
Abstract
Reactive oxygen species (ROS) and ROS-induced oxidative stress are associated with prostate cancer (PCa) development and castrate-resistant tumor progression. This is in part through the activation of the androgen receptor (AR) signaling. However, the molecular underpinning of ROS to activate AR remains poorly understood. Here, we report that the thioredoxin domain-containing 9 (TXNDC9) is an important regulator of ROS to trigger AR signaling. TXNDC9 expression is upregulated by ROS inducer, and increased TXNDC9 expression in patient tumors is associated with advanced clinical stages. TXNDC9 promotes PCa cell survival and proliferation. It is required for AR protein expression and AR transcriptional activity under oxidative stress conditions. Mechanistically, ROS inducers promote TXNDC9 to dissociate from PRDX1, but enhance a protein association with MDM2. Concurrently, PRDX1 enhances its association with AR. These protein interaction exchanges result in not only MDM2 protein degradation, but also PRDX1 mediated AR protein stabilization, and subsequent elevation of AR signaling. Blocking PRDX1 by its inhibitor, Conoidin A (CoA), suppresses AR signaling, PCa cell proliferation, and xenograft tumor growth even under androgen-deprived conditions. These tumor-suppressive effects of CoA were further strengthened when in combination with enzalutamide treatment. Together, these studies demonstrate that the TXNDC9-PRDX1 axis plays an important role for ROS to activate AR functions. It provides a proof-of-principle that co-targeting AR and PRDX1 may be more effective to control PCa growth.
Similar content being viewed by others


Introduction
The androgen receptor (AR) signaling is critical to prostate cancer (PCa) development and tumor progression [1], since the first-line treatment for metastatic PCa is androgen-deprivation therapy (ADT) that either blocks androgen synthesis or prevents androgens from activating the AR [2]. Although ADT is initially effective, these tumors will inevitably resume the AR signaling and progress into the lethal castrate-resistant prostate cancer (CRPC) stage [3,4,5]. Even though new generation anti-androgens (e.g., enzalutamide (Enz) and abiraterone) that target the ligand-binding domain (LBD) of AR have shown improved patient survival further strengthening the rationale that more potent inhibitors to block the AR signaling would benefit CRPC patients, resistant to Enz and abiraterone still occur [6]. These findings indicate that alternative therapeutic strategies to block AR signaling other than targeting AR LBD may provide further improvement of patient survival.
Elevated reactive oxygen species (ROS) had been detected in almost all cancers [7, 8]. ROS such as superoxide, hydroxyl free radicals, and non-radical molecules can give rise from endoplasmic reticulum (ER) stress, hypoxia, and metabolic defects within tumors [8]. However, cancer cells are more tolerant of ROS than benign cells by expressing increased levels of antioxidant proteins to detoxify from ROS. While excessive ROS would cause cell death [9], chronic and low doses of ROS had been shown to promote PCa cell growth and tumor progression by stimulating the survival and proliferating signaling cascades such as mitogen-activated protein kinases and Akt kinase pathways [10]. ROS-induced DNA mutations can also activate oncogenes to contribute to carcinogenesis and tumor progression [11, 12]. Furthermore, ROS had been shown to regulate AR-targeted transcription factors (Twist1, NF-κB, and c-myc) [13,14,15] or AR coactivators (Hsp27 and STAT3) [16, 17], implying that ROS may activate AR signaling that plays a significant role to promote prostate tumor progression. However, the molecular mechanisms by which ROS modulates AR signaling are not fully understood.
Scavenging of ROS in cancer cells is mediated by a set of “antioxidant” factors that prevent oxidative damage. These factors include glutathione peroxidase, peroxiredoxins, NADPH, thioredoxin, glutaredoxin, and thioredoxins (TRXs). TRXs are multifunctional, and ubiquitously expressed proteins that maintain cellular redox balance [18]. It is therefore not surprising that overexpression and hyper activities of TRX members were frequently observed in various human malignancies [19, 20]. Emerging evidence indicates that these TRXs can also function beyond being antioxidants. We and others have reported that the TRX family members, thioredoxin domain-containing protein 5 (TXNDC5) [21] and thioredoxin-1 (TRX1) [22], contribute to PCa progression by activating AR or protecting against AR-induced redox stress. Blocking TRX1 activity by its inhibitor PX-12 had also been reported to enhance ROS production, resulting in upregulation of AR expression in PCa under androgen-deprived condition, further supporting the idea that ROS can enhance the AR signaling during prostate tumor progression [22].
The thioredoxin domain-containing protein 9 (TXNDC9), also called APACD or PhLP3, is a member of TRX family. It has a central domain homologous to thioredoxin [23] implicating that it functions to sustain the redox state of cells. From the GEO38643 database, we found that increased TXNDC9 expression was more dramatic in PCa cells under ROS inducer treatments than other TRX family members [24], suggesting that TXNDC9 may play a more prominent role in maintaining redox balance in PCa cells. In addition, gene amplification of TXNDC9 was also reported in hepatocellular carcinoma [25]. TXNDC9 promotes hepatocellular carcinoma cell proliferation through MYC and FoxA1 [26, 27] signaling, both of which were known for their impacts on the AR signaling in PCa. These findings together implied that TXNDC9 may not only detoxify ROS, but also stimulate AR signaling in PCa. In this study, we demonstrate for the first time that TXNDC9 mediates ROS induced AR activation in contributing to PCa progression. This TXNDC9 action is mediated through a decreased association of TXNDC9 from PRDX1 but an increased association of TXNDC9 with MDM2 protein, resulting in MDM2 protein degradation. Concurrently, PRDX1 strengthens its protein interaction with AR protein to increase its protein stabilization and elevated AR signaling. Blocking PRDX1 alone or in combination with AR inhibition strongly suppresses PCa cell growth and xenograft progression, emphasizing that co-targeting PRDX1 and AR may provide improved PCa patient survival.
Results
Upregulation of TXNDC9 can be induced by ROS inducer and is associated with PCa at advanced clinical stage
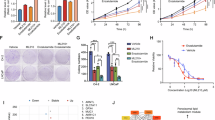
Previous gene studies (GEO 38643 database [24]) had shown that TXNDC9 is one of the most significantly upregulated TRX family members by the ROS inducer, tunicamycin (TM) (Fig. 1a). Our results showed consistent results that TM induced TXNDC9 mRNA levels in both LNCap and PC3 cell models (Fig. 1b, c). Furthermore, TXNDC9 protein levels were observed to be significantly upregulated by TM or thapsigargin (TG) in LNCap, VCap, and C4-2B cell models in both time- and dose-dependent manners (Fig. 1d, e). In contrast, no visible changes were observed in benign prostatic epithelial RWPE-1 cells treated with TM or TG (Fig. S1a, b). These results support the idea that PCa cells are more reactive than being prostate cells to express TXNDC9 in response to ROS exposure.

Upregulation of TXNDC9 can be induced by ROS inducer and is associated with PCa at advanced clinical stage. a Analysis of Trx family member expression in GSE38643. Trx family member expression was validated by real-time qPCR in LNCap (b) and PC3 cells (c). Cells were treated with TM for the indicated amount of time. d, e TXNDC9 protein levels were measured by western blot in LNCap, VCap, and C4-2B cells following incubation with TM or TG at the indicated doses for 12 h (d) or various amounts of time (e). f The expression of TXNDC9 compared in primary PCa samples and CRPC samples in GSE32269. g, h TXNDC9 expression in PCa subgroups in GSE21032 with different pathological stage (g) and Gleason scores (h). i Representative images of immunohistochemical staining of TXNDC9 in the Qilu cohort with benign prostatic hyperplasia (BPH), localized PCa, and castration-resistant prostate cancer (CRPC). j Percentages of different TXNDC9 expression (out of 100%) for localized PCa and CRPC samples. *P < 0.05, **P < 0.01, based on Student’s t test
To investigate whether TXNDC9 expression is also upregulated in prostate tumors in patients, we first analyzed TXNDC9 mRNA levels using the published dataset (GSE32269). TXNDC9 expression was significantly higher in CRPC than primary PCa tissues (Fig. 1f). TXNDC9 mRNA levels were positively correlated with clinical pathology stage (Fig. 1g) and Gleason scores (Fig. 1h). To further confirm these findings and measure TXNDC9 protein expression in patient PCa samples, we performed immunohistochemistry on tissue microarrays (TMA). Totally 177 tumor cores were stained with TXNDC9 antibody. These tissue cores include benign prostatic hyperplasia (BPH) tissues (n = 20), clinically localized PCa cases (n = 132), and CRPC cases (n = 25). Representative immunohistochemistry images are presented (Fig. 1i). We found that TXNDC9 localized mainly in the cytoplasmic compartment and its expression level was higher in PCa tissues than that in BPH. Furthermore, 92 (69.7%) of localized tumors showed negative or weak staining (scores 0 and 1), and only 40 (30.3%) had moderate or strong staining (scores 2 and 3). In contrast, 18 (72.0%) CRPC cases showed moderate to strong, whereas only 7 (28.0%) showed negative or weak staining of TXNDC9 (Fig. 1j). Together, both gene microarray studies and our IHC studies consistently indicate that upregulation of TXNDC9 is associated with PCa progression to advanced clinical stages.
TXNDC9 promotes PCa cell growth and tumor progression
To evaluate the biological functions of TXNDC9 in PCa cells, we first designed two siRNAs, both of which can significantly depleted TXNDC9 expression (Fig. S2a). Under basal ROS conditions where ROS inducer was absent, TXNDC9 silencing resulted in reduced cell viability in LNCap cells (Fig. S2b). However, no visible changes were observed in RWPE-1 cells when transfected with si-TXNDC9 or overexpression TXNDC9 (Fig. S2c). Before performing the functional assays in the presence of the ROS inducer, we first analyzed the effects of TM or TG on AR activity and cell viability. Cell proliferation was inhibited by treatment with TM (1 ng/mL) or TG (10uM) (Fig. S2d), and cell death did not differ significantly when compared with the controls. In addition, TM (1 ng/mL) or TG (10 uM) exposure induced the expression of PSA mRNA in LNCap cells (Fig. S2e). When LNCap cells were exposed to ROS inducers TM and TG at 1 ng/ml and 10 uM, respectively, cell viability was also reduced (Fig. 2a). However, these inhibitory effects of TM and TG were further enhanced by TXNDC9 silencing, but compromised by TXNDC9 overexpression (Fig. 2b). We showed that both TM and TG increased LNCap cell apoptosis rates measured by Annexin X staining and by BAX expression, which impacts were further enhanced by TXNDC9 silencing but reduced by TXNDC9 overexpression (Fig. 2c, d and S2f). Consistent with the effects in PCa cells, the viability decreased in RWPE-1 cells in the present with TM or TG treatment (Fig. S2g). Importantly, the inhibitory effects of ROS inducers on cell viability could not hardly be disturbed by silencing TXNDC9 (Fig. S2h). These findings were consistent to our LNCap xenograft studies showing that gain-of-function of TXNDC9 promoted tumor growth (Fig. 2e). LNCap-TXNDC9 tumors grew faster than LNCap-Ctrl (83.2 mm3/week vs 40.3 mm3/week). These results indicated that TXNDC9 promotes prostate tumor growth through enhancing PCa cell survival and proliferation.

TXNDC9 promotes PCa cell growth and tumor progression. a LNCap cells were transfected with siTXNDC9 or a negative control (siNC). Cell viability was assessed by cck-8 assay after treatment with TM (1 ng/mL) or TG (10uM) at different time points. b LNCap cells were introduced with TXNDC9 (ectopic overexpression TXNDC9) or empty plasmids (vector). Cell viability was assessed by cck-8 assay after treatment with TM (1 ng/mL) or TG (10uM) at different time periods. c, d After silencing or overexpressing TXNDC9 in LNCap cells as described in A and B and treated with TM or TG for 12 h, rate of cell apoptosis was measured using flow cytometry. e Subcutaneous xenograft assay was used to monitor tumor growth formed by LNCap-Vector and LNCap-TXNDC9 cells in vivo. f TXNDC9 protein levels in LNCap were measured by western blot following androgen-deprivation treatment at different time periods. g LNCap cells were introduced with TXNDC9 or its control and then cultured under androgen- deprivation conditions. Cell viability was measured by cck-8 assays. h C4-2B cells with stable expression of Vector or TXNDC9 subcutaneously injected into nude mice. The tumor volume was measured in nude mice with castration. *P < 0.05, **P < 0.01, ***P < 0.001, based on Student’s t test
In addition, it is observed that silencing TXNDC9 could inhibit cell viability (Fig. S2i) and proliferation (Fig. S2j) in hepatocellular carcinoma cell line HepG2 and cervical cancer cell line HeLa, indicating it is also involved in tumor progression in cancers other than PCa. However, the exact role of TXNDC9 remains to be determined.
Since the AR signaling is critical for PCa cell survival and growth, we further showed that androgen depletion induced TXNDC9 protein expression in LNCP cells in a time-dependent manner (Fig. 2f). Conversely, TXNDC9 enhanced LNCap cell viability under the androgen depletion conditions (Fig. 2g). Furthermore, TXNDC9 enhanced C4-2B xenograft growth in castrated mice, with tumor volumes increased more rapidly at 80 mm3/week in C4-2B-TXNDC9 tumors as compared with C4-2B-control tumors at 32.3 mm3/week after castration surgery to the mice (Fig. 2h). Together, these results indicate that TXNDC9 promotes PCa cell/xenograft growth and counteracts ROS in suppressing PCa cell viability and enhancing cell apoptosis. More importantly, androgen deprivation conditions upregulate TXNDC9 expression, which can confer PCa cells accelerated proliferation under androgen deprived conditions, and castrate-resistant xenograft progression, implying its potential roles in regulating AR signaling.
TXNDC9 can activate AR signaling in PCa cells
To determine whether TXNDC9 regulates AR signaling, we observed that TXNDC9 overexpression increased, while TXNDC9 silencing reduced AR protein levels in both LNCap, VCap and C4-2B cancer cells (Fig. 3a). TXNDC9 induced AR expression resulted in more AR protein localization in nucleus as shown by nuclear protein lysis fragmentation assays (Fig. 3b). Moreover, the mRNA levels of AR target genes including PSA, KLK2, S100P, TMPRSS2, and FKBP5 were all downregulated by TXNDC9 silencing in LNCap cells under R1881 treatment (Fig. 3c). Consistent results were also obtained in immunoblotting assays that showed reduced PSA protein levels by TXNDC9 silencing (Fig. 3d). These results together indicate that TXNDC9 regulates AR protein expression and AR target transcription in the presence of androgen.

TXNDC9 can activate AR signaling in PCa cells. a Western blot analysis of TXNDC9 and AR expression with TXNDC9 overexpression or knockdown in LNCap, VCap and C4-2B cells. b Cytoplasmic and nuclear AR protein levels were analyzed in siTXNDC9- or siNC -transfected LNCap, VCap and C4-2B cells with or without R1881 stimulation under hormone-deprived condition. GAPDH and Lamin A/C were used as cytoplasmic and nuclear protein-loading controls, respectively. c SiTXNDC9- or siNC -transfected LNCap cells were stimulated with R1881 or ethanol for16 h. AR target gene (PSA, KLK2, TMPRSS2, FKBP5, and S100P) expression was analyzed by real-time qPCR. d Western blot analyses were performed comparing PSA protein expression levels in siTXNDC9 LNCap cells in the absence or presence of R1881 (10 nM). e AR protein level was measured by western blot in LNCap and C4-2B cells transfected with siTXNDC9 or siCtrl prior to TM (ng/mL) or TG (uM) treatment. f Changes of nuclear/cytoplasmic expression of AR was assayed by western blot in siTXNDC9- or siNC-transfected LNCap and C4-2B cells following treatment with TM (1ng/mL). g AR target genes expression (PSA and KLK2) was measured by real-time qPCR in LNCap cells after siTXNDC9 prior to TM or TG treatment for 12 h. *P < 0.05, **P < 0.01, ***P < 0.001, based on Student’s t test
Under oxidative stress conditions, we first showed that either TM or TG induced AR protein expression (Fig. S3a) and PSA mRNA levels (Fig. S3b) in a dose-dependent manner. When cells were treated with cycloheximide (CHX) to block protein synthesis, we observed that TM treatment increased AR protein stability resulting in slower AR protein decay comparing vehicle-treated cells (Fig. S3c). To test whether TXNDC9 is required for ROS inducers in controlling AR protein stability, LNCap and C4-2B cells were treated with control or TXNDC9 siRNA in the presence or absence of TM or TG. We showed that TM and TG lost their capability to induce AR protein levels and AR protein nuclear localization when TXNDC9 was depleted in the cells (Fig. 3e, f). TM and TG induced PSA and KLK2 mRNA levels were also diminished by TXNDC9 silencing (Fig. 3g). These results indicated that TXNDC9 is essential for ROS to enhance AR protein stability. However, we did not identify any protein interaction between TXNDC9 and AR (Fig. S3d), suggesting that TXNDC9 regulation of AR protein stability is indirect, and involves other protein factors.
TXNDC9 coordinates with PRDX1 to regulate the activity of AR signaling in PCa
To define the molecular mechanism by which TXNDC9 regulates AR protein stability, we have performed GST-pull down assays followed by Mass Spectrometry to profile the interactome of TXNDC9 in PCa cells (Fig. S4a). These studies led to the discovery of PRDX1 and MDM2 are the two major interacting proteins of TXNDC9. To confirm these findings, we applied co-immunoprecipitation assays using LNCap and VCap cell models and showed that TXNDC9, PRDX1, and MDM2 form a protein complex (Fig. S3b). Interestingly, no physical interaction between TXNDC9 and PRDX1 was detected by Co-IP and IF analysis in HeLa and HepG2 cells (Figs. S3c and S4d). PRDX1 was known as a family member of peroxidases that involves in eliminating peroxides [28]. PRDX1 forms into polymers and functions as a molecular chaperone upon exposure to oxidative stress and heat shock [29, 30]. In the presence of hypoxia or reoxygenation, PRDX1 could act as a chaperone to interact and activate AR independent of its antioxidant activity in PCa [31]. Interestingly, MDM2 was an E3 ubiquitin ligase that controls AR protein stability [32]. These findings together prompt us to focus on characterizing the TXNDC9–PRDX1–MDM2 complex in regulating AR signaling under oxidative stress conditions in PCa cells. In consistent to its role as an antioxidant, overexpression of PRDX1 reduced ROS level in LNCap cells under basal levels of ROS conditions. However, such PRDX1 action was abolished when cells were exposed to oxidative stress inducers (Fig. 4a). Instead, PRDX1 reduced its association with TXNDC9 but increased its association with AR in both LNCap, VCap and C4-2B cells (Fig. 4b). These results suggest that there likely exists a functional switch of PRDX1 from an antioxidant to an AR chaperone when cells are treated with an ROS inducer. Indeed, PRDX1 can prolong AR protein decay induced by CHX when LNCap and C4-2B cells are under TM treatment (Fig. 4c). Knocking down PRDX1 reduced, while PRDX1 overexpression enhanced AR protein levels in the presence of TM (Fig. 4d). To further evaluate the impacts of PRDX1 on AR functions, we showed that PRDX1 did not increase the transcription of these AR targeted PSA and KLK2 genes unless ROS inducer TM was applied to the cells (Fig. 4e). Furthermore, we showed that reduced AR activity by TXNDC9 knockdown was attenuated when PRDX1 was introduced into the cells (Fig. 4f, left) in the presence of TM. Vice versa, TXNDC9 induced AR activation was abolished when PRDX1 was depleted (Fig. 4f, right). Together, these results indicated that oxidative stress conditions stimulate PRDX1 disassociation from TXNDC9 but enhance PRDX1 association with AR. These dynamic protein interaction changes are required to stabilize AR proteins and regulate AR transcriptional activity.

TXNDC9 coordinates with PRDX1 to regulate the activity of AR signaling in PCa. a LNCap cells were transfected with PRDX1 for 36 h and then incubated with TM (1 ng/mL) for another 6 h. ROS levels were measured using a FACStar flow cytometer. b LNCap,VCap, and C4-2B cells were incubated with TM (1 ng/mL). The cell lysates were subjected to immunoprecipitation with either preimmune rabbit IgG or anti-PRDX1 antibody and the resulting precipitates were examined by immunoblot analysis with anti-TXNDC9 and anti-AR antibodies. c PRDX1 was overexpressed in LNCap and C4-2B cells for 36 h, followed by incubation with TM (1 ng/mL) for another 6 h. AR protein levels were examined by western blot in the absence or presence of CHX. d PRDX1 was overexpressed or silenced in LNCap and C4-2B cells, AR level was examined by western blot. e LNCap cells were transfected with PRDX1 for 36 h and then treated with TM (1 ng/mL) for 6 h. AR target gene (PSA) expression was measured by real-time qPCR. f After overexpressing or silencing TXNDC9 for 24 h, LNCap cells were transfected with PRDX1 or siPRDX1 for another 24 h. The transfected cells were then incubated with TM (1 ng/mL) for 6 h. The mRNA levels of AR target gene PSA were measured by real-time qPCR. *P < 0.05, **P < 0.01, ***P < 0.001, based on Student’s t test
PRDX1 regulates TXNDC9 interaction with MDM2 and MDM2 protein expression
To study the role of MDM2 in the TXNDC9–PRDX1–MDM2 complex, we performed immunoprecipitation (IP) assays to show that oxidative stress increased MDM2 association with TXNDC9, but reduced interaction with AR (Fig. 5a) in PCa cells. Both oxidative inducers TM and TG dowregulated MDM2 protein levels in a dose-dependent manner (Fig. 5b). In addition, TXNDC9 overexpression reduced, while TXNDC9 knockdown increased MDM2 proteins (Fig. 5c). Oxidative stress downregulated MDM2 protein levels, which effects were abolished when TXNDC9 was depleted (Fig. 5d). These results indicated that TXNDC9 interacts with MDM2 and downregulates MDM2 protein expression.

PRDX1 regulates TXNDC9 interaction with MDM2 and MDM2 protein expression. a LNCap, VCap, and C4-2B cells were untreated or treated with TM (1 ng/mL) for 6 h. Cell lysates were subjected to immunoprecipitation with MDM2 antibody. The resulting precipitates were examined by immunoblot analysis with antibodies to TXNDC9 and AR. b MDM2 expression was measured by western blot in LNCap, VCap and C4-2B cells treated with TM (ng/mL) or TG (uM) for 12 h. c MDM2 protein level was measured by western blot in PCa cells with TXNDC9 knockdown or overexpression. d LNCap, VCap, and C4-2B cells were transfected with siTXNDC9 for 36 h, followed by incubation with TM(1 ng/mL) for another 6 h. AR protein levels were examined by western blot. e, f LNCap and C4-2B cells were transfected with siPRDX1/siMDM2 for 48 h and then untreated or treated with TM (1 ng/mL) for 6 h. Cell lysates were subjected to immunoprecipitation with MDM2/PRDX1 antibody. The resulting precipitates were examined by immunoblot analysis with antibodies to TXNDC9 and AR. g LNCap and C4-2B cells were transfected with increasing amounts of plasmids encoding Flag-PRDX1 (0, 2, or 4 ug) in the presence of TM (1 ng/mL). The amount of DNA in each transfection was kept constant by the addition of an appropriate amount of empty vector. Precipitated proteins by anti-AR antibody were analyzed by immunoblotting using anti-PRDX1 and anti-MDM2 antibodies. *P < 0.05, **P < 0.01, ***P < 0.001, based on Student’s t test
Since MDM2 mediates AR protein degradation through the ubiquitin-proteasome pathway [32], TXNDC9 may possibly prevent AR from protein degradation through MDM2. To test this hypothesis, we performed IP assays with MDM2 antibody to show that under oxidative stress conditions MDM2 protein interaction with AR was downregulated in both LNCap, VCap and C4-2B cells (Fig. 5a). These findings were consistent to our observations that oxidative stress enhances AR protein expression. These oxidative stress effects required the presence of PRDX1, since PRDX1 depletion increased MDM2 interaction with AR when comparing to control siRNA treatment condition (Fig. 5e). Concurrently, MDM2 protein interaction with TXNDC9 was increased by oxidative stress condition, which changes were abolished by PRDX1 deletion. Our IP assays with the PRDX1 antibody indicated that oxidative stress enhanced PRDX1 interaction with AR regardless of MDM2 depletion (Fig. 5f), which results were consistent to that oxidative stress stimulates MDM2 disassociation with AR protein. To examine whether PRDX1 competes with MDM2 for interaction with AR, increasing amounts of plasmid expressing Flag-PRDX1 was transfected into LNCap and C4-2B cells. The data shows that the binding between MDM2 with AR decreased steadily as PRDX1 expression increased in PCa cells in the presence of TM. Collectively, these results suggest that PRDX1 could compete with MDM2 to bind to AR (Fig. 5g), which may facilitate the interaction between TXNDC9 and MDM2, and subsequent protein degradation of MDM2.
PRDX1 inhibitor is effective to block PCa cell growth and tumor progression
Based on the results from above, we hypothesized that TXNDC9 may promote PCa progression in a PRDX1-dependent manner. We have applied Conoidin A (CoA), a PRDX1 inhibitor [33, 34] to this hypothesis. CoA could significantly suppress TXNDC9 induced expression of the AR target genes, KLK2 and PSA (Fig. 6a). CoA also compromise TXNDC9 enhanced cell viability of LNCap cells under basal ROS conditions (Fig. S5a). When nude mice carrying LNCap-Vector or LNCap-TXNDC9 xenografts were treated with CoA, we found that the impacts of TXNDC9 in promoting tumor growth were significantly diminished by CoA (Fig. 6b). Further analysis showed that combining CoA treatment with TXNDC9 knockdown showed stronger suppression to LNCap cell viability (Fig. S5b). Importantly, CoA showed stronger inhibition to cell viability under androgen-deprived conditions (Fig. 6c, d). Consistently, we observed that CoA treatment could also significantly inhibit cell proliferation, and enhance PCa cell apoptosis even when TXNDC9 is overexpressed in CRPC model C4-2B cells, a condition frequently occurred in patient prostate tumors (Fig. 6e, left and Fig. 6f, left). In addition, CoA treatment showed stronger suppression to PCa cell proliferation and increase apoptosis when TXNDC9 was knocked down (Fig. 6e, right and Fig. 6f, right). To evaluate the effects of CoA to C4-2B xenograft models, we showed that (Fig. 6g) CoA treatment reduced tumor growth more effectively in castrated mice as compared with ones without castration. To further support the involvement of PRDX1 in TXNDC9-mediated CRPC progression, C4-2B overexpressed with TXNDC9 were treated with CoA under androgen-deprived conditions. The increase of cell viability induced by TXNDC9 overexpression could be attenuated by CoA treatment. Similar inhibition was also observed with treatment of the inhibitor of AR, Enz. More importantly, CoA in combination with Enz could reduce the cell viability more significantly when compared with CoA or Enz treatment alone (Fig. 6h). These data suggest that the combined inhibition of AR and PRDX1 synergistically delays PCa progression, especially in cases with high TXNDC9 expression. Taken together, our results suggest that blocking PRDX1 might represent a novel treatment strategy for PCa.

PRDX1 inhibitor is effective to block PCa cell growth and tumor progression. a mRNA levels of AR target genes (PSA and KLK2) were analyzed by real-time qPCR after silencing or overexpressing TXNDC9 in LNCap with or without CoA (8uM) treatment for 12 h. bPhotographs of tumor growths with LNCap-Vector and LNCap-TXNDC9 implantation and treatment with DMSO or CoA. The vector and TXNDC9 groups are the same as Fig. 2e. Cell proliferation was examined by cck-8 (c) and EdU (d) after overexpressing TXNDC9 in CoA (8uM)-treated LNCap under androgen-depletion conditions. For EdU assays, representative photographs were shown at original magnification: ×200. The proliferation rate was calculated by EdU-positive cells compared with total cell number. After the same treatment as in d, cell proliferation (e) and cell apoptosis (f) were examined by cck-8 and flow cytometry. g The tumor volume was measured in nude mice with castration as the indicated treatment. The vector and TXNDC9 groups are the same as Fig. 2h. h Cell viability was detected by cck-8 assay in C4-2B cells after introduction of TXNDC9 in prior to the addition of Enz plus CoA (8uM). i A putative model illustrating the role of TXNDC9 during PCa progression. ROS inducer promotes TXNDC9 expression, PRDX1 dissociates with TXNDC9 and activates AR signaling, and facilitates TXNDC9-mediated degradation of MDM2. *P < 0.05, **P < 0.01, ***P < 0.001, based on Student’s t test
Discussion
Cancer cells are characterized by enhanced ROS production [35]. Although excessive ROS leads to cell death, it can induce mutations in DNA, genomic instability and release of tumorigenic signals, which finally promoted cancer cell survival, proliferation, and adaptation to hypoxia, chemotherapy or radiation [8]. Recently, an increasing body of evidence suggested that ROS is closely associated with the initiation and progression of PCa [36]. In response to ADT, elevated ROS is also involved in CRPC progression [35, 37]. Importantly, AR is responsive to ROS in hydrogen peroxide-resistant LNCap cells. The increase of AR expression may be due to the protective effects of ROS on its mRNA or protein stability. In addition, AR signaling plays a central role for PCa and targeting its cofactors, such as P160 SRC (steroid receptor coactivator) family genes [38] or HSP90 [39], maybe effective for PCa treatment. Therefore, identifying key factors in maintaining ROS balance for AR activation would be helpful to elucidating the pathological mechanism of PCa and also providing novel treatment target.
In this study, it is observed that TXNDC9 overexpression predicts poor prognosis of PCa patients. Functionally, it could promote tumor growth even in castrated models. Importantly, the side effects of ROS inducer or ADT on PCa cell survival and proliferation could be compromised by overexpressing TXNDC9. These facts supported that TXNDC9 is required for PCa to cope with ADT-induced adverse atmosphere during disease progression. Recently, Samaranayake et al. described a protective role of TRX1 against ROS-associated cytotoxicity in PCa cells [22]. Intriguingly, pharmacologic TRX1 inhibition elevated ROS and AR levels. Meanwhile, AR depletion could mitigate both TRX1 inhibition-mediated ROS production and cell death. These facts further support the likely reciprocal regulation between AR and cellular ROS during PCa progression even though the exact mechanism remains obscure. Here, our study demonstrated that TXNDC9 is required for the activation of AR signaling in the presence of ROS inducer and TXNDC9 may behave as an effector for the crosstalk between ROS and AR.
PRDX1 belongs to the redox-regulating peroxiredoxin family and plays an anti-ROS role in its homodimer form [28]. However, its catalytic ability is relatively lower when compared with those of catalases or glutathione peroxidases and could be easily inactivated during the H2O2-catalytic process [29, 30]. Besides, we and other studies have confirmed that PRDX1 could form polymer and switch to a chaperone role upon exposure of oxidative stress or heat shock [29, 30]. It is been reported that, with the stimulation of hypoxia/reoxygenation, PRDX1 could interact with AR and enhance its activity in PCa [31]. Interestingly, similar effects of PRDX1 on AR was observed in PCa following detachment from TXNDC9 and PRDX1 could stabilized AR protein in turn. Of note, we identified MDM2, an E3 ubiquitin-protein ligase that promotes AR ubiquitination and degradation, as another interactor for TXNDC9. Detailed analysis showed that, besides the direct binding and activation of PRDX1 toward AR, PRDX1 also provides a paracrine avenue to activate AR signaling by promoting TXNDC9-mediated MDM2 degradation. These facts support that the dissociation of PRDX1 from TXNDC9 could therefore represent an upstream event to favor the subsequent activity of AR. PRDX1-based machinery involving TXNDC9 and MDM2 may be required for ROS to activate AR signaling, and can resist the detrimental effects of excessive ROS on cell survival and proliferation.
Our cellular and molecular experiments indicated that targeting PRDX1 may hold therapeutic merit for PCa treatment. CoA is a PRDX1 inhibitor that could inactivate two mammalian peroxiredoxin homologs (PrxI and PrxII) [33, 34]. As expected, CoA could efficiently inactivate AR signaling and decrease cell viability and tumor growth. Such therapeutic effect of CoA on PCa was more obvious under androgen-deprived and castrated condition. Furthermore, the combination of blocking TXNDC9-PRDX1 and AR signaling may be more beneficial for PCa treatment than using the AR inhibition strategy alone.
In conclusion, our data reveal a novel dynamic complex involving TXNDC9, MDM2, and PRDX1 that mediates ROS-induced activation of AR signaling. More importantly, blocking the crosstalk between ROS and AR using an PRDX1 inhibitor may provide a novel strategy for treating PCa.
Materials and methods
Patients and tissue specimens
A total of three TMA were constructed representing 20 patients with BPH and 132 clinically localized PCa patients who underwent radical prostatectomy as a primary therapy between 2012 and 2015 at Qilu Hospital of Shandong University (Jinan, China). Twenty-five PCa patients with CRPC treated by transurethral resection of the prostate were also included. Three cores (0.6 mm in diameter) were taken from each representative tumor focus and the morphology was confirmed by two pathologists (B.H., X.W.). This study was conducted in accordance with the International Ethical Guidelines for Biomedical Research Involving Human Subjects. This study protocol was approved by Shandong University Medical Research Ethics Committee according to the Declaration of Helsinki. The informed written consent was obtained from the patients.
Immunohistochemistry (IHC) and immunofluorescence (IF)
Immunohistochemistry and semiquantitative analysis of the staining intensity score were described as previously reported [40]. For immunofluorescence, proteins were detected using goat anti-mouse Alexa Fluor 647 and goat anti-rabbit Alexa Fluor 488 (Beyotime, P0176, P0191). The information of antibodies is summarized in Supplementary Table 1. Images were processed under a confocal microscope (FV3000, OLYMPUS, Tokyo, Japan).
Bioinformatics analysis
Datasets of GSE38643, GSE32269, and GSE21032 were downloaded from the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo).
Cell culture and reagents
LNCap and C4-2B were kindly provided by Dr Jidan Yu (Northwestern University, Chicago, USA) in 2015. VCap, 22RV1, benign prostatic cell line RWPE-1 were purchased from American Type Culture Collection (ATCC) (Rockville, MD, USA) between 2012 and 2015 and authenticated again by short tandem repeat analysis before and after our study. Hela and HepG2 cells were obtained from Wei Tang (Shandong University). All of the newly revived cells were tested free of mycoplasma contamination by Hoechst 33258 staining (Beyotime, Jiangsu, China). The information of reagents is summarized in Supplementary Table 2.
Transient transfection and viral transduction
Human MDM2, TXNDC9, and PRDX1 plasmids were purchased from Vigene (CH821314, CH840719, CH885311). siTXNDC9, siMDM2, siPRDX1, and negative controls were designed and synthetized by Ribobio (Guangzhou, China). The target sequences are listed in Supplementary Table 3. To avoid off-target effects, we co-transfection of two RNAs with better interference efficiency. The transfection procedure was performed as previously described [41]. Human Lenti-TXNDC9-EGF and its control Lenti-EGFP was obtained from Genecopoeia. To acquire a stable cell line, single-cell clonal isolates were selected by puromycin.
Cell viability
Cell viability was performed as previously described [41].
RNA isolation and qRT-PCR analysis
Total RNA was acquired from PCa cells using the Trizol reagent (Invitrogen,15596-018) according to the manufacturer’s instructions. RNA was reverse-transcribed with ReverTra Ace qPCR RT kit (Toyobo, PCR-311). RT-qPCR was performed using SYBR Green mix (Toyobo, QPK-201). The primer sequences are listed in Supplementary Table 4.
Western blot and IP assay
Western blot and IP assays were performed as described [41]. The information of antibodies is summarized in Supplementary Table 1. The cytoplasmic and nuclear proteins were extracted using the ProteoExtract subcellular proteome extraction kit (Millipore, 539791). For the protein stability assay, cells were treated with cyclohexane (CHX, 5ug/mL) for the indicated time points.
Flow cytometric analyses
Intracellular ROS was detected by 5- (and -6)-chloromethyl-2′,7′-dichlorofluorescein diacetate (AAT Bioquest, 16050) as described in the manufacturer’s protocol. The apoptosis assay was performed as previously described [42]. Both the apoptosis and ROS assay were generated and analyzed on a BD FACS Calibur Cytometer and the included software.
Identification of TXNDC9-binding proteins
LNCap cells were transfected with GST-TXNDC9 plasmids for 36 h and then treated with TM for 6 h. Total protein from cell lysates were incubated with GST magnetic-beads (Cell Signaling Technology, 11847) at 4 °C for overnight. The protein-bead complexes were washed four times with washing buffer containing 0.1% NP40. After boiling at 100 °C in the loading buffer, proteins were loaded onto SDS–PAGE gel and visualized with Coomassie blue R350. The potential interacting proteins (specific bands on the gel) were evaluated by mass spectrum analysis.
In vivo tumor formation
The experimental protocols were performed in accordance with the Ethical Animal Care and Use Committee of Shandong University. Five-week-old male athymic nude mice (nu/nu) were purchased from Weitonglihua Biotechnology (Beijing, China). We suspended 1 × 107 (n = 6/group) stable TXNDC9-overexpression LNCap or its control LNCap cells suspended in 100% Matrigel and then inoculated subcutaneously into the nude mice. Animals were randomized into a control or treatment group for intraperitoneal injection of PBS or CoA (10 mg/kg) every 3 days. Tumor size was measured every 3 days by Vernier calipers and tumor volume was calculated as V(mm3) = 0.5 × length × (width)2. At the end of experiments, mice were killed by sodium pentobarbital anesthesia injections (50 mg/kg) and tumors were excised and weighed. The CRPC model (n = 7/group) was studied as previously described [40].
Statistics analysis
GraphPad Prism 7 Software (GraphPad Software Inc.) was used for all statistical analysis unless otherwise specified. All results are presented as the mean and the standard error of the mean (SEM). P < 0.05 was considered statistically significant.
References
Fizazi K, Smith MR, Tombal B. Clinical development of darolutamide: a novel androgen receptor antagonist for the treatment of prostate cancer. Clin Genitourin Cancer. 2018;16:332–40.
Huggins C, Hodges CV. Studies on prostatic cancer: I. The effect of castration, of estrogen and of androgen injection on serum phosphatases in metastatic carcinoma of the prostate. 1941. J Urol. 2002;168:9–12.
Scher HI, Sawyers CL. Biology of progressive, castration-resistant prostate cancer: directed therapies targeting the androgen-receptor signaling axis. J Clin Oncol. 2005;23:8253–61.
Sharifi N. Mechanisms of androgen receptor activation in castration-resistant prostate cancer. Endocrinology. 2013;154:4010–7.
Visakorpi T, Hyytinen E, Koivisto P, Tanner M, Keinanen R, Palmberg C, et al. In vivo amplification of the androgen receptor gene and progression of human prostate cancer. Nat Genet. 1995;9:401–6.
Groner AC, Brown M. Role of steroid receptor and coregulator mutations in hormone-dependent cancers. J Clin Invest. 2017;127:1126–35.
Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nat Rev Cancer. 2011;11:85–95.
Gorrini C, Harris IS, Mak TW. Modulation of oxidative stress as an anticancer strategy. Nat Rev Drug Discov. 2013;12:931–47.
Schafer ZT, Grassian AR, Song L, Jiang Z, Gerhart-Hines Z, Irie HY, et al. Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment. Nature. 2009;461:109–13.
Sabharwal SS, Schumacker PT. Mitochondrial ROS in cancer: initiators, amplifiers or an Achilles’ heel? Nat Rev Cancer. 2014;14:709–21.
Martindale JL, Holbrook NJ. Cellular response to oxidative stress: signaling for suicide and survival. J Cell Physiol. 2002;192:1–15.
Leslie NR, Bennett D, Lindsay YE, Stewart H, Gray A, Downes CP. Redox regulation of PI 3-kinase signalling via inactivation of PTEN. EMBO J. 2003;22:5501–10.
Shiota M, Yokomizo A, Tada Y, Inokuchi J, Kashiwagi E, Masubuchi D, et al. Castration resistance of prostate cancer cells caused by castration-induced oxidative stress through Twist1 and androgen receptor overexpression. Oncogene. 2010;29:237–50.
Grad JM, Dai JL, Wu S, Burnstein KL. Multiple androgen response elements and a Myc consensus site in the androgen receptor (AR) coding region are involved in androgen-mediated up-regulation of AR messenger RNA. Mol Endocrinol (Baltim, Md). 1999;13:1896–911.
Gloire G, Legrand-Poels S, Piette J. NF-kappaB activation by reactive oxygen species: fifteen years later. Biochem Pharm. 2006;72:1493–505.
Zoubeidi A, Zardan A, Beraldi E, Fazli L, Sowery R, Rennie P, et al. Cooperative interactions between androgen receptor (AR) and heat-shock protein 27 facilitate AR transcriptional activity. Cancer Res. 2007;67:10455–65.
Yamamoto T, Matsuda T, Junicho A, Kishi H, Saatcioglu F, Muraguchi A. Cross-talk between signal transducer and activator of transcription 3 and estrogen receptor signaling. FEBS Lett. 2000;486:143–8.
Jacquot JP, Gelhaye E, Rouhier N, Corbier C, Didierjean C, Aubry A. Thioredoxins and related proteins in photosynthetic organisms: molecular basis for thiol dependent regulation. Biochem Pharm. 2002;64:1065–9.
Kakolyris S, Giatromanolaki A, Koukourakis M, Powis G, Souglakos J, Sivridis E, et al. Thioredoxin expression is associated with lymph node status and prognosis in early operable non-small cell lung cancer. Clin Cancer Res. 2001;7:3087–91.
Grogan TM, Fenoglio-Prieser C, Zeheb R, Bellamy W, Frutiger Y, Vela E, et al. Thioredoxin, a putative oncogene product, is overexpressed in gastric carcinoma and associated with increased proliferation and increased cell survival. Hum Pathol. 2000;31:475–81.
Wang L, Song G, Chang X, Tan W, Pan J, Zhu X, et al. The role of TXNDC5 in castration-resistant prostate cancer-involvement of androgen receptor signaling pathway. Oncogene. 2015;34:4735–45.
Samaranayake GJ, Troccoli CI, Huynh M, Lyles RDZ, Kage K, Win A, et al. Thioredoxin-1 protects against androgen receptor-induced redox vulnerability in castration-resistant prostate cancer. Nat Commun. 2017;8:1204.
Gaudet R, Bohm A, Sigler PB. Crystal structure at 2.4 angstroms resolution of the complex of transducin betagamma and its regulator, phosducin. Cell. 1996;87:577–88.
Guha P, Kaptan E, Gade P, Kalvakolanu DV, Ahmed H. Tunicamycin induced endoplasmic reticulum stress promotes apoptosis of prostate cancer cells by activating mTORC1. Oncotarget. 2017;8:68191–207.
Chen D, Zou J, Zhao Z, Tang X, Deng Z, Jia J, et al. TXNDC9 promotes hepatocellular carcinoma progression by positive regulation of MYC-mediated transcriptional network. Cell Death Dis. 2018;9:1110.
Parolia A, Cieslik M, Chu SC, Xiao L, Ouchi T, Zhang Y, et al. Distinct structural classes of activating FOXA1 alterations in advanced prostate cancer. Nature. 2019;571:413–8.
Xu B, Song B, Lu X, Kim J, Hu M, Zhao JC, et al. Altered chromatin recruitment by FOXA1 mutations promotes androgen independence and prostate cancer progression. Cell Res. 2019. [Epub ahead of print].
Rhee SG, Kang SW, Chang TS, Jeong W, Kim K. Peroxiredoxin, a novel family of peroxidases. IUBMB Life. 2001;52:35–41.
Kumsta C, Jakob U. Redox-regulated chaperones. Biochemistry. 2009;48:4666–76.
Jang HH, Lee KO, Chi YH, Jung BG, Park SK, Park JH, et al. Two enzymes in one; two yeast peroxiredoxins display oxidative stress-dependent switching from a peroxidase to a molecular chaperone function. Cell. 2004;117:625–35.
Park SY, Yu X, Ip C, Mohler JL, Bogner PN, Park YM. Peroxiredoxin 1 interacts with androgen receptor and enhances its transactivation. Cancer Res. 2007;67:9294–303.
Lin HK, Wang L, Hu YC, Altuwaijri S, Chang C. Phosphorylation-dependent ubiquitylation and degradation of androgen receptor by Akt require Mdm2 E3 ligase. EMBO J. 2002;21:4037–48.
Haraldsen JD, Liu G, Botting CH, Walton JG, Storm J, Phalen TJ, et al. Identification of conoidin a as a covalent inhibitor of peroxiredoxin II. Org Biomol Chem. 2009;7:3040–8.
Liu G, Botting CH, Evans KM, Walton JA, Xu G, Slawin AM, et al. Optimisation of conoidin A, a peroxiredoxin inhibitor. ChemMedChem. 2010;5:41–45.
Best CJ, Gillespie JW, Yi Y, Chandramouli GV, Perlmutter MA, Gathright Y, et al. Molecular alterations in primary prostate cancer after androgen ablation therapy. Clin Cancer Res. 2005;11:6823–34.
Shiota M, Yokomizo A, Naito S. Oxidative stress and androgen receptor signaling in the development and progression of castration-resistant prostate cancer. Free Radic Biol Med. 2011;51:1320–8.
Tam NN, Gao Y, Leung YK, Ho SM. Androgenic regulation of oxidative stress in the rat prostate: involvement of NAD(P)H oxidases and antioxidant defense machinery during prostatic involution and regrowth. Am J Pathol. 2003;163:2513–22.
Nakka M, Agoulnik IU, Weigel NL. Targeted disruption of the p160 coactivator interface of androgen receptor (AR) selectively inhibits AR activity in both androgen-dependent and castration-resistant AR-expressing prostate cancer cells. Int J Biochem Cell Biol. 2013;45:763–72.
Harashima K, Akimoto T, Nonaka T, Tsuzuki K, Mitsuhashi N, Nakano T. Heat shock protein 90 (Hsp90) chaperone complex inhibitor, radicicol, potentiated radiation-induced cell killing in a hormone-sensitive prostate cancer cell line through degradation of the androgen receptor. Int J Radiat Biol. 2005;81:63–76.
Wang L, Song G, Zhang X, Feng T, Pan J, Chen W, et al. PADI2-mediated citrullination promotes prostate cancer progression. Cancer Res. 2017;77:5755–68.
Feng T, Zheng L, Liu F, Xu X, Mao S, Wang X, et al. Growth factor progranulin promotes tumorigenesis of cervical cancer via PI3K/Akt/mTOR signaling pathway. Oncotarget. 2016;7:58381–95.
Liu F, Zhang W, Yang F, Feng T, Zhou M, Yu Y, et al. Interleukin-6-stimulated progranulin expression contributes to the malignancy of hepatocellular carcinoma cells by activating mTOR signaling. Sci Rep. 2016;6:21260.
Funding
This work was supported by the National Key Research And Development Program of China (No. 2018YFC0114703), the National Natural Science Foundation of China (grant nos. 81572544, 81772760, 81472417, 81672554), The Key Research and Development Project of Shandong (grant no. 2016GSF201166), The Shandong Taishan Scholarship (grant no. tsqn20161076).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Feng, T., Zhao, R., Sun, F. et al. TXNDC9 regulates oxidative stress-induced androgen receptor signaling to promote prostate cancer progression. Oncogene 39, 356–367 (2020). https://doi.org/10.1038/s41388-019-0991-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41388-019-0991-3
- Springer Nature Limited