
Abstract
In this study, leaf nitrogen (N) and phosphorus (P) stoichiometry were used as indicators of nitrogen saturation and to assess ecosystem nutrient limitations. Schima superba, a representative and widely distributed dominant evergreen broadleaf tree species of the subtropical forests in southern China, was used for this purpose. A nutrient-addition experiment and a field survey were conducted to test the responses of trees from different provenances to N deposition. The relationships between leaf N and P stoichiometry and biomass, nutrient limitation, and soil N:P were analyzed. There was a relationship between leaf N, P, N:P, soil N:P and plant dry biomass. A threshold leaf N:P ratio (16.3) divided the five provenances into different nutrient-limitation classes that were related to the soil N:P ratio or N deposition. The leaf N:P ratio provided an indication of P limitation. A higher soil P level reduced the N deposition effect on plant growth. The leaf N:P ratio of individuals from different provenances can be used as a predictor of nutrient limitation, and this was related to the soil N:P ratio.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
The nitrogen (N) and phosphorous (P) levels in plant tissues, especially in the leaves, have often been studied, and the N:P ratio is considered an important indicator of nutrient limitation within individual species as well as for the entire ecosystem1,2,3,4,5,6,7,8. In wetland ecosystems, the leaf N:P ratio is <14 under N-limited conditions and >16 under P-limited conditions9. However, in upland ecosystems, the threshold value of the leaf N:P ratio varies considerably (ranging from 6.7 to 16 under N-limited conditions and 12.5 to 26.3 under P-limited conditions), and vegetation growth is limited by P10. Numerous studies have shown that leaf N and P stoichiometry is affected by factors such as habitat, functional group, and growth stage4,5,6,11,12,13. An important environmental factor influencing plant biomass and the leaf N:P ratio is the N:P ratio of the soil solution4,7. A species that occurs in different areas could have different leaf N:P ratios based on soil N:P ratio variation14. In southern China, a higher leaf N:P ratio occurs in woody plants in response to a shortage of soil P6,13.
N deposition is high in the subtropical forests of southern China. Zhu et al. reported that the average deposition fluxes of N and P in China were 13.69 ± 8.69 kg ha−1 a−1 and 0.21 ± 0.17 kg ha−1 a−1, respectively8. The N:P ratios of atmospheric wet deposition (average = 77 ± 40) were found to be negatively correlated with current soil N:P ratios. In addition to the low soil P concentration, such N:P ratio imbalances will disturb nutrient availability and increase P limitation8,15. This deposition trend is expected to increase in future decades, and the production of reactive N will reach 270 Tg N a−1 by 20508,16,17,18,19,20,21,22. We evaluated how a widespread species of different provenances would respond to soil N:P ratios and determined the biomass patterns corresponding to the N:P ratios of this species under N deposition in China. Schima superba, a representative and widely distributed broadleaf evergreen tree species of subtropical forests in southern China, was used as the study material23. Based on uniform levels of simulated N deposition in several provenances of this species, we found that the root system was more developed, and P absorption efficiency (PAE) was higher under N deposition, which was related to soil nutrition24. From these results, we inferred that there would be a relationship between biomass and the leaf N:P ratios and that changes would occur in the leaf N and P stoichiometry patterns in this species under N deposition. We also inferred that leaf N:P in S. superba could be a predictor of nutrient limitation in nursery breeding or in forest management. Our objectives were to (a) compare the seedling growth, leaf N, P, and N:P ratio in S. superba from different provenances in P-limited and P-rich soil under simulated N deposition treatments; (b) determine whether seedlings from different provenances with different N and P availability exhibited the same nutrient limitation; (c) compare the N:P ratio of different forest sites (Table 1, Fig. 1) using results of (a); and (d) determine the relationship between the leaf N:P ratio, the soil N:P ratios, and the biomass of S. superba, and assess N deposition effects on leaf N and P stoichiometry.

Geographical distribution of the study provenances of S. superba in southern China.
Results
Leaf N and P concentrations and the N:P ratios under N deposition
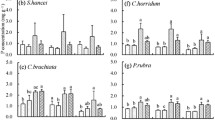
There was no interaction between provenances and nutrient input level, and only main effect responses for all variables were observed under the experimental treatment (Table 2). Regardless of the P effect, the leaf N concentration (16.8 mg g−1, p < 0.05) and N:P ratio (19.5, p < 0.05) in the N200 treatment were significantly higher than in the other three treatments (N0: 10.0 mg g−1 and 9.9; N50: 10.2 mg g−1 and 13.1; N100: 12.4 mg g−1 and 17.0, respectively). However, the highest leaf P concentration was measured in the N0 treatment (1.06 mg g−1) compared to N50 (0.86 mg g−1); N100 (0.88 mg g−1), and N200 (0.77 mg g−1). The biomass, leaf P concentration, and N:P ratios all significantly differed among provenances, P, and N treatments. The leaf N did not differ significantly among the provenances (Tables 2, 3).
The leaf N, P and N:P ratios exhibited large variations across forest sites, ranging from 13–18 mg g−1 for N, 0.2–0.4 mg g−1 for P, and 35–85 for the N:P ratios (Table 4). The mean values of the leaf N, P and N:P ratio for all forest sites were 14.7 mg g−1, 0.34 mg g−1, and 50.0, respectively.
The leaf N:P ratios increased with N deposition level both in the experimental treatments (Table 3) and forest field sites (Table 4).
Relationship between biomass and leaf N, leaf P, and N:P ratio
Both aboveground biomass and total biomass were significantly positively correlated with leaf N and P, and they were more strongly correlated with leaf N under experimental N deposition treatment (Table 5). The aboveground biomass and the leaf N:P ratio had a correlation coefficient of 0.44, p < 0.001 (Table 5), and a regression curve (Waboveground Biomass = −0.014(ratioN:P)2 + 0.4552 ratioN:P + 0.4318, R2 = 0.2958, p < 0.001, n = 192) was fitted. The highest biomass value was measured at a leaf N:P ratio of 16.3.
Although the field forest leaf N:P ratio was higher than that recorded in the experimental N deposition study, there was a similar trend between the biomass and the leaf N:P ratio (Tables 1, 4).
The aboveground biomass was significantly negatively correlated with the soil N:P ratio and the correlation coefficient was −0.6, p < 0.001 (Table 5). The relationship between the forest field biomass and the soil N:P (Table 3) was the same as that shown for the experimental results (Table 4). The five provenances in the experiment had a higher aboveground biomass when the soil N:P value was less than 2. JO had the highest biomass, which was associated with a leaf N:P ratio of 14.9 (Table 3). In the forest field site, JO-F also had the highest biomass, and its soil N:P ratio was only 1.5 (Table 4).
The principal components model using nutrient element, biomass, and environmental variables from five forest sites established four main principal components with a degree of explanation >87% (Table 6). The first component explained 41.10% of the variability, the second component explained 21.42%, the third one explained 14.61%, and the fourth one explained 10.12% (Table 6). Principal component 1 was related to leaf N:P, sample sites, N deposition, and biomass (Table 7). Soil N:P was correlated with principal component 2, and it was correlated statistically (p < 0.05) with annual temperature and tree height (Table 7).
Discussion
Leaf N, P, and N:P under N deposition
We conducted an analysis of the leaf N and P stoichiometry from five different provenances inhabited by S. superba, a widely distributed and dominant subtropical broad-leaved evergreen in southern China.
Leaf N and leaf N:P ratio were increased but leaf P was decreased with the addition of N. This result is similar to the results of Mo et al. for two species (S. superba and Cryptocarya concinna) in China18. Increases in N availability under N deposition are typically associated with increases in plant N concentration25,26 and decreases in P concentration27,28,29,30. This is related to the increase in N supply, and the greater N uptake results in an increased uptake of other nutrients such as P. However, these other nutrients may become limiting over time. After long-term N deposition, the concentration of P, Ca, Mg, or other cations in soils tends to decrease28,30, and deficiency of these elements in plants is often observed14,29,30,31,32,33. Chronic increases in N deposition increase the soil and leaf N:P ratios, limiting plant growth24,34,35,36 and photosynthetic rates32,37 and leading to less diverse ecosystems. The abundance of nitrophilous species may increase38. A higher soil P concentration could reduce the negative effect of N deposition, increase the leaf N and P concentration, and decrease the leaf N:P (Low-P: WAB = 2.5 g, LN = 9.8 mg g−1, LP = 0.7 mg g−1, L N:P = 15.5; High-P: WAB = 4.5 g, LN = 16.6 mg g−1, LP = 1.3 mg g−1, L N:P = 13.8). The leaf N:P increased from the southwest to the northeast. The soil P and N:P were related to temperature in principal component 2 (Table 7). The soil P concentration increases from the southeastern coastal areas to the inland areas of southern China39,40, and this is due to temperature leading to soil P decomposition by weathering. Therefore, soil P concentration is an important factor affecting plant development, especially plant height, and it reflects a bio-geographical gradient pattern at local scales2,13.
A previous study found that leaf N and P stoichiometry could be used as indicators of nutrient limitation in the ecosystem1,2,3,4,5,6,7,8. We found that the leaf N:P ratio was a good predictor of nutrient limitation of individual species such as S. superba. Young seedlings in the experiment had a leaf N:P ratio (15.3) that was similar to 149 evergreen tree species (15.2) and 255 broadleaf tree species (15.1) in China13, and 74 evergreen woody species (13.4) worldwide41. This suggested that the leaf N:P was weakly affected by the seedling age and species difference and had a similar value as large categories of woody species42. However, in our forest site survey, the average value of the leaf N:P ratio was 49.9, which is 3.4 times greater than the average value found in previous studies. A possible cause of the substantially higher leaf N:P ratio may be the different soil and growth conditions of S. superba. Roughgarden predicted that the level of phenotypic variance is often associated with niche width43. Forest ecosystems have higher N and P deposition ratios (91:1) than other ecosystem types8. At the same time, larger crowns and greater leaf areas intercept more N from N deposition, and S. superba growing in forest sites absorbs more inorganic N through the leaves44. We found that the soil N concentration was high in the five forest sites, and this may be maintained by a deep litter layer and high organic matter content (Table 3). Thus, high levels of inorganic N and leaching from foliar tissue or directly from N deposition may be transported into the soil and lead to soil N saturation. In addition, an increased N supply and uptake can result in the soil deficiency of P and other elements14,28,29,30,31,32. This means that niche size, with respect to nutrient cycling, would be in the range of mineral element concentrations in environments in which the species can grow and reproduce42. The correlation and principal component analysis between leaf N:P and the environment confirmed that N deposition had a considerable effect on leaf N:P (r2 = 0.874, p = 0.019), and the leaf N:P ratio can be used as an indicator for individual species to assess the degree of N deposition (Table 7).
Leaf N:P, biomass, and nutrient limitation threshold
The threshold value indicated by the regression curve between the biomass and leaf N:P ratio was 16.3. When the leaf N:P ratio was lower than this value, plant growth was limited by N. In contrast, at a N:P ratio higher than 16.3, plant growth was limited by P. Leaf N:P ratios were considerably higher in the forest field sites, and these trees were limited by P. This may be caused by the very low available soil P in southern China36,37. However, different results were obtained from the experiment using one-year seedlings originating from the same provenances. For example, N limitations were becoming evident in seedlings from HZ, LQ, and JO (the leaf N:P ratio was lower than 16.3) (Table 2). N is present in forms such as amino acids or proteins and thus incurs a metabolic cost for storage45. Therefore, plants with rapid N metabolism undergo constant turnover, which creates a large N demand for plant growth46. N deposition satisfies the N demand, and trees growing in areas under long-term and high N deposition were found to have faster turnover47. If these plants had grown in areas without N deposition, they may have been N limited.
The extent to which the plant N:P ratio reflects that of the soil solution is a function of the homeostatic regulation exerted by the plant over its N:P stoichiometry4,7. For example, there was a greater increase in plant biomass in response to P addition compared with N addition. This suggested that the growth of S. superba at the five forest sites was limited by P-deficiency. Aerts and Chapin showed that P-deficient plants increase their P uptake rate and reduce the rate of N uptake to regulate the leaf N:P ratio, whereas N-deficient plants reduce the P uptake rate and increase the N uptake rate to maintain stoichiometric balance2. Therefore, there is a complex relationship between the soil N:P and the leaf N:P, and both of these ratios affect plant biomass. Based on the relationship between soil N:P and biomass, we found that when the soil N:P ratio was less than 1.8, the plant biomass increased. However, when this ratio was higher than 2.3, biomass was lower (Tables 1, 3 and 4). This suggests that a range of soil N:P can improve plant growth, and 1.8 may be an important threshold value for S. superba. Zhu et al. provided a formula for soil N:P ratios and atmospheric deposition N:P ratios, which was: N:Psoil = −0.0108 N:Patmospher + 3.3552 (R2 = 0.8, P = 0.007)8. We also propose that atmospheric deposition N:P ratios ranging from 97.7 to 144 would promote S. superba growth. Further studies are required to test this hypothesis.
Conclusions
We found that a higher soil P concentration could weaken the negative effect of N deposition. Plant leaf N:P ratios of S. superba from different provenances were affected by habitat, and this ratio could be an effective predictor of the nutrient limitation level of the habitat. Leaf N:P could be used as an nutrient index in tree breeding in addition to its traditional ecological significance. Soil N:P ratios had a complex relationship with leaf N:P and they were found to affect plant biomass. These results can guide the selection of optimal nutrient values for nursery tree breeding and forest management.
Materials and Methods
This study was conducted by the Forestry Genetic and Breeding Lab at the Research Institute of Subtropical Forestry, CAF, State Forestry Administration, China (RISF-CAF). The State Forestry Administration is responsible for national parks and other protected areas. No specific permission was required for these locations/activities, as they were based on non-destructive collection of plant material. The species is not endangered or protected, and the locations were not privately owned or protected by law.
Determination of the plant N:P ratio in forests
The five study sites for this project were in natural forests or plantations of S. superba. They were Hangzhou (HZ-F) and Longquan in Zhejiang province (LQ-F), Jianou in Fujian province (JO-F), Xinfeng in Jiangxi province (XF-F), and Guiyang in Hunan province (GY-F). The forest ages ranged from 12 to 30 years (Table 1). The N deposition in these areas over an approximate 30-year period (from 1980 to 2007) was 10–20 kg ha−1 a−1 in Hunan, 20–30 kg ha−1 a−1 in Jiangxi, 30–45 kg ha−1 a−1 in Fujian, and more than 45 kg ha−1 a−1 in Zhejiang province20 (Table 1, Fig. 1). The mature leaf samples for N and P analysis in this study were randomly collected from 30 trees in each of the forest sites in June 2017. The samples collected were the third or fourth leaf from the top of the current year branch, and 10 leaves were collected per tree. Soil samples (1 kg, collected from a 0–20 cm depth), collected from five different locations in each forest site, were pooled. The plant material and soil were dried and sieved through a 1 mm sieve to determine the mineral composition of the samples. They were then digested with H2SO4 and the catalyst H2O2, and diluted with deionized water to 50 mL after cooling. Total N was determined using the standard Kjeldahl method48. Total P was determined using the Mo-Sb anti-spectrophotography method49.
Experimental assessment
The experiment included five provenances (HZ, LQ, JO, XF, and GY) of S. superba seedlings, P-limited soil, and four levels of N addition using NH4NO3. The levels were N0 = 0 kg N ha−1 year−1, N50 = 50 kg N ha−1 year−1, N100 = 100 kg N ha−1 year−1, and N200 = 200 kg N ha−1 year−1. These N levels were determined based on current and historical N deposition in southern China8,17,19,20,50. The comparison set consisted of three provenances (HZ, JO, and XF), P-rich soil, and four levels of N addition. There were 12 replicates per treatment, with a total of 384 individuals. In November 2017, six plants were randomly chosen from each treatment; these plants were harvested and their biomass was determined. The soil in the six containers was mixed and the composition was determined.
The P-limited soil (P-L) used in this experiment was regular forest soil obtained from Zhejiang province. The pH value of this soil was 5.06 (potentiometric method, LY/T 1239–1999); total N was 0.41 g kg−1 (semi-micro Kjeldahl method, LY/T 1228–1999), hydrolyzable N was 46.43 mg kg−1 (alkaline hydrolysis diffusion method, LY/T 1229–1999), available P was 13 mg kg−1 (hydrochloric acid-sulfuric acid extraction method, LY/T 1233–1999), available potassium (K) was 130.08 mg kg−1 (1 mol L−1 ammonium acetate extraction–flame photometry method, LY/T 1236–1999), and organic matter was 6.68 g kg−1 (potassium dichromate oxidation–external heating method, LY/T 1237–1999). The P-rich soil (P-H) received KH2PO4 until the available P measured 60 mg kg−1.
The N deposition experiment was carried out from March to November 2017 in a glass house without climate control. The temperature in the glass house ranged from 5 °C to 15 °C from December to the following February and 26 °C to 34 °C during the period from July to September 2017. The S. superba seeds from the five provenances were germinated in March 2017. The seeds were collected from Hangzhou (HZ) and Longquan (LQ) in Zhejiang province, Jianou (JO) in Fujian province, Guiyang (GY) in Hunan province, and Xinfeng (XF) in Jiangxi province (Table 1). When the seedlings had two cotyledons and were 2–3 cm tall, they were planted in plastic containers (15 cm diam. × 18 cm high) in April 2017, with one seedling per container. A NH4NO3 solution was sprayed from the foliage to the soil once every two weeks for one month, and once a month for the next six months. The spraying began on 20th April 2017 when the leaves started to bud. The concentrations of the NH4NO3 solution were 0.0016 mol L−1 for N50, 0.0031 mol L−1 for N100 and 0.0062 mol L−1 for N200. The seedlings were watered every two days with distilled water until they were harvested during the first week of November 2017. Pests and weeds were controlled manually.
Data analysis
The data were analyzed using the SAS statistical program (version V8, SAS Institute, Cary, NC) and SPSS (PASW statistics 18). The N:P ratio of each plant and the soil was determined by dividing the N concentration by the P concentration. The biomass, leaf N concentration, leaf P concentration, and N:P ratio of the different N and P treatments were analyzed for statistical differences. Three-way analysis of variance (ANOVA) using a generalized linear model (GLM) was performed at a significance level of 0.05, with P, N, and provenance as the independent factors. All variables were normally distributed and did not need to be transformed. Duncan’s multiple range test was carried out to determine whether significant (p < 0.05) differences existed between different levels of the N and P treatments. The correlation coefficients between biomass, leaf N and P, and the N:P ratio for each treatment were calculated (p < 0.001). We computed the arithmetic mean of the N:P ratio and biomass of different provenances and then fitted regression equations to determine the relationship between the N:P ratios and biomass.
References
Fenn, M. E., Poth, M. A. & Johnson, D. W. Evidence for nitrogen saturation in the San Bernardino Mountains in southern California. Forest Ecology and Management. 82, 211–230 (1996).
Aerts, R. & Chapin, F. S. III. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research. 30, 1–67 (2000).
Reich, P. B. & Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences, USA 101, 11001–11006 (2004).
Güsewell, S. N. Pratios in terrestrial plants: variation and functional significance. New Phytologist 164, 243–260 (2004).
Kang, H. Z. et al. Variation in leaf nitrogen and phosphorus stoichiometry in Picea abies across Europe: An analysis based on local observations. Forest Ecology and Management 261, 195–202 (2011).
Wu, T. G., Yu, M. K., Wang, G. G., Dong, Y. & Cheng, X. R. Leaf nitrogen and phosphorus stoichiometry across forty-two woody species in southeast China. Biochemical Systematics and Ecology 44, 255–263 (2012).
Wu, T. G., Dong, Y., Yu, M. K., Wang, G. G. & Zeng, D. H. Leaf nitrogen and phosphorus stoichiometry of Quercus species across China. Forest Ecology and Management 284, 116–123 (2012).
Zhu, J. X. et al. Imbalanced atmospheric nitrogen and phosphorus depositions in China: Implications for nutrient limitation. Journal of Geophysical Research-Biogeoscience, 121, https://doi.org/10.1002/2016JG003393 (2016).
Koerselman, W. & Meuleman, A. F. M. The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology 33, 1441–1450 (1996).
Tessier, J. T. & Raynal, D. J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. Journal of Applied Ecology 40, 523–534 (2003).
Cunningham, S. A., Summerhayes, B. & Westoby, M. Evolutionary divergences in leaf structure and chemistry, comparing rainfall and soil nutrient gradients. Ecological Monographs. 69, 569–588 (1999).
Sterner, R. W., J. J. Elser. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton, NJ, USA: Princeton University Press. (2002).
Han, W. X., Fang, J. Y., Guo, D. L. & Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist 168, 377–385 (2005).
Sardans, J. & Peñuelas, J. The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiology 160, 1741–1761 (2012).
National soil survey office of China. Soil of China. Beijing, China: Chinese Agriculture Press, 483–486. (1998).
Xu, Y. G., Zhou, G. Y., Lou, T. S., Wu, Z. M. & He, Z. C. Soil solution chemistry and element budget in the forest ecosystem in Guangzhou. Acta Ecological Sinica 21, 1760–1861 (2001).
Zheng, L. X., Liu, X. J. & Zhang, F. S. Amospheric deposition of organic nitrogen: A review. Acta Ecologica Sinica 27, 3828–383 (2007).
Mo, J. M., Li, D. J. & Gundersen, P. Seedling growth response of two tropical tree species to nitrogen deposition in southern China. European Journal of Forest Research 127, 275–283 (2008).
Hu, Z. H. et al. Effects of simulated nitrogen deposition on soil respiration in northern subtropical deciduous broad-leaved forest. Environmental Science 31, 1726–1732 (2010).
Ti, C. P. & Yan, X. Y. Estimation of atmospheric nitrogen wet deposition in China’s mainland from based on N emission data. Journal of Agro-Environment Science 29, 1606–1611 (2010).
Liu, X. et al. Enhanced nitrogen deposition over China. Nature 494(7438), 459–462 (2013).
Yu, H. L. et al. Acid deposition in precipitation of Chinese natural ecosystems: Evidences from national-scale network monitoring. Journal of Geophysical Research-Atmosphere, 121, https://doi.org/10.1002/2015JD02441 (2016).
Luo, W. J. Forest germplasm resources conservation and utility. Zhejiang Science and Technology Press China 238–242 (2009).
Zhang, R., Zhou, Z. C., Luo, W. J., Wang, Y. & Feng, Z. P. Effects of nitrogen deposition on growth and phosphate efficiency of Schima superba of different provenances grown in phosphorus-barren soil. Plant and Soil 370, 435–445 (2013).
Baron, J. S. et al. Ecosystem responses to nitrogen deposition in the Colorado front range. Ecosystems (N Y). 3, 352–368 (2000).
Peñuelas, J. & Filella, I. Herbaria century record of increasing eutrophication in Spanish terrestrial ecosystems. Global Change Biology 7, 427–433 (2001).
Duquesnay, A., Dupouey, J. L., Clement, A., Ulrich, E. & Le, T. F. Spatial and temporal variability of foliar mineral concentration in beech (Fagus sylvatica) stands in northeastern France. Tree Physiology. 20, 13–22 (2000).
Kowalenko, C. G. The effect of nitrogen and boron fertilizer applications on Willamette red raspberry growth, and on applied and other nutrients in the plant and soil over two growing seasons. Canadian Journal of Plant Science 86, 213–225 (2006).
Sardans, J., Rivas-Ubach, A. & Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: a review and perspectives. Perspectives in Plant Ecology Evolution and Systematics 14, 33–47 (2012).
Lu, X. et al. Plant acclimation to long-term high nitrogen deposition in an N-rich tropical forest. PNAS www.pnas.org/cgi/doi/10.1073/pnas.1720777115 (2018).
Menge, D. N. L. & Field, C. B. Simulated global changes alter phosphorus demand in annual grassland. Global Change Biology 13, 2582–2591 (2007).
Lund, N., Christensen, T. R., Mastepanov, M., Lindroth, A. & Ström, L. Effects of N and P fertilization on greenhouse gas exchange in two northern peatlands with contrasting N deposition rates. Biogeosciences 6, 2135–2144 (2009).
Gilliam, F. S., May, J. D. & Adams, M. B. Response of foliar nutrients of Rubus allegheniensis to nutrient amendments in a central Appalachian hardwood forest. Forest Ecology and Management 411, 101–107 (2018).
Güsewell, S. High nitrogen:phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytologist 166, 537–550 (2005).
Granath, G., Strengbom, J. & Rydin, H. Direct physiological effects of nitrogen on Sphagnum: a greenhouse experiment. Functional Ecology. 26, 353–364 (2012).
Zhang, R., Wang, Y., Jin, G. Q., Zhou, Z. C. & Feng, Z. P. Nitrogen addition affects root growth, phosphorus and nitrogen efficiency of three provenances of Schima superba in barren soil. Acta Ecologica Sinica 33(12), 3611–3621 (2013).
Zhang, R. et al. The Effects of Nitrogen Deposition on Leaf Physiological and Photosynthetic Characters of Schima superba Seedlings from Three Provenances. Forest Research 26(2), 207–213 (2013).
Jung, K., J. H. Kwak, F. S. Gilliam, S. X. Chang. Simulated N and S deposition affected soil chemistry and understory plant communities in a boreal forest in western Canada. Journal of Plant Ecology https://doi.org/10.1093/jpe/rtx030 (2018).
Wang, T., Yang, Y. H. & Ma, W. H. Storage, patterns and environmental controls of soil phosphorus in China. Acta Scientiarum Naturalium Universitatis Pekinensis 44, 945–952 (2008).
Yan, J. L. Spatial heterogeneity research of soil phosphorus along different altitudes in Wuyi Mountain. Dissertation for Master’s degree of Fujian Agriculture and Forestry University. (2012)
Aerts, R. Nutrient resorption from senescing leaves of perennials: are there general patterns? Journal of Ecology. 84, 597–608 (1996).
Garten, C. T. Multivariate perspectives on the ecology of plant mineral element composition. Am. Natural 112, 533–544 (1978).
Roughgarden, J. Evolution of niche width. Amer. Nature 106, 683–718 (1972).
Uscola, M., Salvador, P. V., Oliet, J. & Warren, C. R. Foliar absorption and root translocation of nitrogen from different chemical forms in seedlings of two Mediterranean trees. Environmental and Experimental Botany 104, 34–43 (2014).
Chapin, F. S., Schulze, E. D. & Mooney, H. A. The ecology and economics of storage in plants. Annual Review of Ecology and Systematics. 21, 423–447 (1990).
Ostertag, R. Foliar nitrogen and phosphorus accumulation responses after fertilization: an example from nutrient-limited Hawaiian forests. Plant and Soil 334, 85–98 (2010).
Gilliam, F. S., C. A. Walter, M. B. Adams, W. T. Peterjohn. Nitrogen (N) dynamics in the mineral soil of a central appalachian hardwood forest during a quarter century of whole watershed N additions. Ecosystems https://doi.org/10.1007/s10021-018-0234-4 (2018).
Bremner, J. M., C. S. Mulvaney. Nitrogen-total. In: Page, A. L., Miller, R. H. & Keeney, D. R. eds Methods of soil analysis. Part 2: chemical and microbial properties, Agronomy Monograph 9. Agronomy Society of America, Madison. 595–624 (1982).
Anderson, J. M. & J. S. I., Ingram. Tropical soil biology and fertility: a handbook of methods. 2nd edn. CAB International Wallingford press (1993).
He, Y. Q. The status and utilization approach of forest soil fertility in tropical and subtropical China. Red SoilEcological Research (Volume II), Nanchang, China: Jiangxi Science and Technology Press, 16–22 (1993).
Cheng, Y. W., Hong, C. Z. & Wu, X. H. Qi. Distribution characters of Schima superba aboveground biomass and its productivity. Chinese Journal of Applied & Environmental Biology. 15(3), 318–322 (2009).
Acknowledgements
We thank Jiazhong Yu, Zhongcheng Li, Liuying Chen, Xiuhua Wang, and Pingsheng Zeng for their assistance in the field. This study was supported by the Special Fund for Forest new varieties breeding in Zhejiang (2016C02056), the Fifth Stages of Planting and Seedling Science and Technology of Fujian Province, and Key projects in Jiangxi Province (201503).
Author information
Authors and Affiliations
Contributions
R.Z. conceived and designed the study; R.Z. and H.P. performed the experiments, were involved in the analysis, and reviewed and edited the manuscript; B.H. and H.C. managed the field data; Z.Z. contributed reagents and materials; R.Z. drafted the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, R., Pan, H., He, B. et al. Nitrogen and phosphorus stoichiometry of Schima superba under nitrogen deposition. Sci Rep 8, 13669 (2018). https://doi.org/10.1038/s41598-018-32031-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-32031-y
- Springer Nature Limited
Keywords
This article is cited by
-
Leaf functional traits and ecological strategies of common plant species in evergreen broad-leaved forests on Huangshan Mountain
Journal of Forestry Research (2024)
-
Root morphology and physiology responses of two subtropical tree species to NH4+-N and NO3−-N deposition in phosphorus-barren soil
New Forests (2022)
-
Long-term nitrogen addition regulates root nutrient capture and leaf nutrient resorption in Larix gmelinii in a boreal forest
European Journal of Forest Research (2021)
-
Ecological stoichiometry and biomass responses of Agropyron michnoi under simulated N deposition in a sandy grassland, China
Journal of Arid Land (2020)
-
Seedling growth and nutrition responses of two subtropical tree species to NH4+-N and NO3−-N deposition
New Forests (2019)