
Abstract
ErbB3, a member of the ErbB family of receptor tyrosine kinases, is a potent activator of phosphatidyl inositol-3 kinase (PI3K) and mammalian target of rapamycin (mTOR) signaling, driving tumor cell survival and therapeutic resistance in breast cancers. In luminal breast cancers, ErbB3 upregulation following treatment with the antiestrogen fulvestrant enhances PI3K/mTOR-mediated cell survival. However, the mechanism by which ErbB3 is upregulated in fulvestrant-treated cells is unknown. We found that ErbB3 protein levels and cell surface presentation were increased following fulvestrant treatment, focusing our attention on proteins that regulate ErbB3 at the cell surface, including Nrdp1, NEDD4 and LRIG1. Among these, only LRIG1 correlated positively with ERα, but inversely with ErbB3 in clinical breast cancer data sets. LRIG1, an estrogen-inducible ErbB downregulator, was decreased in a panel of fulvestrant-treated luminal breast cancer cells. Ectopic LRIG1 expression from an estrogen-independent promoter uncoupled LRIG1 from estrogen regulation, thus sustaining LRIG1 and maintaining low ErbB3 levels in fulvestrant-treated cells. An LRIG1 mutant lacking the ErbB3 interaction motif was insufficient to downregulate ErbB3. Importantly, LRIG1 overexpression improved fulvestrant-mediated growth inhibition, whereas cells expressing the LRIG1 mutant were poorly sensitive to fulvestrant, despite effective ERα downregulation. Consistent with these results, LRIG1 expression correlated positively with increased disease-free survival in antiestrogen-treated breast cancer patients. These data suggest that ERα-dependent expression of LRIG1 dampens ErbB3 signaling in luminal breast cancer cells, and by blocking ERα activity with fulvestrant, LRIG1 is decreased thus permitting ErbB3 accumulation, enhanced ErbB3 signaling to cell survival pathways and blunting therapeutic response to fulvestrant.
Similar content being viewed by others


Introduction
ERBB3, the gene encoding the ErbB receptor tyrosine kinase (RTK) family member ErbB3, is expressed at relatively high levels in breast cancers that are HER2-amplified,1 and at even higher levels in Luminal A breast cancers,2 which often express and are dependent on estrogen receptor (ER)-α. Nearly 20% of all Luminal breast tumors exhibit ERBB3 gene copy number gains, causing increased ERBB3 expression, and correlating with decreased disease-free survival.2 ErbB3 expression is increased further in luminal breast cancers in response to treatment with the selective ERα modulator tamoxifen or the selective ERα downregulator fulvestrant,3, 4, 5 two drugs clinically prescribed to patients with luminal breast cancer. Little is known regarding the role of ErbB3 in luminal breast tumors, largely because of a historical description of ErbB3 as a kinase-impaired heterodimeric partner of epidermal growth factor receptor (EGFR) and ErbB2.1, 6, 7 However, increasing evidence suggests that ErbB3 is a key therapeutic target in luminal breast cancers.
Because ErbB3 harbors six phospho-tyrosine motifs that bind to the p85 regulatory subunit of phosphatidyl inositol-3-kinase (PI3K),1, 8, 9, 10, 11 ErbB3 is a potent activator of PI3K signaling.12 Once active, PI3K generates PIP3, a potent second messenger that recruits PI3K pathway effectors to the plasma membrane (for example, serine-threonine kinases PDK-1 and Akt), resulting in their activation. As Akt sits atop a signaling cascade that enhances cell growth and survival,13 ErbB3 signaling promotes survival of luminal breast cancer cells. Thus, ErbB3 upregulation in response to tamoxifen or fulvestrant increases PI3K-Akt signaling within 24 h of treatment, increasing tumor cell survival and blunting the therapeutic benefit of these endocrine inhibitors.2, 3, 5, 9 However, combination of endocrine inhibitors with strategies designed to target ErbB3 may be used to circumvent the compensatory upregulation of ErbB3-PI3K-Akt signaling, and improve luminal breast cancer cell killing. The therapeutic ErbB3 antibody U3-1287, for example, significantly impaired PI3K signaling and survival in fulvestrant-treated luminal breast tumor cells.2
The mechanism(s) contributing to ErbB3 upregulation in response to fulvestrant are poorly understood. However, recent findings demonstrated a twofold upregulation of ERBB3 transcripts in a panel of luminal breast cancer cells 24 h after fulvestrant treatment.2 Thus, it is likely that additional mechanisms of ErbB3 regulation contribute to enhanced ErbB3 protein expression in fulvestrant-treated luminal breast cancer cells. We found that estrogen-dependent expression of the ErbB family regulatory factor LRIG1 maintains ErbB3 expression at low levels under basal conditions, and that fulvestrant disrupts ERα-dependent LRIG1 expression, permitting ErbB3 accumulation and survival signaling in fulvestrant-treated cells.
Results
The ErbB3 antibody A4 neutralizes fulvestrant-mediated ErbB3 upregulation
Previous studies demonstrate that humanized anti-ErbB3 monoclonal antibody A4 binds the ErbB3 ectodomain and effectively downregulates ErbB3 in HER2-amplified breast cancer cells through receptor internalization.14, 15 However, the utility of A4 in luminal breast cancers has not yet been described. We tested the impact of A4 on ErbB3 protein levels in two luminal breast cancer cell lines, MCF7 and T47D cells. Western blot analysis confirmed ErbB3 expression in serum-starved MCF7 and T47D cells (Figure 1a), but showed that ErbB3 tyrosine phosphorylation at Y1289 was not detected under these conditions. Recombinant heregulin α1 (HRG, 2 ng/ml) induced ErbB3 Y1289 phosphorylation in control cells treated with nonspecific human IgG. In contrast, pre-treatment of cells with A4 for 24 h before addition of HRG diminished HRG-induced ErbB3 phosphorylation at Y1289 in MCF7 and T47D, and decreased total ErbB3 levels.

Fulvestrant-mediated ErbB3 upregulation is neutralized by the ErbB3 antibody A4. (a) Western blot analysis of whole-cell lysates collected from serum-starved MCF7 and T47D cells treated 24 h with antibody A4 (2–5 μg/ml) or nonspecific human IgG, and for the final 10 min of culture with recombinant HRGβ1 (2 pg/ml). (b) Western blot analysis of whole-cell lysates cultured in 10% serum and treated 24 h with fulvestrant (1 μM) or antibody A4 (2 μg/ml). (c) Cells cultured for 3 days in 10% serum in the presence or absence of fulvestrant (1 μM) or antibody A4 (2 μg/ml) were collected by trypsinization and counted. Average cell number (±s.d.) is shown. N=5, each counted in duplicate. Student’s t-test. (d) Left panel: representative images of crystal violet-stained cells after 14-day culture in 10% serum in the presence or absence of fulvestrant (1 μM) and antibody A4 (2 μg/ml). Right panel: crystal violet fluorescence was measured on the Odyssey scanner and used as a relative measure of total cell number, setting the average fluorescence value for cells cultured in IgG/DMSO equal to 1. Average±s.d. is shown. N=3, each assessed in triplicate. Student’s t-test.
Luminal breast cancer cells use ErbB3 upregulation in response to ERα targeting to increase tumor cell growth and survival.2, 8, 9 Therefore, we tested the ability of antibody A4 to neutralize ErbB3 upregulation in luminal breast cancer cells treated with the selective ERα downregulator fulvestrant. MCF7 and T47D cells were treated for 24 h with fulvestrant in the presence or absence of A4. As expected, fulvestrant reduced expression of ERα while upregulating expression of ErbB3 in control IgG-treated cultures (Figure 1b), consistent with previous findings.2, 3 However, treatment of luminal breast cancer cells with antibody A4 in combination with fulvestrant-neutralized fulvestrant-mediated ErbB3 upregulation. In MCF7 and T47D cells treated with the combination of antibody A4 and fulvestrant, phosphorylation of Akt and ribosomal protein S6 was downregulated to a greater extent than what was seen in cells treated with either agent alone.
To determine the impact of A4 on growth of luminal breast cancer cells, we treated 1 × 106 MCF7 and T47D cells with A4 for 3 days in media containing 10% serum. Treatment of cells with A4 resulted in a modest but significant decrease in cell number after 3 days of treatment (Figure 1c), consistent with previous reports using another ErbB3-specific antibody U3-1287.2 Treatment of cells with fulvestrant for 3 days decreased cell number. However, the combination of A4 with fulvestrant reduced cell number to a greater extent than either fulvestrant or A4 alone, confirming that ErbB3 upregulation in fulvestrant-treated cells limits fulvestrant-mediated growth inhibition, and that targeting ErbB3 using the ErbB3 antibody A4 works in combination with fulvestrant to inhibit tumor cell growth through inhibition of ErbB3 expression. We investigated how ErbB3 targeting with antibody A4 affects the formation of fulvestrant-resistant colonies by seeding MCF7 and T47D cells at low density. Cells were cultured for 14 days in the presence of fulvestrant (1 μM) or dimethylsulfoxide (DMSO). Fulvestrant profoundly reduced colony formation in MCF7 and T47D cells (Figure 1d). Antibody A4 also decreased colony formation in MCF7 and T47D cells as a single agent. However, the combination of fulvestrant+A4 caused superior growth inhibition as compared with fulvestrant alone or A4 alone.
Increased ErbB3 protein correlates with decreased ERα in luminal breast cancer cells
Because fulvestrant downregulates ERα while upregulating ErbB3 protein levels in cultured luminal breast cancer cell lines, we investigated the inverse relationship between ErbB3 and ERα in clinical luminal breast cancer specimens curated and analyzed by the Cancer Genome Atlas (N=321). We found high ErbB3 protein levels in 13.1% (42 of 321) of tumors from the combined Luminal A and Luminal B subgroups as measured by reverse phase protein array (RPPA, Supplementary Figure S1A). Interestingly, the RPPA score for total ERα (ESR1) protein was significantly downregulated in tumors with high ErbB3 protein levels (Figure 2a), consistent with the inverse relationship between ERα and ErbB3 protein levels seen in cultured luminal breast cancer cells. Furthermore, phosphorylation of ERα at S118 (which marks ERα transcriptional activity16) was substantially lower in luminal tumors with high ErbB3 protein levels (Supplementary Figure S1B). In contrast, ERBB3 transcripts were upregulated in 12% (37/324) luminal breast cancer specimens, but primarily in tumors distinct from those with ErbB3 protein upregulation (Figure 2b), demonstrating that ErbB3 mRNA may not predict ErbB3 protein levels in tumors. In contrast to the inverse relation observed between the RPPA scored for ErbB3 and ERα, no relationship was observed between ERα-Ser118 and ERBB3 mRNA levels (Supplementary Figure S1C), whereas a positive relationship was seen between ERBB3 mRNA and total ERα protein (Figure 2a). These data suggest that ErbB3 protein and ERRB3 mRNA correlate with ERα in distinct ways. This observation is consistent with the idea that ErbB3 protein expression, but not mRNA expression, is highest when ERα transcriptional activity is low.

ErbB3 protein levels correlate inversely with ERα, but ERBB3 mRNA levels do not. (a) Using the Cancer Genome Atlas (TCGA)-curated luminal breast cancer data sets, the RPPA protein score for ESR1 (ERα) was plotted for those tumors with high ErbB3 protein expression or high ERBB3 mRNA expression as compared with the remaining luminal tumors. Midlines represent average, whiskers indicate s.d. Analysis was performed on cBioPortal.org. (b) Oncoprint generated using the cBioportal software (www.cbioportal.org) to analyze TCGA-curated Luminal A/B tumors for tumors with high ErbB3 RPPA scores or high ERBB3 mRNA expression. Unaltered cases were removed to demonstrate the distinct compartments where ErbB3 mRNA and protein upregulation fail to overlap. (c) Western blot analysis of whole-cell lysates or avidin–biotin pull-downs from biotin-labeled MCF7 cells (lanes 1–2 and 5–6). Cells without biotin labeling were used as negative controls for avidin–biotin pull downs. (d) Western blot analysis of whole-cell lysates harvested from cells cultured in 10% serum for 24 h with DMSO or fulvestrant (1 μM), and for the final 0–120 min of culture with CHX.
We used avidin-mediated pull-down of biotinylated cell surface proteins followed by western blot analysis to determine whether ErbB3 levels were upregulated at the cell surface of fulvestrant-treated MCF7 breast cancer cells. After treatment for 24 h, ErbB3 pull-down increased more than threefold in fulvestrant-treated cells as compared with cells treated with DMSO (Figure 2c). ErbB3 was not pulled down by avidin beads in non-biotinylated cells, demonstrating the specificity of the assay. Thus ErbB3 protein presentation at the cell surface is upregulated in response to fulvestrant.
We further investigated fulvestrant-mediated ErbB3 upregulation in four fulvestrant-treated luminal breast cancer cell lines, including lines with PIK3CA-H1047R-activating mutations (T47D), PIK3CA-E545K-activating mutations (MCF7 and MDA-MB-361), HER2 overexpression (MDA-MB-361) or with wild-type (WT) PIK3CA and HER2 (HCC1428). Fulvestrant caused ErbB3 upregulation in each of these cell lines within 24 h (Figure 2d). Using cycloheximide (CHX) to block new protein synthesis, we found that ErbB3 protein levels decreased at time points following the addition of CHX to DMSO-treated cells and were not detected after 120 min of CHX (Figure 2d). In contrast, fulvestrant decreased ERα expression but increased ErbB3 expression over what was seen in DMSO-treated cells. In fulvestrant-treated HCC1428, MDA-MB-361 and T47D cells, ErbB3 remained abundant through 120 min after CHX treatment. In fulvestrant-treated MCF7 cells, however, ErbB3 diminished at a rate similar to what was seen in DMSO-treated cells. However, because baseline expression of ErbB3 was higher in fulvestrant-treated cells before adding CHX, ErbB3 levels were still evident 120 min after CHX treatment. Thus, protein stabilization of ErbB3 may occur in response to fulvestrant in many luminal breast cancer cells, independently of PIK3CA mutation or HER2 amplification; however, other mechanisms may also lead to ErbB3 protein upregulation in response to fulvestrant.
LRIG1 decreases expression of ErbB3
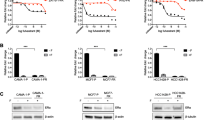
To understand the molecular events that increase cell surface ErbB3 protein levels in fulvestrant-treated luminal breast cancer cells, we turned our attention to proteins involved in ErbB3 trafficking and degradation, including LRIG1,17 Nrdp118, 19 and NEDD4,20 which were overexpressed in 14% (33/235), 9% (22/235) and 8% (19/235) of Luminal A breast cancers, respectively. RPPA data from the Cancer Genome Atlas-curated luminal breast cancers did not reveal a report for total ErbB3 levels in tumors queried for LRIG1 or NRDP1 mRNA upregulation, although total ErbB3 RPPA sores were reported on query for NEDD4 upregulation, revealing no association between ErbB3 protein levels and NEDD4 transcript expression (Supplementary Figure S2). However, the RPPA score for phospho-ErbB3 Y1289, a phospho-tyrosine induced by treatment of breast cancer cells with HRG (Figure 1a) or fulvestrant (Figure 1b), was reported in queries for LRIG1, Nrdp1 and NEDD4. Although no association was observed between P-ErbB3 Y1289 RPPA scores and either Nrdp1 or NEDD4, we found a statistically significant inverse relationship between LRIG1 and P-ErbB3 Y1289 (Figure 3a). Therefore, we focused further attention on the relationship between LRIG1 and ErbB3 in luminal breast cancer cells treated with and without fulvestrant.

LRIG1 decreases expression of ErbB3. (a) Using TCGA-curated luminal breast cancer data sets, the RPPA protein score for ErbB3 phospho-Tyr1289 was plotted for those tumors with high LRIG1, NRDP1 and NEDD4 mRNA expression as compared with the remaining luminal tumors. Midlines represent average, whiskers indicate s.d. Analysis was performed on cBioPortal.org. (b) Western blot analysis of whole-cell lysates harvested from cells stably expressing the empty pcDNA3.1 vector, LRIG1ΔEcto (a dominant-negative LRIG1 mutant), LRIG1-WT and LRIG1ΔCyto (an active LRIG1 mutant).
We stably expressed a myc-epitope-tagged LRIG1 construct17 in MCF7, HCC1428 and T47D cells (Figure 3b). In addition to WT LRIG1, we also expressed a myc-tagged LRIG1 deletion mutant lacking the large ErbB3 interaction motif in the LRIG1 ectodomain (LRIG1ΔEcto),21 and an LRIG1 mutant lacking the short regulatory/inhibitory cytoplasmic domain (LRIG1ΔCyto).21 Cells were grown in 10% serum, and we assessed cell lysates using western blot analysis for ErbB3. We found that LRIG1WT overexpression modestly decreased ErbB3 protein levels in each cell line as compared with cells expressing the empty vector pcDNA3, whereas expression of LRIG1ΔEcto, which lacks the ErbB3 interaction motif, increased ErbB3 protein levels in each cell line tested. In contrast, ErbB3 was profoundly downregulated in cells expressing LRIG1ΔCyto, consistent with the reported increased ErbB-targeting activity of LRIG1ΔCyto.
LRIG1 inhibits growth of luminal breast cancer cells through ErbB3 downregulation
Previous studies demonstrate that LRIG1 suppresses growth of cancer cells.22, 23, 24, 25 We used Kaplan–Meier Plot (kmplot.com) to examine patient survival in data sets of tamoxifen-treated ER+ breast cancers (N=665), bifurcating these tumors into those expressing high or low levels of LRIG1. The patients in this combined data set were treated with tamoxifen. Similar to fulvestrant, tamoxifen blocks the action of ERα, albeit through a distinct mechanism that warrants careful consideration in the context of the presented data. However, we used this publically available meta-analysis tool to identify potentially important information regarding an association between LRIG1 and response to endocrine-treated breast cancers. This approach identified a statistically significant decrease in disease-free survival for patients with tumors expressing lower LRIG1 levels (Figure 4a). In contrast, expression of NRDP1 or NEDD4 did not correlate with disease-free survival of tamoxifen-treated ER+ breast cancer patients (Supplementary Figure S4). These data suggest that LRIG1 expression and activity promotes a favorable outcome following tamoxifen treatment, and conversely that lower LRIG1 levels may predict a worse outcome for tamoxifen-treated patients.

ErbB3 upregulation drives increased growth of cells lacking LRIG1 activity. (a) Transcript levels of LRIG1 were assessed in tamoxifen-treated breast cancer expression microarray data sets and used to generate a Kaplan–Meier survival curve using KmPlot.com. Analysis included all tumors treated with or without chemotherapy, and excluded all samples not treated with tamoxifen. The probes used for analysis are indicated at the top of panel. (b) Western blot analysis of whole-cell lysates harvested from cells stably expressing pcDNA3.1, LRIG-WT or LRIG1-ΔEcto, and cultured for 24 h in 10% serum with antibody A4 (2 μg/ml) or IgG. (c) Cells cultured for 3 days in 10% serum in the presence or absence of antibody A4 (2 μg/ml) were collected by trypsinization and counted. Average cell number (±s.d.) is shown. N=3, each counted in duplicate. Student’s t-test. (d) Western blot analysis of whole-cell lysates harvested form cells transiently transfected with ErbB3 siRNA sequences or with non-targeting siRNA sequences. Twenty-four hours after transfection cells were seeded and cultured for 72 h in 10% serum before collection for western blot analysis. (e) Twenty-four hours after transient transfection with ErbB3 siRNA sequences or with non-targeting siRNA sequences, equal number of cells were seeded and cultured in 10% serum for 3 days. Cells were collected by trypsinization and counted. Average cell number (±s.d.) is shown. N=5, each counted in duplicate. Student’s t-test. (f) Western blot analysis of whole-cell lysates harvested from cells transiently transfected with LRIG1 siRNA sequences or non-targeting siRNA sequences. Seventy-two hours after transfection, cells were cultured in 10% serum with antibody A4 (2 μg/ml) or IgG for 24 h. (g) Twenty-four hours after transient transfection with LRIG1 siRNA sequences or with non-targeting siRNA sequences, cells were cultured in 10% serum with antibody A4 (2 μg/ml) or IgG for 3 days. Cells were collected by trypsinization and counted. Average cell number (±s.d.) is shown. N=5, each counted in duplicate. Student’s t-test.
To study this more directly, and to determine the role of ErbB3 in LRIG1-mediated growth inhibition, we measured the impact of exogenous LRIG1 expression in luminal breast cancer cells. Overall, 2.0 × 106 MCF7 and HCC1428 cells stably expressing LRIG1WT, LRIG1ΔEcto or the empty expression vector pcDNA3.1 were seeded and cultured with and without the ErbB3 antibody A4 for 3 days. Western blot analysis confirmed that ErbB3 levels were reduced on A4 treatment in cells expressing the empty vector or expressing LRIG1ΔEcto as compared with these same cells treated with a control IgG (Figure 4b). Although ErbB3 levels were already substantially reduced in cells expressing LRIG1WT, ErbB3 protein levels were further reduced by A4. Cell counting revealed fewer cells in IgG-treated samples expressing LRIG1WT as compared with cells expressing the empty pcDNA3.1 vector, but increased cells in cultures expressing LRIG1ΔEcto (Figure 4c). Cell growth was substantially reduced in LRIG1ΔEcto cultures treated with antibody A4, suggesting that cells expressing LRIG1ΔEcto have a growth advantage over cells expressing the empty vector due in large part to expression of ErbB3. A4 did not further decrease the growth of cells expressing LRIG1WT. These results were confirmed using short interfering RNA (siRNA)-mediated ErbB3 downregulation. MCF7 cells expressing LRIG1WT, LRIG1ΔEcto or the empty vector were transiently transfected with siRNA against ErbB3. At 24 h after transfection, 1.0 × 105 cells were seeded in six-well dishes, and cultured for 3 days in 10% serum, at which time parallel wells were assessed using western blot analysis or by cell counting. Western blot analysis of whole-cell lysates confirmed decreased expression of ErbB3 in cells expressing LRIG1WT, and increased ErbB3 levels in cells expressing LRIG1ΔEcto as compared with cells harboring the empty vector. Cells transfected with siRNA sequences directed against ErbB3 demonstrated a reduction in ErbB3 levels (Figure 4d). Cell counting revealed that knockdown of Erbb3 reduced the growth of MCF7 cells expressing LRIG1ΔEcto and MCF7 cells expressing the empty vector (Figure 4e). Although cells expressing LRIG1WT displayed reduced growth as compared with cells expressing empty pcDNA3.1, knockdown of ErbB3 did not decrease growth of these cells further. These observations are consistent with the idea that the tumor suppressor LRIG1 decreases growth of luminal breast cancer cells through downregulation of ErbB3.
To understand the consequences of LRIG1 depletion, we used two distinct siRNA sequences directed against human LRIG1 to knockdown LRIG1 expression. Western blot analysis confirmed knockdown of LRIG1 following transient transfection of sequences targeting LRIG1, but not control siRNA sequences (Figure 4f). ErbB3 levels were increased in MCF7 and HCC1428 cells on LRIG1 knockdown but were reduced in the presence of antibody A4. To assess the impact of LRIG1 knockdown on growth of luminal breast cancer cells, we seeded 2.0 × 106 MCF7 and HCC1428 cells 24 h after siRNA transfection, and then cultured cells for an additional 3 days in 10% serum, with and without antibody A4. Cell counting after 3 days revealed increased growth of cells transfected with LRIG1 siRNA as compared with cells transfected with control siRNA sequences (Figure 4g). Antibody A4 blocked the growth advantage conferred to MCF7 and HCC1428 cells by LRIG1 knockdown. Together, these data suggest that LRIG1 is a key mechanism restraining ErbB3-mediated growth in luminal breast cells.
Decreased LRIG1 levels in fulvestrant-treated luminal breast cancer cells
Previous reports demonstrate that estrogen-activated ERα transactivates the LRIG1 gene.22 Consistent with these findings, luminal breast cancers with high ERα RPPA scores displayed significantly higher expression of LRIG1 mRNA (Figure 5a; N=235, P=0.003). In contrast, ERα RPPA scores did not correlate with Nrdp1 or NEDD4 expression in luminal breast cancers. Similarly, the RPPA score for P-ERα s-118 correlated positively with LRIG1 mRNA levels but showed no correlation with NEDD4 or NRDP1 mRNA levels (Supplementary Figure S4). These data are consistent with the idea that LRIG1 is an ERα-activated factor in luminal breast cancers.22 Therefore, we tested the hypothesis that ERα downregulation in response to fulvestrant causes decreased expression of LRIG1. LRIG1 downregulation was seen in MCF7, MDA-MB-361, HCC1428 and T47D cells cultured for 24 h with fulvestrant as compared with LRIG1 levels seen in DMSO-treated cells (Figure 5b). Although downregulation of ERα was nearly complete, LRIG1 downregulation, as measured using western blot analysis, was partial in MCF7 and MDA-MB-361 cells, although LRIG1 was more substantially downregulated in T47D and HCC1428 cells. We measured LRIG1 mRNA levels in MCF7, T47D, HCC1428 and MDA-MB-361 cells cultured for 24 h in the presence or absence of fulvestrant, revealing a substantial reduction in LRIG1 mRNA levels in fulvestrant-treated cells (Figure 5c). Further, MCF7 and MDA-MB-361 xenografts from mice treated with fulvestrant for 7 days (N=7) expressed a reduced level of LRIG1 transcripts as compared with control-treated tumors (Figure 5d). Conversely, MCF7 and T47D cells cultured in 10% charcoal-stripped serum and treated with 20 pM 17β-estradiol for 24 h upregulated LRIG1 protein levels while concurrently downregulating ErbB3 protein levels (Figure 5e). This suggests that ERα positively regulates LRIG1 transcript levels in luminal breast cancer cells, although additional mechanisms may contribute to total LRIG1 protein levels.

LRIG1 decreases growth of luminal breast cancer cells. (a) Using TCGA-curated luminal breast cancer data sets, the RPPA protein score for ESR1 (ERα) was plotted for those tumors with high LRIG1, NRDP1 and NEDD4 mRNA expression as compared with the remaining luminal tumors. Midlines represent average, whiskers indicate s.d. Analysis was performed on cBioPortal.org. (b) Western blot analysis of whole-cell lysates harvested from cells cultured for 24 h in 10% serum with fulvestrant (1 μM) or equal volume of DMSO. (c) LRIG1 transcripts were measured in whole-cell RNA harvested from cells cultured for 24 h in 10% serum with fulvestrant (1 μM) or DMSO. Values represent average relative mRNA levels (±s.d.), N=3, each analyzed in triplicate. Student’s t-test. (d) LRIG1 transcripts were measured in whole-tumor RNA harvested from MCF7 and MDA-MB-361 xenografts treated for 7 days with fulvestrant or vehicle. Values represent average relative mRNA levels (±s.d.), N=4, each analyzed in triplicate. Student’s t-test. (e) Western blot analysis of whole-cell lysates harvested from cells cultured for 24 h in 10% charcoal-stripped serum with estrogen (20 pM) or equal volume of ethanol. Values represent average relative mRNA levels (±s.d.), N=3, each analyzed in triplicate. Student’s t-test.
Uncoupling LRIG1 from ERα sustains ErbB3 downregulation in fulvestrant-treated breast cancer cells
To determine whether exogenous expression of LRIG1WT could maintain ErbB3 protein downregulation in fulvestrant-treated cells, we treated MCF7 cells expressing LRIG1WT with fulvestrant for 24 h. Although LRIG1 levels were decreased in vector-control cells treated with fulvestrant, MCF7-LRIG1WT cells maintained high LRIG1 expression levels following fulvestrant treatment (Figure 6a). Importantly, ErbB3 levels, which were elevated in fulvestrant-treated MCF7-pcDNA3 cells, remained low in fulvestrant-treated MCF7-LRIG1WT cells. We also used biotinylation of cell surface proteins to demonstrate that fulvestrant-mediated upregulation of cell surface ErbB3 is blocked in cells expressing LRIG1WT as compared with MCF7-pcDNA3 cells (Figure 6b). Because LRIG1 inhibits growth of luminal breast cancer cells, and fulvestrant decreases expression of LRIG1, we wanted to test the hypothesis that LRIG1 downregulation in response to fulvestrant results in a growth advantage that partially masks the tumor cell growth inhibition that could be achieved by fulvestrant. We tested this hypothesis in fulvestrant-treated MCF7 cells. Fulvestrant decreased the growth of MCF7-pcDNA3 cells over 7 days (Figure 6c). However, fulvestrant-mediated growth inhibition was significantly greater in MCF7-LRIGWT and MCF7-LRIGΔCyto cells as compared with MCF7-pcDNA3.1 cells. In contrast, MCF7-LRIG1ΔEcto cells were relatively resistant to the growth inhibitory effects of fulvestrant, indicating that the ectodomain of LRIG1 is necessary for LRIG1-mediated growth inhibition. These data suggest that sustained expression of LRIG1 improves the response of luminal breast cancer cells to fulvestrant, and conversely that loss of ERα-dependent LRIG1 expression weakens tumor growth inhibition in response to fulvestrant. Western blot analysis confirmed continued upregulation of ErbB3 at 7 days of fulvestrant treatment in MCF7 cells expressing pcDNA3 (Figure 6d). Taken together, these data suggest that ERα drives expression of the growth inhibitory protein LRIG1, a protein necessary for ErbB3 downregulation (Figure 6c). Upon loss of ERα and hence loss of LRIG1, ErbB3 levels accumulate at the cell surface, contributing to growth in fulvestrant-treated cells.

LRIG1 downregulation drives fulvestrant-mediated ErbB3 upregulation. (a) Western blot analysis of whole-cell lysates harvested from cells stably expressing pcDNA3.1 or LRIG1-WT. Cells cultured in 10% serum and with fulvestrant (1 μM) or DMSO for 24 h. (b) Western blot analysis of whole-cell lysates or avidin–biotin pull-downs from cells labeled with or without biotin for 30 min following 24-h treatment with fulvestrant or DMSO. (c) Cells expressing pcDNA3, LRIG1-WT, LRIG1-ΔEcto or LRIG1-ΔCyto were cultured in 10% serum with fulvestrant (1 μM) or DMSO for 7 days. Cells were collected by trypsinization and counted. Average cell number (±s.d.) is shown. N=5, each counted in duplicate. Student’s t-test. (d) Western blot analysis of whole-cell lysates harvested from cells stably expressing pcDNA3.1 or LRIG1-WT. Cells cultured in 10% serum for 7 days, and with fulvestrant (1 μM) for the final 3 days or 7 days, or cultured with DMSO for 7 days. (e) Schematic of how ERα-induced expression of LRIG1 maintains ErbB3 at low levels in luminal breast cancer cells. Endocrine inhibitors, such as fulvestrant, tamoxifen or aromatase inhibitors would cause reduced LRIG1 expression levels, thus allowing ErbB3 accumulation at the cell surface.
Discussion
We have shown previously that ErbB3 drives growth and survival of the normal luminal mammary epithelium through MAPK and PI3K pathways, respectively.26 Similarly, luminal breast cancers also use ErbB3 to increase cell survival in a PI3K-dependent manner,27 particularly in fulvestrant-treated cells,2, 3 making the combination of therapeutic ErbB3 antibodies with fulvestrant an attractive approach to treat luminal breast cancers.
Several reports suggest that genetically enforced ErbB3 overexpression enhances endocrine resistance in luminal breast cancer cells.2, 3 More recently, it was shown that ErbB3 upregulation in response to the loss of ER activity is not a genetic event acquired over time, but instead is a mechanism inherent to many luminal breast cancer cells.2 We demonstrate herein that ERα-dependent expression of LRIG1,22, 28 a cell surface protein involved in ErbB family RTK trafficking and/or degradation,17, 25, 29 is downregulated in fulvestrant-treated luminal breast cancer cells, allowing ErbB3 to accumulate on the surface of breast cancer cells (Figure 2), where it sustains cell growth. These data are consistent with previous reports demonstrating that estrogen-activated ERα controls expression of LRIG1, such that estrogen deprivation or ERα targeting reduce expression of LRIG1 in luminal breast cancer cells.22 These findings explain the relationships shown here between ErbB3 protein (but not mRNA), ERα protein and LRIG1 mRNA in luminal breast cancers.
Importantly, decreased LRIG1 transcript levels correlated with decreased disease-free survival in tamoxifen-treated luminal breast cancers (Figure 5). Because LRIG1 negatively regulates cell surface levels of ErbB family RTKs, it is possible that increased signaling through ErbB family RTKs contributes to the worse outcome for those luminal breast tumors with low LRIG1. The ErbB RTK family comprises four members, including EGFR, ErbB2, ErbB3 and ErbB4.1, 30 Although our studies focus on ErbB3, previous studies demonstrate that enforced overexpression of EGFR, ErbB2 or certain ErbB4 splice variants increases cell growth and survival in tamoxifen-treated luminal breast cancer cells.3, 4, 5, 8, 31 While we have not ruled out the potential roles of EGFR, ErbB2, and ErbB4 in regulating the response of luminal breast tumors to fulvestrant, we found that antibody-based ErbB3 targeting neutralized ErbB3 upregulation in fulvestrant-treated cells, resulting in profound growth inhibition. Blockade of post-therapeutic ErbB3 upregulation by antibody A4 has been reported in Raf/Mek inhibitor-treated melanomas15, 32 and EGFR inhibitor-treated non-small cell lung cancers.33 Moreover, we found that the ErbB3 antibody blocked cell growth caused by LRIG1 knockdown or dominant-negative LRIG1ΔEcto expression.
In summary, we have shown herein that the ERα-regulated gene LRIG1 suppresses cell surface ErbB3 expression and signaling in luminal breast cancers, such that ErbB3 rapidly accumulates on the cell membrane following treatment with fulvestrant. Previous studies demonstrate the profound impact of ErbB3 upregulation on PI3K-Akt-mTOR signaling in fulvestrant-treated breast cancers.2, 3 Although mammalian target of rapamycin (mTOR) inhibitors are approved in combination with endocrine therapy to prevent the consequences of this feedback inhibition,34 mTOR inhibitors are often poorly tolerated. Because ErbB3-targeting antibodies block resurgent signaling through the PI3K-Akt pathway in fulvestrant-treated cells,2 these data suggest that ErbB3 antibodies might be a therapeutic alternative for use in combination with endocrine inhibitors.
Materials and methods
Cell culture
All human breast cancer cell lines (MCF7, T47D, HCC1428 and MDA-MB-361) were obtained from American Tissue Type Collection (Rockville, MD, USA) and were cultured in Dulbecco’s Modified Eagle’s Medium supplemented with 10% fetal bovine serum (Atlanta Biologics, Norcross, GA, USA) except where indicated as serum-free. Fulvestrant (1 μM; Sigma-Aldrich, St Louis, MO, USA) and Heregulin-1β epidermal growth factor-like domain (2 ng/ml, R&D Systems, Minneapolis, MN, USA), and the humanized ErbB3 monoclonal antibody A4,14, 15 were used where indicated. Colony assays were seeded in six-well dishes (105 cells/dish) and cultured for 14 days in Dulbecco’s Modified Eagle’s Medium/10% fetal bovine serum. Cells were fixed in formalin, stained with crystal violet, imaged on a flatbed scanner, converted to binary images and quantified using the Image J software (US National Institutes of Helath, Bethesda, MD, USA). Cells (1 × 105) were transfected using Xfect Reagent (Clontech, Mountain View, CA, USA) according to the manufacturer’s directions with 2 μg plasmid DNA: pcDNA3.1 (Life Technologies/InVitrogen, Carlsbad, CA, USA), pcDNA3.1-LRIG1, pcDNA3.1-LRIG1-ΔEcto and pcDNA3.1-LRIG1-ΔCyto.21, 22 LRIG1 siRNA and nonspecific siRNA sequences were purchased (Dharmacon, Lafayette, CO, USA) and used for transfection of cells with Dharmafect (Dharmacon) according to the manufacturer’s directions.
Western blot analysis and immunoprecipitation
Cells were homogenized in ice-cold lysis buffer (50 mM Tris pH 7.4, 100 mM NaF, 120 mM NaCl, 0.5% NP-40, 100 μM Na3VO4 and 1 × protease inhibitor cocktail (Roche Life Science, Basel, Switzerland)). Lysates (1 mg) were used for immunoprecipitation with ErbB3 (Santa Cruz Biotechnologies, C-17, Santa Cruz, CA, USA) or 9E10 (Sigma-Aldrich) at 1:200 (v/v) for 2 h, and then pulled down with Protein A/G+ agarose (Santa Cruz Biotechnologies) at 1:20. immunoprecipitations or cell lyates (20 μg) were denatured in reducing sample buffer, resolved by 4–12% SDS–PAGE (Novex, Life Technologies, Carlsbad, CA, USA) and transferred to nitrocellulose membranes (iBlot, Life Technologies, Carlsbad, CA, USA). Membranes were blocked and probed with antibody as previously described26 using the following antibodies: ErbB3 (C-17, Santa Cruz Biotechnology; 1:1000); Y1289 P-ErbB3 (Cell Signaling; 1:1000); 9E10 (Sigma-Aldrich; 1:2000); LRIG1 (Sigma-Aldrich; 1:500) and α-actin (Sigma-Aldrich; 1:20 000).
Biotinylation of cell surface proteins
Cells were cultured to 80–90% confluence, and then incubated for 30 min at 4 °C with EZ-Link Sulfo-NHS-SS-Biotin in the Pierce Cell Surface Protein Isolation Kit (Thermo Scientific and Life Technologies, Carlsbad, CA, USA) according to the manufacturer’s specifications. Cells were lysed in Pierce Lysis Buffer (Thermo Scientific) supplemented with protease inhibitor cocktail (Roche Life Science), sonicated five times at low power on ice for 1 s each and clarified by centrifugation. Cells were collected, washed and eluted with dithiothreitol using NeutrAvidin Agarose beads provided with the Pierce Cell Surface Protein Isolation Kit (Thermo Scientific) according to the manufacturer’s directions. Eluates were analyzed using western blot analysis.
Human breast tumor gene expression analysis
The Cancer Genome Atlas35 Invasive Breast Cancer data set was clustered according to PAM50 subtypes and corresponding data on ERBB3, LRIG1, NEDD4 and NRDP1 mRNA, patient survival and ErbB3, ErbB3 PY1289, ESR1 and ESR1 P-S118 were analyzed using publically available software on the cBio Portal website.36 Meta-analysis of expression array data sets of tamoxifen-treated Luminal A breast cancers were assessed for correlations between disease-free survival and expression of LRIG1 (211596_s_at), NRDP1 (201961_s_at) and NEDD4 (213012_at) using KmPlotter (KmPlot.com).
RNA extraction and quantitative reverse transcription–polymerase chain reaction (RT–PCR)
Using RNeasy (Qiagen, Limburg, Germany), total RNA was harvested from cell lysates passed through a QiaShredder column (Qiagen) or from flash-frozen tumors homogenized using a tissue lyser. Total RNA was quantified and 2 μg RNA was used to synthesize cDNA (High Capacity cDNA Reverse Transcription kit, Applied Biosystems and Life Technologies, Carlsbad, CA, USA). Quantitative RT–PCR was performed on a Bio-Rad iCycler iQ5 machine (BioRad and Life Technologies, Carlsbad, CA, USA) using the following primer sequences:
Human LRIG1 F: 5′-TAACTCGAGAACCCACTGCTTACT-3′
Human LRIG1 R: 5′-CTAGAAGGCACACTCGAGGCTGAT-3′.
Xenograft experiments
Estrogen pellets (14-day extended release, 0.17 mg β estradiol, Innovative Research of America) were implanted subcutaneouly into 3- to 4-week-old female Balb/C athymic nude mice (The Jackson Laboratories, Bar Harbor, ME, USA). Right and left inguinal mammary fatpads were injected with 106 MCF7 or MDA-MB-361 in 100 μl growth factor-reduced Matrigel. Tumors were measured with calipers twice weekly. Tumor-bearing mice were randomized into treatment arms to receive fulvestrant (Novartis) delivered once weekly by intramuscular injection. All animal experimentations were performed in AAALAC-approved facilities at the Vanderbilt University Medical Center. All animal use protocols were reviewed and approved before experimentation by the Institutional Animal Care and Use Committee at the Vanderbilt University, Nashville, TN, USA.
Statistical overview
To assess statistical significance in experiments directly comparing an experimental group to a control group, Student’s unpaired, two-tail t-test was used. P<0.05 was considered significantly different from the null hypothesis.
Change history
03 March 2016
This article has been corrected since Advance Online Publication and a corrigendum is also printed in this issue.
References
Stern DF . ERBB3/HER3 and ERBB2/HER2 duet in mammary development and breast cancer. J Mammary Gland Biol Neoplasia 2008; 13: 215–223.
Morrison MM, Hutchinson K, Williams MM, Stanford JC, Balko JM, Young C et al. ErbB3 downregulation enhances luminal breast tumor response to antiestrogens. J Clin Invest 2013; 123: 4329–4343.
Hutcheson IR, Goddard L, Barrow D, McClelland RA, Francies HE, Knowlden JM et al. Fulvestrant-induced expression of ErbB3 and ErbB4 receptors sensitizes oestrogen receptor-positive breast cancer cells to heregulin beta1. Breast Cancer Res 2011; 13: R29.
Tovey S, Dunne B, Witton CJ, Forsyth A, Cooke TG, Bartlett JM . Can molecular markers predict when to implement treatment with aromatase inhibitors in invasive breast cancer? Clin Cancer Res 2005; 11: 4835–4842.
Liu B, Ordonez-Ercan D, Fan Z, Edgerton SM, Yang X, Thor AD . Downregulation of erbB3 abrogates erbB2-mediated tamoxifen resistance in breast cancer cells. Int J Cancer 2007; 120: 1874–1882.
Kraus MH, Issing W, Miki T, Popescu NC, Aaronson SA . Isolation and characterization of ERBB3, a third member of the ERBB/epidermal growth factor receptor family: evidence for overexpression in a subset of human mammary tumors. Proc Natl Acad Sci USA 1989; 86: 9193–9197.
Kim HH, Vijapurkar U, Hellyer NJ, Bravo D, Koland JG . Signal transduction by epidermal growth factor and heregulin via the kinase-deficient ErbB3 protein. Biochem J 1998; 334: 189–195.
Hamburger AW . The role of ErbB3 and its binding partners in breast cancer progression and resistance to hormone and tyrosine kinase directed therapies. J Mammary Gland Biol Neoplasia 2008; 13: 225–233.
Sutherland RL . Endocrine resistance in breast cancer: new roles for ErbB3 and ErbB4. Breast Cancer Res 2011; 13: 106.
Hellyer NJ, Cheng K, Koland JG . ErbB3 (HER3) interaction with the p85 regulatory subunit of phosphoinositide 3-kinase. Biochem J 1998; 333: 757–763.
Kim HH, Sierke SL, Koland JG . Epidermal growth factor-dependent association of phosphatidylinositol 3-kinase with the erbB3 gene product. J Biol Chem 1994; 269: 24747–24755.
Klempner SJ, Myers AP, Cantley LC . What a tangled web we weave: emerging resistance mechanisms to inhibition of the phosphoinositide 3-kinase pathway. Cancer Discov 2013; 3: 1345–1354.
Wong KK, Engelman JA, Cantley LC . Targeting the PI3K signaling pathway in cancer. Curr Opin Genet Dev 2010; 20: 87–90.
Aurisicchio L, Marra E, Luberto L, Carlomosti F, De Vitis C, Noto A et al. Novel anti-ErbB3 monoclonal antibodies show therapeutic efficacy in xenografted and spontaneous mouse tumors. J Cell Physiol 2012; 227: 3381–3388.
Belleudi F, Marra E, Mazzetta F, Fattore L, Giovagnoli MR, Mancini R et al. Monoclonal antibody-induced ErbB3 receptor internalization and degradation inhibits growth and migration of human melanoma cells. Cell Cycle 2012; 11: 1455–1467.
Ali S, Metzger D, Bornert JM, Chambon P . Modulation of transcriptional activation by ligand-dependent phosphorylation of the human oestrogen receptor A/B region. EMBO J 1993; 12: 1153–1160.
Laederich MB, Funes-Duran M, Yen L, Ingalla E, Wu X, Carraway KL 3rd et al. The leucine-rich repeat protein LRIG1 is a negative regulator of ErbB family receptor tyrosine kinases. J Biol Chem 2004; 279: 47050–47056.
Diamonti AJ, Guy PM, Ivanof C, Wong K, Sweeney C, Carraway KL 3rd . An RBCC protein implicated in maintenance of steady-state neuregulin receptor levels. Proc Natl Acad Sci USA 2002; 99: 2866–2871.
Qiu XB, Goldberg AL . Nrdp1/FLRF is a ubiquitin ligase promoting ubiquitination and degradation of the epidermal growth factor receptor family member, ErbB3. Proc Natl Acad Sci USA 2002; 99: 14843–14848.
Huang Z, Choi BK, Mujoo K, Fan X, Fa M, Mukherjee S et al. The E3 ubiquitin ligase NEDD4 negatively regulates HER3/ErbB3 level and signaling. Oncogene 2014; 34: 1105–1115.
Rafidi H, Mercado F 3rd, Astudillo M, Fry WH, Saldana M, Carraway KL 3rd et al. Leucine-rich repeat and immunoglobulin domain-containing protein-1 (Lrig1) negative regulatory action toward ErbB receptor tyrosine kinases is opposed by leucine-rich repeat and immunoglobulin domain-containing protein 3 (Lrig3). J Biol Chem 2013; 288: 21593–21605.
Krig SR, Frietze S, Simion C, Miller JK, Fry WH, Rafidi H et al. Lrig1 is an estrogen-regulated growth suppressor and correlates with longer relapse-free survival in ERalpha-positive breast cancer. Mol Cancer Res 2011; 9: 1406–1417.
Miller JK, Shattuck DL, Ingalla EQ, Yen L, Borowsky AD, Young LJ et al. Suppression of the negative regulator LRIG1 contributes to ErbB2 overexpression in breast cancer. Cancer Res 2008; 68: 8286–8294.
Shattuck DL, Miller JK, Laederich M, Funes M, Petersen H, Carraway KL 3rd et al. LRIG1 is a novel negative regulator of the Met receptor and opposes Met and Her2 synergy. Mol Cell Biol 2007; 27: 1934–1946.
Stutz MA, Shattuck DL, Laederich MB, Carraway KL 3rd, Sweeney C . LRIG1 negatively regulates the oncogenic EGF receptor mutant EGFRvIII. Oncogene 2008; 27: 5741–5752.
Balko JM, Miller TW, Morrison MM, Hutchinson K, Young C, Rinehart C et al. The receptor tyrosine kinase ErbB3 maintains the balance between luminal and basal breast epithelium. Proc Natl Acad Sci USA 2012; 109: 221–226.
Cook RS, Garrett JT, Sanchez V, Stanford JC, Young C, Chakrabarty A et al. ErbB3 ablation impairs PI3K/Akt-dependent mammary tumorigenesis. Cancer Res 2011; 71: 3941–3951.
Ghasimi S, Haapasalo H, Eray M, Korhonen K, Brannstrom T, Hedman H et al. Immunohistochemical analysis of LRIG proteins in meningiomas: correlation between estrogen receptor status and LRIG expression. J Neurooncol 2012; 108: 435–441.
Wong VW, Stange DE, Page ME, Buczacki S, Wabik A, Itami S et al. Lrig1 controls intestinal stem-cell homeostasis by negative regulation of ErbB signalling. Nat Cell Biol 2012; 14: 401–408.
Wiseman SM, Makretsov N, Nielsen TO, Gilks B, Yorida E, Cheang M et al. Coexpression of the type 1 growth factor receptor family members HER-1, HER-2, and HER-3 has a synergistic negative prognostic effect on breast carcinoma survival. Cancer 2005; 103: 1770–1777.
Sundvall M, Iljin K, Kilpinen S, Sara H, Kallioniemi OP, Elenius K . Role of ErbB4 in breast cancer. J Mammary Gland Biol Neoplasia 2008; 13: 259–268.
Fattore L, Marra E, Pisanu ME, Noto A, de Vitis C, Belleudi F et al. Activation of an early feedback survival loop involving phospho-ErbB3 is a general response of melanoma cells to RAF/MEK inhibition and is abrogated by anti-ErbB3 antibodies. J Transl Med 2013; 11: 180.
Noto A, De Vitis C, Roscilli G, Fattore L, Malpicci D, Marra E et al. Combination therapy with anti-ErbB3 monoclonal antibodies and EGFR TKIs potently inhibits non-small cell lung cancer. Oncotarget 2013; 4: 1253–1265.
Baselga J, Campone M, Piccart M, Burris HA 3rd, Rugo HS, Sahmoud T et al. Everolimus in postmenopausal hormone-receptor-positive advanced breast cancer. N Engl J Med 2012; 366: 520–529.
Comprehensive molecular portraits of human breast tumours. Nature. [Research Support, N.I.H., Extramural Research Support, Non-U.S. Gov't Research Support, U.S. Gov't, Non-P.H.S.] 2012; 490: 61–70.
Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov 2012; 2: 401–404.
Acknowledgements
This work was supported by the National Institutes of Health R01CA143126 (RSC) and Susan G. Komen for the Cure grant KG100677 (RSC).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogene website
Supplementary information
Rights and permissions
About this article

Cite this article
Morrison, M., Williams, M., Vaught, D. et al. Decreased LRIG1 in fulvestrant-treated luminal breast cancer cells permits ErbB3 upregulation and increased growth. Oncogene 35, 1143–1152 (2016). https://doi.org/10.1038/onc.2015.169
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2015.169
- Springer Nature Limited
This article is cited by
-
LRIG1 is a pleiotropic androgen receptor-regulated feedback tumor suppressor in prostate cancer
Nature Communications (2019)