
Abstract
Maximizing cost-efficiency of biological surveys is of great importance to assess threats and monitor changes in tropical landscapes, particularly when survey costs are high as in Neotropical small mammals. Species detectability is mostly affected by local abundance but can further vary according to the forest size in fragmented landscapes. For example, species detection in smaller fragments may be higher due to either spatial constrains or forest vertical compression. Here, trade-offs between duration of trapping sessions and the cost of obtaining accurate small mammal individual and species data are investigated across a range of forest sizes, while accounting for potential sampling bias. Using a combination of live (Sherman and wire mesh) and pitfall traps, patterns of cumulative capture–recapture rates and community composition were examined across 16 night trapping sessions in 42 transects nested within 14 insular forest fragments of different sizes, and two continuous forest sites in the Central Brazilian Amazon. Based on 40,254 trap-nights, 604 captures were recorded from 22 identifiable species. Cumulative number of captures gradually increased along the entire trapping session, and on average, nearly half of all species required more than eight trapping nights to be detected at each sampling transect. Sampling bias was responsible for increased cumulative recapture rates in smaller fragments, but did not affect cumulative capture rates, or time to first detection of each species. Instead, time to first detection was mostly positively affected by species abundance. Our results highlight the higher effectiveness of long trapping sessions (i.e. > 10 nights) to detect uncommon species, increase sample sizes and recapture rates. Overall, if small mammal species abundance is high, sampling intensity can be reduced. However, regardless of species abundance, large fragments and continuous forest sites should be more intensively sampled to reduce any potential sampling bias.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Maximizing the cost-efficiency of biological surveys is of great importance to assess threats and monitor changes in forest landscapes prone to human-induced disturbance (Gardner et al. 2008). The costs of obtaining ecological data are particularly high when either species detectability or density is low, making individuals difficult to sample (Green and Young 1993; Gu and Swihart 2004). Moreover, such sampling costs per individual can also vary according to spatial scale and habitat characteristics (Brennan et al. 2002).
In fragmented landscapes—where the original vegetation is reduced to habitat remnants—species richness typically increases with habitat amount or fragment size (Watling and Donnelly 2006; Fahrig 2013), mostly due to the occurrence of forest-dependent or specialist species in areas with higher amount of habitat or larger fragment area (Fig. 1a). These forest-specialist species generally occur at low densities (Devictor et al. 2008), hence for a given sampling effort, failure in species detection will be more frequent towards larger fragment areas (Fig. 1b). This sampling bias would lead to underestimate species richness and extinction rates. Improving sampling completeness of biological communities in larger fragments then becomes critical to derive inferences about their conservation value (Ewers and Didham 2006). Conversely, additional unaccounted factors might favor detectability. For example, sampling bias might be responsible for higher species detectability in small fragments, because spatial constrains may lead individuals to pass nearby traps more often, disturbed forest structure and reduced habitat quality may turn trapping baits more attractive, and may induce arboreal species to move into lower forest strata, thereby coming into contact with traps more often than in higher quality forest (Fig. 1b). In this way, sampling bias increases the detectability of individuals and species, so that a higher abundance and species richness are observed in either small or poor-quality fragments, further masking species responses to habitat loss and fragmentation (Haila 2002). Quantifying such bias is, therefore, important to appropriately adapt sampling designs, for example, by intensifying sampling effort at larger forest areas (Fig. 1c).

Conceptual framework used to investigate sampling bias in small mammal surveys considering a a range of fragment sizes and b variable species detectability across that range due to unaccounted factors. In c are described our expectations if sampling bias occurs across the range of fragment sizes and, if so, how to allocate sampling effort
In Neotropics, small mammals comprise the vertebrate group with the highest associated survey costs in multitaxon studies (Gardner et al. 2008). These species play several ecological roles, feeding on a wide variety of resources and acting as seed consumers and seed dispersers (Terborgh et al. 2001). Information on small mammal assemblages is, therefore, valuable to elucidate broad ecological processes, such as forest regeneration (Galetti et al. 2015) and arthropod control (Carvalho et al. 2005). To minimise survey costs and maximise the amount of data acquisition, many methodological studies have evaluated the efficiency of different trap types (i.e., mostly pitfall and live traps, such as Sherman and Tomahawk), configuration (i.e., line transect or grid), position across the vertical forest strata and trapping season (e.g., Laurance 1992; Lambert et al. 2006; Umetsu et al. 2006; Vieira et al. 2014; Santos-Filho et al. 2015; Palmeirim et al. 2019). More recently, Bovendorp et al. (2017) assessed the effects of varying sampling effort on observed species richness of small mammals across the Brazilian Atlantic Forest, demonstrating that observed species richness is highly dependent on the sampling effort used. However, studies on how to allocate sampling effort in terms of trapping duration (length of each trapping session), intensity (number and density of traps used) and frequency (the number of trapping sessions) remain scarce. Consequently, researchers frequently establish a standard and fixed effort for every sampling unit (fragment and site within a large continuous area), even when a pilot study is performed.
In cost-efficiency terms, trapping duration, intensity and frequency should be balanced considering the rate of new individuals captured–recaptured and species detected in relation to the time spent in setting each live or pitfall trap in their positions, heavy loads, and sufficient manpower to dig many holes for pitfall buckets, in addition to the time spent checking each trap at least once daily. In terms of sampling duration, small mammals are usually sampled using trapping sessions of four to eight consecutive nights (Pardini et al. 2009; Delciellos et al. 2015; Borges-Matos et al. 2016). This number of nights is thought to be sufficient to capture a reasonable number of individuals and species (Larsen 2016), while subsequent trapping nights would only increase the number of recaptured individuals. These assumptions favouring short trapping sessions (i.e., four to eight trapping nights) were rarely tested (but see Moura et al. 2008). Nevertheless, the number of trapping nights may influence the number of small mammal species and individuals. In addition to low-density species, some species are “trap-shy” (i.e., individuals avoiding traps), and thereby undetected during the first trapping nights (Moura et al. 2008). Therefore, the detection of both “trap-shy” species and those occurring at low abundances can be compromised under relatively short trapping sessions.
We aim at determining the most efficient sampling design to survey small mammal assemblages in fragmented landscapes. To do so, the number of trapping nights was related to the cost of obtaining accurate individual and species data, while accounting for potential sampling bias across the range of fragment sizes. Patterns of cumulative capture–recapture rates and community composition were analysed across 16 trapping nights. This unusually long trapping session employed here provide a rare opportunity to evaluate the sampling effort needed to maximize sample size and records of uncommon species. Over that period, we expect (1) the cumulative number of captures to increase over the first trapping week, and cumulative number of recaptures to increase over the second week; and (2) most species to be detected over the first trapping week, while only a few species should be detected over the second week, possibly comprising less abundant species. The relative effects of the length of the trapping session and fragment area were further assessed on cumulative capture-recapture rates, in addition to the influence of fragment area and species abundance on the time to first detection of a species. Finally, due to potential sampling bias, we expect (1) higher capture and recapture rates, and/or (2) shorter periods until first detection of a species in smaller fragments (Fig. 1). Implications of potential sampling bias on small mammals sampling design are discussed.
Materials and methods
Study area
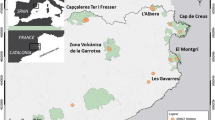
This study was carried within and around the archipelagic landscape of the Balbina Hydroelectric Reservoir, in Central Brazilian Amazonia (1°48′S, 59°29′W; Fig. 2). The reservoir was created 28 years prior to this study following the damming of the Uatumã River, a left-bank tributary of the Amazon River. In total, 312,900 ha of primary forest was flooded within the 443,772-ha hydroelectric reservoir (FUNCATE/INPE/ANEEL 2000), leaving 3546 islands corresponding to the former hilltops. Mainland continuous forest and most islands consist of dense closed-canopy terra firme forest. The local mean annual temperature is 28 °C and mean annual rainfall is ~ 2376 mm (IBAMA 1997). Small mammal assemblages were surveyed across 42 sampling transects nested in 16 sites: 14 islands and 2 sites in mainland continuous forest. Hereafter, fragment area is used to refer to the area of both islands and mainland continuous forest sites.

Location of the study area and 42 sampling transects (indicated by red dots) distributed within 14 forest islands and 2 mainland continuous forest sites in the Balbina archipelagic landscape of Central Brazilian Amazonia. The inset illustrates the sampling design of each transect: an array of three pitfall traps followed by nine stations of live traps (i.e., Sherman and wire-mesh traps), each station including three traps deployed at the forest floor, understorey and (sub)canopy. Distances between traps are indicated in the figure
Small mammal sampling
Due to their diversity of locomotion habits and diets, Neotropical small mammals are usually surveyed using different trap types deployed across the entire vertical forest strata (Larsen 2016). Here, both live and pitfall traps were deployed along linear transects during sessions of 16 consecutive nights over 2 field seasons, from April to November 2014, and from April to November 2015. Each transect consisted of a set of nine stations of live traps (hereafter, LTs), followed by an array of three pitfalls. Stations of LTs were 20-m apart, and included two Sherman traps (23 × 9 × 8 cm, H. B. Sherman Traps, Inc., Tallahassee, Florida) and one wire-mesh trap (30 × 17.5 × 15 cm, Metal Miranda, Curitiba, Paraná). At each LT station, one trap was set on the ground, one in the understory (~ 1.5 m high), and one in the (sub)canopy (> 10 m high). Live traps of different types were placed alternatively on the ground and in the understory across consecutive stations, but only Sherman traps were placed in the canopy due to logistic limitations (see Fig. 2). To set traps in canopy, an adaptation of the method described by Lambert et al. (2006) was used. LTs were baited with a mix of bananas, peanut powder, sardines and oatmeal. Pitfall traps (100L) were also spaced at 20-m intervals and connected by a plastic drift fence 50-cm high and 10-cm underground, with 10 m of fence extending beyond the two terminal pitfalls.
More sampling transects were deployed on larger islands and continuous forest sites. The number of transects on each island ranged from one to four according to the following classes of island size: 10–50 ha, 50–200 ha, 200–500 ha and > 500 ha. The number of transects in continuous forest sites (CF1 and CF2) was four and six, respectively (Table A.1). A total 40,254 trap-nights were conducted across 42 transects. All traps were inspected daily and whenever individuals could not be identified in the field, a maximum of five specimens per species per site were collected during the first trapping session, and deposited at the Mammal Collection of the Instituto Nacional de Pesquisas da Amazônia (INPA), in Manaus, Brazil. The number of collected specimens for identification purposes accounted to 14.7% of all individuals recorded. Yet, this is not expected to affect recapture rates since those are based on the number of individuals marked and released (Fig. A.1). All other individuals recorded were weighted and tagged (small mammals ear tag, size 1; National Band and Tag Co., Newport, Kentucky), so that any subsequent recaptures could be distinguished. Tissue samples from the ear were collected from all individuals recorded and deposited at INPA, but we were not always able to identify at the species-level records of Proechimys spp. (P. cuvieri and P. guyanensis) and Oecomys spp. 1 (O. roberti and O. bicolor) at all sites. Because these congeners are ecologically very similar (Jones et al. 2009), those taxa are referred as ecospecies, and hereafter species is used to refer to both species and ecospecies. The American Society of Mammalogists guidelines were followed in this study (Sikes 2016), which was further approved by the appropriate Brazilian institutional animal care and use committee (SISBIO license no. 39,187-4).
Data analysis
Individuals from the two trapping sessions were pooled together at each transect due to overall low capture rates. All data analyses were performed in R version 3.3.3 (R Development Core Team 2017).
Cumulative individual capture-recapture rates
To compare cumulative individual capture–recapture rates between the first and second trapping weeks, we performed Generalized Linear Mixed Models (GLMMs) for each trapping week separately. GLMMs were performed using the hglm R package (Rönnegård et al. 2010). The response variables were the cumulative number of captures (i.e., considering the first capture of new individuals; log10x) and recaptures (i.e., subsequent captures of the same individual; log10x), which were related to the night of the trapping session (explanatory variable). GLMMs were performed using a Gaussian error structure with an identity-link function. The distribution of each response variable was checked prior to the analysis and log-transformed whenever needed to meet the normality criteria. This procedure was further performed in the following modelling analyses. Since multiple sampling transects were grouped within islands and continuous forest sites, our data presented a nested structure. To account for such potential spatial autocorrelation (Bolker et al. 2009), site identity was considered as a random term.
First and second detection of species
The number of species detected for the first (i.e., species capture) and second time (i.e., species recapture) were quantified over the first and second trapping weeks. To do so, the bootstrapped average number of nights to the first and second detection of each species were determined. A bootstrap approach was used due to considerable variation in number of transects where each species was detected at least once. Indeed, with the exception of Echimys chrysurus for which only one individual was recorded once throughout the study, other species were detected across 2–34 transects (mean ± SD 11.6 ± 8.5). The bootstrapped number of nights to first and second detection of each species was obtained from the same number of randomly drawn transects where each species was detected at least once and detected at least twice, respectively. The average number of trapping nights was given by the average of the estimates obtained on the basis of 1000 sub-samples (mean ± SD), using the boot package within R (Canty and Ripley 2017).
Potential sampling bias
Variation in the cumulative individual capture–recapture rates is expected to be due to the night of the trapping session, depending on local species abundance. If sampling bias occurs, we further expect increased cumulative individual capture-recapture rates in decreasingly smaller patches. To account for such sampling effects, the cumulative number of captures (log10x) and recaptures (log10x) in each transect were the response variables in GLMMs with trapping night and forest fragment area (log10x) being the explanatory variables. Since increased cumulative individual capture-recapture rates can simply result from higher species abundance, species abundance (i.e., number of individuals at each sampling transect) was included as a random variable. Site identity was not included as a random variable, because any potential spatial autocorrelation was considered to be already accounted for by including local species abundance as a random variable. GLMMs were performed using a Gaussian error structure with an identity-link function. Fragment (island) area was calculated using ArcMap 10.1 (ESRI 2012), based on high-resolution multi-spectral RapidEye© imagery (5-m resolution with 5-band colour imagery) of the entire study landscape. Because fragment area could not be obtained for CF sites, values closely approximate one order of magnitude larger than our largest island were assigned to those sites.
More abundant species are expected to be detected earlier in the trapping session (date of first capture). In addition, the smaller the island, the sooner any given species is expected to be detected because of more intensive passive sampling. To determine the importance of the two above-cited sampling bias effects, the number of nights to first detection at each transect was used as response variable in GLMMs with fragment area (log10x) and species abundance (log10x) as predictors. Species and site identity were included as random variables, using a Gaussian error structure and an identity-link function. Time to first detection for individual species was then analysed in separate, by performing similar GLMMs, but considering only site identity as the random term. As a reasonable number of transects with species detection is required for statistical inference, these species-specific analyses were only possible to be carried out for nine species recorded in at least ten transects.
Results
A total of 604 captures and 535 recaptures from 22 identifiable species, representing 13 rodents and 9 marsupials, were recorded across the 42 sampled transects. This resulted in a trapping success of 2.8% and 1.5% including and excluding recaptures, respectively. The most abundant species, which also matched those most commonly recaptured, were the marsupials Marmosa demerarae, Didelphis marsupialis and Philander opossum, and the rodent Hylaeamys megacephalus, collectively accounting for 64.1% of all individuals recorded (Fig. A.1).
Cumulative individual capture-recapture rates
The cumulative number of captures increased significantly not only over the first week (βnight = 0.088 ± 0.004, P < 0.001), but also over the second week (βnight = 0.028 ± 0.003, P < 0.001). Similar results were observed in terms of recapture rates, which also increased significantly over both the first (βnight = 0.077 ± 0.005, P < 0.001) and second weeks (βnight = 0.053 ± 0.006, P < 0.001; Table A.2; Fig. 3). Cumulative captures and recaptures combining all transects further illustrate the increasing trapping success over the entire trapping session (Fig. 4). The initial increase in the cumulative number of captures is followed by an increase in the cumulative number of recaptures. Consequently, overall trapping success in the second week is more than twice that observed in the first week (Fig. 4).

Cumulative numbers captures and recaptures (log10x) analyzed (a, b) by transect, and (c) pooling all 42 transects across 16 trapping nights in the Balbina landscape. Boxplots (a, b) indicate the median, 1st and 3rd quartiles, and minimum and maximum values of cumulative number of captures and recaptures across 42 sampling transects, respectively; vertical dashed line indicates the end of the first trapping week and the beginning of the second

Average (± SD) trapping night of first detection (i.e., capture; solid black dots) and second detection (i.e., recapture; grey dots) across the trapping session of 16 nights at 42 sampled transects in Balbina landscape. The vertical dashed line indicates the end of the first trapping week and the beginning of the second
First and second detection of species
The continuous increase in number of captures over the whole trapping session goes along with the detection of different species. Eleven species were recorded over the first week, while nine species were recorded over the second week (time to first detection ± SD for each species is illustrated in Fig. 4). Recaptures were obtained for 11 species, starting on the 6th trapping night (time to first recapture ± SD for each species is illustrated in Fig. 4). Four and seven species were recaptured over the first and second weeks, respectively.
Potential sampling bias
Fragment area did not additionally affect the cumulative number of captures (Fig. 5a). Yet, cumulative number of recaptures was further significantly affected by fragment area, with more individuals being recaptured at smaller fragments (βfragment area = − 0.029 ± 0.011, P = 0.010; Fig. 5a).

Effect of fragment area (log10x) on the cumulative number of a captures (log10x) and b recaptures (log10x) across 16 trapping nights at 42 sampled transects in the Balbina landscape. Transparency colour was applied to data points to allow overlapping points to be distinguished. In b, the regression line was adjusted given the significant effect of fragment area (P < 0.05), the shaded area representing the 95% confidence region
Considering all species, time to first detection significantly decreased with increasing species abundance. Yet, time to first detection was not affected by the fragment size (Table 1). When nine species recorded in at least 10 sampling transects were considered separately, time to first detection was significantly shorter when species abundance was higher for four species: Didelphis marsupialis, Hylaeamys megacephalus, Marmosa demerarae and Proechimys sp. (Table 1).
Discussion
The unusually long trapping sessions carried out in this study allowed us to improve our understanding of small mammal capture-recapture dynamics along a range of forest fragment sizes. Contrary to our expectations, the cumulative number of captures gradually increased along the entire trapping session, and nearly half of all species on average required more than eight trapping nights to be detected at each sampling transect. Sampling bias effects were responsible for higher cumulative recapture rates observed in smaller forest fragments, but were not important for cumulative capture rates, or for the time to first detection.
How many trapping nights?
Representative sampling of small mammals depends on the total number of trap-nights carried out (Bovendorp et al. 2017). Thus, to record a reasonable number of individuals, it is necessary to balance the duration (i.e., number of trap-nights) and frequency (i.e., number of repeats) of trapping sessions. Information on the trapping success throughout the trapping session is crucial to guide efficient sampling design (Larsen 2016). Most small mammal studies are based on trapping sessions of four to eight consecutive nights that are usually repeated three or four times (Pardini et al. 2009; Delciellos et al. 2015; Borges-Matos et al. 2016), or monthly over longer periods (i.e., ≥ 1 year; Grelle 2003; Mesquita and Passamani 2012; Vieira et al. 2014), while only few studies carried out trapping sessions extending over 8–10 nights (Lambert et al. 2005; Santos-Filho et al. 2015). Our results based on 16 consecutive trapping nights, however, did not find short trapping sessions to be cost-efficient, as new individuals were recorded throughout the entire trapping session. Therefore, to obtain a meaningful sample of small mammals in relatively rich assemblages (e.g., Balbina: > 22 species), longer trapping sessions seems the more efficient strategy. This is particularly relevant when trapping success is low but species richness is high, as is often the case in the Neotropics (e.g., 5%, Woodman et al. 1996; 3.1%, Lambert et al. 2005; 2.4%, Santos-Filho et al. 2012), and if a limited number of traps is used during the trapping session.
The need of longer trapping sessions to reach representative samples is further supported by the time to first detection, which was longer than eight consecutive nights for nearly half of the 22 species. Species recorded later in the trapping session corresponded to those occurring at low abundances, as noticed from the longer time to first detection whenever species abundance was lower. Moreover, this group of rare species, which was largely detected over the second trapping week, includes arboreal Echimyidae rodents, Isothrix pagurus and Echimys chrysurus, as well as other arboreal rodents Guerlinguetus aestuans and Rhipidomys leucodactylus, which are more often captured in the understorey and canopy traps, where trapping success is lower due to the three-dimensional space (Lambert et al. 2005). Some of these species recorded later in the trapping session may also correspond to ‘trap-shy’ species, which likely avoid traps during the first nights (Moura et al. 2008).
Potential sampling bias
Small mammal species abundances tend to increase towards smaller forest fragments surrounded by either a terrestrial (Pardini et al. 2005; Vieira et al. 2009) or aquatic matrix (Glanz 1990). This negative effect of fragment size on species abundance was also observed for small mammals across the Balbina landscape, particularly for non-forest-dependent species (Palmeirim et al. 2018). Higher species abundance in smaller fragments is likely to result from the absence of predators (Terborgh et al. 1997) or density compensation in the absence of other small mammal species (Fonseca and Robinson 1990). Given that overall probability of species detection increased with species abundances, sampling bias is likely to negatively affect species detection at larger fragments where species abundance is lower. Furthermore, sampling bias could potentially favour recording higher species abundance in smaller fragments due to bait attractiveness or spatial constrains (e.g., smaller areas and vertical forest compression) therein. As we did not detect any evidence of sampling bias on the cumulative number of individuals we concluded that individuals encounter traps in large fragments as quickly as in small fragments. However, the cumulative number of recaptures increased faster in smaller fragments, suggesting that once individuals enter a trap for the first time, they tend to enter again more often in smaller fragments. This sampling bias could still be due to higher bait attractiveness in smaller fragments where habitat quality, including the availability of trophic resources, is lower (Adler and Lambert 1996). Likewise, it is possible that smaller fragments and vertical forest compression contribute to some extent for individuals to repeatedly pass near traps, which would increase the cumulative number of recaptures following prolonged trap exposure. This explanation is further supported by evidence on altered small mammal movements in fragmented habitats, which may result in either increased or decreased home ranges and use of space (Pires et al. 2002; Lira et al. 2007; Delciellos et al. 2017).
Recommendations for future studies
Given that resources available for conservation studies are usually scarce, and that strategic allocation of resources is extremely important (Cleary 2006), we recommend using appropriate sampling designs in terms of trapping session duration, frequency and intensity. Assuming the labour and financial costs of deploying live and pitfall traps, adopting longer but fewer trapping sessions could improve the cost efficiency of Neotropical small mammal surveys. This strategy is particularly adequate for studies focusing on the community-wide species composition, since it would allow experts to obtain a better cost-efficient detection of uncommon or rare species. If the focus of future studies is on population ecology, longer trapping sessions could also increase sample size and recapture rates, increasing the power of statistical analysis. Conversely, such studies may require estimates of population size relying on the assumption that populations can be considered mostly constant during the sampling period, without gains or losses, as assumed by statistical closure (reviewed in Williams et al. 2002). The longer the sampling period, the more likely are population gains and losses to occur, and this assumption could be more severely violated. Previous knowledge of the environment and species should be used to infer whether the risks of significant changes in gains and losses within a two-week sampling periods outweigh the advantages of a higher statistical power.
As time to first detection mainly depended on local species abundances, previous knowledge on the local small mammal abundance and richness, if any, should be considered when deciding the trapping session duration and number of repeats. Moreover, our results further highlight the importance of taking into account potential sampling bias noticed towards smaller fragments. One way to overcome this issue would be intensifying sampling effort at larger forest sites. So far, proportional sampling effort has been rarely adopted in previous studies evaluating the effects of habitat loss and fragmentation on small mammal assemblages (e.g., Santos-Filho et al. 2012; Delciellos et al. 2015; Borges-Matos et al. 2016). Still, adopting proportional sampling effort would be useful to account for (1) greater habitat variability of larger forest areas (Schoereder et al. 2004) and (2) apparently higher trapping success intrinsic to smaller forest areas resulting from unaccounted factors (e.g., higher bait attractiveness and spatial constrains). Additionally, local species abundance and potential sampling bias across fragmented landscapes should be further considered to improve the cost efficiency of small mammal surveys according to its specific aims.
References
Adler GH, Seamon JO (1996) Distribution of four-eyed opossum, Philander opossum (Marsupialia, Didelphidae) on small islands In Panama. Mammalia 60:91–100. https://doi.org/10.1515/mamm.1996.60.1.91
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MHH, White JSS (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol 24:127–135. https://doi.org/10.1016/j.tree.2008.10.008
Borges-Matos C, Aragón S, Silva MNF, Fortin MJ, Magnusson WE (2016) Importance of the matrix in determining small-mammal assemblages in an Amazonian forest-savanna mosaic. Biol Conserv 204:417–425. https://doi.org/10.1016/j.biocon.2016.10.037
Bovendorp RS, McCleery RA, Galetti M (2017) Optimising sampling methods for small mammal communities in Neotropical rainforests. Mamm Rev 47:148–158. https://doi.org/10.1111/mam.12088
Brennan JM, Bender DJ, Contreras TA, Fahrig E (2002) Chapter 3: Focal patch landscape studies for wildlife management: optimizing sampling effort across scales. In: Jianguo L, Taylor WW (eds) Integrating landscape ecology into natural resource management. Cambridge University Press, Cambridge, pp 68–83
Canty A, Ripley B (2017) Package ‘boot’. https://cran.r-project.org/web/packages/boot/boot.pdf. Accessed 20 June 2018.
Carvalho FMV, Fernandez FAS, Nessimian JL (2005) Food habits of sympatric opossums coexisting in small Atlantic Forest fragments in Brazil. Mamm Biol 70:366–375. https://doi.org/10.1016/j.mambio.2005.08.003
Cleary D (2006) The questionable effectiveness of science spending by international conservation organisations in the tropics. Conserv Biol 20:733–738. https://doi.org/10.1111/j.1523-1739.2006.00413.x
Delciellos AC, Vieira MV, Grelle CE, Cobra P, Cerqueira R (2015) Habitat quality versus spatial variables as determinants of small mammal assemblages in Atlantic Forest fragments. J Mamm 97:253–265. https://doi.org/10.1093/jmammal/gyv175
Delciellos AC, Ribeiro SE, Vieira MV (2017) Habitat fragmentation effects on fine-scale movements and space use of an opossum in the Atlantic Forest. J Mamm 98:1129–1136. https://doi.org/10.1093/jmammal/gyx043
Devictor V, Julliard R, Jiguet F (2008) Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 117:507–514. https://doi.org/10.1111/j.0030-1299.2008.16215.x
ESRI (2012) ArcMap 10.1. Environmental Systems Research Institute, Inc., Redlands
Ewers RM, Didham RK (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev 81:117–142. https://doi.org/10.1017/S1464793105006949
Fahrig L (2013) Rethinking patch size and isolation effects: the habitat amount hypothesis. J Biogeograph 40(9):1649–1663
Fonseca GA, Robinson JG (1990) Forest size and structure: competitive and predatory effects on small mammal communities. Biol Conserv 53:265–294. https://doi.org/10.1016/0006-3207(90)90097-9
FUNCATE, INPE, ANEEL (2000) Mapeamento por satélite das áreas inundadas por reservatórios de hidrelétricas brasileiras. Unpublished Report. Convênio FUNCATE/INPE/ANEEL, São Paulo
Galetti M, Guevara R, Galbiati LA, Neves CL, Rodarte RR, Mendes CP (2015) Seed predation by rodents and implications for plant recruitment in defaunated Atlantic forests. Biotropica 47:521–525. https://doi.org/10.1111/btp.12246
Gardner TA, Barlow J, Araujo IS, Ávila-Pires TCS, Bonaldo AB, Costa JE, Espósito MC, Ferreira LV, Hawes J, Hermandez MI, Hoogmoed M, Leite RN, Lo-Man-Hung NF, Malcolm JR, Martins MB, Mestre LAM, Miranda-Santos R, Nunes-Gutjahr A, Oveal WL, Parry LTW, Peters SL, Ribeiro-Júnior MA, Silva MNF, Silva-Motta C, Peres CA (2008) The cost-effectiveness of biodiversity surveys in tropical forests. Ecol Lett 11:139–150. https://doi.org/10.1111/j.1461-0248.2007.01133.x
Glanz WE (1990) Neotropical mammal densities: how unusual is the community on Barro Colorado Island, Panama. In: Gentry AH (ed) Four neotropical rainforests. Yale University Press, New Haven, pp 287–313
Green RH, Young RC (1993) Sampling to detect rare species. Ecol Appl 3:351–356. https://doi.org/10.2307/1941837
Grelle CEV (2003) Forest structure and vertical stratification of small mammals in a secondary Atlantic forest, southeastern Brazil. Stud Neotrop Fauna Environ 38:81–85. https://doi.org/10.1076/snfe.38.2.81.15926
Gu W, Swihart RK (2004) Absent or undetected? Effects of non-detection of species occurrence on wildlife–habitat models. Biol Conserv 116:195–203. https://doi.org/10.1016/S0006-3207(03)00190-3
Haila Y (2002) A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecol Appl 12:321–334. https://doi.org/10.1890/1051-0761(2002)012[0321:ACGOFR]2.0.CO;2
IBAMA (1997) Plano de manejo fase 1: Reserva Biológica do Uatumã. Eletronorte Ibama. Brasília/DF. https://www.icmbio.gov.br/portal/images/stories/docsplanos-de-manejo/rebio_uatuma_pm.pdf. Accessed 10 Sep 2017
Jones KE et al (2009) PanTHERIA: a species-level database of life history, ecology, and geography of extant and recently extinct mammals. Ecology 90:2648. https://doi.org/10.1890/08-1494.1
Lambert TD, Malcolm JR, Zimmerman BL (2005) Variation in small mammal species richness by trap height and trap type in southeastern Amazonia. J Mamm 86:982–990. https://doi.org/10.1644/1545-1542(2005)86[982:VISMSR]2.0.CO;2
Lambert TD, Malcolm JR, Zimmerman BL (2006) Amazonian small mammal abundances in relation to habitat structure and resource abundance. J Mamm 87:766–776. https://doi.org/10.1644/05-MAMM-A-261R1.1
Larsen TH (2016) Core standardized methods for rapid biological field assessment. Conservation International, Arlington
Laurance WF (1992) Abundance estimates of small mammals in Australian tropical rainforest: a comparison of four trapping methods. Wildl Res 19:651–655. https://doi.org/10.1071/WR9920651
Lira PK, Fernandez FAS, Carlos HSA, Curzio PL (2007) Use of a fragmented landscape by three species of opossum in south-eastern Brazil. J Trop Ecol 23:427–435. https://doi.org/10.1017/S0266467407004142
Mesquita AO, Passamani M (2012) Composition and abundance of small mammal communities in forest fragments and vegetation corridors in Southern Minas Gerais, Brazil. Rev Biol Trop 60:1335–1343
Moura MC, Grelle CEV, Bergallo HG (2008) How does sampling protocol affect the richness and abundance of small mammals recorded in tropical forest? An example from the Atlantic Forest, Brazil. Neotrop Biol Cons 3:51–58
Palmeirim AF, Benchimol M, Vieira MV, Peres CA (2018) Small mammal responses to Amazonian forest islands are modulated by their forest dependence. Oecologia 187:191–204. https://doi.org/10.1007/s00442-018-4114-6
Palmeirim AF, Benchimol M, Peres CA, Vieira MV (2019) Moving forward on the sampling efficiency of neotropical small mammals: insights from pitfall and camera trapping over traditional live trapping. Mamm Res. https://doi.org/10.1007/s13364-019-00429-2
Pardini R, Souza SM, Braga-Neto R, Metzger JP (2005) The role of forest structure, fragment size and corridors in maintaining small mammal abundance and diversity in an Atlantic forest landscape. Biol Conserv 124:253–266. https://doi.org/10.1016/j.biocon.2005.01.033
Pardini R, Faria D, Accacio GM, Laps RR, Mariano-Neto E, Paciencia ML, Dixo M, Baumgarten J (2009) The challenge of maintaining Atlantic forest biodiversity: a multi-taxa conservation assessment of specialist and generalist species in an agro-forestry mosaic in southern Bahia. Biol Conserv 142:1178–1190. https://doi.org/10.1016/j.biocon.2009.02.010
Pires AS, Lira PK, Fernandez FAS, Schittini GM, Oliveira LC (2002) Frequency of movements of small mammals among Atlantic Coastal Forest fragments in Brazil. Biol Conserv 108:229–237. https://doi.org/10.1016/S0006-3207(02)00109-X
R Development Core Team (2017) A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Austria
Rönnegård L, Alam M, Shen X (2010) The hglm package (Version 2.0). https://cran.r-project.org/web/packages/hglm/hglm.pdf. Accessed 20 June 2018.
Santos-Filho MD, Peres CA, Da Silva DJ, Sanaiotti TM (2012) Habitat patch and matrix effects on small-mammal persistence in Amazonian forest fragments. Biodivers Conserv 21:1127–1147. https://doi.org/10.1007/s10531-012-0248-8
Santos-Filho MD, Lázari PRD, Sousa CPFD, Canale GR (2015) Trap efficiency evaluation for small mammals in the southern Amazon. Acta Amaz 45:187–194. https://doi.org/10.1590/1809-4392201401953
Schoereder JH, Galbiati C, Ribas CR, Sobrinho TG, Sperber CF, De Souza O, Lopes-Andrade C (2004) Should we use proportional sampling for species—area studies? J Biogeogr 31:1219–1226. https://doi.org/10.1111/j.1365-2699.2004.01113.x
Sikes RS (2016) 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J Mamm 97:663–688. https://doi.org/10.1093/jmammal/gyw098
Terborgh J, Lopez L, Nuñes PV, Rao M, Shahabuddin G, Orihuela G, Riveros M, Ascanio R, Adler GH, Lambert TD, Balbas L (2001) Ecological meltdown in predator-free forest fragments. Science 294:1923–1926. https://doi.org/10.1126/science.1064397
Terborgh J, Lopez L, Tello S (1997) Bird communities in transition: the Lago Guri islands. Ecology 78:1494–1501. https://doi.org/10.1890/0012-9658(1997)078[1494:BCITTL]2.0.CO;2
Umetsu F, Naxara L, Pardini R (2006) Evaluating the efficiency of pitfall traps for sampling small mammals in the Neotropics. J Mamm 87:757–765. https://doi.org/10.1644/05-MAMM-A-285R2.1
Vieira ALM, Pires AS, Nunes-Freitas AF, Oliveira NM, Resende AS, Campello EFC (2014) Efficiency of small mammal trapping in an Atlantic Forest fragmented landscape: the effects of trap type and position, seasonality and habitat. Braz J Biol 74:538–544. https://doi.org/10.1590/bjb.2014.0075
Vieira MV, Olifiers N, Delciellos AC, Antunes VZ, Bernardo LR, Grelle CE, Cerqueira R (2009) Land use vs. fragment size and isolation as determinants of small mammal composition and richness in Atlantic Forest remnants. Biol Conserv 142:1191–1200. https://doi.org/10.1016/j.biocon.2009.02.006
Watling JI, Donnelly MA (2006) Fragments as islands: a synthesis of faunal responses to habitat patchiness. Conserv Biol 20:1016–1025. https://doi.org/10.1111/j.1523-1739.2006.00482.x
Williams BK, Conroy MJ, Nichols JD (2002) Analysis and management of animal populations. Academic, San Diego
Woodman N, Timm RM, Slade NA, Doonan TJ (1996) Comparison of traps and baits for censusing small mammals in Neotropical lowlands. J Mamm 77:274–281. https://doi.org/10.2307/1382728
Acknowledgements
We thank all 15 volunteers/field assistants who assisted in the field; the Reserva Biológica do Uatumã and its staff for logistical support; M.N.F. da Silva for help with species identification. This study was funded by the Amazon Region Protected Areas Program (ARPA); Amazonas Distribuidora de Energia S.A.; Associação Comunidade Waimiri Atroari; Rufford Foundation (Grant number 13,675-1); Idea Wild. AFP was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) scholarship; MVV was funded by Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ CNE 201.344/2014) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq Grant 308.974/2015-8).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling editor: Heiko Rödel.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Palmeirim, A.F., Peres, C.A. & Vieira, M.V. Optimizing small mammal surveys in Neotropical fragmented landscapes while accounting for potential sampling bias. Mamm Biol 100, 81–90 (2020). https://doi.org/10.1007/s42991-020-00012-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-020-00012-2