
Abstract
The endosymbiotic bacterium Wolbachia occasionally increases host fitness or manipulates host reproductions to enhance vertical transmission. Multiple Wolbachia strains can coinfect the same host individual, which alters the density as well as phenotypes of the bacteria. However, the effects of Wolbachia coinfection on host fitness remain largely unknown. Here, we examined the effects of three phylogenetically distinct Wolbachia strains, wHm-a, wHm-b, and wHm-c, on host fitness by comparing non-infected, singly infected, and triply infected Homona magnanima lines within a fixed genetic background. By examining the effects of Wolbachia on host longevity, survivorship, and reproduction, we demonstrated that single infection with either wHm-b or wHm-c reduced host reproduction, but the triple infection led to the highest intrinsic growth rate. Susceptibility to the natural pathogens such as viruses and fungi was not different among the lines regardless of Wolbachia infection status. Cellular and humoral immunities were not affected by Wolbachia in females, whereas phenoloxidase activity was suppressed in males of all Wolbachia-infected lines, implying that it was a result of the mother’s curse hypothesis or a strategy of Wolbachia to increase their horizontal transmission efficiency. Although how the host’s genetic diversity affects the Wolbachia fitness effects is yet unknown, our findings indicated that the effects of Wolbachia are deeply influenced by infection status and that Wolbachia could change symbiotic strategy depending on host sex and transmission route.
Similar content being viewed by others

Introduction
Wolbachia, an Alphaproteobacteria, is the most widespread endosymbiotic bacterium in arthropods and reside in 40–60% insect species (Duron et al. 2008; Werren et al. 2008; Zug and Hammerstein 2012). Since it localises in the host cytoplasm and is mostly transmitted vertically from the mother to offspring, Wolbachia is considered to have developed strategies to compensate host fitness and to manipulate host reproduction (Werren et al. 2008).
Wolbachia confers fitness advantages to its host by establishing mutualistic relationship. In Bemisia tabaci, Wolbachia shortens host developmental duration and alters larval survival (Xue et al. 2012). In the case of Drosophia suzukii, Wolbachia infection increases the number of eggs in the female hosts (Mazzetto et al. 2015). These characteristics are related to the intrinsic growth rate (r), a population dynamics measure, of hosts (Birch 1948). Furthermore, Wolbachia enhances host resistance to natural pathogens; in the case of D. melanogaster, Wolbachia helps induce resistance to RNA viruses (Teixeira et al. 2008). Certain Wolbachia strains reduce susceptibility to RNA viruses in dipteran insect hosts (Moreira et al. 2009; Stevanovic et al. 2015). Wolbachia also enhances humoral immune responses, such as phenoloxidase (PO) activity, and stimulates the expression of genes related to early immunity in dipteran insects (Thomas et al. 2011; Rancès et al. 2012).
Conversely, association with Wolbachia can sometimes incur costs for hosts. Some Wolbachia strains negatively affect host fitness by shortening adult longevity and reducing female host fecundity (Hoffmann et al. 1990; Fleury et al. 2000). However, it is considered that parasitic Wolbachia has evolved to form more mutualistic associations with their hosts (Weeks et al. 2007). Other costly traits are reproductive manipulations, such as male killing (a son killing phenomenon) and cytoplasmic incompatibility (CI), whereby a cross between males infected with Wolbachia and females uninfected or infected with different Wolbachia strain produces no or few offspring (Werren et al. 2008). Such reproductive manipulations enhance the prevalence of Wolbachia in host populations but could be costly, especially for male hosts, resulting in the development of resistance against the male-killing phenotype of Wolbachia (Hornett et al. 2006).
Furthermore, Wolbachia interacts with other Wolbachia strains or microbes within a single host individual, which affects the intensity of reproductive manipulations as well as population dynamics of Wolbachia, resulting in the evolution of host–symbiont interactions (Ikeda et al. 2003; Dobson et al. 2004; Kondo et al. 2005; Dean 2006; Mouton et al. 2006; Narita et al. 2007; Watanabe et al. 2011; Lu et al. 2012). Therefore, the host–bacteria and bacteria-bacteria interactions generate new outcomes and are important for the biology of Wolbachia.
Homona magnanima (Tortricidae, Lepidoptera), a serious tea pest, harbours three Wolbachia strains, wHm-a, wHm-b, and wHm-c, that influence host fitness and reproductions: wHm-a does not induce any reproductive phenotype, wHm-b causes CI, and wHm-c shortens the larval development time and increases pupal weight in female hosts (Arai et al. 2019; Arai et al. 2020; Takamatsu et al. 2021). Theoretically, multiple Wolbachia infections can be maintained in a host population if the coinfected hosts have more fitness advantages than the singly infected or uninfected hosts (Frank 1998, Vautrin and Vavre 2009). In this study, we hypothesise that the Wolbachia triple infection is more advantageous to its host H. magnanima than a single infection or a non-infection. To test this hypothesis, we compared multiple life history parameters of isogenic H. magnanima lines with different Wolbachia infection status and determined the advantages and strategies of triple infection of Wolbachia for its successful transmission in H. magnanima.
Materials and methods
Insects
Five lines of H. magnanima, namely Wabc, Wa, Wb, Wc, and W-, established in the laboratory as described by Arai et al. (2019), were used. The Wabc line, harbouring wHm-a, wHm-b, and wHm-c strains, was collected from Akiruno city, Tokyo, Japan, in 1999. The Wa, Wb, and Wc lines, established from the Wabc line via antibiotic treatments, are singly infected with wHm-a, wHm-b, and wHm-c, respectively. The W- line, free of any Wolbachia strains, was generated by subjecting the Wabc line to antibiotic treatment. We used the Wabc line from 116–240 generations Wa from 60–84 generations, Wb and Wc from 24–48 generations, W- from 96–120 generations for the experiments. All the larvae hatched from an egg mass were mass-reared as previously described in Arai et al. (2019), under laboratory conditions (16: 8 h light/dark cycle, 25 °C, and 60% relative humidity). We used males of the W- line to mate with females of each host line to homogenise their genetic backgrounds. The rearing and mating treatments were followed in each generation.
Fitness traits of H. magnanima
To investigate the longevity and fecundity of female adults, egg hatchability, and egg development time, a virgin female within 24 h after emergence and two males of the same host line were mated in a 120 mL plastic cup for oviposition. The mating sets comprised 50, 46, 40, 35 and 35 cups for the W−, Wa, Wb, Wc and Wabc lines, respectively. The areas of egg masses collected every day were determined, and the estimated total number of eggs per female oviposition was calculated from the total egg mass area, as described previously (Arai et al. 2019). Egg development time was defined as the number of days from oviposition to hatching. After hatching, pharate larvae in each egg mass were counted. The embryonic developmental ratio of eggs per female was calculated as the total number of pharate larvae divided by the estimated total number of eggs. The hatchability of pharate larvae was calculated as the number of hatched larvae divided by the total number of pharate larvae. The number of hatched larvae was determined as the difference between the total number of pharate larvae and unhatched pharate larvae. Survival of the female adults was recorded every day. After the death of female adults, they were dissected and spermatophore presence was checked to confirm mating. Non-mating females were excluded from the oviposition data. The longevity of the female adult was defined as the number of days from emergence till death. To determine the longevity of male adults, males within 24 h after emergence were individually reared in a plastic cup. Male adult longevity was defined as the number of days from emergence till death.
The pupation rates were recorded using 105 individuals (35 × 3 replicates) of each host line. The neonates hatched within 24 h were individually reared on an artificial diet INSECTA LF (Nosan Co., Ltd., Yokohama, Japan). The pupal duration and emergence rate were calculated using another 105 individuals (35 × 3 replicates) from each host line. The final instar larvae were individually reared using INSECTA LF until their eclosion. Pupal duration was recorded as the number of days from pupation to eclosion; the eclosion rate was calculated as total number of adults divided by the number of pupae.
The net reproduction rate (R0), the mean generation time (T), and the intrinsic growth rate (r) in each host line were calculated as follows (Birch 1948).
where lx is the survival rate of the female adult at x day old, and mx is the mean oviposition number (the number of pharate larvae) per day. The sex ratio is assumed to be 1:1 (Arai et al. 2019). The value of x and lx were defined as follows. Mean larval development times in each host line were cited from Arai et al. (2019).
x = (mean egg development time) + (mean larval development time) + (mean pupal duration) + (days after emergence).
lx = 0.5 (sex ratio) × (mean hatchability) × (mean pupation rate) × (mean eclosion rate) × (survival rate of the female adult at x days old).
To perform statistical analysis, Jackknife estimate of r (rj) was calculated as follows (Meyer et al. 1986):
where rall is the intrinsic growth rate calculated from the mean oviposition number per day of all females (mx, all), ri is the intrinsic growth rate calculated from the mean oviposition number per day of all females, excluding the i number individual (mx, i), and n is the number of all females. Jackknife estimates of R0 and T were also calculated using a similar equation as used for the calculation of rj (Maia et al. 2000).
Susceptibility of H. magnanima against viruses and fungi
The double-stranded DNA viruses Adoxophyes honmai entomopoxvirus (AHEV, Poxviridae) and Homona magnanima granulovirus (HomaGV, Baculoviridae) were isolated from diseased larvae collected from tea fields in Mizuho (Tokyo) and Kagoshima Prefecture Tea Experimental Station (Kagoshima), respectively. Occlusion bodies (OBs) of AHEV and HomaGV were purified as described previously (Takatsuka et al. 2010; Tsuruta et al. 2018). The concentration of AHEV OBs was determined via phase-contrast microscopy, using a Thoma hemocytometer, and that of Homa GV OBs was determined via Transmission electron microscopy using Latex beads (0.3 µm in diameter, Sigma Aldrich, St. Louis, MO). The purified OB suspensions of AHEV and HomaGV were adjusted to 2.0 × 107 OBs mL−1 and 2.0 × 106.5 OBs mL−1, respectively, using sterilised distilled water. Neonates (within 24 h of hatching, three replicates with 5–35 individuals) were allowed to feed on droplets of a viral suspension containing 10% sucrose and 5% red dye, with 1.0 × 107 OBs mL−1 for AHEV and 1.0 × 106.5 OBs mL−1 for HomoaGV, as described previously (Ishii et al. 2003). The droplets without viral OBs served as controls. Larvae that ingested the suspension were individually reared on an artificial diet and observed every day until they pupated or died. The dead insects were checked for symptoms of lethality by viruses.
The insect pathogenic fungus Beauveria bassiana was isolated from BotaniGard wettable powder (Arysta Lifescience Corporation, Tokyo, Japan). BotaniGard wettable powder was diluted ten times with sterilised distilled water, plated on to the Sabouraud agar medium, and incubated under a 16 h: 8 h light/dark cycle at 25 °C until conidia formation. Fungal conidia were scraped and suspended in sterilised 0.02% Tween 80, followed by centrifugation at 3000 × g for 10 min. Pellets were suspended in 10 mL sterilised 0.02% Tween 80. The concentration of conidia was adjusted to 2.0 × 109 mL−1. Fourth instar larvae (within 24 h of moulting, three replicates with 9–34 individuals) were soaked in the conidial suspension for 10 s, and sterilised 0.02% Tween 80 served as the control. After air-drying, the larvae were individually reared on an artificial diet and observed every day for mortality until they pupated or died under conidial infestation.
Immune traits of H. magnanima
Haemolymph was withdrawn from the fifth instar larvae from each host line after 3 days of moulting using a thin needle. To evaluate total haemocyte count (THC), the haemolymph of each larva was immediately diluted five times with 1 × PBS (pH 7.0), and haemocyte number per μL of diluted haemolymph was estimated by counting haemocyte with a light microscope using a Thoma haemocytometer. The THC (cells μL−1) of undiluted haemolymph was estimated as haemocyte number per μL of diluted haemolymph multiplied by the dilution rate.
To evaluate PO activity, the haemolymph of each larva was diluted ten times with 0.1 M potassium phosphate buffer (pH 6.0) and centrifuged at 9000 × g and 4 °C for 1 min. Then, 50 μL of the supernatant was dispensed into two wells of 96-well plate, and either 2 μL of ethanol or 5% phenylthiourea (PTU) in ethanol was added to the wells. Following incubation at room temperature (15–25 °C) for 15 min, 48 μL of 0.02 M L-dopa solution in 0.1 M potassium phosphate buffer was added to each well as the substrate. After incubation for 10 min at 30 °C, the absorbance at 490 nm was measured every 5 min for 2 h. PO activity was defined as the difference in absorbance between the two wells containing either ethanol or ethanol with PTU. The absorbance variation during the first 5 min was used for analysis because all samples showed a linear change for ~40 min.
Total PO activity was evaluated using the modified method of Bailey and Zuk (2008). After centrifuging the diluted haemolymph in PBS (pH 7.0), 50 μL each of the supernatants was dispensed into two wells of a 96-well plate containing 7 μL α- chymotrypsin solution (1.3 mg/mL DW, Sigma Aldrich C7762, St. Louis, Missouri, USA). After either ethanol or a 5% PTU and 0.02 M L-dopa solution in PBS was added, the absorbance was measured as mentioned above. Absorbance variation during the first 5 min was used for analysis because all samples showed a linear change by about 5 min. This experiment was done only for males because we could not get enough female larvae.
To examine the induced antibacterial activity of larval haemolymph, we conducted a growth inhibition assay. We injected 1 μL of a suspension of Escherichia coli (gram-negative) or Micrococcus luteus (gram-positive) (1.0 × 108 cells mL−1 in PBS) into the haemocoel of the fifth instar larvae from each host line (2 days after moulting), using a hand micro-applicator (Kiya Kogyo Seisakusho, Tokyo, Japan) fitted with a 50 μL micro-syringe (Ito Co., Shizuoka, Japan). After 24 h of individual rearing, haemolymph was collected on ice and centrifuged at 9000 × g and 4 °C for 1 min. Then, 5 μL of the supernatant was pipetted onto a 5 mm diameter filter paper, air-dried, and inoculated in to LB soft agar medium containing the same bacteria as mentioned above. After 24 h of incubation at 37 °C for E. coli or 30 °C for M. luteus, the area of growth inhibition zone was measured using Image J (https://imagej.nih.gov/ij/).
Statistical analysis
The survival data of H. magnanima larvae against viruses (AHEV and HomaGV) and fungi (B. bassiana), and adult longevity were analysed using the Kaplan-Meier survival estimates and pairwise log-rank tests between the curves, with p-values adjusted by Bonferroni using the “survival” package in R ver. 4.0.2 (R Core Team 2021). All data sets obtained in this study were tested for normality using Shapiro-Wilk test, and the data showing normal distributions (i.e., female emergence rate, female THC, and growth inhibition data) were analysed using a parametric Tukey-Kramer test. Other data sets (total number of eggs and pharate larvae per female, embryonic developmental ratio, hatchability, pupation rate, male emergence rate, pupal duration, adult longevity, T, R0, r, male THC, PO activity and total PO activity) were analysed using a non-parametric Steel-Dwass test. The Shapiro-Wilk, Tukey-Kramer, and Steel-Dwass tests were performed using JMP9 (SAS Institute Inc., Cary, NC, USA).
Results
Triple infection of Wolbachia resulted in high fitness advantages for H. magnanima
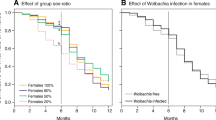
The Wabc line showed higher number of eggs per female individual than the Wc line (p = 0.042, Table 1, Fig. 1a) and a higher number of pharate larvae than the Wb and Wc lines (p < 0.05, Fig. 1b). Hatchability was lower in the Wb line than in other lines (p < 0.05, Table 1). Pupal duration was shorter in females of the W- line than in those of the other lines and was longer in the males of the Wabc line than those of the W-, Wa, and Wc lines (p < 0.05, Table 1). No significant difference was observed in the embryonic developmental ratio, egg development time, pupation rate, emergence rate, and female adult longevity between the host lines (Log-rank test, p > 0.05, Table 1, Fig. 2). Notably, no analysis was carried out on the data for adult male longevity because we could only observe very low number of males.

a Number of eggs per female and b the number of pharate larvae per female. Different letters indicate significant differences between lines (p < 0.05). In each box-and-whisker plot, the centre line indicates the median. The upper and lower boundaries of the box indicate the upper quartile and lower quartile, respectively. The points on plots indicate each sample.

Kaplan-Meier curves of Homona magnanima females after emergence.
The intrinsic growth rate (r) was higher in the Wabc line than that in the singly infected or non-infected H. magnanima lines and the lowest in the Wb line harbouring only CI-inducing wHm-b (p < 0.01, Table 2). For parameters relating to the r value, the net reproduction rate (R0) was higher in the Wabc line than in the Wb and Wc lines (p < 0.01). Mean generation time (T) was shorter in the Wabc line than in the other lines (p < 0.01), except for the Wc line (p > 0.05), and longest in the Wb lines. All statistical data are shown in Tables S1 and S2.
Wolbachia did not protect hosts from natural pathogens but altered male immunity
Regardless of Wolbachia infection status, we observed no difference in larval susceptibility to AHEV, HomaGV, and B. bassiana among host lines (Log-rank test, p > 0.05, Fig. 3). Regarding immunity-related factors, the THC (cellular immunity) and induced antibacterial activity to E. coli of larval haemolymph (humoral immune response) were not different among host lines and sexes (p > 0.05, Fig. 4). Larval haemolymph of both sexes from each line (n = 3) did not inhibit the growth of M. luteus. No difference was observed in PO activity (humoral immune responses) among the female host lines (p > 0.05, Fig. 5a), while male larvae of all host lines harbouring Wolbachia showed lower PO activities than those in the W- line (Fig. 5b). In contrast, there was no difference in total PO activity in the haemolymph of male hosts among the lines, regardless of Wolbachia infection (p > 0.05, Fig. 5c).

Kaplan-Meier curves of the larvae after each treatment in the inoculation tests of a AHEV and b HomaGV, and the fourth instar larvae for days post-inoculation of c B. bassiana.

The area of growth inhibition zone of E. coli was measured using host larval haemolymph after 24 h of E. coli injection. The centre line and the upper and lower boundaries of the box indicate median, upper quartile, and lower quartile, respectively. The points on plots indicate each sample. The sample size is indicated in parentheses below host lines.

a Total haemocyte count and b phenoloxidase activities in female and male larval haemolymph, c total phenoloxidase activity in male larval haemolymph. Different letters indicate significant differences between lines (Steel-Dwass test, p < 0.05). The centre line and the upper and lower boundaries of the box indicate median, upper quartile, and lower quartile, respectively. The points on plots indicate each sample. The sample size is indicated in parentheses below host lines.
Discussion
In the present study, we demonstrated that females of the triply infected H. magnanima showed higher intrinsic growth rates (r) than other non-infected or singly-infected host lines. Although Wolbachia infection did not protect the hosts from natural pathogen attack, it specifically altered the PO activity in males but not in females. These findings suggest that triple Wolbachia infection is more advantageous for H. magnanima females than for other singly or non-infected females (Table 3, Fig. S1).
We previously demonstrated that wHm-b induced CI in the H. magnanima collected from Tokyo (Arai et al. 2019). The CI phenotype enabled rapid spreads of CI-inducing Wolbachia in its host population (Turelli and Hoffmann 1991; Ballad et al. 1996). Takamatsu et al. (2021) reported a high wHm-b infection frequency (approximately over 90%) in Shizuoka population, wherein triple infection was more frequent than wHm-b single infection. In this study, we found that wHm-b single infection reduced female fitness parameters, such as egg number and hatchability, resulting in reduced net reproduction rate (R0) and intrinsic growth rate (r). This result suggests that wHm-b single infection is not preferable for the host. Even if a Wolbachia strain affects its host negatively, other mutualistic and parasitic microbes would compensate the infection costs and increase the overall fitness advantage (Gómez-Valero et al. 2004, Frank 1998, Engelstädter et al. 2004), which contributes to the persistence of microbes in host populations (Hoffmann et al. 1990; Vautrin and Fabrice 2009). In H. magnanima, CI intensity was lower in the triply infected line than in the wHm-b singly infected line, corresponding to wHm-b density (Arai et al. 2019). We speculate that the lowered wHm-b density resulted in reduced fitness costs in the triply infected line. The hosts with wHm-c triple and single infection exhibited fitness advantages in insect growth, while only wHm-c single infection reduced the number of eggs laid per female host. Host fecundity is a crucial factor for Wolbachia transmission, but several studies have reported that Wolbachia infection reduces egg number (Hoffmann et al. 1990; Fleury et al. 2000). Generally, pupal weight and fecundity are positively correlated in lepidopteran insects such as Cydia pomonella (Tortricidae) (Deseo 1971; Danthanarayana 1975; Hough and Pimentel 1978). However, wHm-c infection increased larval growth and pupal weight (Arai et al. 2019) and reduced the number of eggs, probably by negatively affecting oocyte maturation or oviposition behaviour in female hosts. Similar to that of wHm-b, the lowered wHm-c density (Arai et al. 2019) may reduce the negative effects in the triply infected line. Otherwise, the presence of wHm-a may recover the negative effects in the triply infected line. Although the mechanism by which triply infected Wolbachia led to high intrinsic growth in their hosts is unknown, Wolbachia interactions may contribute to recover the negative effects observed in singly infected host lines. Future studies using double-infected lines would provide more information on how Wolbachia facilitates intrinsic growth rate.
We also identified the difference in H. magnanima immunity caused between the sexes by Wolbachia. Notably, the Wolbachia-infected H. magnanima males showed reduced PO activities (Fig. 5b). As total PO activity, all quantities of inactive precursors of PO (pro-PO) activated by α-chymotrypsin, did not differ regardless of the presence or absence of Wolbachia, reduced quantity of pro-PO activity may not be the cause of reduced PO activity in males. Wolbachia presence probably suppressed the pro-PO activation pathway, wherein pro-PO is cleaved by a proteolytic enzyme to form PO. Wolbachia sometimes increases host fitness by enhancing resistance to pathogens (Moreia et al. 2009; Stevanovic et al. 2015). Contrastingly, Wolbachia sometimes suppresses host immunity; the CI-inducing wDil strain suppresses host immunity by reducing phagocytosis and PO activities in males the terrestrial isopods Porcellio dilatatus (Pigeault et al. 2014). Wolbachia is transmitted mostly vertically from female hosts to offspring but not from male hosts; therefore, suppressed immunities in male hosts is not adaptive. This can be a result of natural selection explained by the mother’s curse hypothesis: inheritance through cytoplasm. If the elements inherited through cytoplasm (such as mitochondria) have mutation with severe effects for males but only mild effects for females, the mutation will undergo natural selection in females. The mutations increase to relatively a high frequency in a population because males do not transmit the mutation and male-specific phenotypes have no fitness consequences for the elements inherited through cytoplasm (Frank and Hurst 1996). Therefore, the inheritance through cytoplasm can result in sexually antagonistic traits. Since Wolbachia is also inherited through the cytoplasm of the female host, suppressed PO activity in only male hosts in H. magnanima would be a trait that has no fitness consequences for Wolbachia and does not support natural selection. Another possible interpretation is that the adaptation of Wolbachia for horizontal transmission from male hosts. In the manner of vertical transmission, infection in male host is a dead-end for Wolbachia. Accordingly, Wolbachia is thought to have evolved reproductive manipulations, such as male-killing, feminisation, and CI (Werren et al. 2008). But for the reproductive manipulations, Wolbachia would have developed horizontal transmission from males to other insects. It is unclear whether the horizontal transmission of Wolbachia occurs in H. magnanima, but several studies have suggested that parasitoid wasps harbour identical Wolbachia strains in their hosts and can transmit Wolbachia to a new insect from the original host (Vavre et al. 1999; Ahmed et al. 2015). The pro-PO activation in insect host is triggered by recognising the peptidoglycan layer of the cell wall of gram-negative bacteria (Takehana et al. 2002). Although a gram-negative Wolbachia does not have a proper cell wall, some studies have suggested the presence of peptidoglycan in Wolbachia (Vollmer et al. 2013; Voronin et al. 2014; Zug and Hammerstein 2015), which possibly triggers the pro-PO activation pathway. Despite of the presence of peptidoglycan, Wolbachia may prevent pro-PO activation in H. magnanima males with unknown manners and avoid elimination by the host immunity system, which could enhance its horizontal transmission from male host to other insects.
In conclusion, we demonstrated that triple infection of Wolbachia is more advantageous than single or non-infection in H. magnanima, which confirmed the prediction by Frank (1998). Our results imply that the Wolbachia interactions or triple infection plays a mutualistic role in females but a potentially parasitic role in males to maximise the transmission efficiency. The various strategies utilised by Wolbachia depending on the sex of the host and transmission route were revealed using isogeneic host lines. However, we have to take in to account the possibility that the genetic diversity of hosts as well as Wolbachia within or between populations can diverge their interaction (Hornett et al. 2006, Fry et al. 2004; Dean 2006; Capobianco et al. 2018). The complex H. magnanima-Wolbachia as well as Wolbachia-Wolbachia interactions may contribute to the maintenance of Wolbachia coinfections in nature. Further investigations on traits in other H. magnanima populations with wHm-a, wHm-b, and wHm-c will need to understand the effects of the host’s genetic diversity on the interactions with Wolbachia.
Data archiving
References
Ahmed MZ, Li SJ, Xue X, Yin XJ, Ren SX, Jiggins FM et al. (2015) The Intracellular bacterium Wolbachia uses parasitoid wasps as phoretic vectors for efficient horizontal transmission. PLoS Pathog 11:1–19
Arai H, Hirano T, Akizuki N, Abe A, Nakai M, Kunimi Y et al. (2019) Multiple infection and reproductive manipulations of Wolbachia in Homona magnanima (Lepidoptera: Tortricidae). Microb Ecol 77:257–266
Arai H, Lin SR, Nakai M, Kunimi Y, Inoue MN (2020) Closely related male-killing and nonmale-killing Wolbachia strains in the oriental tea tortrix Homona magnanima. Microb Ecol 79:1011–1020
Bailey NW, Zuk M (2008) Changes in immune effort of male field crickets infested with mobile parasitoid larvae. J Insect Physiol 54:96–104
Ballad JWO, Hatzidakis J, Karr TL, Kreitman M (1996) Reduced variation in Drosophila simulans mitochondrial DNA. Genetics 144:1519–1528
Birch LC (1948) The intrinsic rate of natural increase of an insect population. J Anim Ecol 17:15–26
Capobianco IIIF, Nandkumar S, Parker JD (2018) Wolbachia affects survival to different oxidative stressors dependent upon the genetic background in Drosophila melanogaster. Physiol Entomol 43:239–244
Danthanarayana W (1975) Factors determining variation in fecundity of the light brown apple moth, Epiphyas postvittana (Walker) (Tortricidae). Aust J Zool 23:309–319
Dean MD (2006) A Wolbachia-associated fitness benefit depends on genetic background in Drosophila simulans. Proc R Soc B 273:1415–1420
Deseo KV (1971) Study of factors influencing the fecundity and fertility of codling moth (Laspeyresia pomonella L., Lepidoptera, Tortricidae). Acta Phytopathol Hun 6:243–252
Dobson SL, Rattanadechakul W, Marsland EJ (2004) Fitness advantage and cytoplasmic incompatibility in Wolbachia single-and superinfected Aedes albopictus. Heredity 93:135–142
Duron O, Bouchon D, Boutin S, Bellamy L, Zhou L, Engelstädter J, Hurst GD (2008) The diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol 6:1–12
Engelstädter J, Telschow A, Hammerstein P (2004) Infection dynamics of different Wolbachia-types within one host population. J Theor Biol 231:345–55
Fleury F, Vavre F, Ris N, Fouillet P, Boulétreau M (2000) Physiological cost induced by the maternally-transmitted endosymbiont Wolbachia in the Drosophila parasitoid Leptopilina heterotoma. Parasitology 121:493–500
Frank SA (1998) Dynamics of cytoplasmic incompatibility with multiple Wolbachia infections. J Theor Biol 192:213–18
Frank SA, Hurst LD (1996) Mitochondria and male disease. Nature 383:224–224
Fry AJ, Palmer MR, Rand DM (2004) Variable fitness effects of Wolbachia infection in Drosophila melanogaster. Heredity 93:379–389
Gómez-Valero L, Soriano-Navarro M, Pérez-Brocal V, Heddi A, Moya A, García-Verdugo JM, Latorre A (2004) Coexistence of Wolbachia with Buchnera Aphidicola and a secondary symbiont in the aphid Cinara cedri. J Bacteriol 186:6626–33
Hoffmann AA, Turelli M, Harshman LG (1990) Factors affecting the distribution of cytoplasmic incompatibility in Drosophila simulans. Genetics 126:933–948
Hornett EA, Charlat S, Duplouy AMR, Davies N, Roderick GK, Wedell N et al. (2006) Evolution of male-killer suppression in a natural population. PLoS Biol 4:1643–1648
Hough JA, Pimentel D (1978) Influence of host foliage on development, survival, and fecundity of the gypsy moth. Environ Entomol 7:97–102
Ikeda T, Ishikawa H, Sasaki T (2003) Infection density of Wolbachia and level of cytoplasmic incompatibility in the Mediterranean flour moth, Ephestia kuehniella. J Invertebr Pathol 84:1–5
Ishii T, Nakai M, Okuno S, Takatsuka J, Kunimi Y (2003) Characterization of Adoxophyes honmai single-nucleocapsid nucleopolyhedrovirus: morphology, structure, and effects on larvae. J Invertebr Pathol 83:206–214
Kondo N, Shimada M, Fukatsu T (2005) Infection density of Wolbachia endosymbiont affected by coinfection and host genotype. Biol Lett 1:488–491
Lu P, Bian G, Pan X, Xi Z (2012) Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl Trop D 6:1–8
Maia AHN, Luiz AJB, Campanhola C (2000) Statistical inference on associated fertility life table parameters using jackknife technique: computational aspects. J Econ Entomol 93:511–518
Mazzetto F, Gonella E, Alma A (2015) Wolbachia infection affects female fecundity in Drosophila suzukii. Bull Insectol 68:153–157
Meyer JS, Ingersoll CG, McDonald LL, Boyce MS (1986) Estimating uncertainty in population growth rates: jackknife vs. bootstrap techniques. Ecology 67:1156–1166
Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, Hedges LM et al. (2009) A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 139:1268–1278
Mouton L, Henri H, Bouletreau M, Vavre F (2006) Effect of temperature on Wolbachia density and impact on cytoplasmic incompatibility. Parasitology 132:49–56
Narita S, Nomura M, Kageyama D (2007) Naturally occurring single and double infection with Wolbachia strains in the butterfly Eurema hecabe: transmission efficiencies and population density dynamics of each Wolbachia strain. FEMS Microb Ecol 61:235–245
Pigeault R, Braquart-Varnier C, Marcadé I, Mappa G, Mottin E, Sicard M (2014) Modulation of host immunity and reproduction by horizontally acquired Wolbachia. J Insect Physiol 70:125–133
Rancès E, Ye YH, Woolfit M, McGraw EA, O´Neill SL (2012) The relative importance of innate immune priming in Wolbachia-mediated dengue interference. PLoS Pathog 8:e1002548. https://doi.org/10.1371/journal.ppat.1002548
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Stevanovic AL, Arnold PA, Johnson KN (2015) Wolbachia -mediated antiviral protection in Drosophila larvae and adults following oral infection. Appl Environ Micro 81:8215–8223
Takamatsu T, Arai H, Abe N, Nakai M, Kunimi Y, Inoue MN (2021) Coexistence of two male-killers and their impact on the development of oriental tea tortrix Homona magnanima. Microb Ecol 81:193–202
Takehana A, Katsuyama T, Yano T, Oshima Y, Takada H, Aigaki T et al. (2002) Overexpression of a pattern-recognition receptor, peptidoglycan-recognition protein-LE, activates imd/relish-mediated antibacterial defense and the prophenoloxidase cascade in Drosophila larvae. Proc Natl Acad Sci USA 99:13705–13710
Takatsuka J, Okuno S, Ishii T, Nakai M, Kunimi Y (2010) Fitness-related traits of entomopoxviruses isolated from Adoxophyes honmai (Lepidoptera: Tortricidae) at three localities in Japan. J Invertebr Pathol 105:121–131
Teixeira L, Ferreira Á, Ashburner M (2008) The Bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol 6:e1000002. https://doi.org/10.1371/journal.pbio.1000002
Thomas P, Kenny N, Eyles D, Moreira LA, O´Neill SL, Asgari S (2011) Infection with the wMel and wMelPop strains of Wolbachia leads to higher levels of melanization in the hemolymph of Drosophila melanogaster, Drosophila simulans and Aedes aegypti. Dev Comp Immunol 35:360–365
Tsuruta K, Wennmann JT, Kunimi Y, Inoue MN, Nakai M (2018) Morphological properties of the occlusion body of Adoxophyes orana granulovirus. J Invertebr Pathol 154:58–64
Turelli M, Hoffmann AA (1991) Rapid spread of an inherited incompatibility factor in California Drosophila. Nature 353:440–442
Vautrin E, Vavre F (2009) Interactions between vertically transmitted symbionts: cooperation or conflict? Trends Microbiol 17:95–99
Vavre F, Fleury F, Lepetit D, Fouillet P, Boulétreau M (1999) Phylogenetic evidence for horizontal transmission of Wolbachia in host- parasitoid associations. Mol Biol Evol 16:1711–1723
Vollmer J, Schiefer A, Schneider T, Jülicher K, Johnston KL, Taylor MJ et al. (2013) Requirement of lipid II biosynthesis for cell division in cell wall-less Wolbachia, endobacteria of arthropods and filarial nematodes. Int J Med Microbiol 303:140–149
Voronin D, Guimarães AF, Molyneux GR, Johnston KL, Ford L, Taylor MJ (2014) Wolbachia lipoproteins: abundance, localization and serology of Wolbachia peptidoglycan associated lipoprotein and the Type IV Secretion System component, VirB6 from Brugia malayi and Aedes albopictus. Parasite Vector 7:462
Watanabe M, Miura K, Hunter MS, Wajnberg E (2011) Superinfection of cytoplasmic incompatibility-inducing Wolbachia is not additive in Orius strigicollis (Hemiptera: Anthocoridae). Heredity 106:642–648
Weeks AR, Turelli M, Harcombe WR, Reynolds KT, Hoffmann AA (2007) From parasite to mutualist: rapid evolution of Wolbachia in natural populations of Drosophila. PLoS Biol 5:0997–1005
Werren JH, Baldo L, Clark ME (2008) Wolbachia: Master manipulators of invertebrate biology. Nat Rev Microbiol 6:741–751
Xue X, Li S, Ahmed MZ, Barro PJ, Ren S, Qiu B (2012) Inactivation of Wolbachia reveals its biological roles in whitefly host. PLoS One 7:e48148. https://doi.org/10.1371/journal.pone.0048148
Zug R, Hammerstein P (2012) Still a host of hosts for Wolbachia: analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS One 7:e38544. https://doi.org/10.1371/journal.pone.0038544
Zug R, Hammerstein P (2015) Wolbachia and the insect immune system: what reactive oxygen species can tell us about the mechanisms of Wolbachia-host interactions. Front Microbiol 6:1–16
Acknowledgements
We thank Dr. T. Hiraoka (Tokyo University of Agriculture and Technology, Tokyo, Japan) for teaching the method to evaluate PO activity and Dr. S. Okuno (Arysta Lifescience Corporation, Tokyo, Japan) for providing the BotaniGard wettable powder. We also thank anonymous reviewers for their valuable suggestions.
Author information
Authors and Affiliations
Contributions
MU conducted experiments, data analysis, and prepared the manuscript. HA contributed to the discussion and revision of the manuscript. KM conducted data analysis. MN contributed to discussion. MNI supervised all experiments, prepared the manuscript, and contributed to the discussion.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Associate editor: Darren Obbard.
Supplementary information
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ueda, M., Arai, H., Masaike, K. et al. Distinct effects of three Wolbachia strains on fitness and immune traits in Homona magnanima. Heredity 130, 22–29 (2023). https://doi.org/10.1038/s41437-022-00574-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41437-022-00574-6
- Springer Nature Switzerland AG