
Abstract
This study explored the isolation and screening of an osmotolerant yeast, Wickerhamomyces anomalus BKK11-4, which is proficient in utilizing renewable feedstocks for sugar alcohol production. In batch fermentation with high initial glucose concentrations, W. anomalus BKK11-4 exhibited notable production of glycerol and arabitol. The results of the medium optimization experiments revealed that trace elements, such as H3BO3, CuSO4, FeCl3, MnSO4, KI, H4MoNa2O4, and ZnSO4, did not increase glucose consumption or sugar alcohol production but substantially increased cell biomass. Osmotic stress, which was manipulated by varying initial glucose concentrations, influenced metabolic outcomes. Elevated glucose levels promoted glycerol and arabitol production while decreasing citric acid production. Agitation rates significantly impacted the kinetics, enhancing glucose utilization and metabolite production rates, particularly for glycerol, arabitol, and citric acid. The operational pH dictated the distribution of the end metabolites, with glycerol production slightly reduced at pH 6, while arabitol production remained unaffected. Citric acid production was observed at pH 6 and 7, and acetic acid production was observed at pH 7. Metabolomic analysis using GC/MS identified 29 metabolites, emphasizing the abundance of sugar/sugar alcohols. Heatmaps were generated to depict the variations in metabolite levels under different osmotic stress conditions, highlighting the intricate metabolic dynamics occurring post-glucose uptake, affecting pathways such as the pentose phosphate pathway and glycerolipid metabolism. These insights contribute to the optimization of W. anomalus BKK11-4 as a whole-cell factory for desirable products, demonstrating its potential applicability in sustainable sugar alcohol production from renewable feedstocks.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sugar alcohols or polyols are a group of acyclic hydrogenated carbohydrates, but they are not classified as sugars [1]. Sugar alcohols have several applications in the food, pharmaceutical and chemical industries and are typically used as substitutes for table sugar in food and beverages because they are sweet but have fewer calories than table sugar. In addition, sugar alcohols are used in several pharmaceutical applications due to their excellent functional properties and health benefits. Some sugar alcohols, such as xylitol and sorbitol, have potential applications as building blocks for producing high-value chemicals [2]. Expansion of the global market for sugar alcohols is therefore expected. Sugar alcohols are present in small amounts in plants, fungi, and algae. The low sugar content hinders the extraction of sugar alcohols from natural sources [1]. Sugar alcohols are industrially produced by the catalytic hydrogenation of sugars under harsh conditions. The chemical processes are performed under high pressure and temperature, and many purification steps are required to recover the products, which results in a low product yield. Microbial fermentation is an environmentally benign process that utilizes renewable feedstocks [2]. The whole-cell catalytic process is therefore emerging as the predominant alternative for the production of sugar alcohols such as xylitol, erythritol, mannitol, sorbitol, and D-threitol [1, 2].
Glycerol or propane-1,2,3-triol is a simple sugar alcohol that is commonly known as glycerin. Glycerol has mild antimicrobial and antiviral properties; therefore, it is widely used in several consumer products, mainly as a supplement in personal care and pharmaceutical products. Glycerol can also be used to produce high-value chemical intermediates, including 1,3-propanediol, epichlorohydrin, acrolein, glycerol carbonate, and glycidol [3, 4]. Industrial chlorination of propane to allyl chloride generates approximately 25% of the glycerol needed to meet the global demand, while the remaining 75% is from the saponification of fats in the detergent industry [5, 6]. With the growth of the biodiesel industry, crude glycerol is produced as a major byproduct that contributes approximately 10% of the biodiesel production capacity [7, 8]. Crude glycerol usually contains impurities from the transesterification process, including methanol, alkali salts, and free fatty acids; therefore, purification steps are needed to eliminate those impurities. The Renewable Energy Directive (RED II) was announced by the European Union in 2018 to limit the consumption of plant-based biofuel [3]. This regulation has significantly affected biodiesel production and subsequently increased the global demand for glycerol.
Microbial fermentation using nonedible renewable feedstocks has therefore become a promising alternative. A previous study reported that Saccharomyces cerevisiae can convert monosaccharides to glycerol [9]. Several initiatives and studies have been conducted for improving fermentation process performance. Bisulfite or steering agents added in the fermentation media reportedly improve the glycerol yield by binding alcohol dehydrogenase (ADH) to inhibit the conversion of aldehydes to ethanol, increasing the amount of NADH available for the conversion of dihydroxyacetone phosphate (DHAP) to glycerol. An operating pH higher than 7 was also reported to improve glycerol production [10]. Genetic manipulation of the glycerol-producing pathway was conducted. The targeted enzymes, glyceraldehyde-3-phosphate dehydrogenases (GPD1 and GPD2), were overexpressed. By genetic manipulation, the byproducts from the overexpression mutants, including acetaldehyde, acetic acid, and acetoin, were produced and were likely eliminated [9]. In addition to S. cerevisiae, osmotolerant yeasts such as Candida, Debaryomyces, Hansenula, Pichia, Schizosaccharomyces, Saccharomyces, Torulaspora, and Zygosaccharomyces species can also produce sugar alcohols, including glycerol [11].
D-arabitol is another promising biobased chemical with versatility in industrial applications. D-arabitol has a low caloric content (0.2 kcal/g), moderate sweetness and a low glycemic index. Therefore, it can be used as a substitute for high-calorie sweeteners in the food and beverages consumed by diabetic patients. D-arabitol can also serve as a building block molecule to produce industrially important compounds and drug intermediates, including xylitol, HIV protease inhibitors, and immunosuppressive glycolipids. The US Department of Energy has listed D-arabitol as one of the 12 platform chemicals [12, 13]. Arabitol is currently produced by the catalytic hydrogenation of arabinonic acid and lactones at 100 °C in the presence of Raney nickel catalysts. The high operating temperature and the consumption of costly catalysts make the chemical production of arabitol environmentally and economically unsustainable [14]. For this reason, the biotechnological production of arabitol from renewable feedstocks has become an attractive option. Like glycerol, arabitol can be produced by osmotolerant yeasts, including Zygosaccharomyces, Candida, Debaryomyces, and Metschnikowia species.
Sugar alcohol production by osmotolerant yeasts requires operation under osmotic stress generated by increasing osmolarity in the surrounding environment, which results in hyperosmotic stress and loss of intracellular water. In response to the loss of intracellular water, osmotolerant yeasts accumulate sugar alcohols such as glycerol, erythritol, xylitol, arabitol, and mannitol in the cytoplasm. The operating conditions can be varied to manipulate the metabolic shift of sugar substrates toward the specific production routes for the desired end metabolites. Typically, fermentation by osmotolerant yeasts yields multiple end products. In recent years, the coproduction of chemical intermediates has attracted increasing interest because the yield from substrate conversion to more than one end metabolite is greater [15].
In this study, we investigated the coproduction of glycerol and D-arabitol by an osmotolerant yeast under various stress conditions. The isolation and screening of osmotolerant yeasts were conducted to determine the appropriate isolate, which was subsequently used in fermentation to produce both glycerol and D-arabitol. The use of a one-pot process for the coproduction of glycerol and D-arabitol was explored. This approach revealed a promising technique for generating high-value products via a sustainable bioprocess.
Materials and methods
Isolation and screening of osmotolerant yeasts
Fruit samples with high sugar content, including watermelon, ripe mango, pineapple, preserved sweet tamarind, dried sweetened fruits, and rotten fruits, were collected from local markets in Bangkok, Thailand. One gram of each sample was transferred into sterile enrichment medium (pH 3.5) that contained (per liter) 10 g of glucose (Dongxiao, China), 5 g of peptone (Sigma, Germany), and 3 g of yeast extract (Sigma, Germany). Antibiotics, including 250 mg/L penicillin G (Sigma, Germany) and 250 mg/L streptomycin (Sigma, Germany), were added to the enrichment medium to prevent bacterial growth. The culture was incubated at 30 °C and 150 rpm for 3 days. The culture broth was transferred and streaked onto a plate containing yeast extract-peptone-dextrose (YPD) agar medium, containing (per liter) 5 g of peptone, 3 g of yeast extract, 10 g of glucose, and 20 g of agar (Patanasin Enterprise, Thailand). The culture was incubated at 30 °C for 3 days. A single colony from the YPD agar plate was transferred onto a new YPD agar plate for further colony purification. The purified colony was selected for further screening of the osmotolerant yeast. The purified colony was transferred onto a new YPD agar slant for short-term storage. For long-term storage, a single colony was transferred to sterile YPD broth and incubated at 30 °C and 150 rpm for 24 h before being transferred to sterile 30% (v/v) glycerol (Sigma, Germany) in a 1.5 mL cryotube and kept at -80 °C.
Primary screening for osmotolerant yeasts
The osmotolerant yeast was screened by transferring a purified colony into 2 types of osmophilic yeast selective media. The purified colony was first transferred onto YEG50 medium containing (per liter) 5 g of yeast extract and 500 g of glucose. Notably, the YEG50 medium had a water activity of 0.91, and a previous study reported that osmotolerant yeasts could survive under a water activity as low as 0.89–0.95 and a high sugar content of 40–70% [16]. The growing colony was transferred to another selective medium (OM medium) that contained (per liter) 285 g of glucose, 15 g of fructose (Sigma, Germany), 5 g of yeast extract, 10 g of malt extract (Sigma, Germany), 5 g of meat peptone (Sigma, Germany), 100 mL of glycerol, 50 g of KCl (Merck, Germany), 1 g of MgSO4⋅7H2O (Riedel-de Haen, Germany), 2 g of (NH4)2HPO4 (Merck, Germany), 1 g of (NH4)2SO4 (Merck, Germany), and 1.3 mg of thiamine (Sigma, Germany). The OM medium also had a water activity of 0.91 [17]. The colonies that grew on both selective media were identified as osmotolerant isolates.
Secondary screening for polyol-producing isolates
The previously screened osmotolerant yeast isolates were further screened for their ability to produce polyols such as glycerol and arabitol. All isolates were transferred onto a new sterile YPD agar slant (pH 5.0) and incubated at 30 °C. After 24 h of incubation, 1 mL of sterile deionized (DI) water was added to the culture slant before vortexing thoroughly to obtain the cell suspension. The cell suspension was diluted approximately 50-100-fold with sterile DI water. The optical density at 600 nm (OD600) of the diluted cell suspension was spectrophotometrically measured. The actual OD600 of the cell suspension was calculated by multiplying the OD600 reading of the diluted cell suspension by the dilution factor. The inoculum was prepared by diluting the cell suspension with sterile DI water to an OD600 of 30–40. The inoculum (0.5 mL) was transferred into 49.5 mL of sterile synthetic medium (SM) containing (per liter) 250 g of glucose, 1.5 g of (NH4)2SO4, 1 g of KH2PO4 (Merck, Germany), 0.5 g of MgSO4⋅7H2O, and 0.2 g of CaCl2 (Merck, Germany). The culture was incubated at 30 °C and 200 rpm for 384 h. Fermentation samples were collected every 24 h for further analyses.
Morphological, physiological, and biochemical characterization
The morphology of the selected osmotolerant yeast isolates that produced sugar alcohols was investigated based on colony appearance. Each isolate was subcultured on a YPD agar plate and incubated at 30 °C for 3 days to allow full development of single colonies. Single colonies were observed under a microscope for morphological characterization. The modified methylene blue staining method was used for colony staining. Briefly, methylene blue was dissolved in sodium citrate solution (2% w/v) to a final concentration of 0.01% (w/v). The yeast suspension (0.5 mL of 107 cells/mL) was mixed with 0.5 mL of citrate methylene blue solution. The suspension was observed under a microscope after 5 min. Live cells remained unstained, whereas dead cells were stained blue [18]. The growth temperature (20–45 °C) and pH (1.5–12.0) ranges were determined for cultures grown in YPD broth. In addition, biochemical characterization was performed using the API 20 C AUX assay kit (bioMerieux, Marcyl’ Etoile, France) following an instruction manual (https://www.mediray.co.nz/media/15815/om_biomerieux_test-kits_package_insert-20210.pdf). Briefly, the API 20 C AUX strip consisted of 19 capsules that contained 19 different dehydrated carbon substrates and one capsule without substrate. The capsules were added to a semisolid minimal medium and a fresh yeast suspension with a turbidity of 2 McFarland before incubation at 29 °C ± 2 °C for 48–72 h (±6 h). The yeast only grew if it could utilize the specific substrate in each capsule as the sole carbon source. The observations were interpreted by comparison with the growth controls (a capsule with glucose as the positive control and the capsule without a carbon substrate as the negative control). Identification was performed by referring to the Analytical Profile Index. A suspension that was more turbid than the negative control indicated a positive reaction.
Molecular identification
The selected osmotolerant yeast isolates that could produce sugar alcohols were identified based on the D1/D2 domain of the 26S rRNA sequence. The isolate was transferred onto a YPD agar plate and incubated at 30 °C for 24 h. A single colony was dissolved in 0.1X SSC buffer. The cell suspension was mixed with a polymerase chain reaction (PCR) mixture containing the DNA primers NL-1 (5’GCATATCAATAAGCGGAGGAAAAG-3’) and NL-4 (5’GGTCCGTGTTTCAAGACGG-3’) to amplify the D1/D2 domain of the 26 S rRNA gene sequence [19]. The PCR products were purified with a QIAquick PCR Purification Kit (Qiagen, Tokyo, Japan) before gene sequencing analysis by Macrogen, Inc. (Korea). The DNA sequence was aligned by the BioEdit Sequence Alignment Editor version 7.2 (https://bioedit.software.informer.com/7.2/). The phylogenetic tree was constructed using the neighbor-joining method and the maximum-likelihood method in MEGA version 7 (https://www.megasoftware.net). The nucleotide sequences (26 S rRNA gene sequences) of the isolates were deposited in the NCBI GenBank (https://www.ncbi.nlm.nih.gov/genbank/) under the accession numbers OK458717.1 for isolate BKK12-1 and OK458718.1 for isolate BKK11-4.
Genome sequencing and variance analysis
The isolate BKK11-4 was grown in YPD medium and incubated at 30 °C for 24 h. Genomic DNA was extracted using a Wizard® Genomic DNA Purification Kit (Promega, A1120, USA) according to the manufacturer’s protocol. The DNA purity was investigated by gel electrophoresis. The concentration and purity of the DNA were measured from the ratio of the absorbance at 260 nm to that at 280 nm using a Nanodrop spectrophotometer (DS-11FX+, DeNovixÒ, USA). Illumina SBS technology was used to sequence the whole genome. A total of 1,599,864,328 nucleotides were sequenced, and the genome was created from the entire sequence. To reduce bias, the adapter sequences and poor-quality reads were removed using the Trimmomatic Program (v.0.38) (http://www.usadellab.org/cms/index.php?page=trimmomatic). The overall quality of the whole-genome sequence was examined using the Fast QC2 program (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/). The whole-genome sequence information can be accessed in the NCBI database (https://www.ncbi.nlm.nih.gov/genbank/) (accession number: JAPKVJ000000000). Paired-end assembly was performed using filtered reads and SPAdes version 3.15.4 (http://bioinf.spbau.ru/en/spades) with the cautious option. The scaffolds from the SPAdes program were polished using Pilon software (https://github.com/broadinstitute/pilon).
The genome sequence of the wild-type strain Wickerhamomyces anomalus Y-366T was used as a reference genome for genetic variation analysis of the isolate BKK11-4. Reads were aligned to the reference sequences using the Burrows‒Wheeler Aligner (BWA) program (v0.7.17) (https://github.com/lh3/bwa) to determine the genetic variants, including single-nucleotide polymorphisms (SNPs), insertion mutations, deletion mutations, base change mutations, transitions, and transversions, in all mapped samples.
Glycerol and arabitol fermentation by Wickerhamomyces anomalus BKK11-4
W. anomalus BKK11-4 was subcultured onto YPD agar plates (pH 5) and incubated at 30 °C for 24 h. A total of 2–3 colonies were propagated in 5 mL of preculture medium (Seed 1). Seed 1 was incubated at 30 °C and 120 rpm for 24 h. Then, 1.5 mL of Seed 1 was transferred into the preculture medium in a 250 mL baffled flask (Seed 2) at 3% inoculum volume and incubated at 30 °C and 120 rpm for 6 h. Seed 2 was transferred into the fermentation medium in a 250 mL flask at 10% inoculum volume. The culture was incubated at 30 °C and 200 rpm. Samples were collected every 24 h for a total of 240 h for analyses of cell biomass, glucose, and metabolites excreted during fermentation, including glycerol, arabitol, acetic acid, and ethanol. The precultures contained (per liter) 10 g of glucose (Dongxiao, China), 3 g of yeast extract (Sigma, Germany), 1.5 g of (NH4)2SO4 (Merck, Germany), 1 g of KH2PO4 (Merck, Germany), 0.5 g of MgSO4⋅7H2O (Riedel-de Haen, Germany), and 0.2 g of CaCl2 (Merck, Germany). The fermentation medium contained (per liter) 200 g of glucose (Dongxiao, China), 1.5 g of (NH4)2SO4 (Merck, Germany), 1 g of KH2PO4 (Merck, Germany), 0.5 g of MgSO4⋅7H2O (Riedel-de Haen, Germany), and 0.2 g of CaCl2 (Merck, Germany). In addition to the basal fermentation medium containing 200 g/L glucose, media with glucose concentrations varying from 50 g/L to 300 g/L were prepared to observe the metabolite profiles of W. anomalus BKK11-4. The trace element solution was added to the basal fermentation medium at 1 mL/L to observe metabolite production by W. anomalus BKK11-4. The trace element solution at 1000x contained (per liter) 500 µg of H3BO3 (Merck, Germany), 40 µg of CuSO4 (Fluka, France), 200 µg of FeCl3 (Fluka, France), 400 µg of MnSO4 (Riedel-de Haen, Germany), 100 µg of KI (Merck, Germany), 200 µg of H4MoNa2O4 (Fluka, France), and 400 µg of ZnSO4 (Merck, Germany).
Optimization of process conditions in a 7 L fermenter
W. anomalus BKK11-4 was subcultured on YPD agar plates (pH 5) and incubated at 30 °C for 24 h. A total of 2–3 colonies were propagated in 5 mL of preculture medium (Seed 1). Seed 1 was incubated at 30 °C and 120 rpm for 24 h. Then, 1.5 mL of Seed 1 was transferred into the preculture medium in a 250 mL baffled flask (Seed 2) at 3% inoculum volume and incubated at 30 °C and 120 rpm for 6 h. Sufficient Seed 2 was transferred into fermentation medium in a 7 L stirred tank fermenter (Sartorius Biostat® B) at 10% inoculum volume. The initial working volume was 3.5 L. Before inoculation with Seed 2, the bioreactor containing the fermentation medium was autoclaved at 121 °C and 15 psig for 30 min. After sterilization, the fermenter was cooled before starting the control system. Before inoculation, the operating temperature was set at 30 °C. The dissolved oxygen (DO) sensor (InPro6820, Mettler Toledo) was calibrated with nitrogen and air. After inoculation, the pH was automatically controlled at the set point using 5 M NaOH and 5 M HCl. The agitation rate and the pH were varied. Fermentation samples were collected every 24 h for 168 h for further analyses.
Sample analyses
The fermentation broth sample (1 mL) was centrifuged at 10,000 × g for 5 min. The supernatant was collected for analyses of the remaining glucose and metabolites using high-performance liquid chromatography (HPLC). The solid cell biomass was resuspended in 1 mL of deionized water and vortexed thoroughly before the OD600 was measured spectrophotometrically. To convert the OD600 to the dry cell biomass concentration (g/L), the correlation between the cell biomass concentration and the OD reading was plotted, where an OD600 reading of 3 was equivalent to 1 g/L dry cell weight.
The remaining glucose and metabolites formed during fermentation were analyzed using HPLC (Shimadzu-LC-20 A, Japan). The cell-free supernatant (1 mL) was diluted with double-distilled water and filtered through a cellulose acetate membrane. The diluted sample (15 µL) was automatically injected (Shimadzu-SIL-20 A) into an organic acid analysis column (Bio-Rad, Aminex HPX-87 H ion exclusion organic acid column; 300 mm × 7.8 mm), maintained at 45 °C in a column oven (Shimadzu-CTO-20 A) and eluted with 5 mM H2SO4 at a flow rate of 0.6 mL/min (Shimadzu-LC-20AT). A refractive index detector (Shimadzu-RID-20 A) was used to detect the organic compounds. Standards containing 0–2 g/L each component (glucose, glycerol, arabitol, acetic acid, and ethanol) were injected as references to determine the sample concentration from the respective peak areas.
The concentration was used to determine yield and productivity. The product yield (Yp/s) was calculated from the ratio of the amount of product formed to the amount of glucose consumed during fermentation. Volumetric productivity was defined as the total amount of product formed per unit volume per time.
Metabolomic analyses
Metabolomic analysis was employed to investigate the acquisition of extracellular metabolites. This involved transferring 200 µL of cell-free supernatant, along with an internal standard of 2 g/L xylose, into dried glass ampoules. After 3–4 h of drying with a Labconco CentriVap Aqueous Concentrator System (USA), the samples were incubated overnight in a Labconco® Laboratory Freeze-drying instrument (USA) at -50 °C. Each sample was chemically derivatized using methoxamine chloride dissolved in pyridine (20 mg/mL) (Merck, Germany) and incubated in a water bath at 50 °C for one hour, with vigorous mixing on a vortex at intervals. Subsequently, the samples were treated with 200 mL of N-methyl-N-(trimethylsilyl) trifluoroacetamide (MSTFA) + 1% TCMS (TCI, Japan), incubated for 30 min at 60 °C, and filtered through 0.22-µm PTFE hydrophobic filters before being subjected to gas chromatography‒mass spectrometry (GC‒MS) analysis [20]. GC‒MS/MS analysis was performed with an Agilent A 7890B system with a 7000 C GC/MS Triple Quad mass selective detector. Derivatized samples (0.2 µL) were injected into an HP-5ms capillary column with a 1:50 split ratio, utilizing helium as the helium carrier gas at a constant flow rate of 1 mL/min. The temperature program involved a gradual increase from 100 °C to 310 °C, with specific rate adjustments, and the electron impact ionization technique at -70 eV was employed for full scan detection in the mass range of m/z 33–800. For metabolite identification, Mass Hunter Quantitative Analysis Version B.6.00 software from Agilent Technologies was utilized, involving comparisons with the NIST2011 mass spectral library and validation using in-house authentic standards to ensure precise and reliable results.
Statistical analyses
The experimental data of this study were statistically analyzed using GraphPad Prism software (version 9.2.0) (https://www.graphpad.com/features). The results are presented as the mean±SD of triplicate experimental runs. For multiple comparisons between datasets, Tukey’s method for one-way analysis of variance (ANOVA) was used to determine significant differences.
Results and discussion
Isolation and characterization of osmotolerant yeast isolates
A total of 64 yeast isolates were screened from dried fruits, fermented fruits, rotten fruits, and high-sugar fruits. All 64 isolates were tested for their ability to survive and grow at high osmotic pressure. High sugar concentrations reduced water availability and therefore impeded regular yeast fermentation and growth. Only 7 yeast isolates could grow on both selective media (YEG50 and OM) that contained a high glucose concentration with low water activity. The 7 osmotolerant yeast isolates included TM2, BKK9-1, BKK10-2, BKK11-1, BKK11-4, BKK12-1, and BKK14-1.
The results of morphological, biochemical, and physiological characterization are given in Table 1. The colonies of the 7 isolates were circular, convex, and cream, white, or whitish cream colored. Bud formation was observed for all isolates. None of the 7 isolates showed true mycelia, and no pellicle formation was observed. All isolates showed good growth at temperatures up to 40 °C. As the temperature increased to 45 °C, the growth of the yeast isolates decreased. The inability of the yeast isolates to grow at 45 °C in this study is consistent with the mesophilic character of yeasts widely reported in the literature [21]. All isolates grew in the pH range from 1.5 to 10.5 except for TM2, which grew in a wider pH range (from 1.5 to 12.0). The effect of pH on biomass and glycerol production was observed in W. anomalus HH16. No significant difference in growth or glycerol production was observed in the culture in the pH range from 4 to 8 [22]. Debaryomyces hansenii, another osmotolerant yeast, grows in a narrow pH range from 3.5 to 6. A significant growth reduction was observed when this yeast was cultivated at a pH lower than 3.5 [23]. Typically, yeasts grow and multiply more rapidly at a pH value greater than 5.0, with the optimal growth rate observed at pH 5.5, whereas fermentation is more effective at a pH values less than 5.0. Therefore, maintaining a suitable pH that promotes growth and enhances fermentation performance is crucial. A comparison of all 7 isolates in this study with other osmotolerant yeasts reported in the literature revealed that the osmotolerant yeasts isolated in this study could survive in a wide pH range, especially at acidic pH values; therefore, low-pH conditions favor metabolite production. All the isolates could ferment and assimilate D-glucose, D-saccharose, and glycerol. L-arabinose, D-xylose, adonitol, xylitol, D-galactose, inositol, D-lactose, and D-raffinose were not fermented or assimilated by any of the isolates. Methyl-α-D-glucopyranoside and D-maltose were utilized by all the isolates except TM2. Only TM2 fermented and assimilated calcium 2-keto-gluconate. N-acetyl-glucosamine and D-cellobiose were fermented and assimilated by only BKK9-1. D-sorbitol could be fermented and assimilated by BKK9-1 and TM2. Only BKK11-4 could utilize D-trehalose, whereas BKK11-1 could utilize D-melezitose. Similar results were observed for W. anomalus HH16. This isolate was unable to ferment lactose and galactose, while it could assimilate other sugars [22]. The results of this study, together with those reported by Hawary et al. (2019), suggested that W. anomalus could assimilate a wide range of carbon substrates compared with Saccharomyces cerevisiae. For example, some strains of S. cerevisiae, such as MTCC170, only ferment D-glucose and D-galactose [21].
Selection of the polyol-producing isolates
After assessing morphological, physiological, and biochemical traits, all the yeast strains were tested to identify potential glycerol-arabitol-producing strains. Among the 64 isolates, 7 strains could produce glycerol and arabitol from glucose in SM medium at final concentrations ranging from 5.55 to 66.50 g/L glycerol and 4.50–36.43 g/L arabitol (Table 2). Among the 7 isolates, BKK11-4 and BKK12-1 exhibited good fermentation performance when grown in SM medium. A high glucose consumption rate was observed during fermentation by these 2 isolates (P < 0.05). This revealed that there was no substrate inhibition when a high initial glucose concentration was present in the fermentation medium. BKK11-4 produced glycerol at a final titer of 66.50 g/L with a yield and productivity of 0.27 g/g and 0.13 g/L⋅h, respectively. This isolate also exhibited high arabitol production, with a final titer of 36.43 g/L, yield of 0.11 g/g, and productivity of 0.09 g/L⋅h. BKK12-1 also exhibited good sugar alcohol production. This isolate produced glycerol at a final titer of 42.25 g/L with a corresponding yield and productivity of 0.19 g/g and 0.14 g/L⋅h, respectively. Arabitol was also produced by this isolate at a final titer of 32.93 g/L, with a corresponding yield and productivity of 0.16 g/g and 0.09 g/L⋅h, respectively.
The glucose uptake rate was the metabolic bottleneck under osmotic stress conditions. Glucose uptake at high concentrations without an inhibitory effect was desirable for fermentation by the osmotolerant yeasts. The presence of glycerol as an osmolyte in fermentation using a high glucose concentration regulated external osmotic pressure and maintained homeostasis [24]. The high consumption of glucose for the production of the sugar alcohols glycerol and arabitol indicated good fermentation performance. Compared with BKK12-1, BKK11-4 consumed more glucose, resulting in higher sugar alcohol production. Therefore, BKK11-4 was selected for further strain characterization and fermentation optimization for sugar alcohol production.
Phylogenetic tree and whole-genome sequence of the potential sugar alcohol-producing isolate
Molecular identification based on the D1/D2 domain of the 26 S rRNA gene sequence was conducted. A neighbor-joining phylogenetic tree showed that the isolates BKK12-1 and BKK11-4 were 99% genetically related to the type strain (W. anomalus NRRL Y-366T (AY046144)) (Fig. 1). The sequence of isolate BKK12-1 was 99% identical (565/567) to that of the type strain with no gap (0/567). Isolate BKK11-4 also shared 99% sequence identity with the type strain with a 1/538 gap (0.185%). The phylogenetic tree revealed that the BKK11-4 and BKK12-1 isolates are genetically related and seem to have descended from W. anomalus NRRL Y-366T as their common ancestor. W. anomalus was first identified in 1891 as Saccharomyces anomalus, a fermentative brewer’s yeast strain with an ovoid to sausage-like shape. S. anomalus formed a pellicle during cultivation, emitted an ester-like odor, and generated hat-shaped ascospores. In 1904, S. anomalus was assigned to the newly established genus Hansenula based on specimens with hat-shaped ascospores. Due to the lack of a genuine type species, Hansenula anomala NRRL Y-366 was chosen as the neotype for the genus Hansenula in 1951 for preservation. H. anomala was later reassigned to the genus Pichia based on the clear morphological difference between the two genera, i.e., Hansenula utilized nitrate as the sole nitrogen substrate, while Pichia was unable to use nitrate. Recently, a multigene sequence analysis (D1/D2) of LSU rRNA, SSUrRNA, and EF-1 split the genus Pichia into multiple phylogenetically defined taxa, leading to the transfer of Pichia anomala to W. anomalus (with conservation of the species name “anomalus”) [25, 26].

Phylogenetic tree based on the sequences of the D1/D2 domain of the 26 S rRNA gene showing the positions of W. anomalus BKK11-4, W. anomalus BKK12-1 and W. anomalus NRRL Y-366T with respect to closely related species
The genome shuffling technique was employed in the investigation of P. anomala TIB-x229. P. anomala is an osmotolerant sister strain of the genus Wickerhamomyces. An investigation showed that P. anomala, also known as W. anomalus, produces more sugar alcohols than its parent strain [27]. W. anomalus has recently been used as a second-generation bioethanol producer in both simultaneous saccharification and fermentation (SSF) and separated hydrolysis and fermentation (SHF) processes. The fermentation results demonstrated that the wastes were almost completely liquefied by enzymes, leading to a high ethanol yield of 141.06 g per 1 kg of waste, which was equivalent to 0.40 g of ethanol per g of carbohydrate consumed [28]. W. anomalus prevented blue mold disease caused by Penicillium expansum contamination in grape [29]. In weaned pigs, W. anomalus AR2016 was used to enhance the gut microbiota, improve intestinal barrier function, decrease the occurrence of diarrhea, and promote growth and health [26]. In addition, Fernandes et al. (2023) reported the production of biosurfactants with inhibitory and bactericidal activities in W. anomalus CCMA 0358 [30]. The European Food Safety Authority (EFSA) has granted W. anomalus the qualified presumption of safety (QPS) status and assigned it to the risk group in the biosafety level classification. This indicates that W. anomalus poses a low risk to people and the environment and is therefore suitable for use in the food industry [31]. A previous study reported that W. anomalus AN2-64 grew under high sugar concentrations. The accumulation of glycerol and arabitol was observed in fermentation using a high initial sugar concentration [32]. W. anomalus is also known as a deadly yeast due to the potential production and secretion of toxins that have antagonistic effects on infectious bacteria, fungi, protozoa, and viruses. It was also reported that W. anomalus could tolerate physicochemical and biotic stresses, i.e., osmotic stress, oxidative stress, nutritional starvation, anaerobiosis, pH shock, chemical inhibitors, cold/heat shock, mechanical shear, and hydrostatic pressure [31]. Due to several essential characteristics of W. anomalus, this yeast has become of interest for biotechnological applications [33, 34].
To elucidate the differentiation of the 2 isolates from their type strains, whole-genome sequencing was conducted using paired-end Illumina technology. An overview of the genome assembly statistics is shown in Table 3. In total, 2363 scaffolds with approximately 1000 base pairs each were created from 20 million base reads. The scaffolds had a predicted GC content of 34.57% and a combined length of 20,627,646 base pairs. These values were comparable to those of other W. anomalus strains reported in the literature, including W. anomalus NRRL Y-366 (35% GC content), W. anomalus DSM 6766 (33.1% GC content), and W. anomalus Y-1 (34.56% GC content) [35, 36]. Analysis of the genetic variance of W. anomalus BKK11-4 was performed by mapping with the genome reference strain W. anomalus Y-366T. The mapped sites of 13,491,688 base pairs were compared with the reference genome (14,145,566 base pairs). The mapped reads contained 9,074,433 base pairs, while the total reads contained 9,675,282 base pairs. Using variant calling, two types of variations, i.e., SNPs and InDels, were identified. There were 479,511 SNPs in the genome sequence that accounted for 328,475 transition and 151,010 transversion base changes. The InDel mutations comprised 10,351 insertions and 8,414 deletions. Whole-genome analysis revealed genetic divergence between W. anomalus BKK11-4 and W. anomalus NRRL Y-366T. The results were consistent with those indicated in the phylogenetic analysis.
Medium optimization for sugar alcohol production by W. anomalus BKK11-4
Figure 2 depicts the effects of trace elements on the fermentation kinetics of W. anomalus BKK11-4. With the trace elements in the fermentation medium, cell biomass production almost doubled, while metabolite production and glucose consumption seemed to be greater in the fermentation medium without trace elements (Fig. 2). At 240 h, glycerol was obtained at 21.41 g/L with a yield of 0.17 g/g, and arabitol was obtained at 23.63 g/L with a yield of 0.10 g/g. On the other hand, the cultivation of W. anomalus BKK11-4 in fermentation medium without trace elements yielded a final glycerol concentration of 50.63 g/L with a yield of 0.31 g/g and a final arabitol concentration of 37.38 g/L with a yield of 0.23 g/g. The results suggested that trace elements were not necessary for the coproduction of glycerol and arabitol, but they enhanced cell biomass production. Previous research reported that trace amounts of mineral salts, including MgSO4, FeSO4, ZnSO4, MnSO4, Na2HPO4, and KH2PO4, promoted the production of citric acid and lipids to facilitate cell formation in Yarrowia lipolytica SK7 [37]. In addition, KH2PO4, FeCl3, and CoCl2 exhibited a significant impact on cell proliferation in Saccharomyces sp [38].. The results of this study are consistent with the findings that fermentation medium without trace elements was essential for improving the sugar alcohol production performance, while cell biomass production was enhanced during cultivation using fermentation medium supplemented with trace elements [37, 38].

Effects of trace elements in the fermentation medium on the fermentation performance of W. anomalus BKK11-4; (A). glucose consumption, (B). cell biomass production, (C). glycerol production, and (D). arabitol production. Shake flask fermentation was conducted at 30 °C and 200 rpm for 240 h. The data are expressed as the mean±SD of triplicate runs
Figure 3 shows the kinetics of glucose uptake for cell biomass and metabolite production by W. anomalus BKK11-4 cultivated in fermentation medium containing different initial glucose concentrations. At a low initial concentration (50 g/L), glucose was mainly consumed within 72 h for cell biomass, arabitol, and citric acid production (Fig. 3A and B, and 3D). Increasing the initial glucose concentration directed glucose consumption toward glycerol and arabitol production, while citric acid production decreased (Table 4). Glycerol production was observed during fermentation with initial glucose concentrations of 150 g/L and above, while arabitol was detected during fermentation with initial glucose concentrations ranging from 50 g/L to 300 g/L (Fig. 3C and D). The results suggested that a high concentration of glucose was necessary for driving glucose flux toward glycerol and arabitol (Table 4). This phenomenon could be explained by osmoregulation occurring as a homeostatic process in yeast under osmotic stress, such as excessive salt and sugar concentrations. S. cerevisiae recognizes osmotic fluctuations through a plasma membrane-localized histidine kinase (Sln1). Sln1 was active under ambient conditions and inhibited stress signaling. When turgor pressure was decreased, Sln1 was inactivated, whereas the mitogen-activated protein (MAP) kinase cascade was activated, and the MAP kinase high osmolarity glycerol 1 (Hog1) was phosphorylated. The active Hog1 that accumulated in the nucleus regulated 2 HOG-targeted genes encoding glycerol-producing enzymes (GPD1 and GPP2). In summary, Hog1 activation induced the synthesis of glycerol as an osmolyte that increased the intracellular osmotic pressure [39]. As shown in Table 4, the activation of Hog1 in W. anomalus BKK11-4 was confirmed when the initial glucose concentration in the fermentation medium reached 150 g/L, as observed from glycerol production. Further increasing the initial glucose concentration resulted in increased glycerol production. W. anomalus BKK11-4 produced arabitol at all the initial glucose concentrations tested in this study. This is typical for W. anomalus, which produces glycerol and arabitol as the major polyols during fermentation using high sugar concentrations [32]. Complete glucose consumption was observed during fermentation with an initial glucose concentration of 50–200 g/L. Further increasing the initial glucose concentration led to incomplete glucose consumption. The remaining glucose at the end of fermentation affected downstream product recovery and purification as well as increased effluent treatment load [40]. Therefore, fermentation medium containing an initial glucose concentration of 200 g/L was selected for further process optimization in this study.

Fermentation kinetics of W. anomalus BKK11-4 cultivated in fermentation medium with different initial glucose concentrations; (A). glucose consumption, (B). cell biomass production, (C). glycerol production, and (D). arabitol production. Shake flask fermentation was conducted at 30 °C and 200 rpm for 240 h. The data are expressed as the mean±SD of triplicate runs
Optimization of conditions for enhancing the production of sugar alcohols in a stirred fermenter
Figure 4 depicts the fermentation kinetics of W. anomalus BKK11-4 cultivated in a laboratory-scale stirred fermenter. A significant effect of agitation on glucose consumption and metabolite production was observed when the aeration rate was kept constant at 1.0 vvm during fermentation. Increasing the agitation rate from 300 rpm to 500 rpm resulted in complete glucose consumption (Fig. 4A) and increased cell biomass production (Fig. 4B). The glucose consumption rate increased with increasing agitation speed. Cell biomass production increased with increasing agitation speed from 300 rpm to 500 rpm. Further increasing the agitation rate from 500 rpm to 700 rpm did not significantly affect cell biomass production (Table 5). Glycerol production was enhanced upon further increasing the agitation rate from 500 rpm to 700 rpm, while arabitol production increased when the agitation rate was increased from 300 rpm to 500 rpm (Fig. 4C and D). Citric acid production noticeably increased with increasing agitation rate (Table 5). It was reported that the increased agitation rate facilitated mixing and directed glucose consumption toward the tricarboxylic acid (TCA) cycle, which was coupled with the electron transport chain (ETC) when sufficient oxygen was available for ATP regeneration. The TCA cycle is also involved in biosynthesis. With a sufficient ratio of carbon to nitrogen (C: N) in substrates, high agitation reportedly promoted cell biomass production. On the other hand, a high C: N ratio can lead to citric acid accumulation during fermentation, as observed in this study [41]. Therefore, controlling the agitation rate at a certain level to provide sufficient mixing for a well-mixed fermentation broth and gas transfer is essential [40]. A previous study reported the positive effect of oxygen on cell growth and glycerol synthesis by the osmotolerant yeast Candida magnoliae I2B [42]. In addition, previous research reported an increase in arabitol production with increasing agitation rate in Zygosaccharomyces rouxii JM-C46 [43]. Mixing is typically employed to distribute gases, specifically oxygen, during aerobic fermentation. Under aerobic conditions, oxygen is involved in the bioconversion of glucose to cell biomass and metabolites as well as in the cellular maintenance process. ATP is regenerated by the oxidative phosphorylation of glucose via glycolysis, the TCA cycle, and the ETC. At the inner membrane of the mitochondria, the ETC system, which includes complex proteins such as the NADH dehydrogenase complex, cytochrome b-c1 complex, and cytochrome oxidase complex, oxidizes high-energy electrons from NADH and FADH2. With oxygen as a final electron acceptor in the ETC system, ADP is phosphorylated by ATP synthase to ATP to provide high proton energy for cellular metabolism. Vigorous mixing at 700 rpm provided an adequate supply of oxygen that drove ATP regeneration to provide sufficient energy for the complete consumption of glucose to produce end metabolites, including glycerol, arabitol, and citric acid (Table 5).

Effect of agitation on the production of sugar alcohols by W. anomalus BKK11-4 in a stirred fermenter; (A) glucose consumption, (B) cell biomass production, (C) glycerol production, and (D) arabitol production. Fermentation was conducted at 30 °C and pH 5 with 1.0 vvm air for 168 h. The data are expressed as the mean±SD of triplicate runs
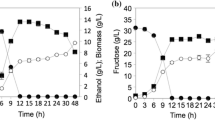
The operating pH also played a role in controlling the production of end metabolites by W. anomalus BKK11-4. Figure 5 shows the fermentation kinetics of W. anomalus BKK11-4 cultivated at different pH values. Complete glucose consumption was observed with the different profiles of the end metabolites. Cell biomass production was strongly promoted at pH 5, with a decreasing trend observed as the operating pH increased (Fig. 5B). Glycerol production was slightly lower at pH 6, while pH did not seem to affect arabitol production (Fig. 5C and D). Citric acid was detected at pH 6 and 7, and at pH 7, W. anomalus BKK11-4 produced acetic acid, while at lower pH, no acetic acid was observed during fermentation (Fig. 5E; Table 6). The effect of pH on glycerol production was found to vary by microbial species. A previous study reported that a slightly alkaline pH (pH 8) had a positive effect on glycerol production by W. anomalus HH16 [22]. In addition, previous research has shown no significant effect of pH on glycerol production by Candida glycerinogenes, but the authors concluded that acidic conditions were preferred for fermentation in their study [11]. The 2 osmotolerant yeast isolates C. magnoliae and C. glycerinogenes preferred pH 5 for cell growth and glycerol production [44]. The effect of pH on arabitol production was also reported. During fermentation by Z. rouxii JM-C46 at various initial pH values, the D-arabitol yield decreased significantly when the initial pH was greater than 5. Therefore, the optimal pH for this study was 5 [43]. It was reported that the arabitol yield from M. reukaufii AJ14787 was greater than 60% when fermentation was carried out at a low pH of approximately 3.5 to 5, and pH 5 was the optimal pH for arabitol production [45]. To promote the production of sugar alcohols and to limit the production of citric acid and acetic acid by W. anomalus BKK11-4 in this study, the optimal pH was 5. At this pH, maximal production of glycerol and arabitol was observed without the production of acetic acid and with low production of citric acid (Table 6). This would subsequently lower the load during product recovery and purification and effluent treatment [46].

Effect of pH on the production of sugar alcohols by W. anomalus BKK11-4 in a stirred fermenter; (A) glucose consumption, (B) cell biomass production, (C) glycerol production, (D) arabitol production, and (E) acetic acid. Fermentation was conducted at 30 °C and 700 rpm with 1.0 vvm air for 168 h. The data are expressed as the mean±SD of triplicate runs
Analysis of metabolite dynamics
Metabolomics plays a crucial role in monitoring bioprocesses and validating hypotheses by employing high-throughput techniques to measure extracellular metabolite levels. In this study, GC/MS analysis was utilized to detect primary and secondary metabolites, including amino acids, organic acids, sugar phosphates, alcohols, alkanes, and sugar/sugar alcohols, in samples categorized based on various osmotic stress levels determined by varying glucose concentrations. Heatmaps were generated to visualize differences in metabolite levels, revealing 29 identified metabolites, including L-alanine, acetic acid, propanoic acid, xylulose, DL-glyceraldehyde, butanal, butane, glycerol, D-arabinose, D-lyxose, D-arabitol, ribitol, L-arabitol, xylitol, β-D-galactofuranose, D-fructose, D-galactopyranose, D-glucose, D-allose, D-sorbitol, D-mannitol, maltose, D-mannose, talose, glycoside, myo-inositol, D-trehalose, D-cellobiose, and β-gentiobiose (Fig. 6). The sugar/sugar alcohol group exhibited the highest abundance of products produced by BKK11-4. Furthermore, this study explored the metabolic dynamics following glucose uptake and entry into glycolysis/gluconeogenesis, revealing redirections in pathways such as the pentose phosphate pathway, glycerolipid metabolism, fructose and mannose metabolism, inositol phosphate metabolism, and propanoate metabolism. The intricate network involving pentose and glucuronate interconversions highlighted key metabolites such as D-arabitol, D-xylulose, ribitol, D-arabinose, and D-mannitol. When exposed to high initial glucose concentrations, W. anomalus BKK11-4 exhibited a significant reversal of glucose catabolism, favoring the pentose phosphate pathway and glycerolipid metabolism. This finding aligns with the use of W. anomalus AN2-64 for ethanol production under high sugar concentrations. Glycerol and arabitol have emerged as the predominant sugar alcohols in the fermentation process [32]. In response to environmental stress, Candida albicans accumulates polyols such as D-arabitol and glycerol [47]. This study provides comprehensive insights into the versatile nature of glucose metabolism in W. anomalus BKK11-4, contributing to our understanding of cellular metabolism with potential applications in metabolomics and metabolic engineering for future studies.

Comparison of the metabolic profiles of W. anomalus BKK11-4 during fermentation with various initial glucose concentrations. Red indicates a relatively high quantity of metabolites, while blue indicates a comparatively low abundance of metabolites
Comparison of the sugar alcohol production performance of osmotolerant yeasts
Several osmotolerant yeast isolates with various final product titers have been reported as glycerol producers. These include isolates of Candida magnoliae, Candida glycerinogenes, Candida krusei, Z. rouxii, Kodamaea ohmeri, Pichia farinose, Pichia manchurica, and W. anomalus (Table 7). Some of these isolates could also produce arabitol during fermentation. C. magnoliae I2B produced glycerol with markedly high production performance. At the end of the batch cultivation, a final glycerol titer of 80.00 g/L with a 0.32 g/g yield and 0.65 g/L⋅h productivity was obtained without the production of arabitol. The highest glycerol production rate was observed during the late exponential phase of growth. Therefore, the glycerol production kinetics followed the mixed model, where glycerol could be considered a growth- or non-growth-associated product [18]. Another Candida sp. that exhibited significant glycerol production performance was C. glycerinogenes. A glycerol concentration within the range 110.00–130.00 g/L was obtained in batch fermentation under the optimized conditions from the shake flask scale to the 30 L stirred fermenter scale, with a corresponding yield of 0.58 g/g and productivity of 1.69 g/L⋅h. In a 50,000 L airlift fermenter with an average glucose concentration of 240.60 g/L, a final glycerol concentration of 121.90 g/L and a corresponding yield of 50.67% (w/w) were obtained. A low concentration of arabitol (4.50 g/L) was also detected. This study was the first to report fermentation performance that could be applied on an industrial scale [11].
Unlike Candida sp., Zygosaccharomyces rouxii, Kodamaea ohmeri, Pichia manchurica, Metschnikowia reukaufii, and W. anomalus typically produce arabitol as the major product. Z. rouxii NRRL27624 and Z. rouxii JM-C46 produced arabitol as the major product (80.00–83.40 g/L), while a low concentration of glycerol (10.00–15.10 g/L) with a low productivity of 0.07–0.08 g/L⋅h was obtained [43, 48]. K. ohmeri NH-9, a novel yeast isolate obtained from natural osmophilic sources, also produced arabitol as the primary metabolite from glucose (81.20 g/L final titer and 0.41 g/g yield), while glycerol was produced at a low rate (15.10 g/L final titer, 0.08 g/g yield, and 0.21 g/L⋅h productivity) [49]. P. manchurica is another osmotolerant yeast that was reported to produce both glycerol and arabitol, but its production performance was low, with final titers of 8.00 g/L glycerol and 27.60 g/L arabitol [50]. Among the arabitol producers with osmotolerance, the highest arabitol production was observed in the well-optimized batch fermentation by M. reukaufii AJ14787. A final arabitol concentration of 206 g/L with 17 g/L glycerol was obtained within 100 h [45].
W. anomalus exhibited remarkable characteristics compared to other osmotolerant yeasts previously reported. W. anomalus can utilize feedstocks for the production of a variety of essential intermediates, including ethanol, ethyl acetate, glycerol, and arabitol [34, 51]. This yeast predominates during the early stage of alcoholic fermentation, and it is widely recognized as a flavor enhancer for wine. This yeast is physiologically versatile and capable of growing on various carbon substrates and under various operating conditions [52]. In this study, the end metabolites and their final concentrations differed with changes in the operating conditions, i.e., initial glucose concentration, agitation rate, and pH. Arabitol was found to be the primary product of W. anomalus BKK11-4 in this study. The glycerol concentration continued to increase with increasing glucose concentration and agitation in response to high osmotic pressure. After UV mutagenesis, W. anomalus HH16 utilized a seawater-based medium for glycerol production without arabitol as a major byproduct. This isolated mutant produced glycerol at 73.33 g/L with a yield of 0.43 g/g and a productivity of 1.20 g/L⋅h [22]. With proper operating conditions and strain improvement, one can manipulate W. anomalus for the production of the target end metabolites glycerol and arabitol. The results of this study and those reported in a previous study indicated that W. anomalus has good characteristics for use as an industrial osmotolerant yeast, as it grew well and produced the desired product with acceptable process performance under the optimized process conditions in a simple medium [22].
Conclusions
The experimental data from this study indicate the potential of W. anomalus BKK11-4 as a producer of sugar alcohols such as glycerol and arabitol. This isolate was characterized using traditional taxonomic, biochemical, molecular, and genomic methods. W. anomalus was classified as an isolate in biosafety category risk group 1 and granted QPS status by the European Food Safety Authority; therefore, it is safe for use in industrial bioprocessing. Different medium formulations and processing conditions were employed during batch fermentation by W. anomalus BKK11-4 to demonstrate the osmotolerance characteristics and to preliminarily explore glucose metabolism to produce end metabolites from this isolate. Arabitol and glycerol were found to be the key end metabolites of W. anomalus BKK11-4. This isolate exhibited a significantly high production performance for both glycerol and arabitol based on certain optimized operating conditions. Therefore, with detailed optimization using systems biology and process engineering, it is believed that this isolate can be further modified for improved production performance for sugar alcohols and other promising metabolites that can be synthesized using the osmotolerant yeast platform.
Data availability
All the data generated or analyzed during this study are included in this article.
Abbreviations
- SM:
-
Synthetic medium
- OM:
-
Osmotolerant selective medium
- QPS:
-
Qualified presumption of safety
- SNPs:
-
Single-nucleotide polymorphisms
References
Abbasi AR, Liu J, Wang Z, Zhao A, Hanjie Y, Qu L, Alam AMd, Xiong W, Xu J, Lv Y (2021) Recent advances in producing sugar alcohols and functional sugars by engineering Yarrowia Lipolytica. Front Bioeng Biotechnol 9:648382. https://doi.org/10.3389/fbioe.2021.648382
Park Y-C, Oh EJ, Jo J-H, Jin Y-S, Seo J-H (2016) Recent advances in biological production of sugar alcohols. Curr Opin Biotechnol 37:105–113. https://doi.org/10.1016/j.copbio.2015.11.006
Attarbachi T, Kingsley MD, Spallina V (2023) New trends on crude glycerol purification: a review. Fuel 340:127485. https://doi.org/10.1016/j.fuel.2023.127485
Kong PS, Aroua MK, Daud WMAW (2016) Conversion of crude and pure glycerol into derivatives: a feasibility evaluation. Renew Sust Energ Rev 63:533–555. https://doi.org/10.1016/j.rser.2016.05.054
Ciriminna R, Pina CD, Rossi M, Pagliaro M (2014) Understanding the glycerol market. Eur J Lipid Sci Technol 116:1432–1439. https://doi.org/10.1002/ejlt.201400229
Lopes AP, Souza PR, Bonafé EG, Visentainer JV, Martins AF, Canesin EA (2019) Purified glycerol is produced from the frying oil transesterification by combining a pre-purification strategy performed with condensed tannin polymer derivative followed by ionic exchange. Fuel Process Technol 187:73–83. https://doi.org/10.1016/j.fuproc.2019.01.014
Nda-Umar UI, Ramli I, Taufiq-Yap YH, Muhamad EN (2019) An overview of recent research in the conversion of glycerol into biofuels, fuel additives and other bio-based chemicals. Catalysts 9:15. https://doi.org/10.3390/catal9010015
Quispe CAG, Coronado CJR, Carvalho JA (2013) Glycerol: production, consumption, prices, characterization and new trends in combustion. Renew Sust Energ Rev 27:475–493. https://doi.org/10.1016/j.rser.2013.06.017
Tilloy V, Cadière A, Ehsani M, Dequin S (2015) Reducing alcohol levels in wines through rational and evolutionary engineering of Saccharomyces cerevisiae. Int J Food Microbiol 213:49–58. https://doi.org/10.1016/j.ijfoodmicro.2015.06.027
Wang Z, Zhuge J, Fang H, Prior BA (2001) Glycerol production by microbial fermentation: a review. Biotechnol Adv 19:201–223. https://doi.org/10.1016/S0734-9750(01)00060-X
Zhuge J, Fang HY, Wang ZX, Chen DZ, Jin HR, Gu HL (2001) Glycerol production by a novel osmotolerant yeast Candida glycerinogenes. Appl Microbiol Biotechnol 55:686–692. https://doi.org/10.1007/s002530100596
Kordowska-Wiater M (2015) Production of arabitol by yeasts: current status and future prospects. J Appl Microbiol 119:303–314. https://doi.org/10.1111/jam.12807
Li X, Zhang Y, Zabed HM, Yun J, Zhang G, Zhao M, Ravikumar Y, Qi X (2023) High-level production of D-arabitol by Zygosaccharomyces rouxii from glucose: metabolic engineering and process optimization. Bioresour Technol 367:128251. https://doi.org/10.1016/j.biortech.2022.128251
Filippousi R, Tsouko E, Mordini K, Ladakis D, Koutinas AA, Aggelis G, Papanikolaou S (2022) Sustainable arabitol production by a newly isolated Debaryomyces prosopidis strain cultivated on biodiesel-derived glycerol. Carbon Resour Convs 5:92–99. https://doi.org/10.1016/j.crcon.2022.02.002
Shankar K, Kulkarni NS, Sajjanshetty R, Jayalakshmi SK, Sreeramulu K (2020) Co-production of xylitol and ethanol by the fermentation of the lignocellulosic hydrolysates of banana and water hyacinth leaves by individual yeast strains. Ind Crops Prod 155:112809. https://doi.org/10.1016/j.indcrop.2020.112809
Jermini MFG, Geiges O, Schmidt-Lorenz W (1987) Detection, isolation and identification of osmotolerant yeasts from high-sugar products. J Food Prot 50:468–472. https://doi.org/10.4315/0362-028X-50.6.468
Pfannebecker J, Schiffer-Hetz C, Frohlich J, Becker B (2016) Culture medium optimization for osmotolerant yeasts by use of a parallel fermenter system and rapid microbiological testing. J Microbiol Methods 130:14–22. https://doi.org/10.1016/j.mimet.2016.08.021
Smart KA, Chambers KM, Lambert I, Jenkins C, Smart CA (1999) Use of methylene violet staining procedures to determine yeast viability and vitality. J Am Soc Brew Chem 57:18023. https://doi.org/10.1094/ASBCJ-57-0018
Kurtzman CP, Robnett CJ (1998) Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Anton Leeuw Int J G 73:331–371. https://doi.org/10.1023/a:1001761008817
Kato H, Izumi Y, Hasunuma T, Matsuda F, Kondo A (2012) Widely targeted metabolic profiling analysis of yeast central metabolites. J Biosci Bioeng 113:665–673. https://doi.org/10.1016/j.jbiosc.2011.12.013
Ali MN, Khan MM (2014) Screening, identification and characterization of alcohol tolerant potential bioethanol producing yeasts. Curr Res Microbiol Biotechnol 2:316–324
Hawary H, Rasmey AHM, Aboseidah AA, El-Morsi ES, Hafez M (2019) Enhancement of glycerol production by UV-mutagenesis of the marine yeast Wickerhamomyces Anomalus HH16: kinetics and optimization of the fermentation process. 3 Biotech 9:1–14. https://doi.org/10.1007/s13205-019-1981-4
Marquina D, Barroso J, Santos A, Peinado JM (2001) Production and characteristics of Debaryomyces hansenii killer toxin. Microbiol Res 156:387–391. https://doi.org/10.1078/0944-5013-00117
Hagman A, Säll T, Piškur J (2014) Analysis of the yeast short-term Crabtree effect and its origin. FEBS J 281:4805–4814. https://doi.org/10.1111/febs.13019
Kurtzman CP (2011) Phylogeny of the ascomycetous yeasts and the renaming of Pichia anomala to Wickerhamomyces Anomalus. Anton Leeuw Int J G 99:13–23. https://doi.org/10.1007/s10482-010-9505-6
Ma Y, Sun Z, Zeng Y, Hu P, Sun W, Liu Y, Hu H, Rao Z, Tang Z (2021) Isolation, identification and function of Pichia anomala AR2016 and its effects on the growth and health of weaned pigs. Animals 11:1–22. https://doi.org/10.3390/ani11041179
Zhang G, Lin Y, Qi X, Wang L, He P, Wang Q, Ma Y (2015) Genome shuffling of the nonconventional yeast Pichia anomala for improved sugar alcohol production. Microb Cell Factories 14:1–10. https://doi.org/10.1186/s12934-015-0303-8
Ntaikou I, Antonopoulou G, Lyberatos G (2021) Sustainable second-generation bioethanol production from enzymatically hydrolyzed domestic food waste using Pichia anomala as biocatalyst. Sustainability 13:1–16. https://doi.org/10.3390/su13010259
Godana EA, Yang Q, Zhao L, Zhang X, Liu J, Zhang H (2021) Pichia anomala induced with Chitosan triggers defense response of table grapes against post-harvest blue mold disease. Front Microbiol 12:704519. https://doi.org/10.3389/fmicb.2021.704519
Fernandes NAT, Rose AL, Simões LA, Dias DR (2023) Chemical and biological evaluation of biosurfactant fractions from Wickerhamomyces Anomalus CCMA 0358. Appl Microbiol Biotechnol 107:7621–7633. https://doi.org/10.1007/s00253-023-12811-x
Walker GM (2011) Pichia anomala: cell physiology and biotechnology relative to other yeasts. Anton Leeuw Int J G 99:25–34. https://doi.org/10.1007/s10482-010-9491-8
Urano N, Ishida M, Naito Y, Endo R, Takei T, Takashio M, Okai M (2021) Ethanol fermentation by high-stress-tolerance aquatic yeasts and their mutants. Adv Microbiol 11:616–629. https://doi.org/10.4236/aim.2021.1111045
Giovati L, Ciociola T, De Simone T, Conti S, Magliani W (2021) Wickerhamomyces yeast killer toxins’ medical applications. Toxins 13:655. https://doi.org/10.3390/toxins13090655
Padilla B, Gil JV, Manzanares P (2018) Challenges of the non-conventional yeast Wickerhamomyces anomalus in winemaking. Fermentation 4:68. https://doi.org/10.3390/fermentation4030068
Schneider J, Rupp O, Trost E, Jaenicke S, Passoth V, Goesmann A, Tauch A, Brinkrolf K (2012) Genome sequence of Wickerhamomyces Anomalus DSM 6766 reveals genetic basis of biotechnologically important antimicrobial activities. FEMS Yeast Res 12:382–386. https://doi.org/10.1111/j.1567-1364.2012.00791.x
Shi X, Wang X, Hou X, Tian Q, Hui M (2022) Gene mining and flavour metabolism analyses of Wickerhamomyces anomalus Y-1 isolated from a Chinese liquor fermentation starter. Front Microbiol 13:891387. https://doi.org/10.3389/fmicb.2022.891387
Kumar LR, Yellapu SK, Tyagi RD, Drogui P (2021) Optimization of trace elements in purified glycerol for microbial lipid and citric acid production by Yarrowia Lipolytica SKY7. Syst Microbiol Biomanufacturing 1:76–89. https://doi.org/10.1007/s43393-020-00006-8
Venkateshwar M, Chaitanya K, Altaf M, Mahammad EJ, Bee H, Reddy G (2010) Influence of micronutrients on yeast growth and β-d-fructofuranosidase production. Indian J Microbiol 50:325–331. https://doi.org/10.1007/s12088-010-0005-1
Klipp E, Nordlander B, Krüger R, Gennemark P, Hohmann S (2005) Integrative model of the response of yeast to osmotic shock. Nat Biotechnol 23:975–982. https://doi.org/10.1038/nbt1114
Thitiprasert S, Kodama K, Tanasupawat S, Prasitchoke P, Rampai T, Prasirtsak B, Tolieng V, Piluk J, Assabumrungrat S, Thongchul N (2017) A homofermentative Bacillus sp. BC-001 and its performance as a potential L-lactate industrial strain. Bioprocess Biosyst Eng 40:1787–1799. https://doi.org/10.1007/s00449-017-1833-8
Prasirtsak B, Thitiprasert S, Tolieng V, Assabumrungrat S, Tanasupawat S, Thongchul N (2017) Characterization of D-lactic acid, spore-forming bacteria and terrilactibacillus laevilacticus SK5-6 as potential industrial strains. Ann Microbiol 67:763–778. https://doi.org/10.1007/s13213-017-1306-y
Sahoo DK, Agarwal GP (2002) Effect of oxygen transfer on glycerol biosynthesis by an osmophilic yeast Candida magnoliae I(2)B. Biotechnol Bioeng 78:545–555. https://doi.org/10.1002/bit.10237
Guo Q, Zabed H, Zhang H, Wang X, Yun J, Zhang G, Yang M, Sun W, Qi X (2019) Optimization of fermentation medium for a newly isolated yeast strain (Zygosaccharomyces rouxii JM-C46) and evaluation of factors affecting biosynthesis of D-arabitol. LWT 99:319–327. https://doi.org/10.1016/j.lwt.2018.09.086
Sivasankaran C, Ramanujam PK, Shanmugam S, Raja Sathendra E, Balasubramanian B, Mani J (2014) Comparative study on Candida sp. for the production of glycerol. Int J Chemtech Res 6:5058–5063
Nozaki H, Suzuki SI, Tsuyoshi N, Yokozeki K (2003) Production of D-arabitol by Metschnikowia reukaufii AJ14787. Biosci Biotechnol Biochem 67:1923–1929. https://doi.org/10.1271/bbb.67.1923
Phanthumchinda N, Thitiprasert S, Tanasupawat S, Assabumrungrat S, Thongchul N (2017) Process and cost modeling of lactic acid recovery from fermentation broths by membrane-based process. Process Biochem 68:205–213. https://doi.org/10.1016/j.procbio.2018.02.013
Kayingo G, Wong B (2005) The MAP kinase Hog1p differentially regulates stress-induced production and accumulation of glycerol and D-arabitol in Candida albicans. J Microbiol 151:2987–2999. https://doi.org/10.1099/mic.0.28040-0
Qi X, Luo Y, Wang X, Zhu J, Lin J, Zhang H, Chen F, Sun W (2015) Enhanced D-arabitol production by Zygosaccharomyces rouxii JM-C46: isolation of strains and process of repeated-batch fermentation. J Ind Microbiol Biotechnol 42:807–812. https://doi.org/10.1007/s10295-015-1603-z
Zhu HY, Xu H, Dai XY, Zhang Y, Ying HJ, Ouyang PK (2010) Production of D-arabitol by a newly isolated Kodamaea ohmeri. Bioprocess Biosyst Eng 33:565–571. https://doi.org/10.1007/s00449-009-0378-x
Sundaramoorthy BA, Gummadi SN (2019) Screening of new yeast Pichia manchurica for arabitol production. J Basic Microbiol 59:256–266. https://doi.org/10.1002/jobm.201800366
Raimondi S, Foca G, Ulrici A, Destro L, Leonardi A, Buzzi R, Candeliere F, Rossi M, Amaretti A (2022) Improved fed-batch processes with Wickerhamomyces anomalus WC 1501 for the production of d-arabitol from pure glycerol. Microb Cell Factories 21:179. https://doi.org/10.1186/s12934-022-01898-y
Atitallah BI, Ntaikou I, Antonopoulou G, Alexandropoulou M, Brysch-Herzberg M, Nasri M, Lyberatos G, Mechichi T (2020) Evaluation of the non-conventional yeast strain Wickerhamomyces anomalus (Pichia anomala) X19 for enhanced bioethanol production using date palm sap as renewable feedstock. Renew Energ 154:71–81. https://doi.org/10.1016/j.renene.2020.03.010
Liu HJ, Liu DH, Zhong JJ (2004) Oxygen limitation improves glycerol production by Candida krusei in a bioreactor. Process Biochem 39:1899–1902. https://doi.org/10.1016/j.procbio.2003.09.017
Chen G, Yao S (2013) The effect of created local hyperosmotic microenvironment in microcapsule for the growth and metabolism of osmotolerant yeast Candida krusei. Biomed Res Int 2013:467263. https://doi.org/10.1155/2013/467263
Saha BC, Sakakibara Y, Cotta MA (2007) Production of D-arabitol by a newly isolated Zygosaccharomyces rouxii. J Ind Microbiol Biotechnol 34:519–523. https://doi.org/10.1007/s10295-007-0211-y
Vijaikishore P, Karanth NG (1986) Factors affecting glycerol production by Pichia farinosa under alkaline conditions. Appl Biochem Biotechnol 13:189–205. https://doi.org/10.1007/BF02798457
Acknowledgements
This study was supported by NSTDA (RDI), the National Research Council of Thailand (Grant No. P-21-50505) and the Thailand Science Research and Innovation Fund, Chulalongkorn University (CU_FRB65_bcg(33)_209_61_01).
Funding
This study was supported by NSTDA (RDI), the National Research Council of Thailand, Research Chair Professor Grants provided by the National Science and Technology Development Agency (NSTDA), and the Thailand Research Fund (Grant No. RTA6280014). JT is the recipient of the Science Achievement Scholarship of Thailand (SAST) provided by the Thailand Research Fund.
Author information
Authors and Affiliations
Contributions
NT and ST developed the idea for the study. JT performed the experiments. JT, ST, and NT conducted the data analysis and prepared the manuscript. All the authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Rosane Freitas Schwan.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Thammaket, J., Srimongkol, P., Ekkaphan, P. et al. Isolation, screening, and characterization of the newly isolated osmotolerant yeast Wickerhamomyces anomalus BKK11-4 for the coproduction of glycerol and arabitol. Braz J Microbiol 55, 2149–2167 (2024). https://doi.org/10.1007/s42770-024-01383-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42770-024-01383-1