
Abstract
Determining the optimum dose of a mutagen that causes a high frequency of favourable mutations with minimum damage to the plant is the vital criterion for the success of a mutation breeding programme. We evaluated the comparative effectiveness, radiosensitivity and relative biological effectiveness (RBE) of three physical mutagens, viz., electron beams, X-rays and gamma-rays, on twenty indigenous aromatic Joha rice cultivars of Assam. We exposed dry uniform seeds of 12.4% moisture content to 16 different mutagens doses ranging from 50 to 800 Gy at an interval of 50 Gy. Key results: Germination percentage and the seedling traits registered a decreasing trend towards increasing the mutagen doses in all the cultivars, showing differential radiosensitivities. The LD50 values ranged from 515 to 615 Gy for electron beams and 421 to 537 Gy for X-rays and 414 to 481 Gy for gamma-rays. The GR50 values based on seedling vigour index I ranged from 125 to 304 Gy for electron beams, 191 to 313 Gy for X-rays and 178 to 267 Gy for gamma-rays. The optimum dose ranges for electron beams, X-rays and gamma-rays are 241 to 337 Gy, 228 to 324 Gy and 250 to 346 Gy, respectively. The cultivar Soru Joha-Tinsukia was the most radio tolerant, followed by Keteki Joha and Local Joha, while the cultivar Kon Joha-Moran showed the least tolerance. Joha rice cultivars showed more radiosensitivity for X-rays, followed by electron beams and gamma-rays. Similarly, a high RBE was evident for X-rays, followed by electron beams and gamma-rays, indicating their decreasing penetration capacity and lethality trend. These results would help plant breeders select the appropriate mutagen dose for mutation breeding in Assam’s least researched aromatic Joha rice.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Joha rice is a unique class of aromatic rice grown as winter rice in Assam. It is trendy and highly valued due to its quality. Joha rice possesses a superfine kernel, unique aroma, better cooking properties and excellent palatability (Das et al. 2010). Joha rice cultivars have tall, weak culms, low tillering habits, poor yielding ability and susceptibility to pests and diseases. The agricultural productivity of Joha rice is the lowest with the demand for production, and the gap is increasing over time. Further, the limited scope for yield improvement of Joha rice through cross-breeding with non-aromatic cultivars (Rutger 1983; Bourgis et al. 2008; Pathirana et al. 2009) demands mutation breeding an effective tool for desirable changes without losing its quality characteristics (Pathirana 2011). Mutations have played a mammoth role in increasing world food security by contributing significantly to the augmentation of crop production (Kharkwal and Shu 2009; Srivastava et al. 2011).
The chief advantage of mutation breeding is improving a single feature in a variety without altering the other desirable agronomic traits. Crop improvement through induced mutagenesis has been most successful in utilizing ionizing radiations. Induced mutation using either physical or chemical mutagen is one way of creating variation in crop plants. The physical mutagens comprise ionizing radiation, particulate (alpha rays, beta rays, fast neutrons and thermal neutrons) and non-particulate, also called electromagnetic radiation (X-rays and gamma-rays). The works of Muller (1927) and Stadler (1928) paved the way for a new field of genetics and plant breeding called induced mutagenesis (Ahloowalia et al. 2004). According to Kovacs and Keresztesa (2002), gamma-rays are the most potent among the physical mutagens such as alpha- and beta rays. Identifying the most effective and efficient mutagen dose is essential to recovering a high frequency and spectrum of beneficial mutations. The success of mutation breeding dramatically depends on the mutation rate, the number of screened plants and the mutation efficiency. Lethal dose 50 (LD50), the specific dose of radiation killing 50% of the test material, is the optimum dose that causes a high frequency of favourable mutations with minimum damage to the plant. Before an experiment on induced modifications starts, fixation of LD50 is crucial as it varies with the biological materials, nature of treatment and subsequent environmental conditions. Determination of optimum dose, radiosensitivity and treatment conditions are essential for genetic manipulation through induced mutation.
Materials and methods
Location of the experiment
The present study was carried out in 2020 at the laboratory of the Department of Plant Breeding and Genetics, College of Agriculture, Assam Agricultural University, Jorhat.
Plant material
Twenty indigenous Joha rice cultivars of Assam (Table 1) obtained from the Department of Plant Breeding and Genetics, Assam Agricultural University, Jorhat, were exposed to various doses of three physical mutagens, namely, electron beams, X-rays and gamma-rays for the assessment of radiation-induced injuries to germination and seedling growth.
Mutagen treatments
Dry uniform healthy seeds of 12.4% moisture content were used for mutagen treatments. The doses irradiated ranged from 50 to 800 Gy at an interval of 50 Gy for each mutagen. The control treatments comprised the non-irradiated seeds of the twenty cultivars. Fifty seeds were used per dose. Gamma irradiation was done from a Cobalt-60 source in the gamma chamber at Bhabha Atomic Research Centre (BARC), Mumbai, India. Seeds were exposed to 9.0 MeV electron beams and 7.3 MeV X-rays at the agricultural radiation processing facility at Electron Beam Centre of Raja Ramanna Centre for Advanced Technology, Indore, India.
Experimental methods
We used folded rectangular sheets (15 cm × 30 cm) of Whatman no. 1 filter paper, each fold measuring 3 cm in width. Twenty-five small holes were made inside the folded sheets’ trenches and placed in germination racks. Seeds were identified with the embryonic portions facing the holes (Plate 1), and the frames were placed in plastic trays with 3–4 cm of water in them, kept at room temperature (Tmax, 26.8 °C). The layout was a completely randomized design with two replications.

Paper folder technique adopted for germination and seedling growth
Observations
The sample size for the seedling traits was a random sample of five seedlings per replication.
Germination percentage
At seven days after sowing (DAS), the seeds with the emergence of coleoptile and radicle above 1 mm length (Lee et al. 1998) were counted and expressed as a percentage of the total number sown (25).
Seedling growth parameters
Seedlings were separated carefully from the folded sheets and wiped properly at 15 DAS, and observed for shoot length (cm), root length (cm), total seedling length (cm), fresh seedling weight (g) and dry seedling weight (g). The estimate of seedling vigour used the formulae of Abdul-baki and Anderson (1973) as follows:
Statistical analysis
Mean data for germination and seedling traits were subjected to analysis of variance as per three-factor completely randomized design (Singh and Chaudhary 1985) in Windostat version 9.2 (http://www.windostat.org). The model for one environment is
where yijkl = observation of the ijklth plot, μ = the overall mean, mi = effect of the ith mutagen, gj = effect of the jth genotypes, dk = effect of the kth dose, (mg)ij = interaction effect of the ith mutagen and the jth genotype, (md)ik = interaction effect of the ith mutagen and the kth dose, (gd)jk = interaction effect of the jth genotype and the kth dose, (mgd)ijk = interaction effect of the ith mutagen, jth genotype and kth dose, and eijkl = error associated with ijklth plot.
Determination of lethal dose 50 (LD50) and growth reduction 50 (GR50)
Probit analysis is a specialized regression model of binomial response variables. The idea of probit analysis was initially proposed by Bliss (1934) and described by Finney (1952), in which the sigmoid dose–response curve is transformed into a straight line. Probit analysis has been an extensively used statistical procedure for understanding the dose–response relationships. Regression is a method of finding the best-fit line to a data set to establish the relationship between the response variable (Y) and the independent variable (X).
where a = y-intercept, b = the slope of the line and e = error term.
Mean data on germination and seedling growth were first expressed as per cent of the control (0 doses). These percentages were transformed into probits using the probit transformation table (Finney 1952). The transformed data on log doses (X-variable) and probits (Y-variable) were subjected to regression analysis in MS Excel 2007 to work out the y-intercept (a) and the slope (b) of the line. The LD50/GR50 was calculated as follows:
Results
Radiosensitivity of the Joha rice cultivars
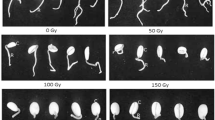
The effective doses of the electron beams, X-rays and gamma-rays for mutation induction in the twenty Joha rice cultivars were evaluated based on different growth parameters using the paper folder technique (Plate 1). Plates 2, 3 and 4 depict seedling length for representative Joha rice cultivars exposed to the various mutagens. The LD50 and GR50 values are routinely determined in any mutation breeding programme to identify the appropriate doses for different genotypes. The analyses of variance for all the traits revealed highly significant differences among the cultivars, mutagens and doses (Table 2). All the characteristics showed highly substantial variations for the cultivars x mutagen, cultivars x doses and cultivars x mutagens x doses interaction components.

Seedling length of representative Joha rice cultivar exposed to electron beam

Seedling length of representative Joha rice cultivar exposed to X-rays

Seedling length of representative Joha rice exposed to Gamma-rays
Lethal dose 50 (LD50) of the cultivars based on germination at 7 DAS
Germination percentage with increasing doses showed a linear decrease, significantly different from the control (Fig. 1). The LD50 values for the twenty cultivars exposed to varying doses of the electron beam, X-rays and gamma-rays are shown in Figs. 2a, 2b. The LD50 values ranged from 367 Gy (Kon Joha 2) to 507 Gy (Soru Joha-Tinsukia) for electron beams and 300 Gy (Kon Joha-Moran) to 467 Gy (Joha-Golaghat) for X-rays, and 381 Gy (Kon Joha-Moran) to 560 Gy (Soru Joha-Tinsukia) for gamma-rays.

Graphical representation of mean germination percentages of 20 indigenous Joha rice cultivars using electron beams, X-rays and gamma-rays


LD50 curves of the twenty Joha rice cultivars exposed to electron beams, X-rays and Gamma-rays
GR50 of the cultivars based on seedling height reduction
The dose of 50% growth reduction (GR50) was calculated based on seedling height reduction expressed as a percentage of control. The seedling length gradually decreased with increasing doses of the electron beam, X-rays and gamma-rays compared to control (Fig. 3). The maximum reduction was visible at higher concentrations of the mutagens. The GR50 doses for electron beams, X-rays and gamma-rays varied from 138 Gy (Joha-Golaghat) to 315 Gy (Kali Jeera), 192 Gy (Kon Joha-Moran) to 301 Gy (Ronga Joha) and 185 Gy (Kon Joha-Moran) to 291 Gy (Local Joha), respectively (Fig. 4).

Graphical representation of seedling length at 15 DAS

Graphical representation of LD50, GR50 and average of the cultivars
In contrast, in X-rays and gamma-rays, GR50 on the vigour index ranged from 191 Gy (Jeera Joha) to 313 Gy (Kon Joha-Teok) and 178 Gy (Kon Joha 4) to 267 Gy (Kon Joha-Teok), respectively (Fig. 5a, 5b). The optimized doses of the mutagens would help initiate a large-scale mutation breeding programme in Joha rice cultivars to bring about a broad spectrum of mutations for selection.


GR50 of the twenty indigenous Joha rice cultivars based on vigour index 1
Radiosensitivity behaviour of the cultivars for various mutagens
The cultivar Soru Joha-Tinsukia showed the highest LD50 and GR50 value, followed by Keteki Joha and Local Joha. In contrast, Kon Joha-Moran showed the lowest value (Fig. 4), indicating that Soru Joha-Tinsukia is radio tolerant compared to the other cultivars. Variations in the physical appearance of seeds, leaf characteristics and growth habits may play a vital role in creating differences in mutagen doses. Soru Joha-Tinsukia possesses long-lender grains, while Kon Joha-Moran has short-bold grains. The average Pearson correlation coefficients of LD50 based on germination and GR50 based on vigour index and seedling length of the three mutagens with thousand seed weight (g) ranged from 0.32 to 0.41, indicating greater radio tolerance of the genotypes with larger seed size. Thus, Soru Joha-Tinsukia with a large grain size could be a reason for its radio tolerance (Fig. 6 and Plate 5).

Graphical representation of grain length and breadth of 20 Joha rice cultivars

Grain/kernel characteristics of the 20 indigenous Joha rice cultivars of Assam
Relative biological effectiveness (RBE) of various mutagens
The RBE is an empirical value that depends on various types of radiation, the energy involved and the biological effects. The quality for estimating the RBE (IAEA/ICRU 2008), gamma-rays (60Co) are the reference mutagen because of their predominant use in plant mutation breeding. Similarly, the RBE of the electron beams, X-rays and gamma-rays was calculated for the twenty cultivars based on LD50 doses by considering the gamma-rays as reference (Table 3). A higher RBE was evident for X-rays, followed by electron beams for all the cultivars.
Discussion
The analyses of variance for all the traits revealed highly significant differences among the cultivars, mutagens and doses. All the seedling growth traits, viz., shoot length, root length, seedling length, fresh seedling weight, dry seedling weight, vigour index I and vigour index II registered a decreasing trend towards increasing the doses of the electron beams, X-rays, and gamma-rays in all the cultivars, with differential radiosensitivity among the cultivars (Fig. 7). Gamma-ray doses generally caused higher radiation injury than the doses of electron beams and X-rays. The findings were like the results obtained by Gowthami et al. (2016), who reported differential responses of the varieties irradiated with different doses of electron beams and gamma-rays. The lethal dose 50 is the optimum dose that causes a high frequency of favourable mutations with minimum damage to the plant and is the radiation dose that kills 50% of the test material (Jency et al. 2016). Since the LD50 value varies with the biological materials, the nature of mutagens and existing environmental conditions (Babaei et al. 2010), its determination becomes inevitable before the bulk mutagenesis of a particular target. Besides a 50% reduction in the survival of the initial target population, LD50 restricts the growth of the surviving fraction. It thus becomes a limitation in obtaining the expected number of mutants, suggesting a significant influence of the mutagens on germination. Similar observations were reported in previous studies on rice by Sareen and Koul (1999), Cheema and Atta (2003), Harding et al. (2012) and Ramchander (2015) for gamma-rays; Gowthami et al. (2016) for gamma-rays and electron beams; and Promnart et al. (2018) for electron beam. Ussuf et al. (1974) assigned the seedling growth reduction following mutagen treatments to physiological injuries, biochemical disturbances and changes in ascorbic acid content. The mutagen effects on the physiological system cause a decrease in root and shoot length (Gaul 1977). In general, the lower doses of the mutagens exhibited stimulatory effects on seedling growth parameters of the cultivars. Zaka et al. (2004) supplemented that the stimulatory effects of irradiation on seedling growth have been due to increased cell division rates and activation of growth hormones. Low-dose irradiation of seeds frequently leads to a range of positive growth effects, such as accelerated germination, increased biomass and development, and enhanced immune responses and tolerance to stress (Pishenin et al. 2021). Ariraman et al. (2014) attributed the reduction in germination at higher doses of the mutagen to disturbances at the cellular level. Seedling height has been a widely used index in determining the biological effects of various physical and chemical mutagens in M1 generation (Konzak et al. 1972). The cultivars exhibited variable responses for the seedling length at different doses of the mutagens. However, seedling length did not increase or decrease in a definite pattern in response to the various electron beam and X-ray doses. Cheema and Atta (2003), Harding et al. (2012) and Kadhimi et al. (2016) also observed that the seedling height in rice decreased with the increasing irradiation doses, the reduction being nonlinear with the increase in mutagen doses, in line with the present findings. However, the dose-seedling length relationship for gamma-rays was linear, as also reported by Wang et al. (1995) and Katoch et al. (1992) for the dependency of seedling height on the doses of physical and chemical mutagens. In agreement with the above, Chauhan et al. (2019) noted a linear dose-dependent relationship for shoot length, root length, total seedling length and vigour index using proton beam irradiation. Since seedling vigour reflects the potential level of activity and performance of the seed or seed lot during germination and seedling emergence, which represents the total of all those properties of the seed (Perry 1978), GR50 doses based on the vigour index would represent optimum doses in mutagenesis experiments. In our study, GR50 values based on the seedling vigour index ranged from 125 Gy (Joha-Golaghat) to 304 (Kali Jeera) Gy in the electron beam. The recovery of a high frequency and range of beneficial mutations, identification of the most effective mutagenic treatment and efficient mutagens are crucial (Jency et al. 2016). LD50 and GR50 for the electron beams, X-rays and gamma-rays might vary with the genotypes due to their differential radiosensitivities. According to van Harten (1998), a 20–30% growth reduction corresponding to a survival rate of 70–80% might produce an optimal mutation yield in cereal crops. Shu et al. (1996) reported a genotype-dependent variation for radiosensitivity to mutagens. Mutagen sensitivity of biological material could be attributed to the level of differentiation and development of the embryo at the time of treatment and the extent of damage to growth processes very similar rate of cell division, cell elongation, various stages of hormone and biosynthetic pathways (Zhu et al. 2008). The optimum dose determination for potent physical mutagens would aid in utilizing these mutagens to improve aromatic rice through mutation breeding programmes. LD50 and GR50 values obtained for twenty aromatic Joha rice cultivars in the present study would be beneficial in determining the range of optimum doses of mutagens. The optimum dose ranges for electron beams, X-rays and gamma-rays are 241 to 337 Gy, 228 to 324 Gy and 250 to 346 Gy, respectively. Amounts between these ranges would be helpful for irradiation of aromatic rice seeds to get the maximum number of valuable mutants with minimum damage to plant survival. The biological tissues deposit the different radiation energy in different ways, which affects the quantity of cellular damage. RBE refers to the ratio of the doses required by two radiations to cause the same effect on biological tissue by depositing per unit of energy (Jones 2015). Similarly, Rani et al. (2016) for X-rays and Sao et al. (2020) for the electron beams, X-rays, gamma-rays and proton beams in rice and Mondal et al. (2017) for electron beam in groundnut reported more than one RBE in tune with the present findings. RBE proceeds as a function of linear energy transfer (LET), which is the energy shifted to the target material per unit length of the track. As the LET increases, the RBE value slowly reaches the maximum and falls due to cell overkill (Willers et al. 2018). Therefore, this study reported higher RBE and LET of X-rays, indicating their profound penetration ability and lethal effects on biological tissues. The present study revealed that the short aromatic Joha rice cultivars show more radiosensitivity for X-rays followed by electron beam and gamma-rays. Also, of the cultivars taken under investigation, Soru Joha-Tinsukia is radio tolerant compared to the other cultivars, owing to its genotypic attributes and genetic constituents. The comparison of radiosensitivity behaviour for the three mutagens mentioned above for aromatic Joha rice makes it a unique study. In future, we can use the optimized doses for all three types of physical mutagens in fragrant rice to conduct mutation breeding programmes and compare the mutagen efficiency and effectiveness in obtaining the optimum number of mutants in the field. In addition, gamma-rays, electron beams and X-rays can also have the utility of creating genetic variability and novel mutants in aromatic rice.

Variation in radiation injury for the seedling traits of the Joha rice cultivars caused by the electron beams, X-rays and gamma-rays
References
Abdul-Baki AA, Anderson JD (1973) Vigor determination in soybean seed by multiple criteria 1. Crop Sci 13(6):630–633
Ahloowalia BS, Maluszynski M, Nichterlein K (2004) Global impact of mutation derived varieties. Euphytica 135(2):187–204
Ariramana M, Gnanamurthy S, Dhanavelb D, Bharathi T, Murugan S (2014) Mutagenic effect on seed germination, seedling growth and seedling survival of Pigeon pea (Cajanus cajan (L.) Millsp). Inter Let Nat Sci 21:41–49
Babaei A, Nematzadeh GA, Avagyan V, Hamid Reza S, Petrodi H (2010) Radio sensitivity studies of the morpho-physiological characteristics in some Iranian rice varieties (Oryza sativa L.) in M1 generation. Afr J Agril Res 5:2124–2130
Bliss CI (1934) The method of probits. Sci 79(2037):38–39. https://doi.org/10.1126/science.79.2037.38
Bourgis F, Guyot R, Gherbi H, Tailliez E, Amabile I, Salse J, Lorieux M, Delseny M, Ghesquière A (2008) Characterization of the major fragrance gene from an aromatic japonica rice and analysis of its diversity in Asian cultivated rice. Theo App Gen 117:353–368
Chauhan A, Kumar V, Iyer PR, Vishwakarma G, Nair JP, Surendran P, Sparrow H, Gupta AK, Shitre AS, Shinde AK, Kunkerkar RL, Sharma D, Patel RRS, Das BK (2019) Effect of proton beam irradiation on survival and seedling growth parameters of Indian rice (Oryza sativa L.) variety ‘Indira Barani Dhan 1.’ Elect J P Breed 10(2):490–499
Cheema AA, Atta BM (2003) Radiosensitivity studies in basmati rice. Pak J Botany 35(2):197–207
Das A, Kesari V, Rangan L (2010) Aromatic Joha rice of Assam–a review. Agril Rev 31(1):1–10
Finney DJ (ed) (1952). Cambridge University Press, Cambridge
Gaul H (1977) Mutagen effects observable in the first generation. 5.1. Plant injury and lethality, 5.2. Cytological effects, 5.3. Sterility In: Manual on mutation breeding. Second Edition. Technical Reports Series No. 119, IAEA, Vienna, Austria, pp. 87–96
Gowthami R, Vanniarajan C, Souframanien J, Arumugam Pillai M (2016) Effect of gamma rays and electron beam on various quantitative traits of rice (Oryza sativa L.) in M1 Generation. Adv Life Sci. 5(5):1876–1882
Harding SS, Johnson SD, Taylor DR, Dixon CA, Turay MY (2012) Effect of gamma rays on seed germination, seedling height, survival percentage and tiller production in some rice varieties cultivated in Sierra Leone. J Expt Agri Intl 2(2):247–255
IAEA/ICRU (2008) Relative biological effectiveness in Ion beam therapy. Jointly sponsored by the international atomic energy agency and the international commission on radiation units and measurements. Technical Rep Series. 461:14.
Jency JP, Ravikesavan R, Sumathi P, Raveendran M (2016) Determination of lethal dose and effect of physical mutagen on germination percentage and seedling parameters in kodomillet variety CO 3. Elect J Plant Breed 7(4):1122–1126
Jones B (2015) A simpler energy transfer efficiency model to predict relative biological effect for protons and heavier ions. Front Oncol 5:184
Kadhimi AA, Alhasnawi AN, Isahak A, Ashraf MF, Mohamad A, Yusoff WMW, Zain CRCM (2016) Gamma radiosensitivity study on MRQ74 and MR269, two elite varieties of rice (Oryza sativa L.). Life Sci J 13(2):85–91
Katoch PC, Massar JE, Plaha P (1992) Effect of gamma irradiation on variation in segregating generations of F2 seeds of rice. Ind J Gen Plant Breed 52(3):213–218
Kharkwal MC, Shu QY (2009) The role of induced mutations in world food security. Induced plant mutations in the genomics era. Food and Agriculture Organization of the United Nations, Rome 33–38
Konzak CF, Wickham IM, Dekock MJ (1972) Advances in methods of mutagen treatment. In: Induced mutations and plant improvement
Kovacs E, Keresztes A (2002) Effect of gamma and UV-B/C radiation on plant cells. Micron 33(2):199–210
Lee SS, Kim JH, Hong SB, Yun SH (1998) Effect of humidification and hardening treatment on seed germination of rice. Kor J Crop Sci 43(3):157–160
Mondal S, Petwal VC, Badigannavar AM, Bhad PG, Verma VP, Goswami SG, Dwivedi J (2017) Electron beam irradiation revealed genetic differences in radio-sensitivity and generated mutants in groundnut (Arachis hypogaea L.). Appl Radiat Isot 122:78–83
Muller HJ (1927) Artificial Transmutation of the Gene. Sci 66:84–87
Pathirana R, Vitiyala T, Gunaratne NS (2009) Use of induced mutations to adopt aromatic rice to low country conditions of Sri Lanka. In: Induced plant mutations in the genomics era. Proceedings of an international joint FAO/IAEA symposium. International atomic energy agency, Vienna, Austria, p. 388–390
Pathirana R (2011) Plant mutation breeding in agriculture. In: Hemming D, editor. Plant sciences reviews. Cambridge: CABI 2012:107–125
Perry DA (1978) Report of the vigor test committee (1974–1977). Seed Sci Technol 6:159–181
Pishenin I, Gorbatova I, Kazakova E, Podobed M, Mitsenyk A, Shesterikova E, Dontsova A, Dontsov D, Volkova P (2021) Free amino acids and methylglyoxal as players in the radiation hormesis effect after low-dose γ-irradiation of barley seeds. Agriculture 11(10):918
Promnart U, Klakhaeng K, Doungsoongnern P, Kaewchuenchai R, Sarsu F (2018) Thai rice breeding for flood tolerance through electron beam induced mutation. Intl J Gen Engg 5(1):1–10
Ramchander S, Ushakumari R, Pillai MA (2015) Lethal dose fixation and sensitivity of rice varieties to gamma radiation. Ind J Agril Res 49(1):24–31
Rani MH, Kamruzzaman M, Ghanim AMA, Azad MAK, Akter MB (2016) Comparative effect of gamma and X-ray irradiations on some characters of rice seedlings of Ashfal and Binadhan-14. Biosci Agric Res 8(2):739–745
Rutger JN (1983) Applications of induced and spontaneous mutation in rice breeding and genetics. Adv Agron 36:383–413
Sao R, Sahu PK, Sharma D, Vishwakarma G, Nair JP, Petwal VC, Das BK (2020) Comparative study of radio-sensitivity and relative biological effectiveness of gamma rays, X-rays, electron beam and proton beam in short grain aromatic rice. Ind J Gen Plant Breed 80(04):384
Sareen S, Koul AK (1999) Mutation breeding in improvement of Plantago ovata Forsk. Indian J Genet 59:334–337
Shu QX, Li WH, Xia YW, Wu DX, Rui JY (1996) Study on the M1 generation of the biological effects of electron beam implanted seeds of rice. J Nucl Agric Sci 17:162–164
Singh RK, Chaudhary BD (1985) Biometrical methods in quantitative genetic analysis. Kalyani Publishers, New Delhi
Srivastava P, Marker S, Pandey P, Tiwari DK (2011) Mutagenic effects of sodium azide on the growth and yield characteristics in wheat (Triticum aestivum L. em. Thell). Asian J Plant Sci 10(3):190–201
Stadler LJ (1928) Genetic effects of X-rays in maize. Proc. Natl Acad Sci USA 14:69–75
Ussuf KK, Nair PM (1974) Effect of gamma irradiation on the indole acetic acid synthesizing system and its significance in sprout inhibition of potatoes. Rad Bot 14(4):251–256
Wang C, Shen M, Chen Q, Xu G (1995) Preliminary study of mutagenic effects of nitrogen ion implantation in rice. Acta Agri Nuc Sin 9(1):13–19
Willers H, Allen A, Grosshans D, McMahon SJ, Neubeck C, Wiese C, Vikram B (2018) Toward a variable RBE for proton beam therapy. Radiother Oncol 128:68–75
Zaka R, Chenal C, Misset MT (2004) Effect of low doses of short-term gamma radiation on growth and development through two generations of Pisum sativum. Sci Total Env 320:121–129
Zhu H, Xu J, Li S, Sun X, Yao S, Wang S (2008) Effects of high-energy-pulse-electron beam radiation on biomacromolecules. Sci China B Chem 51(1):86–91
Acknowledgements
The authors gratefully acknowledge Dr. V. C. Petwal, Scientific Officer from Raja Ramanna Centre for Advanced Technology, Indore, India, for providing irradiation facilities.
Funding
This work was partially supported by the Assam Agricultural University.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Janusz Zimny.
Rights and permissions
About this article

Cite this article
Bordoloi, D., Sarma, D. & Das, B.K. Comparative sensitivity and relative biological effectiveness of gamma-rays, X-rays and electron beams in aromatic Joha rice derived from different locations in Assam state. CEREAL RESEARCH COMMUNICATIONS 52, 57–71 (2024). https://doi.org/10.1007/s42976-023-00373-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42976-023-00373-7