, Miall AD (2002) Litho- and chronostratigraphic relationships of the Santonian–Campanian Milk River Formation in southern Alberta and Eagle Formation in Montana utilising stratigraphy, U–Pb geochronology, and palynology. Can J Earth Sci 39(10):1553–1577
Peng J, Russell AP, Brinkman DB (2001) Vertebrate microsite assemblages (exclusive of mammals) from the Foremost and Oldman Formations of the Judith River Group (Campanian) of southeastern Alberta: an illustrated guide. Curatorial Section, Provincial Museum of Alberta. Nat Hist Occ Pap 25:1–54
Pregill GK, Gauthier JA, Greene HW (1986) The evolution of helodermatid squamates, with description of a new taxon and an overview of Varanoidea. Trans San Diego Soc Nat Hist 21(11):167–202
Prothero DR (1983) The oldest mammalian petrosals from North America. J Paleont 57(5):1040–1046
Prothero DR, Estes R (1980) Late Jurassic lizards from Como Bluff, Wyoming and their palaeobiogeographic significance. Nature 286:484–486
Rasmussen TE, Callison G (1981) A new herbivorous sphenodontid (Rhynchocephalia: Reptilia) from the Jurassic of Colorado. J Paleont 55(5):1109–1116
Renne PR, Fulford MM, Busby-Spera C (1991) High resolution 40AR/39AR chronostratigraphy of the Late Cretaceous El Gallo Formation, Baja California del Norte, Mexico. Geophys Res Lett 18(3):459–462
Reynoso V-H (1995) Lepidosaurian reptiles from the Cantera Tlayua (Albian), Tepexi de Rodríguez, Puebla, Mexico. Abstract from II International Symposium on Lithographic Limestones, Lleida-Cuenca (Spain), 9–16 July 1995:131–132
Reynoso V-H (1996) A primitive lizard from the Early Cretaceous of Mexico and the phylogenetic position of early lizards. J Vertebr Paleontol 16(supplement to 3):60A
Reynoso V-H (1997) A "beaded" sphenodontian (Diapsida: Lepidosauria) from the Early Cretaceous of central Mexico. J Vertebr Paleontol 17(1):52–59
Reynoso V-H (1998) Huehuecuetzpalli mixtecus gen. et sp. nov.; a basal squamate (Reptilia) from the Early Cretaceous of Tepexi de Rodrígues, central Mexico. Philos Trans R Soc Lond B 353:1–23
Reynoso V-H (2000) An unusual aquatic Sphenodontian (Reptilia: Diapsida) from the Tlayua Formation (Albian), Central Mexico. J Paleont 74(1):133–148
Reynoso V-H, Callison G (2000) A new scincomorph lizard from the Early Cretaceous of Puebla, Mexico. Zool J Linn Soc 130:183–212
Richter A (1994) Lacertilia aus der Unteren Kreide von Uña und Galve (Spanien) und Anoual (Marokko). Berliner Geowiss Abh (E) 14:1–147
Rieppel O, Head JJ (2004) New specimens of the fossil snake genus Eupodophis Rage and Escuillié, from Cenomanian (Late Cretaceous) of Lebanon. Memo Soc Ital Sci Nat Mus Civ Stor Nat Milano 32(2):1–26
Rieppel O, Zaher H, Tchernov E, Polcyn MJ (2003) The anatomy and relationships of Haasiophis terrasanctus, a fossil snake with well-developed hind limbs from the mid-cretaceous of the middle east. J Paleont 77(3):536–558
Roberts EM, Deino AL, Chan MA (2005) 40Ar/39 Ar age of the Kaiparowits Formation, southern Utah, and correlation of contemporaneous Campanian strata and vertebrate faunas along the margin of the Western Interior Basin. Cretac Res 26:307–318
Rogers RR, Swisher Iii CC, Horner JR (1993) 40Ar/39Ar age and correlation of the nonmarine Two Medicine Formation (Upper Cretaceous), northwestern Montana, U.S.A. Can J Earth Sci 30(5):1066–1075
Rose KD, Ciffeli RL, Lipka TR (2001) Second triconodont dentary from the Early Cretaceous of Maryland. J Vertebr Paleontol 21(3):628–632
Rowe TB, Cifelli RL, Lehman TM, Weil A (1992) The Campanian Terlingua local fauna, with summary of other vertebrates from the Aguja Formation, Trans-Pecos Texas. J Vertebr Paleontol 12(4):472–493
Sahni A (1972) The Vertebrate Fauna of the Judith River Formation, Montana. Bull Am Mus Nat Hist 147(6):321–412
Sereno PC (1999) Dinosaurian biogeography: vicariance, dispersal, and regional extinction. In: Tomida Y, Rich TH, Vickers-Rich P (eds) Proceedings of the Second Annual Gondwanan Dinosaur Symposium, vol 15, National Science Museum Monographs, pp 249–257
Sloan RE, Van Valen L (1965) Cretaceous Mammals from Montana. Science 148(3667):220–227
Smith KT (2009) A new lizard assemblage from the earliest Eocene (zone Wa0) of the Bighorn Basin, Wyoming, USA: biogeography during the warmest interval of the cenozoic. J Syst Palaeontol 7(3):299–358
Smith KT (2013) New constraints on the evolution of the snake clades Ungaliophiinae, Loxocemidae and Colubridae (Serpentes), with comments on the fossil history of erycineboids in North America. Zool Anz - J Comp Zool 252(2):157–182
Sternberg CM (1951) The lizard Chamops from the Wapiti Formation of Northern Alberta: Polydontosaurus grandis not a lizard. Bull Nat Mus Can 123:256–258
Sullivan RM (1981) Fossil lizards from the San Juan Basin, New Mexico. In: Lucas SG, Rigby JK Jr, Kues BS (eds) Advances in San Juan Basin Paleontology. University of New Mexico Press, Albuquerque, pp 76–88
Sullivan RM (1982) Fossil Lizards from Swain Quarry "Fort Union Formation," Middle Paleocene (Torrejonian), Carbon County, Wyoming. J Paleont 56(4):996–1010
Sullivan RM (1985) A new middle Paleocene (Torrejonian) rhineurid Amphisbaenian, Plesiorhineura tsentasi new genus, new species, from the San Juan Basin, New Mexico. J Paleont 59(6):1481–1485
Sullivan RM (1986) A new fossil anguid lizard Parodaxosaurus sanjuanensis, new genus, new species from the middle Paleocene (Torrejonian) Nacimiento Formation, San Juan Basin, New Mexico. J Herpetol 20(1):109–111
Sullivan RM (1991) Paleocene Caudata and Squamata from Gidley and Silberling Quarries, Montana. J Vertebr Paleontol 11(3):293–301
Sullivan RM, Lucas SG (1986) Annotated list of lower vertebrates from the Paleocene Nacimiento Formation (Puercan-Torrejonian), San Juan Basin, New Mexico. J Herpetol 20(2):202–209
Sullivan RM, Lucas SG (1996) Paleoscincosaurus middletoni, new genus and species (Squamata: ?Scincidae) from the Early Paleocene (Puercan) Denver Formation, Colorado. J Vertebr Paleontol 16(4):666–672
Tchernov E, Rieppel O, Zaher H, Polcyn MJ, Jacobs LL (2000) A fossil snake with limbs. Science 281:2010–2012
Titus AL, Powell JD, Roberts EM, Sampson SD, Pollock SL, Kirkland JI, Albright LB (2005) Late Cretaceous stratigraphy, depositional environments, and macrovertebrate paleontology of the Kaiparowits Plateau, Grand Staircase–Escalante National Monument, Utah. In: Pederson J, Dehler CM (eds.), Interior Western United States: Geological Society of America Field Guide 6, pp. 101–128
Titus AL, Roberts EM, Albright LB (2013) Geologic overview. In: Titus AL, Loewen MA (eds) At the top of the Grand Staircase: The Late Cretaceous of Southern Utah. Indiana University Press, Oxford, pp 13–41
Waldman M (1970) A teiid lizard jaw from the Cretaceous of Alberta, Canada. Can J Earth Sci 7(2):542–547
Wiens JJ, Kuczynski CA, Townsend T, Reeder TW, Mulcahy DG, Sites JW (2010) Combining phylogenomics and fossils in higher-level squamate reptile phylogeny: molecular data change the placement of fossil taxa. Syst Biol 59(6):674–688
Williamson TE, Weil A (2004) First occurence of the Teiid lizard Peneteius from the latest Cretaceous Naashoibito Member, Kirtland Formation, San Juan Basin, New Mexico. N Mex Geol 26(2):65
Wilson JA, Mohabey DM, Peters SE, Head JJ (2010) Predation upon hatchling dinosaurs by a new snake from the Late Cretaceous of India. PLoS Biol 8(3):e1000322
Winkler DA, Murry PA, Jacobs LL (1990) Early Cretaceous (Comanchean) vertebrates of central Texas. J Vertebr Paleontol 10(1):95–116
Yi H-Y, Norell MA (2013) New materials of Estesia mongoliensis (Squamata: Anguimorpha) and the evolution of venom grooves in lizards. Am Mus Nov 3767:1–3
Zaher H, ApesteguÍA S, Scanferla CA (2009) The anatomy of the upper cretaceous snake Najash rionegrina Apesteguía & Zaher, 2006, and the evolution of limblessness in snakes. Zool J Linn Soc 156(4):801–826
Zaher H, Scanferla CA (2012) The skull of the Upper Cretaceous snake Dinilysia patagonica Smith-Woodward, 1901, and its phylogenetic position revisited. Zool J Linn Soc 164(1):194–238
Zanno LE, Makovicky PJ (2011) On the earliest record of Cretaceous tyrannosauroids in western North America: implications for an Early Cretaceous Laurasian interchange event. Hist Biol 23(4):317–325
Acknowledgements
I wish to express my appreciation to Jim Gardner for inviting me to participate in the Insights from the Fossil Record into the Evolution of Extant Amphibians and Reptiles symposium at the World Herpetological Conference in Vancouver as well as his generous offer for me to co-edit this collection of contributions. I would further wish to offer my sincerest gratitude to the very patient Dr. Sinje Weber of the editorial team for Palaeobiodiversity and Palaeoenvironments. She and her family were also very kind hosts during my visit to Frankfurt in the summer of 2012. I have also benefitted greatly from very informative, and at times spirited, conversations about fossil squamates with Anjan Bhullar, Michael Caldwell, Jack Conrad, David DeMar Jr., Bob Denton, Susan Evans, Jacques Gauthier, Jason Head, Johannes Müller, and Krister Smith. Don Brinkman was helpful in sorting out the current stratigraphic interpretations of southern Alberta though any errors herein are my own. Cynthia Crane, Dave DeMar, and Bob Denton generously shared some of their preliminary results. Brent Adrian assisted with the preparation of the figures. Reviews by Krister Smith and Johannes Müller were instrumental in the improvement of this paper. Although I have tried to be comprehensive in this review, it is always possible that a reference may have been unintentionally overlooked. Support for this work was provided in part by Midwestern University Intramural Research Funds.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is a contribution to the special issue “Mesozoic and Cenozoic lissamphibian and squamate assemblages of Laurasia”
Rights and permissions
About this article
Cite this article
Nydam, R.L. Squamates from the Jurassic and Cretaceous of North America. Palaeobio Palaeoenv 93, 535–565 (2013). https://doi.org/10.1007/s12549-013-0129-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-013-0129-5