
Abstract
Giant squirrels are imperilled across their distributional range as a result of anthropogenic disturbances; converting tropical landscapes into mosaics of small habitat patches interspersed within an inhospitable matrix of agricultural land, plantations and human settlement. With limited understanding of Ratufa bicolor’s habitat relations in the lowland tropical forests, this study aimed to understand the correlates of habitat use from an isolated habitat patch and its adjoining matrix in the Brahmaputra valley which represents the northern distribution limit of the species. The ecological dataset comprised of grid-based spatial occurrence data summarized as average squirrel encounter rate (4 temporal replicate transect surveys in each grid), as well as a range of fine spatial scale ecological covariates (14 habitat attributes). We used generalized linear models and principal component analysis to characterize relationships between squirrel abundance and habitat variables representing different aspects of forest, i.e., structure, composition and food availability, anthropogenic disturbance and edge influence. Our study revealed giant squirrels’ selection of habitat patches to be based on concentrated areas of critical habitat attributes with strong support for the influence of canopy connectivity, canopy density, overall tree density and availability of food tree. Further, a quantitative model for habitat use was evaluated using GLM linking squirrel abundance to the predictive parameters representing different forest aspects, with the best fit model highlighting the positive interaction effects of forest structure and forest composition. The findings from the study provide insights into the necessity of maintaining associated habitat matrix to serve as complimentary space, ensuring future survival of arboreal mammals in human-modified landscapes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Indo-Malayan genus Ratufa is represented by the world’s four largest tree squirrels; with all the members having an extensive oriental distribution and each one featuring in Appendix II of CITES, being legally protected in most countries across their distributional range (Thorington et al. 2012). Among these four extant species, R. bicolor (Sparrman, 1778) is the largest of all and has an extensive mainland distribution occurring from South Asia (Northeast India, Nepal and Bangladesh), Southern China into much of Mainland and Maritime Southeast Asia (Fig. 1), inhabiting a variety of tropical and sub-tropical forests (Chatterjee et al. 2020; Duckworth and Molur 2016). Ecologically, they are restricted to the highest arboreal niche in the canopy and their life history has been linked to numerous facets of old growth and complex forests throughout their distributional range to a profound extent (Borges 2008; Johnsingh and Manjrekar 2016). Giant squirrels are often considered an integral component of animal biomass in tropical forest ecosystem (Eisenberg 1980), performing vital ecosystem services with their interactions with fruits and seeds as consumers and dispersers (Thorington and Cifelli 1990). Thus, the tropics, with their complex physiognomic structure of forests, have long served as an ideal habitat for these giant squirrels, shaping their evolution, diversification and endemism (Koprowski and Nandini 2008; Roth and Mercer 2008). However, tropical landscapes are undergoing unprecedented rates of degradation and conversion than any other biome in the world (Hansen et al. 2013; Laurance 2007, 2015; Mayaux et al. 2005).

Global distributional range of giant squirrel, Ratufa bicolor (Sparrman, 1778). Inset: Picture of an adult R. bicolor, commonly known as Malayan giant squirrel in a Dipterocarp Forest (Photo- Samrat Sengupta)
Human land use for agriculture and developmental activities has spread and largely intensified in the last few decades, permanently altering tropical landscapes (Dobrovolski et al. 2011; Gibbs et al. 2010). Presently, they exist as mosaics of isolated habitat patches of varying sizes, interspersed within an inhospitable matrix (i.e., the non-habitat surrounding habitat patches) formed of agricultural land, plantations and human settlements (Fahrig 2013). This has threatened the survival prospects of arboreal mammals that are habitat specialists; confining them to patches with reduced habitat availability and quality in human-modified landscapes (Kakati et al. 2009; Myers and Knoll 2001; Sharma et al. 2012; Taubert et al. 2018). Under these prevailing conditions, the only way forward for conservation practice should be to incorporate not only the habitat patches, but also their associated ‘matrix’ under their existing management goal in a pragmatic manner. Studies suggest a tremendous potential of surrounding matrix to form a continuum of habitat when properly managed, which can mitigate the negative effects of habitat loss and fragmentation (Franklin and Lindenmayer 2009). Also, species capable of efficient use of matrix around small habitat fragments are known to be less susceptible towards stochastic events of local extinctions (de Castro and Fernandez 2004; Laurance 1991). Therefore, understanding the life history and habitat relations of native fauna and the ecological importance of the surrounding matrix is vital for conservation planning and management practices.
However, to date, little is known about the habitat relations of R. bicolor (commonly called as Malayan giant squirrel; hereafter MGS) in the lowland tropical forests of Northeast India, which represents the northern distribution limit of the species (Choudhury 2013) and has been projected to become ecologically ‘unsuitable’, thus incurring a loss of 90% of their present distribution range in the context of changing climate (Chatterjee et al. 2020). Alarmingly, their population have also plummeted through much of its distributional range (suspected to be nearly 30% decline within the last two decades itself, Duckworth and Molur 2016), raising concerns globally. Of the many factors attributed to the cause of this decline, urbanization and agronomic pressure-induced habitat loss and fragmentation were considered particularly critical, making them imperilled across their distributional range (Duckworth and Molur 2016). With a diet primarily based on seeds and fruits, MGS rely heavily on large trees for nesting and foraging (Sengupta 2017; Shyam and Saikia 2012), making them highly sensitive to any kind of forest perturbations. With this rationale in mind, we assessed the patterns of fine-scale space use (habitat utilization) by R. bicolor in an isolated habitat patch including the surrounding matrix, and explored the potential role of habitat profile (structural and compositional attributes of the forest) in regulating their site level occupancy and abundance pattern. We are hopeful that the findings of the study will provide valuable insights into the ecological drivers of their local densities and enable us to identify the suite of habitat covariates that governs the intensity of habitat use. This will help to chalk out conservation plans for the long-term survival and persistence of many locally restricted populations of MGS across such isolated forest patches in a tropical landscape.
Materials and methods
Study area
Hollongapar Gibbon Sanctuary (hereafter HGS) is a small tropical forest patch in the Brahmaputra Valley region (Upper Assam, Northeast India, Fig. 2A, B), located on the southern bank of the river Brahmaputra. Situated between 26º 40′–26º 45′ N latitude and 94º 20′–94º 25′ E longitude (Fig. 2) with an elevation range of 100–120 m.s.l., this forest patch supports a rich assemblage of arboreal mammal community with evidences of their continued survival and persistence, prone from any local extinction (see Sharma et al. 2012, 2014).

Study area showing location of ‘Hollongapar Gibbon Sanctuary’ and its peripheral landscapes in Upper Brahmaputra Valley, Assam. A India, blue rectangle = Upper Assam, Northeast India; B Upper Brahmaputra Valley; C Digital elevation map of the study area derived from DEM (ASTER-DEM) data. D Land use and land cover map of the study area, derived from Landsat data recorded during the period of study
Named after its two flagship species, Western hoolock gibbons (Hoolock hoolock) and Hollong tree (Dipterocarpus retusus), this forest patch has a conservation history dating back to 1881 when it was declared as a “Reserved Forest” which was subsequently upgraded to “Wildlife Sanctuary” in the year 1997 (Choudhury 1999). However, unabated logging and land conversion gradually converted HGS into an “island” forest patch with complete isolation from any nearby patches as sprawling tea gardens (between 1880 and 1920), village settlements (during the 1960s) and linear infrastructures began to emerge (Hazarika and Gupta 2005). HGS has a sub-tropical climate with distinct season: dry and wet season (based on average monthly rainfall > 100 mm, Fig. S1) having maximum and minimum temperature ranging across 28 °C and 19 °C. The topography of the sanctuary gradually descends from southeast to northwest (Fig. 2C) and has numerous small streams, most of which are rainfed and usually dries out in the winter. Basically having semi-evergreen forests and evergreen patches (Fig. 2D) with good canopy contiguity, distinct three-tier forest strata with high diversity of plant species and the year-round availability of fruit-bearing trees (Bujarbarua and Sarma 2004; Sarkar and Devi 2014; Sengupta 2017), this forests patch is a host to diverse biodiversity clustered within just 20.98 km2 of area.
Survey design and data collection
The habitat use of MGS was accessed through direct (animal sightings ~ encounter rate) and indirect (nests) signs with a rationale that the presence of nests and higher animal abundance in an area reflects the quality of the habitat around it (Datta 1993; Krausman 1999; Ransome and Sullivan 1997). For this study, we assumed that all the inhabiting individuals of MGS in the absence of any physical barrier, have the dispersal potential to colonize the entire extent of the study area and thus, any observed differences in the spatial distribution and abundance pattern of these squirrels occurring at the grid level have to be related to the habitat features found therein and not to the presence of dispersal barriers hindering the accessibility of the geographical space to the species.
Squirrel sampling
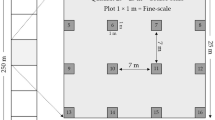
Prior to survey, vegetation maps (land use and land cover) were prepared using a supervised classification approach using Landsat TM and ETM + satellite imagery (30 m resolution) within a 1 km buffer area of the study area, HGS (Fig. 2D). This was overlaid with a 500 m × 500 m grid using the Geographical Information System (GIS) software Arc Map 10.2 (ESRI 2014), dividing the study site into 128 sampling grids. The grid size was ascertained based on the available literature on annual home range studies of giant squirrel (Ratufa genus), reporting it to be in the range of 0.82 ha and 1.2 ha in contiguous forest patches (Borges 1993; Joshua 1992), and even smaller in small, fragmented forest patches (home range compaction effect, see Koprowski 2005).
A total of 104 transects, each measuring 500 m in length were laid spatially covering each grid (some grids could not be surveyed due to logistic constraints), to collect squirrels’ habitat use data and abundance pattern relative to microsite and stand-level characteristics. Four temporal replicates of each transect were undertaken during the dry season between October 2014 and March 2015. All surveys were carried out only during fine weather days (no rain or strong winds), by walking at a slow but constant speed to minimize any potential bias and to maximize the number of detections. Survey time alternately covered two different time frames of the day (between 06:00 and 09:30 h as well as between 13:00 and 16:30 h) as it is known to be the time of peak activity of giant squirrels (Joshua 1992). Data (squirrel sightings) from replicate transect surveys (four temporal replicates) from each grid were then used to generate an ‘average encounter rate’ index for each individual sampling site by calculating the total transect distance walked and the number of squirrels sighted.
Habitat sampling
To account for the potential predictors of habitat use that influence occupancy and the abundance pattern of giant squirrels, we quantified the spatial structure of forests which in our study, refer to the relative spatial arrangement of trees and interconnections between them (see Baskent and Jordan 1996). Apart from forest structure, we also hypothesized the interactive role of forest composition and food availability to determine the same, including the effect of anthropogenic disturbances and forest edge. For this, a total of 14 ecologically meaningful covariates (Table 1) representing both the spatial characteristics and the non-spatial characteristics or composition of patches were measured in each grid using quadrat method of 20 m × 20 m size to account for the variation in occupancy pattern vis-a-vis habitat utilization pattern of giant squirrel. Quadrats were placed in the geographical center of each grid, avoiding sharp edges and forest gap areas. These habitat covariates, which include remotely sensed variables (n = 3) and field-based variables (n = 11), were collected adhering to standard methodologies (Chaturvedi and Khanna 1982; Phillips 1959) and other similar studies (Borges 1993; Herlekar 2010). All the samplings (both habitat and squirrel) were undertaken in the dry season to avoid the confounding effects of seasonal foliage change in influencing the observed distribution pattern.
Data analysis
Giant squirrel’s habitat use and their association with surrounding forest attributes or habitat covariates were tested using generalized linear models (GLM) approach with ‘average encounter rate’ estimated per grid (a surrogate measure of animal abundance) and multiplied by 100 as the response variable (n = 104); tested against the 14 ecological covariates (forest attributes) as predictor variables. We also used the Kruskal–Wallis test to examine the differences in grid-wise squirrel abundance among different habitat types including the matrix.
Since correlation test (Pearson’s correlation) among the collected ecological covariates revealed high degree of association (Table S1) with variance inflation factors (VIFs) exceeding threshold value (VIF: 5, Table S2), principal components (PCs) were extracted from the entire suite of 14 variables (Table S3) by means of a principal component analysis (PCA), a dimensionality reduction technique (Maddala 1977). This allowed the information from the original set of 14 variables to be retained, obtaining new components that are independent from each other (retaining only the components with VIF value below five), thus avoiding problems of “multi-collinearity” reflected in the early exploratory analyses.
However, before performing PCA analysis, we had transformed all the variables into a Z-score to reduce the order of magnitude and allow the numerical optimization for subsequent analytic treatments. The components (first 4) that resulted from the exploratory PCA analysis using all the 14 covariates explained a cumulative variance by 84% (Table S3). However, the pattern of loading across the components (assessed through PCA biplot) made interpretation of this PCA result difficult (Fig. S2). As such, based on ecological considerations and guided by component loadings (see Nath et al. 2019), the entire suite of ecological dataset was split into sub-groups, each one representing different aspects of forest, viz., structural complexity, composition and food abundance, disturbance and edge influence. Subsequently, four separate PCAs were recomputed for each such ecological sub-grouping respectively. We later quantified the relationship between each forest aspect (1. forest structure, 2. forest composition and food availability, 3. forest disturbance and 4. edge influence) and the response variable using exploratory scatter plot analysis and univariate regressions to choose an appropriate function (linear or polynomial) before subsequent modeling.
To evaluate the ecological correlates of squirrels’ habitat use, we built various candidate regression models (generalized linear model with a Gaussian distribution error) using forest structure and its additive as well as their interactions with other remaining forest attributes—forest composition and food availability, forest disturbance and edge influence as independent variables (considering a maximum of only two predictors at a time during each model built) and average squirrel encounter rate as the dependent variable (Table S4). We also constructed the null model (with the intercept only) and used information theoretic approach for model selection following Burnham and Anderson (2002). Model weights (Akaike weight, Wi) were computed to identify comparative explanatory power of models. We estimated regression coefficients and unconditional standard errors of each predictor using MuMIn R package (Bartoń 2020). The best model was used for visualization of predictor effects (ggplot2 R package—Wickham 2016; visreg R package—Breheny and Burcheet 2017). All statistical analyses were carried out using R 3.6.2 (R Core Team 2019).
Results
Spatial distribution pattern
The entire survey of the 104 grid cells spanning across the dry season with 4 temporal replicates in each cell, yielded a total of 182 direct sightings of giant squirrels on the line transects (total effort of 208 km transect walk) with a mean encounter rate (n/l) 0.875/km ± 1.40 (SD). However, replicate surveys failed to locate any evidence of giant squirrel presence in 60 of the sampled grid cells. We obtained significant difference in the spatial distribution pattern of the species across the studied habitat types (Kruskal–Wallis, X2 = 23.86, P = < 0.001, Fig. 3). The spatial distribution pattern of the species along with all their defined sightings during the study period is represented as an illustration (Fig. 4) indicating the distribution of suitable habitat with greater intensity of use in the study area.

Squirrel abundance across the studied habitat types in and around the study area

Sampling regime to collect data on giant squirrel habitat use and distribution pattern in and around the study area. Grid cells indicated by grid lines (500 m × 500 m) superimposed over a land use land cover map. White dots represent all the defined sighting records of R. bicolor during the study period
Ecological correlates of habitat use
The relationship between different habitat variables (14 covariates) and squirrels’ intensity of habitat use depicted using a surrogate measure of abundance, i.e., encounter rate is shown in Fig. S3. We restructured and summarized the original dataset of 14 covariates by means of a principal component analysis (PCA). The first grouping (forest structure) reflecting forest attributes related to its structural complexity (average girth size of trees, overall tree density, canopy connectivity, canopy density and average canopy height) had the first principal component explaining 81% of the variance. The second ecological grouping (forest composition and food availability) combined variables that reflected forest composition and squirrel food availability attributes (i.e., food abundance, food tree diversity, food tree species richness and liana species richness), and yielded one single principal component contributing 82% of the total variance. The third grouping (forest disturbance) summed up forest disturbance attributes (cut marks, NTFP extraction intensity) and the last ecological grouping (edge influence) summarized the edge influence (nearest distance to human habitation, navigable road and forest edge). The first principal component explained 90% and 63% of the total variance respectively, for the last two groupings. The principal component loadings of the ecological dataset with the original 14 variables are shown in Table S3. Collinearity statistics showed variance inflation factor (VIF) of the independent PCA components (the four forest aspects) with two of them exhibiting considerable, but insignificant inflation (“forest structure” is inflated by a factor of 3.6 and “forest composition and food availability” is by 3.4). Forest disturbance and edge influence revealed very less variance inflation (Table 2).
The global regression model with each of these four independent components or forest aspects (PCA sub-groupings) showed squirrel abundance with 95-fold positive relationship with “forest structure” attributes and 48-fold influence with “forest composition and food availability” attributes, with both these components contributing significantly; while insignificant negative relationship was seen with “forest disturbance” (eightfold). Edge influence, even though an insignificant one, exerted a positive influence (fourfold) on the abundance pattern of giant squirrel, implying a tendency of increased abundance with increasing remoteness of sampling site from forest edges (Table 2). Forest disturbance and edge influence components did not show up as important in explaining abundance pattern due to large unconditional standard errors for the parameter estimates. Overall, these variables contributed 62% variation to the model (R2 = 0.62).
Of all the tested candidate models with various combinations of additive and interaction effect of the four different forest aspects (Table S4), the best discrete-choice model explaining the observed variation in squirrel abundance included the interaction effects of forest structure with forest composition and food availability (Table 3). This model accounted for 76% of the AICc weight and indicated a positive linear relationship between animal abundance and the two explanatory predictors (Fig. 5). Model parameter estimates (of best model) and the 95% confidence intervals for the effects of forest structure (β = 139, SE = 9.8; 95% CI = 48–198) and forest composition and food availability (β = 131, SE = 10.2; 95% CI = 32–202) did not overlap zero (Table 4), indicating that the interaction between these variables contributed significantly to model fit.

Scatter plots showing relationship between different forest aspects (principal components, PCs) and squirrel abundance. a forest structure, b forest composition and food availability, c forest disturbance, d edge influence
Discussion
The current study examines the underlying ecological determinants of R. bicolor’s habitat selection and provides a comprehensive quantitative profile for a better understanding of their life history and habitat relations. Quite often, ‘patch quality’ has been reported to play a decisive role in regulating habitat occupancy and its intensity of use (Cudney-Valenzuela et al. 2021). However, patch quality is a complex set of ecological covariates which exerts their influence either individually, or in a synergistic manner (Umapathy and Kumar 2000). In case of arboreal mammals, their life history is intricately linked to numerous structural attributes of the forest (Kays and Allison 2001; Malcolm 2004), and this relationship can have far-reaching conservation implications as choosing the right habitat improves their survival and reproductive fitness (Cody 1985; Marsh et al. 2013). Interestingly, the current study reveals that giant squirrels select patches of usable habitat based on concentrated areas of critical habitats with conducive structural and compositional forest attributes; traits that are usually associated with old growth forest.
Spatial distribution pattern
Our study revealed a very restricted habitation of the animal in the sanctuary (Fig. 4), being absent from 60 out of 128 grid cells surveyed. Repeated surveys failed to locate any of their population in the surrounding monoculture plantations (betel nut and tea gardens) which engulfs almost the entirety of the sanctuary. The distribution pattern of the animal presents a preponderance of occupancy in forest interiors while being absent from forest edges and clearings, potentially indicating their adaptation to functionally important forest attributes. However, they were spotted foraging in home gardens around forest fringe human settlements. These seasonal forage into the adjoining areas largely depended on the availability of suitable and sufficient structures for mobility and opportunistic food resources maintained in human-modified habitats. These home gardens are primarily characterized by the presence of Ficus spp., and other fruiting tress, such as Pomelo (Citrus maxima), both of which are ubiquitous components of home gardens in this landscape.
Sporadic venture of arboreal mammals into the adjoining matrix is a function of the degree of structural contrast existing between the patch-matrix systems (see Driscoll et al. 2013). Conventional monoculture tea plantations typically lack a well-defined, structurally complex multi-tier vertical stratum which fails to integrate with the surrounding forest patches, forming an inhospitable matrix to many obligate canopy dwellers (Chowdhury et al. 2021; Raman et al. 2021). Setting up such plantations has already been established to be detrimental, restricting the movement of habitat specialist like Lion Tailed Macaque, Macaca silenus (Ramachandran and Joseph 2000). This highlights the importance of promoting and maintaining associated habitat matrix around small-fragmented forest patches (especially through structurally complex agricultural habitats like agroforestry plantations), which can serve as complimentary space for populations of giant squirrel species to disperse successfully.
Ecological correlates of habitat use
Studies of habitat selection by R. bicolor are scarce and very patchy in nature. However, there seems to be a consensus in all these studies, including the present study, concerning their obligate arboreal niche (Datta and Goyal 2008; Mackinnon 1978; Payne 1979; Saiful and Nordin 2004; Shyam and Saikia 2012). They prefer densely forested top canopy forest strata (probably helps in camouflaging the nests to avoid predation). Many arboreal mammals exhibit this trait to exploit the advantages of increased visibility for vigilance against arboreal and aerial predators by selectively preferring tall tree stands (Baldellou and Henzi 1992; Burke and Nol 1998).
Additionally, our study also explored the role of underlying site covariates within these tall stands to explain the observed variation in the intensity of habitat use by giant squirrels, and the results showed a strong influence of canopy connectivity (R2 = 0.61), canopy density (R2 = 0.57), overall tree density (R2 = 0.63) and availability of primary food trees (R2 = 0.61). All these findings reaffirm the critical importance of forest physiognomy and its spatial structure for the conservation of giant arboreal squirrels with implications on their ability to restrict or provide access to habitats (Borges 2008; Datta and Goyal 1996; Emmons and Gentry 1983; Verbeylen et al. 2009). For instance, Dodd et al. (2006) emphasized the importance of interlinking canopy crowns with canopy continuity for the persistence and recruitment of arboreal squirrel at the patch scale. Contrarily, some argue the ‘compositional’ elements of a patch to be more influential on the life history of arboreal mammals than forest structural attributes (Adler 2000; Herbers and Klenner 2007; Saiful and Nordin 2004). A study from the tropical region reports similar finding, with diversity of tree species playing a key role in providing high-quality habitat for arboreal tree squirrels (Islam 2002). However, in most studies, the distinction between attributes representing structural and compositional aspects of a forest patch remains blurred and must be addressed if we are to fully understand the species–habitat relationship of arboreal mammals.
Interestingly, our GLM results exploring the different possible combinations of composite independent factors or forest aspects (PCA reduced the 14 individual covariates into 4 composite factors) suggested the crucial interplay of forest composition and food availability (a composite of food resource availability attributes) along with forest structure as the best candidate model in explaining the observed variation within patch level habitat utilization pattern. However, no single habitat variable or suite of individual variables was able to fully explain the observed variation. This clearly demonstrates giant squirrels' sensitivity to forest composition and food availability attributes, beyond habitats’ structural complexity. However, very few studies acknowledge the synergistic action of forest aspects (e.g., canopy connectivity and food availability) in regulating arboreal mammals’ habitat use (Herbers and Klenner 2007; Nandini and Parthasarathy 2008; Ransome and Sullivan 1997; Sridhar et al. 2008). In the current study area, a significant portion of the arboreal complexity was contributed by the entangled lianas providing inter-tree physical connectivity; facilitating unrestricted movement. Apart from lianas, the old and large fig trees also provide a complex branching pattern and are known to act as a ‘fallback food’ reserve for many arboreal life forms (Borges 2008; Dunn et al. 2012; Sengupta 2017), thus providing food availability in these structurally accessible areas.
Of the other two forest aspects considered in the analysis, our study documented a negative relationship between squirrel abundance and anthropogenic disturbance, which mirror the findings generally found across the geographic distribution of the species. Mostly these studies report about populations of giant squirrels becoming sparse, scattered and eventually getting locally extirpated with increasing disturbance regime (Datta and Goyal 2008; Joshua and Johnsingh 1994; Sengupta et al. 2016; Singh et al. 2000). Persistence of anthropogenic disturbance in small isolated forest patch markedly alters the demographic, dietary, ranging and nesting patterns of arboreal mammals (Kays and Allison 2001; Whitworth et al. 2019). This, in turn, has an effect on the individual fitness and reproductive success of the species, being unable to adjust with the ongoing changes in the structure and composition of the forest, which eventually leads to a decline in their population density (Irwin 2008; Koprowski 2005; Selonen et al. 2001; Wauters et al. 1997). Similarly, edge influence, which was estimated using a surrogate index, i.e., proximity to forest edges, also revealed a negative impact with an increasing trend of squirrel abundance away from the forest clearings. This holds true, given the role of edges in modifying plant species diversity and vegetation structure, which has been reported to have mostly negative effects on populations of arboreal mammals (Laurance et al. 2000, 2002; Lehman 2007; Murcia 1995).
Overall, our analysis reveals a new perspective on the response of giant squirrels to stand-level microhabitat characteristics in a lowland tropical forest. The concordance between our findings with available pool of literature validates the importance of habitats with microhabitat features of structurally complex forest (good canopy cover and arboreal connectivity, high food tree species diversity, high basal area) as putative conservation priority sites for long-term survival and persistence of giant squirrel populations across their geographic distribution (Baskaran et al. 2011; Borges 1989; Datta 1993; Datta and Goyal 1996, 2008; Ramachandran 1988; Srinivas et al. 2008). Moreover, our study confirms the sensitive ecological requirements of giant squirrel in a fragmented landscape, along with their inability to inhabit any anthropogenic matrix which lacks structural complexity. This makes them an ideal indicator species for assessing the habitat quality and forest integrity in tropical landscape (see Koprowski 2005).
Conservation implication
Upper Brahmaputra Valley, even though a biological frontier; is a leading tea-producing region in the world. This has resulted in a significant loss of its original forest cover, transforming the landscape into a mosaic of tea plantations, agricultural land and forest fragments of various sizes. This study, conducted in a lowland tropical forest patch which lies in the northern distribution limit of R. bicolor, quantifies its microhabitat use in relation to stand-level characteristics. Our findings shows that the ecological correlates influencing habitat occupancy and the intensity of its use in a small isolated forest patch are mainly the result of the spatial distribution and frequency of occurrence of some microhabitats with which the species is closely associated. These microhabitats are shaped by a few critical structural and forest compositional attributes. Thus, maintaining these extremely subtle microhabitats within the broader habitat type will be critical in ensuring the future survival of giant squirrels in these pockets of far-flung isolated forest patches that typically suffer from reduced food availability and a possible reduction in forage quality for arboreal species (Agetsuma 1995; Taubert et al. 2018) because of the associated habitat loss and reduced density of big trees and plant species richness. Moreover, with the number of species threatened with extinction in tropical forests predicted to escalate (Vamosi and Vamosi 2008; Whitmore and Sayer 1992), the only way forward from here will be to properly manage these disturbed forests to support their populations (see Turner and Corlett 1996; Volenec and Dobson 2020). Having said that, the fitness values associated with each individual habitat feature and their relative importance to giant squirrels might vary across different spatial scale; and as such, an understanding of patch-occupancy requirements and landscape-level distribution patterns of giant squirrels should be the next focus area for research in this type of human-modified tropical landscapes.
Nevertheless, the findings of this study contribute significantly towards our understanding of the autecology of R. bicolor populations across an isolated tropical forest in its northernmost distributional range. It can also aid in the development of a comprehensive conservation action plan for the species, with strong impetus to its associated matrix land use management and habitat restoration in and around such isolated, remnant lowland rain forest fragments of the tropical landscapes.
Data availability
The raw data and the codes used are available from the corresponding author upon request.
References
Adler GH (2000) Tropical tree diversity, forest structure and the demography of a frugivorous rodent, the spiny rat (Proechimys semispinosus). J Zool 250(1):57–74
Agetsuma N (1995) Foraging strategies of Yakushima macaques (Macaca fuscatayakui). Int J Primatol 16(4):595–609
Baldellou M, Henzi SP (1992) Vigilance, predator detection and the presence of supernumerary males in vervet monkey troops. Anim Behav 43(3):451–461
Bartoń K (2020) MuMIn: multi-model inference. R Package Version 1(43):17
Baskaran N, Senthilkumar K, Saravanan M (2011) A new site record of the Grizzled Giant Squirrel Ratufa macroura (Pennant, 1769) in the Hosur forest division, Eastern Ghats, India and its conservation significance. J Threat Taxa 3(6):1837–1841
Baskent EZ, Jordan GA (1996) Designing forest management to control spatial structure of landscapes. Landsc Urban Plan 34(1):55–74
Borges RM (1989) Resource heterogeneity and the foraging ecology of the Malabar Giant Squirrel, Ratufa indica. Doctoral dissertation, University of Miami
Borges RM (1993) Figs, Malabar giant squirrels, and fruit shortages within two tropical Indian forests. Biotropica 25(2):183–190
Borges RM (2008) Why are there so many giants, including giant squirrels, in the Old World tropics? Curr Sci 95:866–870
Breheny P, Burcheet W (2017) Visualisation of regression models using visreg. R Journal 9:56–71
Bujarbarua P, Sarma SK (2004) An assessment of the floristic diversity of Gibbon wildlife sanctuary, Assam, India. Phytotaxonomy 4:124–130
Burke DM, Nol E (1998) Edge and fragment size effects on the vegetation of deciduous forests in Ontario, Canada. Nat Areas J 18(1):45–53
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: a practical information-theoretic approach, 2nd edn. Springer-Verlag, New York, pp 70–71
Chatterjee P, Tripathy B, Chandra K, Saha GK, Mondal K (2020) Climate change alarms the survival of Near Threatened species Malayan giant squirrel (Ratufa bicolour Sparrman, 1778) in India. Mammal Study 45(4):289–302
Chaturvedi AN, Khanna LS (1982) Forest mensuration. International Book Distributors, Dehradun, India
Choudhury A (1999) Primates in gibbon sanctuary Assam, India. Asian Primates 7(1–2):4–6
Choudhury A (2013) The mammals of North east India. Gibbon Books, Guwahati, India
Chowdhury A, Samrat A, Devy MS (2021) Can tea support biodiversity with a few “nudges” in management: evidence from tea growing landscapes around the world. Glob Ecol Conserv 31:1801
Cody ML (1985) Habitat selection in birds. Academic press INC, London
Cudney-Valenzuela SJ, Arroyo-Rodríguez V, Andresen E et al (2021) Does patch quality drive arboreal mammal assemblages in fragmented rainforests? Perspect Ecol Conserv 19(1):61–68
Datta A (1993) Space-use pattern of the Indian giant squirrel (Ratufa indica centralis) in relation to food availability in Bori Wildlife Sanctuary, Madhya Pradesh, India. Doctoral dissertation, M. Rajkot: Saurashtra University
Datta A, Goyal SP (1996) Comparison of forest structure and use by the Indian giant squirrel (Ratufa indica) in two riverine forests of Central India. Biotropica 28:394–399
Datta A, Goyal SP (2008) Responses of diurnal tree squirrels to selective logging in western Arunachal Pradesh. Curr Sci 95:895–902
de Castro EBV, Fernandez FA (2004) Determinants of differential extinction vulnerabilities of small mammals in Atlantic Forest fragments in Brazil. Biol Conserv 119(1):73–80
Dobrovolski R, Diniz-Filho JAF, Loyola RD et al (2011) Agricultural expansion and the fate of global conservation priorities. Biodivers Conserv 20(11):2445–2459
Dodd NL, Schweinsburg RE, Boe S (2006) Landscape-scale forest habitat relationships to tassel-eared squirrel populations: Implications for ponderosa pine forest restoration. Restor Ecol 14(4):537–547
Driscoll DA, Banks SC, Barton PS et al (2013) Conceptual domain of the matrix in fragmented landscapes. Trends Ecol Evol 28(10):605–613
Duckworth JW, Molur S (2016) Ratufa bicolor. The IUCN Red List of Threatened Species2016:e.T19377A22261810
Dunn JC, Asensio N, Arroyo-Rodríguez V et al (2012) The ranging costs of a fallback food: liana consumption supplements diet but increases foraging effort in howler monkeys. Biotropica 44(5):705–714
Eisenberg JF (1980) The density and biomass of tropical mammals. In: Soule ME, Wilcox BA (eds) Conservation Biology. Sinauer, Sunderland, MA, pp 35–55
Emmons LH, Gentry AH (1983) Tropical forest structure and the distribution of gliding and prehensile-tailed vertebrates. Am Nat 121(4):513–524
Environmental Systems Research Institute (ESRI) (2014) ArcGIS Release 10.2. Redlands, CA
Fahrig L (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. J Biogeogr 40(9):1649–1663
Franklin JF, Lindenmayer DB (2009) Importance of matrix habitats in maintaining biological diversity. Proc Natl Acad Sci 106(2):349–350
Gibbs HK, Ruesch AS, Achard F et al (2010) Tropical forests were the primary sources of new agricultural land in the 1980s and 1990s. Proc Natl Acad Sci 107:16732–16737
Hansen MC, Potapov PV, Moore R et al (2013) High-resolution global maps of 21st-century forest cover change. Science 342(6160):850–853
Hazarika R, Gupta AK (2005) Resource sharing by Hoolock Gibbon (Bunopithecus hoolock) with two primate species in Gibbon Wildlife Sanctuary, Assam, India, Conservation of Hoolock Gibbon (Bunopithecus hoolock) in Northeast India. Wildlife Institute of India, Dehradun, pp 233–257
Herbers JIM, Klenner W (2007) Effects of logging pattern and intensity on squirrel demography. J Wildl Manage 71(8):2655–2663
Herlekar IR (2010) Effect of canopy fragmentation on the patterns of habitat use of the grizzled giant squirrel Ratufa macroura in Cauvery Wildlife Sanctuary, Karnataka, India. Dissertation, Tata Institute of Fundamental Research, Bengaluru
Irwin MT (2008) Feeding ecology of Propithecus diadema in forest fragments and continuous forest. Int J Primatol 29(1):95–115
Islam S (2002) Effects of selective logging on tree species diversity, stand structure and physical environment of tropical hill dipterocarp forest of Peninsular Malaysia. Dissertation, University of Kebangsaan, Malaysia
Johnsingh AJT, Manjrekar N (2016) Mammals of South Asia, vol II. University Press, India
Joshua J (1992) Ecology of the endangered grizzled giant squirrel Ratufa macroura in Tamil Nadu, south India. Dissertation, Bharathidasan University
Joshua J, Johnsingh AJT (1994) Impact of biotic disturbances on the habitat and population of the endangered grizzled giant squirrel Ratufa macroura in South India. Biol Conserv 68(1):29–34
Kakati K, Raghavan R, Chellam R et al (2009) Status of Western Hoolock Gibbon (Hoolock hoolock) populations in fragmented forests of eastern Assam. Primate Conserv 24(1):127–137
Kays R, Allison A (2001) Arboreal tropical forest vertebrates: current knowledge and research trends. Plant Ecol 153(1–2):109–120
Koprowski JL (2005) The response of tree squirrels to fragmentation: a review and synthesis. Anim Conserv 8(4):369–376
Koprowski JL, Nandini R (2008) Global hotspots and knowledge gaps for tree and flying squirrels. Curr Sci 95:851–856
Krausman PR (1999) Some basic principles of habitat use. Grazing Behav Livest Wildl 70:85–90
Laurance WF (1991) Edge effects in tropical forest fragments: application of a model for the design of nature reserves. Biol Conserv 57(2):205–219
Laurance WF (2007) Forest destruction in tropical Asia. Curr Sci 93:1544–1550
Laurance WF (2015) Emerging threats to tropical forests, treetops risk challenges glob. Canopy Ecol Conserv 100(3):71–79
Laurance WF, Delamonica P, Laurance SG et al (2000) Rainforest fragmentation kills big trees. Nature 404(6780):836
Laurance WF, Lovejoy TE, Vasconcelos HL et al (2002) Ecosystem decay of Amazonian forest fragments: a 22-year investigation. Conserv Biol 16(3):605–618
Lehman SM (2007) Ecological and phylogenetic correlates to body size in the Indriidae. Int J Primatol 28(1):183–210
MacKinnon KS (1978) Stratification and feeding differences among Malayan squirrels. Malayan Nat J 30:591–608
Maddala GS (1977) Econometrics. McGraw Hills publication Company, New York
Malcolm JR (2004) Ecology and conservation of canopy mammals. In: Lowman MD, Rinker HB (eds) Forest canopies. Elsevier, San Diego, pp 175–211
Marsh LK, Chapman CA, Arroyo-Rodríguez V, Cobden AK, Dunn JC, Gabriel D, Ghai R, Nijman V, Reyna-Hurtado R, Serio-Silva JC, Wasserman MD (2013) Primates in fragments 10 years later: once and future goals. In: Primates in fragments. Springer, New York, pp 505–525
Mayaux P, Holmgren P, Achard F et al (2005) Tropical forest cover change in the 1990s and options for future monitoring. Philos Trans R Soc B Biol Sci 360(1454):373–384
Murcia C (1995) Edge effects in fragmented forests: Implications for conservation. Trends Ecol Evol 10(2):58–62
Myers N, Knoll AH (2001) The biotic crisis and the future of evolution. Proc Natl Acad Sci USA 98(10):5389–5392
Nandini R, Parthasarathy N (2008) Food habits of the Indian giant flying squirrel (Petaurista philippensis) in a rain forest fragment, Western Ghats. J Mammal 89(6):1550–1556
Nath A, Singha H, Haque M et al (2019) Sparrows in urban complexity: macro and micro-scale habitat use of sympatric sparrows in Guwahati City. India Urban Ecosyst 22(6):1047–1060
Payne JB (1979) Synecology of Malayan tree squirrels with special reference to the genus Ratufa. Doctoral dissertation, University of Cambridge
Phillips EA (1959) Methods of vegetation study. Henry Holt, New York, US
R Development Core Team (2019) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria
Ramachandran KK (1988) Ecology and behaviour of Malabar Giant Squirrel Ratufa indica maxima (Schreber, 1788). Report of the Project Wild 04/83. Division of Wildlife Biology, Kerala for Res Inst Peechi Kerala
Ramachandran KK, Joseph GK (2000) Habitat utilization of lion-tailed macaque (Macaca silenus) in silent valley national park, Kerala India. Primate Rep 58:17–26
Raman TS, Gonsalves C, Jeganathan P et al (2021) Native shade trees aid bird conservation in tea plantations in southern India. Curr Sci 121(2):294–305
Ransome DB, Sullivan TP (1997) Food limitation and habitat preference of Glaucomys sabrinus and Tamiasciurus Hudsonicus. J Mammal 78(2):538–549
Roth VL, Mercer JM (2008) Differing rates of macroevolutionary diversification in arboreal squirrels. Curr Sci 95:857–861
Saiful AA, Nordin M (2004) Diversity and density of diurnal squirrels in a primary hill dipterocarp forest. Malaysia J Trop Ecol 20(1):45–49
Sarkar M, Devi A (2014) Assessment of diversity, population structure and regeneration status of tree species in Hollongapar Gibbon Wildlife Sanctuary, Assam, Northeast India. Trop Plant Res 1(2):26–36
Selonen V, Hanski IK, Stevens PC (2001) Space use of the Siberian flying squirrel Ptevomys volans in fragmented forest landscapes. Ecography 24(5):588–600
Sengupta S (2017) Ecology and resource partitioning of sympatric species of squirrels in Hollangapar Gibbon Wildlife Sanctuary, Assam, India. Doctoral dissertation, Assam University, India
Sengupta S, Singha H, Deb P (2016) Ground foraging behaviour of Malayan giant squirrel (Ratufa bicolor). Curr Sci 110(12):2223–2225
Sharma N, Madhusudan MD, Sarkar P et al (2012) Trends in extinction and persistence of diurnal primates in the fragmented lowland rainforests of the Upper Brahmaputra Valley, north-eastern India. Oryx 46(2):308–311
Sharma N, Madhusudan MD, Sinha A (2014) Local and landscape correlates of primate distribution and persistence in the remnant lowland rainforests of the upper Brahmaputra valley, Northeastern India. Conserv Biol 28(1):95–106
Shyam C, Saikia P (2012) Nesting habitat characteristics and food habits of Malayan giant squirrel Ratufa bicolor (Sparrmann, 1778) in Gibbon wildlife sanctuary, Jorhat, Assam, India. Ecoscan 6:73–78
Singh M, Kumara HN, Kumar MA et al (2000) Status and conservation of lion-tailed macaque and other arboreal mammals in tropical rainforests of Sringeri Forest Range, Western Ghats, Karnataka, India. Primate Rep 58:5–16
Sridhar H, Raman TS, Mudappa D (2008) Mammal persistence and abundance in tropical rainforest remnants in the southern Western Ghats, India. Curr Sci 97:748–757
Srinivas V, Venugopal PD, Ram S (2008) Site occupancy of the Indian giant squirrel Ratufa indica (Erxleben) in Kalakad-Mundanthurai Tiger Reserve, Tamil Nadu, India. Curr Sci 95:889–894
Taubert F, Fischer R, Groeneveld J (2018) Global patterns of tropical forest fragmentation. Nature 554(7693):519–522
Thorington RW Jr, Cifelli RL (1990). In: Daniel JC, Serrao JS (eds) Conservation in developing countries: problems and prospects. Oxford University Press, Mumbai, pp 212–219
Thorington RW Jr, Koprowski JL, Steele MA et al (2012) Squirrels of the World. Johns Hopkins University Press, Baltimore, MD
Turner IM, Corlett RT (1996) The conservation value of small, isolated fragments of lowland tropical rain forest. Trends Ecol Evol 11(8):330–333
Umapathy G, Kumar A (2000) The occurrence of arboreal mammals in the rain forest fragments in the Anamalai Hills, south India. Biol Conserv 92(3):311–319
Vamosi JC, Vamosi SM (2008) Extinction risk escalates in the tropics. PLoS One 3(12):e3886
Verbeylen G, Wauters LA, De Bruyn L et al (2009) Woodland fragmentation affects space use of Eurasian red squirrels. Acta Oecologica 35(1):94–103
Volenec ZM, Dobson AP (2020) Conservation value of small reserves. Conserv Biol 34(1):66–79
Wauters LA, Somers L, Dhondt A (1997) Settlement behaviour and population dynamics of reintroduced red squirrels Sciurus vulgaris in a park in Antwerp, Belgium. Biol Conserv 82(1):101–107
Whitmore TC, Sayer JA (1992) Deforestation and species extinction in tropical moist forests. Tropical deforestation and species extinction. Chapman & Hall, London, pp 1–14
Whitworth A, Beirne C, Pillco Huarcaya R et al (2019) Human disturbance impacts on rainforest mammals are most notable in the canopy, especially for larger-bodied species. Divers Distrib 25(7):1166–1178
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York
Acknowledgements
This study was financed in part by WWF small grants programme (2013) and UGC-BSR fellowship (2013–16) during the period of first author’s (SS) doctoral research work at Assam University, Silchar (registration no. Ph.D/2094/2012). Field work at the study site was authorized by Principal Chief Conservator of Forest and Chief Wildlife Warden of Assam (vide permission letter no. WL/FG.31/Pt/2013, dated 14.08.2013). We thank Dr. Anukul Nath (Wildlife Institute of India, Dehradun, India) and Dr. Narayan Sharma (Cotton State University, Guwahati, India) for suggesting valuable reference material and also for providing crucial analytical insights. We would also like to thank Mr. Nilutpal Mahanta and our field assistants Dilip Boruah and Late Noren Bhuyan for accompanying us during the field surveys. We are also highly grateful and appreciate the time and effort that the two anonymous reviewers have dedicated to provide the valuable suggestions on our manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Handling editor: Raquel Monclús.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sengupta, S., Deb, P., Arandhara, S. et al. Ecological correlates of giant squirrel (Ratufa bicolor) microhabitat use in a lowland tropical forest: implications for matrix management. Mamm Biol 103, 107–119 (2023). https://doi.org/10.1007/s42991-022-00329-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-022-00329-0