
Abstract—
The review summarizes the results of recent studies of microbial communities of the Lake Baikal sediments obtained using diverse techniques. In the sediments of the areas of stable sedimentation metabarcoding revealed predominance of members of the phyla Alpha- and Gammaproteobacteria (including Betaproteobacteriales), Bacteroidetes, Acidobacteria, Verrucomicrobia, and Thaumarchaeota, which are also common in other freshwater lakes. In the areas of discharge of gas-bearing mineralized fluids, the structure of microbial communities varied depending on the presence of electron acceptors and intensity and component composition of gas-bearing fluids responsible for microbial migration from the deep zone to the upper sediment layers and vice versa. Methanogenic archaea detected in Baikal sediments belonged to the groups capable of all four known catabolic pathways of methanogenesis: hydrogenotrophic, acetoclastic, methylotrophic, and hydrogen-dependent methylotrophic ones. Predominant members of the Baikal archaeal community, hydrogenotrophic methanogens of the family Methanoregulaceae (genera Methanoregula and Methano-sphaerula, as well as uncultured lineages), hydrogen-dependent methylotrophic archaea of the order Methanomassiliicoccales, and acetoclastic methanogens of the family Methanosaetaceae (genus Methanothrix (Methanosaeta)), were the same as in methanogenic communities of other freshwater lakes. Experimental evidence was obtained for anaerobic methane oxidation (AOM) via the nitrate- and nitrite-dependent pathways by archaea of the ANME-2d subcluster and bacteria of the phylum NC10. Structures of the 16S rRNA genes, mcrA, and pmoA exhibited high identity to those of the known freshwater organisms performing this process. Diversity of microbial communities at the sites of natural oil seepage differed at the order and family levels, as well as by the presence of alkane hydroxylases in the genes of the cultured species.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
SEDIMENTS OF LAKE BAIKAL AS A HABITAT FOR MICROORGANISMS
Lake Baikal is the deepest and the most ancient lake in the world. The total volume of its sediments is approximately 75 thousand km3, and their maximum thickness is more than 9 km (Logachev, 2003). The Baikal depression, with a maximum depth of 1642 m, is located in the center of a tectonically active rift zone; the oldest deposits date back to the Oligocene age (Hutchinson et al., 1992; Mats et al., 2001). The depression is divided into three basins: South, Central, and North. They are separated from each other by the following seamounts: the Buguldeika Saddle and the Academician Ridge (Fig. 1). The sediments in the three deep-water parts of the lake have an identical chemical composition, which is a result of homogenization of fine fractions and incoming terrigenous material carried by steady currents (Gvozdkov, 1998). The entire central part of the lake bottom is occupied by diatomaceous silts; the average concentration of biogenic silicon in the surface silts of the pelagic zone is 19%. Up to 90% of the bottom is covered with sediments containing. more than 10% of biogenic SiO2. (Granina, 2008). In the Lake Baikal sediments, the average Corg concentration is 1.9%; in the sediments of South and Central Baikal, it is 1.5–2.5%, while in the North basin, its concentration is 1.0–1.3% (Vykhristyuk, 1980). These values are comparable to those determined in oceanic sediments in highly productive continental margins (Qiu et al., 1993). According to the data reported by L.A. Vykhristyuk (1980), the composition of the buried organic matter (OM) includes humic acids and cellulose, as well as significantly decomposed remains of higher terrestrial plants and phytoplankton, which are highly resistant to further degradation.

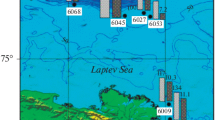
Schematic map of Lake Baikal and areas of research: ⚫, mud volcano; ⚪, methane seep; ◻, oil seep; △, thermal vent.
Geochemical processes in the sediments of Lake Baikal have been the subject of research by many authors (Mizandrontsev, 1975;Pogodaeva et al., 2007, 2017; Granina, 2008; Och et al., 2012). One of the most important characteristics of Lake Baikal is a widespread oxidative stage of early diagenesis (Granina, 2008). The depth of oxygen penetration into the sediments is from <2 to >50 mm; the thickness of the oxidized layer is from 0.2 to >30 cm. The pH values in the sediment are mainly from 6.9 to 7.4, with the range from 6 to >8 (Mizandrontsev, 1975; Granina, 2008). According to I.B. Mizandrontsev (1978), deep penetration of oxygen is due to the slow accumulation of sediments in Lake Baikal, and the surface sediments are oxidized on most of the bottom (Granina, 2008). Sedimentation rates in open Baikal vary from 0.015 to >1 mm per year (Coleman et al., 1993; Vologina et al., 2000).
Pore water of sediments in the areas with normal sedimentation inherits the chemical composition of lake water; it is characterized by a homogeneous salt composition. The most common ions dissolved in Lake Baikal are \({\text{HCO}}_{3}^{ - }\) and Ca2+. Regarding biogeochemical cycles, a very low concentration of dissolved \({\text{SO}}_{4}^{{2 - }}\) in the water of Lake Baikal (0.05 mM) is the most important difference from seawater (28.9 mM) (Aloisi et al., 2019). Other ions that compose the majority of dissolved salts in seawater (Cl–, Na+, Mg2+, and K+) are present in the water of Lake Baikal at very low concentrations (≤0.15 mM) (Pogodaeva et al., 2007, 2017). During diagenetic transformations, the total concentration of ions in the pore water increases with the depth of the sediment due to an increase in the concentrations of bicarbonate and calcium ions. Due to the lack of electron acceptors \({\text{SO}}_{4}^{{2 - }}\) and \({\text{NO}}_{3}^{ - },\) most dissolved inorganic matter, formed after OM decomposition and the subsequent redox reactions, is in the form of CO2, rather than \({\text{HCO}}_{3}^{ - },\) which results in lower pH values in diagenetic environments (Mizandrontsev, 1975). OM replenishment occurs annually due to the massive development of diatoms in spring and autumn (Votintsev et al., 1975) and of picoplankton in summer (Nagata et al., 1994). Using sedimentation traps, the presence of diatom shells and organic matter of autochthonous origin has been shown at all depths of the water column (Vologina and Shturm, 2017), while only 10% of picoplankton reaches the bottom.
MICROBIOLOGICAL STUDIES OF THE SEDIMENTS IN LAKE BAIKAL
Microbiological studies of the sediments of Lake Baikal have been carried out since the beginning of the previous century using approaches and methods that are widely used in aquatic microbiology. Research fields and main results have been reported in monographs and numerous papers (Mikroorganizmy…, 1985; Namsaraev and Zemskaya, 2000; Mikrobiologicheskoe nasledie…, 2004). Leading microbiologists of Russia worked at Lake Baikal, including S.I. Kuz-netsov, A.E. Criss, Yu.I. Sorokin, A.P. Romanova, A.G. Rodina, G.A. Dubinina, V.N. Maksimov, E.A. Maksimova, and B.B. Namsaraev. Their publications provide information about microbial abundance in the Lake Baikal sediments in various geochemical environments. The distribution of microbial physiological groups in different areas and at different depths of the sedimentary layers has been analyzed; the metabolic activity of microbial communities, including those in the areas with increased anthropogenic impact, has been assessed (Mikroorganizmy…, 1985; Namsaraev and Zemskaya, 2000; Mikrobiologicheskoe nasledie…, 2004). A new stage of research into the role of microbial communities in the sediments has begun during the period of large-scale geological and geophysical studies carried out by international scientific teams at Lake Baikal since the late 1990s. During these years, the main attention was paid to the study of microorganisms participating in different stages of OM decomposition and to assessment of the rates of the degradation of organic matter in the sedimentary layers of Lake Baikal (Maksimova and Maksimov, 1989; Namsaraev and Zemskaya, 2000; Pimenov et al., 2014). The development of certain physiological microbial groups in sediments was found to be determined by availability of the substrates, oxidized or reduced conditions at certain depths of the sedimentary strata, and the influx of gas-bearing mineralized fluids from deep sediments. Annual supply of organic matter to the sediments and the low concentrations of sulfate ions determine the predominance of the methane generation process at the final stages of destruction.
MICROBIAL DIVERSITY IN THE SEDIMENTS OF LAKE BAIKAL
Progress in assessment of the diversity of microbial communities has been achieved using molecular biological methods and approaches. Application of the methods for the 16S rRNA and mcrA gene sequencing, data on the diversity and functioning of microorganisms in the sediments with different pore water composition were obtained (Shubenkova et al., 2005; Kadnikov et al., 2012; Chernitsyna et al., 2016; Lomakina et al., 2018). Similarly to other freshwater lakes, the taxonomic composition of microbial communities at different sediment depths in Lake Baikal varied and was determined by the presence of oxidants, the concentration and composition of available OM, as well as of other vital elements (Kuznetsov, 1970; Zavarzin, 2003; Huang et al., 2019; Han et al., 2020). In some cases, microbiomes in the sediments of the same area and similar environments differed in the contribution of the predominant taxa (Zemskaya et al., 2018; Zakharova et al., 2018). In other cases, their similarity to microbiomes from other areas and sediment layers was noted (Shubenkova et al., 2005; Kadnikov et al., 2012; Zemskaya et al., 2015a; Chernitsina et al., 2016; Bukin et al., 2016; Lomakina et al., 2018). Organotrophic bacteria typical of freshwater lakes (Actinobacteria, Proteobacteria, and Verrucomicrobia) predominated in the communities of oxidized sediments and to the lower border of oxygen penetration (2 cm). For these bacteria, participation in the first stages of degradation of organic compounds was shown (Newton et al., 2011). A significant contribution of members of the first two phyla to bacterial communities is characteristic of the oxidized layers of all studied sediments of Lake Baikal, while predominance of the phylum Verrucomicrobia was observed only in some communities (Kadnikov et al., 2012; Zemskaya et al., 2015a, 2018; Zakharova et al., 2018). According to the data on genomic structure of the microorganisms from the same taxonomic groups that were identified in the aerobic zone of Lake Baikal sediments (mainly Alpha-, Beta-, and Gammaproteobacteria, Bacteroidetes, Acidobacteria, Verrucomicrobia, and Planctomycetes), as well as of their closest relatives, the oxidized zone is colonized by aerobic and facultatively anaerobic bacteria that oxidize carbohydrates, amino acids, and fatty acids (Yoon et al., 2008; Pujalte et al., 2014; Sun et al., 2016) and aerobic nitrifying archaea (MGI and SCG) (Spang et al., 2012; Cabello-Yeves et al., 2020; Han et al., 2020). As a rule, at the border of the oxic and anoxic zones, the contribution of the members of Bacteroidetes belonging to the families Flavobacteriaceae and Sphingobacteriaceae, as well as of unclassified microorganisms, increased, with a high contribution of the taxa predominating in the upper sediment layers. Predominant sequences retrieved from the anoxic zone of the sediment, in particular, from the zone of occurrence of the ferromanganese formation (FMF), belonged to organisms of unknown taxonomic position, positively correlating with the concentration of manganese ions, and to Alpha- and Betaproteobacteria, associated with the content of iron ions (Zemskaya et al., 2018). According to Torres et al. (2014), Mn(IV) (and potentially Fe(III)) can act as oxidants in the microbiologically mediated nitrification process. Indeed, in the communities of the core studied by us (below the oxygen penetration limit), the contribution of nitrite-oxidizing bacteria of the phylum Nitrospirae increased, and the presence of archaea of the phylum Thaumarchaeota, which are responsible for the processes of ammonium oxidation to nitrite, was shown. Their participation in nitrogen metabolism was confirmed by metagenomic analysis of genomes (MAGs) from the deep zone of South Baikal (Cabello-Jeves et al., 2020). In the genomes of microorganisms, including those belonging to these taxa, the genes responsible for oxidation of ammonium and nitrite, decomposition of urea and cyanate, or assimilation of nitrates/nitrites, as well as denitrification, were revealed. Members of the phylum Firmicutes, the cultured strains of which were shown to participate in the oxidation of Fe(II) and Mn(II) in Lake Baikal (Zakharova et al., 2010), constituted an insignificant part of the community from the upper layer of the FMF and contributed more significantly to the bottom layer (7.6%). Most of them belonged to the genera Bacillus and Clostridium, the identity of which to the cultured ferromanganese bacteria from Lake Baikal was not confirmed. During the study of the microbial communities from the sediments in Lake Stechlin and five lakes of Central Switzerland, changes in the composition of microbial communities during the change from oxidized to reduced conditions were noted (Wurzbacher et al., 2017; Han et al., 2020). The authors believe that the diversity of communities in these zones was determined by changes in OM sources, respiratory reactions, and selective survival of microorganisms. Therefore, the taxa that were already present in the sediments during sedimentation may be better adapted to the conditions of low availability of energy sources and should predominate with an increase in the sediment depth (Lever et al., 2015; Starnawski et al., 2017; Rissanen et al., 2019). This agrees with our data on the diversity of microbial communities in the sediments of South Baikal. We noted an increase in the contribution of the members of Bacteroidetes, Alpha-, and Betaproteobacteria to the microbial community of the sediment at a depth of 8–9 cm, while in the surface sediment layer they were detected as minor components. According to the data (Kolman et al., 1993; Vologina et al., 2000), the estimated age of the sediment at this depth is 100–500 years, and, probably, members of these taxa proved to be more adapted to the existence in low-energy environments. Cultured members of the taxa from this zone are chemoorganotrophs, chemolithotrophs, and facultative photoheterotrophs; they are found in various biotopes. Noteworthy, the bacterial community in deep (up to 4 m) layers of sediments in Lake Baikal is also diverse (Chernitsyna et al., 2016; Zemskaya et al., 2015a; Bukin et al., 2016; Lomakina et al., 2018).
Archaea are important components of microbial communities in the Lake Baikal sediments. Their presence was recorded starting from the first centimeters of oxidized sediments, as well as their higher diversity in buried oxidized sediments and in the layers with high concentrations of manganese and iron (Zemskaya et al., 2018). The phylum Thaumarchaeota (Marine Group I lineage), which is widespread in soil and aquatic ecosystems and participates in aerobic ammonium oxidation, is one of the predominant taxa in the Lake Baikal sediments (Walker et al., 2010). In the sediments from different areas of Lake Baikal, clusters of sequences related to the cultured species Nitrosopumilus maritimus of the MGI lineage and to uncultured members of this phylum were identified (Lomakina et al., 2018). Representatives of the phyla Euryarchaeota (Uncultured Methanomicrobia), Ba-thyarchaeota, Aigarchaeota, Aenigmarchaeota, Woesearchaeota, Thaumarchaeota, and Hadesarchaeota (former SAGMEG-1) were found in the sediments of the reference areas. Members of the uncultured Marine Benthic Group D (MBG-D) lineage of the order Thermoprofundales, previously related to Thermoplasmatales and widespread in freshwater and marine habitats, were also identified (Schubert et al., 2011). Their significant contribution was noted in the communities of methane seeps (Beal et al., 2009) and sediments from Lake Pavin (Borrel et al., 2012). It is assumed that they can carry out protein degradation in marine sediments, as well as participate in the processes of methanogenesis or anaerobic oxidation of methane (AOM) (Schubert et al., 2011).
A different composition of archaea was observed in sedimentary layers of the zones of seepage of mineralized gas-bearing fluids. Pore water at these sites is characterized as bicarbonate-sulfate, sulfate, or chloride water (Mizandrontsev, 1975; Klerks et al., 2003; Granina, 2008; Minami et al., 2010; Zemskaya et al., 2015a; Pogodaeva et al., 2020). The composition of pore water was significantly affected by the intensity of fluid flows and the specific composition of chemical components (Aloisi et al., 2019). The mechanisms of migratory fluid flows in the areas of mud volcanoes and methane seeps proved to affect also the composition and structure of microbial communities (Shubenkova et al., 2005; Kadnikov et al., 2012; Lomakina et al., 2014, 2018; Zemskaya et al., 2015a; Chernitsyna et al., 2016; Bukin et al., 2016). In particular, in the area of the Krasny Yar methane seep, the inflow of oxygen-saturated surface sediments and aerobic microorganisms to a depth of more than 1 m was shown. In the sediments of the seep on the Posolsk Bank uplift, the deep microflora was found on the sediment surface due to the functioning of the fluid loop (Nauds et al., 2012; Chernitsyna et al., 2016; Pogodaeva et al., 2020). The special routes of migration of fluid flows circulating in the zone of stability of gas hydrates (400 m) explain the presence of significant numbers of sequences assigned to the phyla Chloroflexi and Acidobacteria, as well as to the candidate phyla Aminicenantes and Atribacteria, in the surface layers of sediments, and of the phylum Deinococcus–Thermus in deep sediments. In the zones with especially intense flows of mineralized fluids and gases, formation of microbial mats of two types was noted: with the predominance of colorless sulfur bacteria (Kuznetsov et al., 1991) or methanotrophic bacteria (Zemskaya et al., 2015b). Microbial communities, the functioning of which is based on methanotrophy, are characteristic of forest soils, meadows, rice fields, wetlands, and sediments in freshwater, saline, or alkaline lakes (Ding and Valentine, 2008). The presence of large fields of microbial mats was detected over massive layers of methane hydrates in the bathypelagic zone of Central Baikal using the Mir submersibles, where they provided for the vital activity of a wide range of benthic animals (Zemskaya et al., 2012; Sitnikova et al., 2017). Intense methane fluxes formed due to the dissolution of methane hydrates and the activity of hydrogenotrophic methanogens allowed the development of microbial mats of methanotrophic and methylotrophic bacteria in the oxic layer (Kadnikov et al., 2012; Zemskaya et al., 2015b). These mats were consumed by benthic animals, as was confirmed by the carbon isotopic composition of benthic animals from this area (Zemskaya et al., 2012). In the community of microbial mats, members of the genera Methylobacter, Methylophilus, and Methylotenera, which inhabit various cold-water ecosystems (Dedysh et al., 2001; Pacheco-Oliver et al., 2002) and possess enzyme systems that are responsible for various steps of methane oxidation to CO2 and biomass, were identified. Under the mat layer, the proportion of archaea was approximately 2/3 of the entire microbial community in the surface sediment layers of this area (Kadnikov et al., 2012), including anaerobic methanotrophs “Candidatus Methanoperedens sp.” (ANME-2d) and hydrogenotrophic methanogens of the genus Methano-sphaerula. In the area of intense mineralized flows in other areas of Lake Baikal, microbial mats in which the dominant role belonged to colorless sulfur bacteria of the genus Thioploca were also formed (Kuznetsov et al., 1991; Zemskaya et al., 2001). Similar to marine members of this genus, sulfur bacteria inhabiting Lake Baikal carry out the process of sulfide oxidation coupled to nitrate reduction; they are chemoorganoheterotrophs (Zemskaya et al., 2001). The functioning of the community of bacterial mats in this area, as well as in the areas of marine methane seeps, is provided by chemosynthesis and methanotrophy (Namsaraev and Zemskaya, 2000; Ding and Valentine, 2008).
MICROORGANISMS INVOLVED IN METHANE GENERATION PROCESS
Methane was previously shown to be the main terminal product of OM degradation in the sediments of Lake Baikal (Namsaraev and Zemskaya, 2000; Dagurova et al., 2004), and the processes of its biogenic formation can extend hundreds of meters deep into the sedimentary layer of the lake (Kuz’min et al., 2001). In the zones of hydrocarbon discharge, silts of the low-temperature Frolikha vent, and the sediments of several areas with a high content of organic matter, 5.0–534.7 μL of CH4/kg per day were generated, which corresponded to the rates of methane generation in other oligotrophic and mesotrophic freshwater lakes (Duc et al., 2010; Fuchs et al., 2016; Yang et al., 2020). However, the rate of methanogenesis in the reference regions of the pelagic zone of the lake was lower (0.01–32.29 μL CH4/kg per day) and comparable to that in the sediments of highly productive areas of the ocean and saline meromictic lakes Shira and Shunet (Capone and Kiene, 1988; Kallistova et al., 2006). Similar to many other freshwater lakes, due to low sulfate concentrations in pore water, the upper boundary of the methane generation zone in the sediments of Lake Baikal is determined only by the depth of O2 penetration (Namsaraev and Zemskaya, 2000; Koizumi et al., 2003; Conrad et al., 2007). For instance, in the areas of seeps, mud volcanoes, and an underwater low-temperature vent, active methane production was recorded already in the first centimeter of sediments reduced from the surface (Namsaraev and Zemskaya, 2000; Dagurova et al., 2004), while methane and ethane supplied with a deep fluid and bound in the structure of gas hydrates, were of biogenic, thermogenic, or mixed biogenic-thermogenic origin (Klerks et al., 2003; Kalmychkov et al., 2006). The vertical distribution pattern of methane generation in such areas varied, although high values of methanogenesis rates were often recorded in surface silts to the depths of 20–30 cm, with subsequent bursts of activity in separate deep layers (Namsaraev and Zemskaya, 2000; Dagurova et al., 2004). Studies carried out in the zones unaffected by hydrocarbon discharge showed an almost linear increase in methane content in the sediments up to the depths of 2–2.5 m. This correlated with the content of bicarbonate ion released during diagenesis of organic matter (Pogodaeva et al., 2017). However, in some cases (from 2.5 m to the lowest exposed layers, during the transition to blue-gray clays), an abrupt increase in methane concentrations and rates of methane generation was noted, which also confirmed the presence of active methanogenic communities in the deep sedimentary layers of Lake Baikal (Namsaraev and Zemskaya, 2000).
Methanogenic archaea are known to be able to use a very limited range of substrates formed at the initial stages of anaerobic OM degradation as carbon and energy sources. Among them, H2 : CO2, acetate, and C1-methylated compounds (methanol, methylamines, methyl sulfides, etc.) were noted. These compounds are consumed by hydrogenotrophic, acetoclastic, and methylotrophic methanogens, respectively (Thauer et al., 2008). In the sediments of deep-water zones of stratified lakes, the contribution of acetoclastic methanogens often increases, which is due to the activity of homoacetogenic microorganisms (Schulz and Conrad, 1996; Nozhevnikova et al., 2007). Under psychrophilic conditions, homoacetogens can efficiently compete with hydrogenotrophic methanogens for H2, reducing CO2 to acetate; the latter is accumulated in the sediment and disproportionated to methane by acetoclastic methanogens (Kotsyurbenko et al., 2005). However, in Lake Baikal, predominance of acetoclastic methanogenesis was shown only in the sediments of shallow-water areas. In sediments of the deep-water zone, 75–100% of methane was generated from H2 : CO2 and acetate concentrations varied from 4 to 98 μmol/L (Namsaraev and Zemskaya, 2000). A high contribution of hydrogenotrophic methanogenesis (99%) was noted even in the area of the low-temperature Frolikha vent, in which acetate content can reach 1 mmol/L in pore water and a large proportion (25%) of acetoclastic methanogens was determined among archaea (Lomakina et al., 2018). The exact cause of this phenomenon has not been clarified. Hydrogen concentration in the gas composition of the sedimentary layers of Lake Baikal does not exceed 0.0632 vol % (Kuz’min et al., 2001). Moreover, the temperature of sediments in the deep-water area of the lake (≤4°C) makes syntrophic acetate oxidation thermodynamically unfavorable, which does not explain the prevailing hydrogenotrophic pathway by the additional inflow of hydrogen or utilization of acetate bypassing acetoclastic methanogenesis, as in Lakes Kivu and Kinneret (Nulssein et al., 2001; Pasche et al., 2011). Possibly, the high contribution of CO2 reduction may be due to its greater thermodynamic advantage under the conditions of the sedimentary layers of the pelagic zone (in comparison with acetoclastic methanogenesis), together with the presence of the pool of active psychrotolerant hydrogenotrophic methanogens and acetate-consuming sulfate-reducing bacteria in the communities (Bukin et al., 2018). It should be noted that predominant formation of methane during CO2 reduction occurs also in the sediments of cold-water Antarctic (Wand et al., 2006) and alpine lakes (Mandic-Mulec et al., 2012), as well as in the lakes of the Tibetan Plateau (Liu et al., 2017).
Apart from H2 : CO2, methylated substrates can also play an essential role in methane formation in the sediments of Lake Baikal. During the cultivation of the native sediment from the surface layer under anaerobic conditions in the medium containing methanol, active methane generation from methanol, which involved methylotrophic archaea of the family Methanosarcinaceae, was observed (Bukin et al., 2018).
Using the methods of immunofluorescence staining (Namsaraev and Zemskaya, 2000), cultivation (Pavlova et al., 2014; Bukin et al., 2018), and the 16S rRNA and mcrA gene sequencing (Shubenkova et al., 2005; Kadnikov et al., 2012; Lomakina et al., 2014, 2018; Chernitsyna et al., 2016), it was found that lake sediments contained archaea capable of generating methane via all four catabolic pathways known: hydrogenotrophic (Methanoregulaceae, Methanobacteriaceae, Methanocellaceae, Methanocaldococcaceae, and “Ca. Methanoflorentaceae”), acetoclastic (Methanosaetaceae), methylotrophic (Methanosarcinaseae), and hydrogen-dependent methylotrophic (Methanomassiliicoccales, Methanomethyliaceae, and Methanofastidiosaceae). According to the data on metabarcoding, the largest proportion of detected methanogenic microorganisms in the sediments of Lake Baikal belonged to the hydrogenotrophic representatives of the family Methanoregulaceae (Methanoregula, Methanosphaerula, and uncultured genera), hydrogen-dependent methylotrophic archaea of the order Methanomassiliicoccales, and acetoclastic methanogens of the family Methanosaetaceae (genus Methanothrix (Methanosaeta)). These microorganisms compose the core of methanogenic communities in the sediments of freshwater lakes around the world, regardless of their limnological and hydrochemical characteristics (Wen et al., 2017). In Baikal, members of these taxa were found in the zones of methane seeps and mud volcanoes (Zemskaya et al., 2010; Kadnikov et al., 2012; Chernitsyna et al., 2016; Lomakina et al., 2018), as well as in the sediments of the reference regions (Zemskaya et al., 2018; Zakharova et al., 2018). A large proportion of archaea of the families Methanoregulaceae and Methanosaetaceae may be associated with their high competitiveness under conditions of low substrate concentrations, as well as with their ability to develop stable syntrophic consortia decomposing volatile fatty acids (Borrel et al., 2011). Microorganisms of the order Methanomassiliicoccales (Thermoplasmata) found in the Baikal communities of both the surface and deep layers utilize a wide range of methylated substrates, including methanol, methylamines, and methyl sulfides, the concentrations of which in freshwater ecosystems were previously considered insignificant (Conrad and Claus, 2005). Methylotrophic methanogens are commonly found in the digestive tract of insects and animals, as well as in hypersaline water bodies. In the latter, methylamines, which are formed as a result of the decomposition of osmoprotectants secreted by halophilic proteobacteria, are the main methylated substrates for methylotrophic methanogens (Liu and Whitman, 2008). In the sediments of freshwater ecosystems, methanol that is formed during the degradation of pectin and lignocellulose that compose the cell walls of algae and plants can be the main substrate for them (Sollinger and Urich, 2019). However, although molecular markers of the members of this family were widespread in the communities of the Lake Baikal sediments, they were not detected in methanogenic enrichment cultures when cultivated in the medium containing methanol (Bukin et al., 2018). The inconsistency of the cultivation conditions may be one of the reasons, while another one is the possible use of other C1-methylated compounds instead of methanol by Methanomassiliicoccales in Lake Baikal. Their presence in the upper layers of sediments in the discharge zones of Lake Baikal can be a result of migration of these compounds from deep sediments with the flows of deep highly mineralized fluids. This is in agreement with the isotopic characteristics of dissolved biogenic methane (δ13C1 ≈ –41.0 to –67.3‰; δDC1 = –311 to –300‰), which are characteristic of this gas formed mainly via reduction of the methyl groups (Kalmychkov et al., 2006; Hachikubo et al., 2010). Considering the low rates of acetoclastic methanogenesis in the deep-water zone of the lake, methane generation using methyl groups of C1-methylated compounds is quite probable.
MICROORGANISMS INVOLVED IN ANAEROBIC OXIDATION OF METHANE
The most obvious difference between biogeochemical processes at the sites of seepage of mineralized gas-bearing fluids in Lake Baikal and the processes in oceanic areas is associated with the content of sulfate ions. In the pore water of most regions studied by us, the concentration of sulfate ions was insufficient for the sulfate-dependent AOM (Pimenov et al., 2014). In this regard, the areas of seepage of sulfate- and sulfide-containing abyssal fluids were exceptions. These were Kukuy Canyon and the Malenky mud volcano, in which the sulfate ion content reached 15 mM. The sulfate reduction process was recorded in the sediments of different areas of Lake Baikal directly below the layer of oxidized sediments, although its role in the OM destruction was secondary (Namsaraev and Zemskaya, 2000; Pimenov et al., 2014), which was confirmed by a comparison of the integral rates of methane oxidation and sulfate reduction. Despite insignificant concentrations of sulfate ion (from 0.05 to 0.16 mM), in most sediments in Lake Baikal, analyzed profiles of methane concentrations indicated the occurrence of the AOM process. Comparison of the integral rates of aerobic and anaerobic methane oxidation in the Lake Baikal sediments showed that their values were comparable, while in some areas (the Posolsk Bank methane seep, the Gorevoy Utes (GU) oil seep, and the Bolshoy mud volcano), AOM was significantly higher than the rate of methane oxidation (MO) in the oxic zone. However, no methane-oxidizing archaea of clusters ANME-1, 2, and 3, which are typical of marine sediments, were found in the studied sediments (Zemskaya et al., 2010; Chernitsyna et al., 2016; Lomakina et al., 2018). Possibly, AOM was carried out in the sediments of Lake Baikal by members of other taxa, in particular, archaea of the ANME-2d subcluster and bacteria NC10 (Raghoebarsing et al., 2006; Haroon et al., 2013). Representatives of the subcluster ANME-2d were originally found together with members of the bacterial phylum NC10: “Ca. Methylomirabilis oxyfera” in enrichment cultures obtained from freshwater sediments (Raghoebarsing et al., 2006). These unique bacteria are considered the first anaerobic methanotrophs that are able to produce oxygen from nitrite intracellularly and use it to activate methane, similar to aerobic methanotrophs (Ettwig et al., 2010). Archaea of the ANME-2d subcluster and bacteria of the phylum NC10 are widespread in freshwater ecosystems, soils, rice fields, and wastewater treatment facilities (Raghoebarsing et al., 2006; Haroon et al., 2013; Fu et al., 2016; Welte et al., 2016; Timmers et al., 2017). Previously, their participation in the AOM process was also experimentally confirmed during the cultivation of freshwater sediments (Lakes Chaohu and Taihu, China) and the samples from wastewater reuse lagoons (Luggage Point, Brisbane, Australia) on the medium supplemented with nitrate and nitrite ions (Hu et al., 2009; Ettwig et al., 2010; Haroon et al., 2013). We confirmed that the AOM process via the nitrate-dependent pathway occurred in the sediments of South Baikal by cultivation of the natural microbial community from the Peschanka mud volcano in the medium supplemented with nitrate ions (Lomakina et al., 2018). The experimental samples were characterized by a decrease in methane and the formation of microbial consortia composed of bacteria of the phylum NC10 and archaea of the ANME-2d subcluster, which were identified using fluorescent in situ hybridization with specific probes. Phylogenetic analysis of the 16S rRNA gene libraries showed the similarity of the Baikal archaea of subcluster ANME-2d to the sequences from freshwater sediments of an alpine lake, sediments of Lake Monun, and bioreactors, where AOM processes were also recorded. Members of the phylum NC10 are other participants in the AOM process in the sediments of Lake Baikal. Sequences of this phylum were identified in the microbial communities of all studied sediments, including the Malenky and Kedr mud volcanoes, as well as the Posolsk Bank and Krasny Yar methane seeps. Phylogenetically, members of this taxon were not completely identical; they were grouped into four OTUs and showed similarity to the sequences from the sediments of the lakes Constance and Biwa, as well as to “Ca. Methylomirabilis sp.” from enrichment cultures involved in nitrite-dependent AOM. Anammox bacteria, as well as aerobic ammonium- and nitrite-oxidizing bacteria, are known to participate in the chains of trophic interactions with nitrate- and nitrite-dependent microorganisms carrying out AOM (Welte et al., 2016). As a result of syntrophic interaction in the presence of ammonium in the environment and anammox bacteria or bacteria of the phylum NC10 and nitrite formed in the process of nitrate-dependent AOM, the latter is reduced to gaseous nitrogen (Zhu et al., 2010; Haroon et al., 2013).
It should be noted that the AOM mechanism and the number of its participants in the sediments of Lake Baikal are not fully understood. Even in the areas of discharge of mineralized fluids, the content of nitrate ions in pore water did not exceed 0.0096 mM, and nitrite ions were not detected by the methods used (Zemskaya et al., 2015a; Pogodaeva et al., 2020). It can be assumed that the activity of ammonium-oxidizing archaea of the phylum Thaumarchaeota, which are able to oxidize ammonium to nitrate, compensate for the lack of concentrations of nitrate ions (Zhu et al., 2010). In the sediments of Lake Baikal, members of this phylum indeed composed a significant proportion (up to 26%) in the deep sediments of the areas of gaseous fluid discharge and up to 100% in the deep sediments of the Gorevoy Utes oil seep (Kadnikov et al., 2012; Chernitsyna et al., 2016; Lomakina et al., 2018). The possible involvement of microorganisms in the processes of nitrate- and nitrite-dependent AOM in the sediments of Lake Baikal was confirmed by the presence of marker genes in the total DNA: the mcrA gene for the archaeal subcluster ANME-2d and the pmoA gene for the phylum NC10. The amino acid sequences of the mcrA gene from different areas of Lake Baikal were identical to the sequences of “Ca. Methanoperedens sp.” from the sediments of wastewater treatment plants and bioreactors, while amino acid sequences of the pmoA gene were identical to those of uncultured NC10 members from the sediments of the Lakes Biwa and Constance, as well as from denitrifying reservoirs.
Members of the phylum Bathyarchaeota should not be excluded as other possible participants in the methane cycle in the Lake Baikal sediments. Recent metagenomic studies of two genomes of the phylum Ba-thyarchaeota (formerly Miscellaneous Crenarchaeota Group) showed the presence of the genes encoding the methyl-coenzyme M reductase (MCR) complex, including the mcrA gene (Evans et al., 2015). The authors did not exclude the involvement of Bathyarchaeota not only in the process of methane generation, but also in the process of AOM via the mechanism of reverse methanogenesis. The metabolism of archaea of this phylum in nature, including Lake Baikal sediments, remains to be determined. According to the results of high-throughput sequencing, members of this phylum in the Lake Baikal sediments were responsible for 28–76% of all analyzed archaeal 16S rRNA gene sequences in the areas of mud volcanoes and methane seeps. Moreover, enzymes of methyl-reducing methanogenesis were found in members of the phylum Verstraetearchaeota, which indicates their involvement in methane metabolism (Vanwonterghem et al., 2016). Sequences of the Verstraetearchaeota members, which proved to harbor the mcrA genes, were isolated from a mesophilic methanogenic reactor and from the sediments of the freshwater Lake Pavin (France). However, in the sediments of Lake Baikal, the sequences of this phylum were minor components in the studied microbial communities.
In the publications (Och et al., 2012; Torres et al., 2014; Aloisi et al., 2019), the involvement of Mn(IV) and Fe(III) oxides in the AOM process leading to bicarbonate formation in the sediments of Lake Baikal was considered. Elevated concentrations of iron ions in pore water with the \({\text{SO}}_{4}^{{2 - }}\) and \({\text{NO}}_{3}^{ - }\) contents that are insufficient for the occurrence of AOM were observed in the surface sediments (first centimeters) in different areas of Lake Baikal (Granina et al., 2004; Och et al., 2012; Zemskaya et al., 2015a). This process was previously observed in the sediments of Lake Ørn, where members of ANME-2d were detected in the zones with a low content of nitrates and sulfates in the presence of Fe ions (Norgi et al., 2013; Weber et al., 2017). The microorganisms that are responsible for this process have not been reliably identified, although members of ANME-2d may also participate in the process of iron-dependent AOM (Cai et al., 2018). Therefore, members of the ANME-2d subcluster are presently considered universal anaerobic methanotrophs, which use various ions as electron acceptors in AOM and, thus, play an essential role in methane emission (Shen et al., 2019).
In the oceanic sediments of cold seeps, increased flows of \({\text{HCO}}_{3}^{ - },\) which result from the sulfate-dependent AOM and lead to the widespread deposition of authigenic carbonates in the form of nodules in sediments and carbonate crusts, have been observed (Bohrmann et al., 1998; Luff et al., 2004; Aloisi et al., 2019). In contrast to marine ecosystems, regions of formation of authigenic carbonates are not present throughout Baikal but are locally distributed. In these areas, small (up to 0.5 cm) inclusions of authigenic siderite and rhodochrosite are found (Mizandrontsev, 1975; Granina, 2008; Krylov et al., 2018), which may be a consequence of the low concentration of \({\text{HCO}}_{3}^{ - }\) ions in such zones. The absence of authigenic carbonates over a large area of sediments suggests that the rates of these processes are not high enough to significantly shift the equilibrium of dissolved inorganic carbon in favor of precipitation of the carbonate minerals. Finally, the limited deposition of carbonate minerals in the sediments of cold seeps in Lake Baikal is also associated with the concentration of dissolved calcium, which is 20 times lower than in the ocean. The formation of authigenic carbonates requires increased values of carbonate alkalinity in pore water/fluids, as well as occurrence of the AOM or methane generation processes. In the Baikal sediments containing authigenic carbonates, an increase in alkalinity was noted. The concentration of bicarbonate ions in the pore water was more than 8.19 mM, which is significantly higher than the values recorded for the sediments of the reference regions (1.09 mM) (Zemskaya et al., 2015a; Pogodaeva et al., 2017). Analysis of the 16S rRNA gene libraries from carbonate-containing sediments showed the presence of the sequences of archaea from the ANME-2d subcluster and members of other phyla. For instance, Euryarchaeota (up to 99%) predominated in archaeal communities from oligonite (Fe(Mn, Zn)(CO3)2) collected from the sediments of the Malenky mud volcano, among which 20% of the sequences belonged to the subcluster ANME-2d and 73% were the sequences of the order Thermoplasmatales (Lomakina et al., 2018). In the area where carbonate was identified as siderite (FeCO3), sequences identified as Bathyarchaeota prevailed (from 33 to 76.6%); the proportion of the members of subcluster ANME-2d was approximately 16% (Lomakina et al., 2020). A high percentage of members of the bacterial phyla Chloroflexi and Atribacteria, which are typical members of marine carbonate sediments (Yanagawa et al., 2019), should also be noted in the bacterial communities from the same samples. The data on the taxonomic composition of the communities make it impossible to unravel the mechanism of carbonate formation in the Lake Baikal sediments but can be used for further research in this field.
MICROORGANISMS PARTICIPATING IN THE OXIDATION OF OIL HYDROCARBONS
Apart from its age, depth, and high diversity of biota, natural oil seeps are also a unique feature of Lake Baikal. In addition to Lake Baikal, this phenomenon is characteristic of the deep-water rift Lake Tanganyika and a shallow-water Lake Chapala (Simoneit et al., 2000; Zarate-del Valle et al., 2006). Lake Baikal is known for two areas of natural oil seeps. One area is located at the mouth of the River Bolshaya Zelenovskaya (BZ) and has been known since the end of the 18th century; the second one is situated near Cape Gorevoy Utes (GU) (discovered in 2005). In 2005, oil in the GU area was characterized by an extremely high content of n-alkanes and was identified as non-biodegraded paraffinic oil; in the BZ area, it was described as biodegraded aromatic-naphthenic oil (Kontorovich et al., 2007). Microbiological studies of the oil seeps in the area of the BZ river mouth were started in the 1980s by Taliev et al. (1985). In the water and sediments, the distribution of hydrocarbon-oxidizing microorganisms capable of assimilating hydrocarbons of diesel oil, methane oil, and individual hydrocarbons С3Н8, С4Н10, С5Н12, and С9Н20 was studied (Petrova and Mamontova, 1985; Taliev et al., 1985).
The discovery of a new area of natural hydrocarbon seeps in 2005 allowed online monitoring of the phenomena occurring in this basin, as well as in situ monitoring of qualitative and quantitative changes in the oil composition, structure of microbial communities, and their adaptive capabilities. In the first years of research, an increase in the abundance of hydrocarbon-oxidizing microorganisms (HCOMs) was noted in both the water column and sediments. In the zone of oil slicks formed on the surface of the water column, the number of cultured HCOMs was 12 times higher than the values in the water of the reference regions. Monitoring the abundance of cultured microorganisms that oxidize oil and n-alkanes in the sediments of GU for 15 years has shown its increase in the first two years (2005–2007) (up to 25 ± 2.7 × 104 CFU/g) and a subsequent decrease in 2017–2019 to 9.0 ± 0.1 × 104 CFU/g (Pavlova et al., 2020). An increase in the number of hydrocarbon-oxidizing bacteria after a technogenic oil spill was reported in numerous studies (King et al., 2015). For instance, a month after the accident at the Deepwater Horizon oil rig, the total number of microorganisms was 5.51 ± 0.33 × 104 cells/mL in the deep-water oil plume in the Gulf of Mexico and 2.73 ± 0.33 × 104 cells/mL outside it (Hazen et al., 2010). High indices of the abundance of cultured HCOMs (up to 17.0 ± 1.3 × 104 CFU/g) were recorded in bituminous structures discovered in the area of GU using inhabited Mir deep-submergence vehicles in 2008. These structures are formed of paraffinic oil bitumen and inhabited by a large number of diverse benthic organisms. Their food chain is based on the microorganisms that carry out aerobic and anaerobic hydrocarbon oxidation (Zemskaya et al., 2012; Kadnikov et al., 2013; Sitnikova et al., 2017). Eighty percent of the microbial community colonizing bituminous structures consisted of the members of the phylum Proteobacteria, 40% of which belonged to the uncultured members of Alpha- and Betaproteobacteria. Archaea were represented by typical acetoclastic and hydrogenotrophic methanogens of the orders Methanosarcinales, Methanomicrobiales, and Methanobacteriales (Kadnikov et al., 2013).
High-throughput sequencing analysis of the microbial community structure in the sediments from the GU and BZ areas of oil seeps revealed predominance of the members of the same phyla as in the areas with normal sedimentation: Actinobacteria, Cyanobacteria, and Proteobacteria (Zemskaya et al., 2018). The composition of bacterial communities in the BZ and GU sediments was identical at the level of higher taxa, apart from members of the phylum Actinobacteria, which were more diverse in the GU sediments (Zemskaya et al., 2015a). Among Proteobacteria, members of the classes Alpha-, Beta-, Gamma-, and Deltaproteobacteria prevailed; their presence is characteristic of methanogenic hydrocarbon-oxidizing communities, for instance, in methanogenic oil tailings and hydrocarbon-polluted aquatic ecosystems and sediments (Johnson et al., 2015).
The most obvious differences were observed in the composition of archaeal communities. Members of Thaumarchaeota and Euryarchaeota predominated in the GU sediment communities, which is typical of the Baikal sediments of the methane seep and mud volcano areas. In the oxidized layers of the BZ sediment, members of Crenarchaeota (47%) and Thaumarchaeota (53%) were identified among archaea. In reduced layers, identified sequences belonged to the phyla Eu-ryarchaeota (52%), Thaumarchaeota (26%), Bathyarchaeota (20%), and Crenarchaeota (2%). The phylum Euryarchaeota was represented by hydrogenotrophic archaea of the order Methanomicrobiales and acetoclastic methanogens of the order Methanosarcinales, as well as the sequences of the ANME-2d subcluster (Lomakina et al., 2018). A higher diversity of archaea in the BZ area sediments is most likely associated with a rather long period of their coexistence with oil, which serves as an additional carbon source. This has been previously shown for marine ecosystems in which persistent oil pollution results in a higher microbial diversity due to the induced resistance caused by long-term exposure to hydrocarbons (Jeanbille et al., 2016). It should be noted that members of the phyla Bathyarchaeota and Crenarchaeota were present only in the communities of the BZ area in which oil was highly biodegraded. Members of the phylum Bathyarchaeota may be involved in the AOM process (Evans et al., 2015) and the degradation of aromatic compounds (Jeanbille et al., 2016). The phylum Crenarchaeota in the sediments of the BZ oil seeps was represented by uncultured members of the order Desulfurococcales (class Thermoprotei). Under autotrophic conditions, these microorganisms obtain energy by oxidizing hydrogen using elemental sulfur, thiosulfate, nitrate, or nitrite as electron acceptors and CO2 as a carbon source (Huber and Stetter, 2006). Predominance of members of the phyla Thaumarchaeota and Euryarchaeota in the sediments of the GU oil seeps may be due to a more diverse composition of the oil, which contains n-alkanes, alkylcyclohexanes, isoprenoids, PAHs, and other compounds, as well as to the presence of methane. Members of methanogenic archaea of the families Methanomicrobiaceae, Methanosarcinaceae, “Ca. Methanofastidiosa”, and the order Thermoplasmatales were shown to be capable of generating methane and its anaerobic oxidation, as well as of oil degradation in oil-contaminated soils (Miettinen et al., 2019). Members of the phylum Thaumarchaeota, which are supposed to have a heterotrophic type of metabolism using compounds obtained from crude oil, may also participate in oil biodegradation (Mussmann et al., 2011).
Monitoring the composition of Baikal oil in the area of GU oil seeps for 15 years has indicated oil biodegradation (Gorshkov et al., 2020). Aerobic processes of oil hydrocarbon oxidation prevail in the oxygen-saturated water column and surface sediments with high O2 content. Studies of the microorganisms involved in oil degradation in the anaerobic zone of the sedimentary layers of Lake Baikal are still at the initial stage. The first experimental data on the ability of microbial communities from GU sediments to degrade oil hydrocarbons under anaerobic conditions in the presence of various electron acceptors were obtained. In surface sediments, the conversion of n-alkanes proceeded most intensively in enrichment cultures in the medium supplemented with sulfate ions, while in deep sediments, it occurred most efficiently on a medium with bicarbonate ions. In deep sediments, the microbial community is more focused on anaerobic oxidation of polycyclic aromatic hydrocarbons (according to the high level of their biodegradation, up to 46%), regardless of the electron acceptors present in the medium. Analysis of the clonal libraries of the 16S rRNA genes of bacteria and archaea from enrichment cultures showed the identity (97–99%) of the sequences to those of uncultured microorganisms from thermophilic habitats, sediments of mud volcanoes, and environments polluted with hydrocarbons. The 16S rRNA gene libraries of bacteria were dominated by the members of the phyla Firmicutes, Chloroflexi, Caldiserica (OP5), classes Delta- and Epsilonproteobacteria (Pavlova et al., 2020). The phylogenetic diversity of anaerobic microorganisms involved in oil biodegradation is similar in its composition to that of the microorganisms of the “microbiome core” of oil reservoirs, in which three classes of bacteria (Gammaproteobacteria, Clostridia, and Bacteroidia) and archaea of the class Methanomicrobia were most represented. The functioning of communities in oil reservoirs is provided by syntrophic interaction of these microorganisms (Sierra-Garcia et al., 2017; Pannekens et al., 2019). We do not rule out syntrophic relationships in the microbial communities of Lake Baikal sediments, since microorganisms responsible for interdependent sequential reactions in the general metabolic process were found in all experimental samples.
The ability to degrade hydrocarbons is provided by the genomes of Baikal microorganisms (Lomakina et al., 2014; Likhoshvay et al., 2014). Analysis of the nucleotide sequences of the alk genes encoding alkane-1-monooxygenase showed that most of the cultured hydrocarbon-oxidizing microorganisms (76%) isolated from the water column and sediments in the areas of natural oil seeps in Lake Baikal contained the alkB genes, which are mostly identical to the alkB genes previously identified in Rhodococcus erythropolis (Lomakina et al., 2014). The presence of the alk genes was confirmed by metagenomic analysis of the genomes of microorganisms inhabiting the bathypelagic zone of South Baikal (Cabello-Yeves et al., 2020). A particularly active strain degrading n-alkanes was the strain Rhodococcus erythropolis 4–08 isolated from a bituminous structure of Lake Baikal. The genome of this strain was shown to harbor four types of the alkB genes encoding alkane-1-monooxygenase (Likhoshvay et al., 2013, 2014). The growth rate of Rhodococcus erythropolis increased 2–16 times when cultured on the medium supplemented with a growth stimulant, which makes it promising for biotechnological purposes (Pavlova et al., 2019a, 2019b).
CONCLUSIONS
The sediments of Lake Baikal are inhabited by microorganisms with diverse types of metabolism providing for their participation in the degradation of a wide range of substrates. The latter include photosynthesized OM, as well as gaseous and oil hydrocarbons. The most significant changes in the community composition and the succession of the predominant microbial taxa were observed in the sediments with a pronounced gradient of physicochemical parameters. However, the taxa of bacteria and archaea characteristic of freshwater ecosystems were revealed also in these ecotopes (Newton et al., 2011). Low mineralization of the environment, as well as the availability of carbon and energy sources, are necessary for the growth of microorganisms involved in both the formation of methane and its anaerobic oxidation via the metabolic pathways that are characteristic of the microorganisms from freshwater ecosystems. The diversity of microbial communities in the Lake Baikal sediments overlaps at the phylum level and demonstrates an increasing dissimilarity of microorganisms below the class and genus levels. In this regard, a broader metagenomic analysis of genomes and studies of the metabolism of unique microbial species, including those from the deep zone of Lake Baikal sediments, are required. The ancient age of Lake Baikal, its thick sedimentary layer, and the presence of methane of various origins (including gas hydrates) provide the basis for further studies on the role of microorganisms participating in the cycle of this greenhouse gas, especially during the period of global climate change.
REFERENCES
Aloisi, G., Pogodaeva, T.V., Poort, J., Khabuev, A.V., Kazakov, A.V., Akhmanov, G.G., and Khlystov, O.M., Biogeochemical processes at the Krasniy Yar seepage area (Lake Baikal) and a comparison with oceanic seeps, Geo-Mar. Lett., 2019, vol. 39, pp. 59–75.
Beal, E.J., House, C.H., and Orphan, V.J., Manganese- and iron-dependent marine methane oxidation, Science, 2009, vol. 325, pp. 184–187.
Bohrmann, G., Greinert, J., Suess, E., and Torres, M., Au-thigenic carbonates from the Cascadia subduction zone and their relation to gas hydrate stability, Geology, 1998, vol. 26, p. 647.
Borrel, G., Jézéquel, D., Biderre-Petit, C., Morel-Desrosiers, N., Morel, J.-P., Peyret, P., Fonty, G., and Lehours, A.-C., Production and consumption of methane in freshwater lake ecosystems, Res. Microbiol., 2011, vol. 162, pp. 832–847.
Borrel, G., Lehours, A.C., Crouzet, O., Jézéquel, D., Rockne, K. Kulczak, A., Duffaud, E., Joblin, K., and Fonty, G., Stratification of Archaea in the deep sediments of a freshwater meromictic lake: vertical shift from methanogenic to uncultured archaeal lineages, PLoS One, 2012, vol. 7. e43346. https://doi.org/10.1371/journal.pone.0043346
Bukin, S.V., Pavlova, O.N., Kalmychkov, G.V., Iva-nov, V.G., Pogodaeva, T.V., Galachyants, Yu.P., Bukin, Yu.S., Khabuev, A.V., and Zemskaya, T.I., Substrate specificity of methanogenic communities from Lake Baikal bottom sediments associated with hydrocarbon gas discharge, Microbiology (Moscow), 2018, vol. 87, pp. 549–558.
Bukin, S.V., Pavlova, O.N., Manakov, A.Y., Kostreva, E.A., Chernitsyna, S.M., Mamaeva, E.V., Pogodaeva, T.V., and Zemskaya, T.I., The ability of microbial community of Lake Baikal bottom sediments associated with gas discharge to carry out the transformation of organic matter under thermobaric conditions, Front. Microbiol., 2016, vol. 7, art. 690.
Cabello-Yeves, P.J., Zemskaya, T.I., Zakharenko, A.S., Sakirko, M.V., Ivanov, V.G., Ghai, R., and Rodriguez-Valera, F., Microbiome of the deep Lake Baikal, a unique oxic bathypelagic habitat, Limnol. Oceanogr., 2020, vol. 65, pp. 1471–1488.
Cai, C., Leu, A.O., Xie, G.J., Guo, J., Feng, Y., Zhao, J.X., Tyson, G.W., Yuan, Z., and Hu, S., A methanotrophic archaeon couples anaerobic oxidation of methane to Fe(III) reduction, ISME J., 2018, vol. 12, pp. 1929–1939.
Capone, D.G. and Kiene, R.P., Comparison of microbial dynamics in marine and freshwater sediments: Contrasts in anaerobic carbon catabolism, Limnol. Oceanogr., 1988, vol. 33, pp. 725–749.
Chernitsyna, S.M., Mamaeva, E.V., Lomakina, A.V., Pogodaeva, T.V., Galach’yants, Yu.P., Bukin, S.V., Pimenov, N.V., Khlystov, O.M., and Zemskaya, T.I., Phylogenetic diversity of microbial communities of the Posolsk Bank bottom sediments, Lake Baikal, Microbiology (Moscow), 2016, vol. 85, pp. 672–680.
Conrad, R. and Claus, P., Contribution of methanol to the production of methane and its 13C-isotopic signature in anoxic rice field soil, Biogeochem., 2005, vol. 73, pp. 381–393.
Conrad, R., Chan, O.-C., Claus, P., and Casper, P., Characterization of methanogenic Archaea and stable isotope fractionation during methane production in the profundal sediment of an oligotrophic lake (Lake Stechlin, Germany), Limnol. Oceanogr., 2007, vol. 52, pp. 1393–1406.
Dagurova, O.P., Namsaraev, B.B., Kozyreva, L.P., Zemskaya, T.I., and Dulov, L.E., Bacterial processes of the methane cycle in bottom sediments of Lake Baikal, Microbiology (Moscow), 2004, vol. 74, pp. 202–210.
Dedysh, S.N., Derakshani, M., and Liesack, W., Detection and enumeration of methanotrophs in acidic Sphagnum peat by 16S rRNA fluorescence in situ hybridization, including the use of newly developed oligonucleotide probes for Methylocella palustris, Appl. Environ. Microbiol., 2001, vol. 67, pp. 4850–4857.
Ding, H. and Valentine, D., Methanotrophic bacteria occupy benthic microbial mats in shallow marine hydrocarbon seeps, Coal Oil Point, California, J. Geophys. Res., 2008, vol. 113, G-1. https://doi.org/10.1029/2007jg000537
Duc, N.T., Crill, P., and Bastviken, D., Implications of temperature and sediment characteristics on methane formation and oxidation in lake sediments, Biogeochem., 2010, vol. 100, pp. 185–196.
Ettwig, K.F., Butler, M.K., Le Paslier, D., Pelletier, E., Mangenot, S., Kuypers, M.M., Schreiber, F., Dutilh, B.E., Zedelius, J., de Beer, D., Gloerich, J., Wessels, H.J., van Alen, T., Luesken, F., Wu M.L., et al., Nitrite-driven anaerobic methane oxidation by oxygenic bacteria, Nature, 2010, vol. 464, pp. 543–548.
Evans, P.N., Parks, D.H., Chadwick, G.L., Robbins, S.J., Orphan, V.J., Golding, S.D., and Tyson, G.W., Methane metabolism in the archaeal phylum Bathyarchaeota revealed by genome-centric metagenomics, Science, 2015, vol. 350, pp. 434–438.
Fuchs, A., Lyautey, E., Montuelle, B., and Casper, P., Effects of increasing temperatures on methane concentrations and methanogenesis during experimental incubation of sediments from oligotrophic and mesotrophic lakes, J. Geophys. Res. Biogeosci., 2016, vol. 121, pp. 1394–1406.
Fu, L., Li, S.W., Ding, Z.W., Ding, J., Lu, Y.Z., and Zeng, R.J., Iron reduction in the DAMO/Shewanella oneidensis MR-1 coculture system and the fate of Fe(II), Water Res., 2016, vol. 88, pp. 808–815.
Gorshkov, A.G., Pavlova, O.N., Khlystov, O.M., and Zemskaya, T.I., Fractioning of petroleum hydrocarbons from seeped oil as a factor of purity preservation of water in Lake Baikal (Russia), J. Great Lakes Res., 2020, vol. 46, pp. 115–122.
Granina, L., Muller, B., and Wehrli, B., Origin and dynamics of Fe and Mn sedimentary layers in Lake Baikal, Chem. Geol., 2004, vol. 205, pp. 55–72.
Granina, L.Z., Rannii diagenez donnykh osadkov Baikala (Early Diagenesis in Lake Baikal Bottom Sediments, Novosibirsk: GEO, 2008. Gvozdkov, A.N., Geochemistry of the modern Lake Baikal sediments, Extended Abstract Cand. Sc. (Geol.-Min.) Dissertation, Irkutsk, 1998.
Hachikubo, A., Khlystov, O., Krylov, A., Sakagami, H., Minami, H., Nunokawa, Y., Yamashita, S., Takahashi, N., Shoji, H., Nishio, S., Kida, M., Ebinuma, T., Kalmych-kov, G., and Poort, J., Molecular and isotopic characteristics of gas hydrate-bound hydrocarbons in southern and central Lake Baikal, Geo-Mar. Lett., 2010, vol. 30, pp. 321–329.
Han, X., Schubert, C.J., Fiskal, A., Dubois, N., and Lever, M.A., Eutrophication as a driver of microbial community structure in lake sediments, Environ. Microbiol., 2020, vol. 22, pp. 3446–3462.
Haroon, M.F., Hu, S., Shi, Y., Imelfort, M., Keller, J., Hugenholtz, P., Yuan, Z., and Tyson, G.W., Anaerobic oxidation of methane coupled to nitrate reduction in a novel archaeal lineage, Nature, 2013, vol. 500, pp. 567–570.
Hazen, T.C., Dubinsky, E.A., DeSantis, T.Z., Andersen, G.L., Piceno, Y.M., Singh, N., Jansson, J.K., Probst, A., Borglin, S.E., Fortney, J.L., Stringfellow, W.T., Bill, M., Conrad, M.E., Tom, L.M., Chavarria, K.L., et al., Deep-sea oil plume enriches indigenous oil-degrading bacteria, Science, 2010, vol. 330, pp. 204–208.
Huang, W., Chen, X., Wang, K., Chen, J., Zheng, B., and Jiang, X., Comparison among the microbial communities in the lake, lake wetland, and estuary sediments of a plain river network, Microbiol. Open, 2019, vol. 8. e644. https://doi.org/10.1002/mbo3.644
Huber, H. and Stetter, K.O., Desulfurococcales, in The Prokaryotes, Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., and Stackebrandt, E., Eds., New York: Springer, 2006, pp. 52–68.
Hu, S., Zeng, R.J., Burow, L.C., Lant, P., Keller, J., and Yuan, Z., Enrichment of denitrifying a anaerobic methane oxidizing microorganisms, Environ. Microbiol. Rep., 2009, vol. 1, pp. 377–384.
Hutchinson, D.R., Golmshtok, A.J., Zonenshain, L.P., Moore, T.C., Scholz, C.A., and Klitgord, K.D., Depositional and tectonic flamework of the rift basins of Lake Baikal from multichannel seismic data, Geology, 1992, vol. 20, pp. 589–592.
Jeanbille, M., Gury, J., Duran, R., Tronczynski, J.K, Ghiglione, J.-F., Agogue, H., Said, O.B., Taib, N., Debroas, D., Garnier, C., and Auguet, J.-C., Chronic polyaromatic hydrocarbon (PAH) contamination is a marginal driver for community diversity and prokaryotic predicted functioning in coastal sediments, Front. Microbiol., 2016, vol. 7, art. 1303.
Johnson, J.M., Wawrik, B., Isom, C., Boling, W.B., and Callaghan, A.V., Interrogation of Chesapeake Bay sediment microbial communities for intrinsic alkane-utilizing potential under anaerobic conditions, FEMS Microbiol. Ecol., 2015, vol. 91, no. 2, pp. 1–14.
Kadnikov, V.V., Lomakina, A.V., Likhoshvai, A.V., Gorshkov, A.G., Pogodaeva, T.V., Beletsky, A.V., Mardanov, A.V., Zemskaya, T.I., and Ravin, N.V., Composition of the microbial communities of bituminous constructions at natural oil seeps at the bottom of Lake Baikal, Microbiology (Moscow), 2013, vol. 82, pp. 373–382.
Kadnikov, V.V., Mardanov, A., Beletsky, A.V, Shubenkova, O.V., Pogodaeva, T.N., Zemskaya, T.I., Ravin, N.V., and Skryabin, K.G., Microbial community structure in methane hydrate-bearing sediments of freshwater Lake Baikal, FEMS Microbiol Ecol., 2012, vol. 79, no. 1, pp. 348–358.
Kallistova, A.Y., Kevbrina, M.V., Pimenov, N.V., Rusanov, I.I., Rogozin, D.Y., Wehrli, B., and Nozhevnikova, A.N., Sulfate reduction and methanogenesis in the Shira and Shunet meromictic lakes (Khakasia, Russia), Microbiology (Moscow), 2006, vol. 75, pp. 720–726.
Kalmychkov, G.V., Egorov, A.V., Kuz’min, M.I., and Kh-lystov, O.M., Genetic types of methane from Lake Baikal, Dokl. Earth Sci., 2006, vol. 411, pp. 1462–1465.
King, G.M., Kostka, J.E., Hazen, T.C., and Sobecky, P.A., Microbial responses to the Deepwater Horizon Oil Spill: from coastal wetlands to the deep sea, Annu. Rev. Mar. Sci., 2015, vol. 7, pp. 377–401.
Klerkx, J., Zemskaya, T.I., Matveeva, T.V., Khlystov, O.M., Namsaraev, B.B., Dagurova, O.P., Golobokova, L.P., Vorobyeva, S.S., Pogodaeva, T.P., Granin, N.G., Kalmychkov, G.V., Ponomarchuk, V.A., Shoji, H., Mazurenko, L.L., Kaulio, V.V., et al., Methane hydrates in surface layer of deep-water sediments in Lake Baikal, Dokl. Earth Sci., 2003, vol. 393, pp. 822–826.
Koizumi, Y., Takii, S., Nishino, M., and Nakajima, T., Vertical distributions of sulfate-reducing bacteria and methane-producing archaea quantified by oligonucleotide probe hybridization in the profundal sediment of a mesotrophic lake, FEMS Microbiol. Ecol., 2003, vol. 44, art. 101e108.
Kolman, S.M., Kuptsov, V.M., Dzoins, G.A., and Karter, S.D., Radiocarbon dating of Lake Baikal sediments, Geol. Geofiz., 1993, vol. 34, nos. 10–11, pp. 68–77.
Kontorovich, A.E., Kashirtsev, V.A., Moskvin, V.I., Burshtein, L.M., Zemskaya, T.I., Kostyreva, E.A., Kalmychkov, G.V., and Khlystov, O.M., Petroleum potential of Baikal deposits, Russ. Geol. Geophys., 2007, vol. 12, pp. 1046–1053.
Kotsyurbenko, O.R., Trophic interactions in the methanogenic microbial community of low-temperature terrestrial ecosystems, FEMS Microbiol. Ecol., 2005, vol. 53, pp. 3–13.
Krylov, A.A., Hachikubo, A., Minami, H., Pogodae-va, T.V., Zemskaya, T.I. Krzhizhanovskaya, M.G., Poort, J., and Khlystov, O.M., Authigenic rhodochrosite from a gas hydrate-bearing structure in Lake Baikal, Int. J. Earth Sci., 2018, vol. 107, pp. 2011–2022.
Kuz’min, M.I., Karananov, E.B., Kavai, T., et al., Deep-water drilling at Lake Baikal–main results, Geol. Geofiz., 2001, vol. 42, pp. 8–34.
Kuznetsov, A.P., Strizhov, V.P., Kuzin, V.S., Fialkov, V.A., and Yastrebov, V.S., News on Baikal nature. A community based on bacterial chemosynthesis, Izv. AM SSSR, Ser. Boil., 1991, no. 5, pp. 766–772.
Kuznetsov, S.I., Mikroflora ozer i ee geokhimicheskaya deyatel’nost’ (Microflora of Lakes and Its Geochemical Activity), Leningrad: Nauka, 1970.
Lever, M.A., Rogers, K.L., Lloyd, K.G., Overmann, J., Schink, B., Thauer, R.K., and Jørgensen, B.B., Life under extreme energy limitation: a synthesis of laboratory- and field-based investigations, FEMS Microbiol. Rev., 2015, vol. 39, pp. 688–728.
Likhoshvay, A., Khanaeva, T., Gorshkov, A., Zemskaya, T., and Grachev, M., Do oil-degrading Rhodococci contribute to the genesis of deep water bitumen mounds in Lake Baikal?, Geomicrobiol. J., 2013, vol. 30, pp. 209–213.
Likhoshvay, A., Lomakina, A., and Grachev, M., The complete alk sequences of Rhodococcus erythropolis from Lake Baikal, Springer Plus, 2014, vol. 3, art. 621.
Liu, Y. and Whitman, W.B., Metabolic, phylogenetic and ecological diversity of the methanogenic Archaea, Ann. N.Y. Acad. Sci., 2008, vol. 1125, pp. 171–189.
Liu, Y., Conrad, R., Yao, T., Gleixner, G., and Claus, P., Change of methane production pathway with sediment depth in a lake on the Tibetan plateau, Palaeogeogr. Palaeoclimatol. Palaeoecol., 2017, vol. 474, pp. 279–286.
Logachev, N.A., History and Geodynamics of the Baikal Rift, Geol. Geofiz., 2003, vol. 44, pp. 391–406.
Lomakina, A., Pogodaeva, T., Kalmychkov, G., Chernitsyna, S., and Zemskaya, T., Diversity of NC10 bacteria and ANME-2d archaea in sediments of fault zones at Lake Baikal, Diversity-Basel, 2020, vol. 12. https://doi.org/10.3390/d12010010
Lomakina, A.V., Mamaeva, E.V., Galachyants, Y.P., Petrova, D.P., Pogodaeva, T.V., Shubenkova, O.V., Khabuev, A.V., Morozov, I.V., and Zemskaya, T.I., Diversity of Archaea in bottom sediments of the discharge areas with oil- and gas-bearing fluids in Lake Baikal, Geomicrobiol. J., 2018, vol. 35, pp. 50–63.
Lomakina, A.V., Mamaeva, E.V., Pogodaeva, T.V., Kalmychkov, G.V., Khalzov, I.A., and Zemskaya, T.I., Anaerobic methane oxidation in enrichment cultures from deep sediments of a mud volcano Peschanka (South Baikal), Microbiology (Moscow), 2018, vol. 87, pp. 317–325.
Lomakina, A.V., Pogodaeva, T.V., Morozov, I.V., and Zemskaya, T.I., Microbial communities of the discharge zone of oil- and gas-bearing fluids in low-mineral Lake Baikal, Microbiology (Moscow), 2014, vol. 83, pp. 278–287.
Luff, R., Wallmann, K., and Aloisi, G., Numerical modeling of carbonate crust formation at cold vent sites: significance for fluid and methane budgets and chemosynthetic biological communities, Earth Planet. Sci. Lett., 2004, vol. 221, pp. 337–353.
Maksimova, E.A. and Maksimov, V.N., Mikrobiologiya vod Baikala (Mikrobiology of Baikal Water), Irkutsk: Irkutsk Gos. Univ., 1989.
Mandic-Mulec, I., Gorenc, K., Petrišiš, M.G., Faganeli, J., and Ogrinc, N., Methanogenesis pathways in a stratified eutrophic alpine lake (Lake Bled, Slovenia), Limnol. Oceanogr., 2012, vol. 57, pp. 868–880.
Mats, V.D., Ufimtsev, G.F., Mandel’baum, M.M., Alakshin, A.M., Pospeev, A.V., Shimaraev, M.N., and Khlystov, O.M., Kainozoi Baikal’skoi riftovoi vpadiny: stroenie i geologicheskaya istoriya (Baikal Rift Cenozoic of the Baikal Rift Depression: Structure and Geological History), Novosibirsk: Geo, 2001.
Miettinen, H., Bomberg, M., Nyyssönen, M., Reunamo, A., Jørgensen, K.S., and Vikman, M., Oil degradation potential of microbial communities in water and sediment of Baltic Sea coastal area, PLoS One, 2019, vol. 17. e0218834. https://doi.org/10.1371/journal.pone.0218834
Mikrobiologicheskoe nasledie XX veka (Microbiological Heritage of the 20th Century), Vinogradova, T.P. Ed., Irkutsk: Inst. Geogr. SO RAN, 2004.
Mikroorganizmy v ekosistemakh ozer i vodokhranilishch (Microorganisms in the Ecosystems of Lakes and Reservoirs), Dryukker, V.V., Ed., Novosibirsk: Nauka, 1985.
Minami, H., Pogodaeva, T., Sakagami, H., Hachikubo, A., Krylov, A., Harada, D., Saito, C., Tatsumi, K., Hyakuta-ke, K., Yamashita, S., Nishio, S., Takahashi, N., Shoji, H., Khlystov, O., Zemskaya, T., et al., Traces of original gas hydrate-forming fluid observed in subsurface gas hydrates retrieved from Lake Baikal, Russia, 10th Int. Conf. on Gas in Marine Sediments, Listvyanka, Russia, 2010, p. 129.
Mizandrontsev, I.B., K geokhimii gruntovykh rastvorov (On the Geochemistry of Soil Solutions), Tr. LIN SO AN SSSR, 1975, vol. 21, no. 41, pp. 203–230.
Mizandrontsev, I.B., Osadkoobrazovanie (Sediment Formation), Tr. LIN SO AN SSSR, 1978, vol. 16, no. 36, pp. 33–46.
Mussmann, M., Brito, I., Pitcher, A., Sinninghe Damste, J.S., Hatzenpichler, R., Richter, A., Nielsen, J.L., Nielsen, P.H., Muller, A., Daims, H., Wagner, M., and Head, I.M., Thaumarchaeotes abundant in refinery nitrifying sludges express amoA but are not obligate autotrophic ammonia oxidizers, Proc. Natl. Acad. Sci. U. S. A., 2011, vol. 108, pp. 16771–16776.
Nagata, T., Takai, K., Kawanobe, K., Kim, D.-S., Nakazato, R., Guselnikova, N., Bondarenko, N., Mologawaya, O., Kostornova, T., Drucker, V., Satoh, Y., and Watanabe, Y., Autotrophic picoplankton in southern Lake Baikal: abundance, growth and grazing mortality during summer, J. Plankton Res., 1994, vol. 16, pp. 945–959.
Namsaraev, B.B. and Zemskaya, T.I., Mikrobiologicheskie protsessy krugovorota ugleroda v donnykh osadkakh ozera Baikal (Microbial Processes of the Carbon Cycle in Lake Baikal Bottom Sediments), Novosibirsk: Geo, 2000.
Newton, R.J., Jones, S.E., Eiler, A., McMahon, K.D., and Bertilsson, S., A guide to the natural history of freshwater lake bacteria, Microbiol. Mol. Biol. R., 2001, vol. 75, pp. 14–49.
Norgi, K.A., Thamdrup, B., and Schubert, C.J., Anaerobic oxidation of methane in an iron-rich Danish freshwater lake sediment, Limnol. Oceanogr., 2013, vol. 58, pp. 546–554.
Nozhevnikova, A.N., Nekrasova, V., Ammann, A., Zehnder, A.J.B., Wehrli, B., and Holliger, C., Influence of temperature and high acetate concentrations on methanogenesis in lake sediment slurries, FEMS Microbiol. Ecol., 2007, vol. 62, pp. 336–344.
Och, L.M., Muller, B., Voegelin, A., Ulrich, A., Göttlicher, J., Steiniger, R., Mangold, S., Vologina, E., and Sturm, M., New insight into the formation and burial of Fe/Mn accumulations in Lake Baikal sediments, Chem. Geol., 2012, vol. 330–331, pp. 244–259.
Pacheco-Oliver, M., McDonald, I., Groleau, D., Murrell, J.C., and Miguez, C., Detection of methanotrophs with highly divergent pmoA genes from Arctic soils, FEMS Microbiol. Lett., 2002, vol. 209, pp. 313–319.
Pannekens, M., Kroll, L., Müller, H., Mbow, F.T., and Meckenstock, R.U., Oil reservoirs, an exceptional habitat for microorganisms, New Biotechnol., 2019, vol. 49, pp. 1–9.
Pasche, N., Schmid, M., Vazquez, F., Schubert, C J., Wüest, A., Kessler, J.D., Pack, M.A., Reeburgh, W.S., and Bürgmann, H., Methane sources and sinks in Lake Kivu, J. Geophys. Res., 2011, vol. 116, G03006.
Pavlova, O.N., Adamovich, S.N., Mirskova, A.N., and Zemskaya, T.I., RF Patent 2694593, 2019.
Pavlova, O.N., Adamovich, S.N., Novikova, A.S., Gorshkov, A.G., Izosimova, O.N., Ushakov, I.A., Oborina, E.N., Mirskova, A.N., and Zemskaya, T.I., Protatranes, effective growth biostimulants of hydrocarbon-oxidizing bacteria from Lake Baikal, Russia, Biotechnol. Rep., 2019, vol. 24, e00371.
Pavlova, O.N., Bukin, S.V., Lomakina, A.V., Kalmychkov, G.V., Ivanov, V.G., Morozov, I.V., Pogodaeva, T.V., Pimenov, N.V., and Zemskaya, T.I., Production of gaseous hydrocarbons by microbial communities of Lake Baikal bottom sediments, Microbiology (Moscow), 2014, vol. 83, pp. 798–804.
Pavlova, O.N., Izosimova, O.N., Chernitsyna, S.M., Ivanov, V.G., Pogodaeva, T.V., and Gorchkov, A.G., Process of anaerobic oxidation of oil in bottom sediments of Lake Baikal, Limnol. Freshwater Biol., 2020, no. 3, pp. 1006–1007.
Pavlova, O.N., Izosimova, O.N., Gorshkov, A.G., Novikova, A.S., Bukin, S.V., Ivanov, V.G., Khlystov, O.M., and Zemskaya, T.I., Current state of deep oil seepage near cape Gorevoi Utes (Central Baikal), Russ. Geol. Geophys., 2020, vol. 61, pp. 1007–1014.
Petrova, V.I. and Mamontova, L.M., Changes in bacterial abundance in experiments with oil addition, in Mikroorganizmy v ekosistemakh ozer i vodokhranilishch (Microorganisms in the Ecosystems of Lakes and Reservoirs), Novosibirsk: Nauka, pp. 144–150.
Pimenov, N.V., Zakharova, E.E., Bryukhanov, A.L., Korneeva, V.A., Kuznetsov, B.B., Tourova, T.P., Pogodaeva, T.V., Kalmychkov, G.V., and Zemskaya, T.I., Activity and structure of the sulfate-reducing bacterial community in the sediments of the southern part of Lake Baikal, Microbiology (Moscow), 2014, vol. 83, pp. 47–55.
Pogodaeva, T.V., Lopatina, I.N., Khlystov, O.M., Egorov, A.V., and Zemskaya, T.I., Background composition of pore waters in Lake Baikal bottom sediments, J. Great Lake Res., 2017, vol. 43, pp. 1030–1043.
Pogodaeva, T.V., Poort, J., Aloisi, G., Bataillard, L., Makarov, M.M., Khabuev, A.V., Kazakov, A.V., Chensky, A.G., and Khlystov, O.M., Fluid migrations at the Krasny Yar methane seep of Lake Baikal according to geochemical data, J. Great Lakes Res., 2020, vol. 46, pp. 123–131.
Pogodaeva, T.V., Zemskaya, T.I., Golobokova, L.P., Khlystov, O.M., Minami, Kh., and Sakagami, Kh., Pore water chemical composition in the bottom sediments from different Lake Baikal regions, Geol. Geofiz., 2007, vol. 48, pp. 1144–1160.
Pujalte, M.J., Lucena, T., Ruvira, M.A., Arahal, D.R., and Macian, M.C., The family Rhodobacteraceae, in The Prokaryotes: Alphaproteobacteria and Betaproteobacteria, Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., and Thompson, F., Eds., Berlin: Springer, 2014, pp. 439–512.
Qiu, L., Williams, D.F., Gvorzdkov, A., Karabanov, E., and Shimaraeva, M., Biogenic silica accumulation and paleoproductivity in the northern basin of Lake Baikal during the Holocene, Geology, 1993, vol. 21, pp. 25–28.
Raghoebarsing, A.A., Pol, A., van de Pas-Schoonen, K.T., Smolders, A.J., Ettwig, K.F., Rijpstra, W.I., Schouten, S., Damsté, J.S.S., Op den Camp, J.M., Jetten, M.S., and Strous, M., A microbial consortium couples anaerobic methane oxidation to denitrification, Nature, 2006, vol. 440, pp. 918–921.
Rissanen, A.J., Peura, S., Mpamah, P.A., Taipale, S., Tiirola, M., Biasi, C., Maki, A., and Nykanen, H., Vertical stratication of bacteria and archaea in sediments of a small boreal humic lake, FEMS Microbiol. Lett., 2019, vol. 366, no. 5, fnz044.
Schubert, C.J., Vazquez, F., Losekann-Behrens, T., Knittel, K., Tonolla, M., and Boetius, A., Evidence for anaerobic oxidation of methane in sediments of a freshwater system (Lago di Cadagno), FEMS Microbiol. Ecol., 2011, vol. 76, pp. 26–38.
Schulz, S. and Conrad, R., Influence of temperature on pathways to methane production in the permanently cold profundal sediment of Lake Constance, FEMS Microbiol. Ecol., 1996, vol. 20, pp. 1–14.
Shen, L., Ouyang, L., Zhu, Y., and Trimmer, M., Active pathways of anaerobic methane oxidation across contrasting riverbeds, ISME J., 2019, vol. 13, pp. 752–766.
Shubenkova, O.V., Zemskaya, T.I., Chernitsyna, S.M., Khlystov, O.M., and Triboi, T.I., The first results of an investigation into the phylogenetic diversity of microorganisms in southern Baikal sediments in the region of subsurface discharge of methane hydrates, Microbiology (Moscow), 2005, vol. 74, pp. 314–320.
Sierra-Garcia, I.N., Dellagnezze, B.M., Santos, V.P., Chaves, M.R., Capilla, R., Neto, S., Gray, N., and Oliveira, V.M., Microbial diversity in degraded and non-degraded petroleum samples and comparison across oil reservoirs at local and global scales, Extremophiles, 2017, vol. 21, pp. 211–229.
Simoneit, B.R.T., Aboul-Kassim, T.A.T., and Tiercelin, J.J., Hydrothermal petroleum from lacustrine sedimentary organic matter in the East African Rift, Appl. Geochem., 2000, vol. 15, pp. 355–368.
Sitnikova, T.Ya., Sideleva, V.G., Kiyashko, S.I., Zemskaya, T.I., Mekhanikova, I.V., Khlystov, O.M., and Khal’zov, I.A., Comparative analysis of macroinvertebrates and fish communities associated with methane and oil-methane seeps in Lake Baikal abyssal zone, Usp. Sovr. Biol., 2017, vol. 137, pp. 373–386.
Söllinger, A. and Urich, T., Methylotrophic methanogens everywhere—physiology and ecology of novel players in global methane cycling, Biochem. Soc. Trans., 2019, vol. 47, pp. 1895–1907.
Spang, A., Poehlein, A., Offre, P., Zumbragel, S., Haider, S., Rychlik, N., Nowka, B., Schmeisser, C., Lebedeva, E.V., Rattei, T., Böhm, C., Schmid, M., Galushko, A., Hatzenpichler, R., Weinmaier, T., et al., The genome of the ammonia-oxidizing Candidatus Nitrososphaera gargensis: Insights into metabolic versatility and environmental adaptations, Environ. Microbiol., 2012, vol. 14, pp. 3122–3145.
Starnawski, P., Bataillon, T., Ettema, T.J.G., Jochum, L.M., Schreiber, L., Chen, X., Lever, M.A., Polz, M.F., Jørgensen, B.B., Schramm, A., and Kjeldsen, K.U., Microbial community assembly and evolution in subseafoor sediment, Proc. Natl. Acad. Sci. U. S. A., 2017, vol. 114, pp. 2940–2945.
Sun, L.W., Toyonaga, M., Ohashi, A., Tourlousse, D.M., Matsuura, N., Meng, X.Y., Tamaki, H., Hanada, S., Cruz, R., Yamaguchi, T., and Sekiguchi, Y., Lentimicrobium saccharophilum gen. nov., sp nov., a strictly anaerobic bacterium representing a new family in the phylum Bacteroidetes, and proposal of Lentimicrobiaceae fam. nov., Int. J. Syst. Evol. Microbiol., 2016, vol. 66, pp. 2635–2642.
Taliev, S.D., Kozhova, O.M., and Molozhavaya, O.A., Hydrocarbon-oxidizing microorganisms in biocenoses of some Lake Baikal regions, in Mikroorganizmy v ekosistemakh ozer i vodokhranilishch (Microorganisms in the Ecosystems of Lakes and Reservoirs), Novosibirsk: Nauka, 1985, pp. 64–74.
Thauer, R.K., Kaster, A.K., Seedorf, H., Buckel, W., and edderich, R., Methanogenic archaea: ecologically relevant differences in energy conservation, Nat. Rev. Microbiol., 2008, vol. 6, pp. 579–591.
Timmers, P.H., Welte, C.U., Koehorst, J.J., Plugge, C.M., Jetten, M.S., and Stams, A.J., Reverse methanogenesis and respiration in methanotrophic archaea, Archaea, 2017, vol. 2017, 1654237.
Torres, N.T., Och, L.M., Hauser, P.C., Furrer, G., Brandl, H., Vologina, E., Sturm, M., Bürgmann, H., and Müller, B., Early diagenetic processes generate iron and manganese oxide layers in the sediments of Lake Baikal, Siberia, Environ. Sci., 2014, vol. 16, pp. 879–889.
Vanwonterghem, I., Evans, P.N., Parks, D.H., Jensen, P.D., Woodcroft, B.J., Hugenholtz, P., and Tyson, G.W., Methylotrophic methanogenesis discovered in the archaeal phylum Verstraetearchaeota, Nature Microbiol., 2016, vol. 1, p. 16170.
Vologina, E.G. and Sturm, M., Particulate fluxes in South Baikal: evidence from sediment trap experiments, Russ. Geol. Geophys., 2017, vol. 58, pp. 1045–1052.
Vologina, E.G., Sturm, M., Vorobyova, S.S., and Granina, L.Z., New results of high-resolution studies of surface sediments of Lake Baikal, Terra Nostra, 2000, no. 9, pp. 115–131.
Votintsev, K.K., Meshcheryakova, A.I., and Popovskaya, G.I., Krugovorot organicheskogo veshchestva v ozere Baikal (Organic Matter Turnover in Lake Baikal), Novosibirsk: Nauka, 1975.
Vykhristyuk, L.A., Organic matter in Lake Baikal bottom sediments, in Trudy LIN SO AN SSSR, Novosibirsk: Nauka, 1980, vol. 32, p. 80.
Walker, C.B., de la Torrea, J.R., Klotz, M.G., Urakawa, H., Pinel, N., Arp, D.J., Brochier-Armanet, C., Chain, P.S., Chan, P.P., Gollabgir, A., Hemp, J., Hugler, M., Karr, E.A., Konneke, M., Shin, M., et al., Nitrosopumilus maritimus genome reveals unique mechanisms for nitrification and autotrophy in globally distributed marine crenarchaea, Proc. Natl. Acad. Sci. U. S. A., 2010, vol. 107, pp. 8818–8823.
Wand, U., Samarkin, V.A., Nitzsche, H.-M., and Hubberten, H.-W., Biogeochemistry of methane in the permanently ice-covered Lake Untersee, central Dronning Maud Land, East Antarctica, Limnol. Oceanogr., 2006, vol. 51, pp. 1180–1194.
Weber, H.S., Habicht, K.S., and Thamdrup, B., Anaerobic methanotrophic archaea of the ANME-2d cluster are active in a low-sulfate, iron-rich freshwater sediment, Front. Microbiol., 2017, vol. 8, p. 619.
Welte, C.U., Rasigraf, O., Vaksmaa, A., Versantvoort, W., Arshad, A., Op den Camp, H.J., Jetten, M.S., Lüke, C., and Reimann, J., Nitrate- and nitrite-dependent anaerobic oxidation of methane, Environ. Microbiol. Rep., 2016, vol. 8, p. 941.
Wen, X., Yang, S.Z., Horn, F., Winkel, M., Wagner, D., and Liebner, S., Global biogeographic analysis of methanogenic Archaea identifies community-shaping environmental factors of natural environments, Front. Microbiol., 2017, vol. 8, art. 1339.
Wurzbacher, C., Nilsson, R.H, Rautio, M., and Peura, S., Poorly known microbial taxa dominate the microbiome of permafrost thaw ponds, ISME J., 2017, vol. 11, pp. 1938–1941.
Yanagawa, K., Shiraishi, F., Tanigawa, Y., Maeda, T., Mustapha, N.A., Owari, S., Tomaru, H., Matsumoto, R., and Kano, A., Endolithic microbial habitats hosted in carbonate nodules currently forming within sediment at a high methane flux site in the Sea of Japan, Geosciences, 2019, vol. 9, p. 463.
Yang, Y., Chen, J., Tong, T., Xie, S., and Liu, Y., Influences of eutrophication on methanogenesis pathways and methanogenic microbial community structures in freshwater lakes, Environ. Pollut., 2020, vol. 260, art. 114106.
Yoon, J., Matsuo, Y., Katsuta, A., Jang, J.H., Matsuda, S., Adachi, K., Kasai, H., and Yokota, A., Haloferula rosea gen. nov., sp. nov., Haloferula harenae, sp. nov., Haloferula phyci sp. nov., Haloferula helveola sp. nov and Haloferula sargassicola sp. nov., five marine representatives of the family Verrucomicrobiaceae within the phylum “Verrucomicrobia,” Int. J. Syst. Evol. Microbiol., 2008, vol. 58, pp. 2491–2500.
Zakharova, Y.R., Parfenova, V.V., Granina, L.Z., Kravchenko, O.S., and Zemskaya, T.I., Distribution of iron- and manganese-oxidizing bacteria in the bottom sediments of Lake Baikal, Inland Water Biol., 2010, vol. 3, pp. 313–321.
Zakharova, Y.R., Petrova, D.P., Galachyants, Y.P., Bashenkhaeva, M.Y., Kurilkina, M.I., and Likhosh-way, Y.V., Bacterial and archaeal community structure in the surface diatom sediments of deep freshwater Lake Baikal (Eastern Siberia), Geomicrobiol. J., 2018, vol. 35, pp. 635–647.
Zarate-del Valle, P.F., Rushdi, A.I., and Simoneit, B.R.T., Hydrothermal petroleum of Lake Chapala, Citala Rift, western Mexico: bitumen compositions from source sediments and application of hydrous pyrolysis, Appl. Geochem., 2006, vol. 21, pp. 701–712.
Zavarzin, G.A., Formation of the system of biogeochemical cycles, Paleontol. J., 2003, vol. 37, pp. 576–583.
Zemskaya, T.I., Lomakina, A.V., Mamaeva, E.V., Zakharenko, A.S., Likhoshvai, A.V., Galachyants, Yu.P., and Miller, B., Composition of microbial communities in sediments from southern Baikal containing Fe/Mn concretions, Microbiology (Moscow), 2018, vol. 87, pp. 382–392.
Zemskaya, T.I., Lomakina, A.V., Mamaeva, E.V., Zakharenko, A.S., Pogodaeva, T.V., Petrova, D.P., and Galachyants, Yu.P., Bacterial communities in sediments of Lake Baikal from areas with oil and gas discharge, Aquat. Microbiol. Ecol., 2015a, vol. 75, pp. 95–109.
Zemskaya, T.I., Lomakina, A.V., Shubenkova, O.V., Pogodaeva, T.V., Morozov, I.V., Chernitsina, S.M., Sitnikova, T.Ya., Khlystov, O.M., and Egorov, A.V., Jelly-like microbial mats over subsurface fields of gas hydrates at the St. Petersburg methane Seep (Central Baikal), Geomicrobiol. J., 2015b, vol. 32, pp. 89–100.
Zemskaya, T.I., Namsaraev, B.B., Dul’tseva, N.M., Khanaeva, T.A., Golobokova, L.P., Dubinina, G.A., Dulov, L.E., and Wada, E., Ecophysiological characteristics of the mat-forming bacterium Thioploca in bottom sediments of the Frolikha Bay, Northern Baikal, Microbiology (Moscow), 2001, vol. 70, pp. 335–341.
Zemskaya, T.I., Sitnikova, T.Y., Kiyashko, S.I., Kalmychkov, G.V., Pogodaeva, T.V., Mekhanikova, I.V., Naumova, T.V., Shubenkova, O.V., Chernitsina, S.M., Kotsar, O.V., Chernyaev, E.S., and Khlystov, O.M., Faunal communities at sites of gas- and oil-bearing fluids in Lake Baikal, Geo-Mar. Lett., 2012, vol. 32, pp. 437–451.
Zhu, G., Jetten, M.S.M., Kuschk, P., Ettwig, K.F., and Yin, C., Potential roles of anaerobic ammonium and methane oxidation in the nitrogen cycle of wetland ecosystems, Appl. Microbiol. Biotechnol., 2010, vol. 86, pp. 1043–1055.
Funding
This study was supported by the government contract (project no. 0279-2021-0006 “Investigation of Formation of Hydrate, Oil, and Gaseous Hydrocarbon Systems…” and the Russian Foundation for Basic Research (project no. 18-04-00244).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. This article does not contain any studies involving animals or human participants performed by any of the authors.
Additional information
Translated by A. Panyushkina
Rights and permissions
About this article

Cite this article
Zemskaya, T.I., Bukin, S.V., Lomakina, A.V. et al. Microorganisms in the Sediments of Lake Baikal, the Deepest and Oldest Lake in the World. Microbiology 90, 298–313 (2021). https://doi.org/10.1134/S0026261721030140
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0026261721030140