
Abstract
The biodiversity of filamentous fungi and their ecological relationships in the context of decaying Araucaria angustifolia (an endangered conifer) substrates are still mostly unknown. The present study aimed to investigate the diversity of saprotrophic filamentous fungi, based on morphological identification, associated with A. angustifolia, in addition to assessing possible saprobic/plant affinity relationship, and verifying whether the study areas and substrates affect the composition of the mycobiota. A total of 5000 substrates (decaying needles and twigs) were collected during five expeditions (2014/2015) to two areas: São Francisco de Paula National Forest (FLONA-SFP) and São Joaquim National Park (PARNA-SJ), Brazil. A total of 135 species distributed among 85 genera, 40 families, nine classes, 24 orders, three subphyla, and two phyla were identified. One new genus and five new species that were previously described, and six rare species and five species with affinity for A. angustifolia were also recorded. The twigs showed a community of fungi with greater richness and dominance. Conversely, the values of abundance, Simpson’s diversity index, and evenness were lower than those determined for needles. In terms of the study areas, FLONA-SFP showed higher values of richness, abundance, Simpson’s diversity index, and evenness than PARNA-SJ. Principal coordinate analysis and similarity percentage analysis showed the influence of both substrate factors and areas in the composition of the fungal communities. The presence of new, rare, and affinity-related species reinforces the study of fungi in the context of the conservation of this conifer, as these species are threatened by co-extinction.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The decomposition of plant debris is fundamental for the maintenance of forest ecosystems. The disintegration of complex organic compounds and the reabsorption of the resultant nutrients by plant roots maintain the nutrient cycle [1,2,3]. This process is influenced by several factors: (i) the physical and chemical characteristics of the organic matter—levels of lignin, cellulose, phenolic compounds, and the presence or absence of allelopathic substances; (ii) environmental conditions, such as humidity, precipitation, and temperature; and (iii) the community of decomposing organisms, including microorganisms and microfauna [3,4,5,6].
Saprotrophic fungi are highlighted among the decomposing microorganisms because they have an enzymatic arsenal capable of decomposing diverse plant cell wall polymers, including those of a recalcitrant nature such as lignocellulosic matrices [7]. In fact, they play an extremely important role in ecosystems, decomposing complex plant substrates and, consequently, promoting soil fertility [3, 8]. For instance, in some forest ecosystems, such as the rainforest, the fertility of the soil is maintained by the action of microorganisms in the decomposition of litter (including many fungi), allowing the existence of great botanical diversity [9]. The diversity of fungi associated with the litter is essential for an optimal decomposition process; a single or a consortium of species can be characteristic and important at certain stages of decomposition [10]. In this context, the decrease in the diversity of saprotrophic fungi can directly affect the decomposition process; the reduction or extinction of functional species in the community may consequently impact the entire ecosystem [11, 12]. Of note, a low decomposition rate induces, in turn, a deficit between the amount of substrate in the litter and the mineralization of nutrients, with negative effects on plant growth, vegetation composition, and ecosystem balance [13].
Several factors can influence the diversity and composition of the saprotrophic filamentous fungi community, including the plant species composing the ecosystem and the type of substrates and their spatial and temporal heterogeneity [14,15,16,17]. Only via the investigation of mycodiversity in the context of plant species can the effect of the above factors on the distribution of fungal species be understood [15, 17, 18]. Another important point to be considered is the different levels of affinity relationships between saprotrophic fungi and plants. Over time, several terms have been proposed to characterize fungus/host associations: preference for a host [19], saprobic/plant exclusivity, host specificity, host recurrence, restricted occurrence to host [20], and host affinity [14]. The knowledge of such relationships contributes to better global estimates of fungal species biodiversity [21].
Considering the saprotrophic mycodiversity associated with large groups of phanerogamic plants, studies have revealed a high diversity of species associated with angiosperms [10, 15, 18, 22] and gymnosperms [23,24,25,26,27,28]. Among gymnosperms, Araucaria angustifolia (Bertol.) Kuntze, (Coniferae, Araucariaceae), also known as Brazilian pine, is considered a critically endangered botanical species, according to the International Union for Conservation of Nature [29]. It is the only species of the genus native to Brazil and is mainly distributed in the southern region and some states in the southeast region (Minas Gerais, Rio de Janeiro, and São Paulo) [30, 31]. In addition to Brazil, A. angustifolia is also distributed in Argentina and Paraguay [32, 33]. The community of filamentous fungi associated with decomposition in the context of A. angustifolia is mostly unknown, although some studies have been conducted [34,35,36,37,38,39,40,41]. Most fungal studies in A. angustifolia focused on phytopathogenic fungi [42,43,44,45,46,47] and mycorrhizae fungi [48,49,50,51,52,53].
Given the above, the present study aimed to investigate the diversity and distribution of the saprotrophic filamentous fungi community on decaying needles and twigs from A. angustifolia. We further explored the influence of study areas and substrates in the composition of these communities, as well as possible saprobic/plant ecological relationships.
Material and methods
Study areas
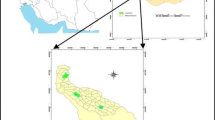
Two areas within the Araucaria Forest Conservation Units in Southern Brazil—the São Francisco de Paula National Forest (FLONA-SFP) (29° 23′ S and 50° 23′ W) and the São Joaquim National Park (PARNA-SJ) (28° 04′ S and 49° 39′ W)—were selected (Fig. 1). The areas have a temperate-type mesothermal climate: extremely humid, without a dry season, with well-distributed rainfall, and at least 1 month with average temperatures below 10 °C (occasional snowfall in winter common in Montanas) [54].

Study areas. South America, Brazil, State of Santa Catarina (SC) and Rio Grande do Sul (RS) in South region of Brazil and the two areas São Francisco de Paula National Forest (FLONA -SFP) and São Joaquim National Park (PARNA -SJ)
The FLONA-SFP, located at São Francisco de Paula Municipality, in the state of Rio Grande do Sul (RS), is a sustainable use Conservation Unit. It has an area of 1615.6 ha and altitudes that can exceed 900 masl [55]. Its vegetation cover is composed of 56% native vegetation, consisting of fragments of dense ombrophilous forest, mixed ombrophilous forest, and 39% of A. angustifolia, Pinus elliottii Engelm, P. taeda Blanco, and Eucalyptus spp. [56]. The altitudes recorded at the collection points were 838, 840, and 883 masl.
The PARNA-SJ, covering the municipalities of Urubici, Grão Pará, Bom Jardim da Serra, and Orleans in the state of Santa Catarina (SC), is an Integral Protection Conservation Unit. It has an area of 49,300 ha and 350–1800 masl, with the presence of four vegetation formations: dense ombrophilous forest, mixed ombrophilous forest, altitude fields, and nebular woods [57, 58]. The altitudes recorded at the collection points were 990 and 1031 masl.
Sampling, processing, and fungal identification
Five collection expeditions were carried out in each study area (Feb, May, Aug, and Nov 2014 and Feb 2015). In each area, 10 equidistant points approximately 200 m apart were selected. At each point, 25 decaying needles (3–7 cm long) and 25 decaying twigs (5 cm long) of A. angustifolia were collected and placed into properly identified paper bags. A total of 5000 samples were analyzed (1250 needles and 1250 twigs per study area). The paper bags were placed in a cardboard box and transported to the Laboratory of Mycology (LAMIC) at the Universidade Estadual de Feira de Santana. The samples were processed up to 5 days after collection as per the washing technique [59] and stored in moist chambers at room temperature (+ / − 25 °C). The substrates were observed daily from the second day of incubation for a period of 60 days [59]. Once reproductive structures of fungi were detected, they were transferred onto slides containing lactic acid and/or PVL resin (polyvinyl alcohol + lactophenol) [60] to make semi-permanent and permanent preparations, respectively. Identification of fungi was based on morphological characteristics of the reproductive structures, which were compared with those described in the specific literature [61]. After morphological identification, the slides were deposited at the Herbarium of Universidade Estadual de Feira de Santana (HUEFS). The species were grouped in higher ranks, such as phylum, class, order, and family, for a broader view of the fungi community present in A. angustifolia.
Ecological analysis
Fungi present in the communities associated with the substrates (needles and twigs) and study areas (FLONA-SFP and PARNA-SJ) were identified. Species abundance was evaluated by counting the number of individuals of each species per substrate [62]. To calculate the expected number of taxa in the communities, the Chao1 richness estimator was used [63]. The diversity between the two substrates and areas studied was compared using the Simpson’s diversity index (1-D), where D means the dominance of taxa that reaches values from 0 (all taxa are equally present) to 1 (one taxon dominates the community completely) and was determined as per the formula:
where ni = number of individuals of taxon i and n = total number of individuals [64].
The species distribution was assessed using the Pielou equitability index [65]. A comparison of the calculated indices between the different substrates and areas was performed with a 95% confidence interval via the bootstrap method [66]. To determine whether the communities were made up of abundant or rare species, a dominance–diversity curve was plotted based on the relative abundance of species in each substrate [67].
The constancy of the species in the different substrates was determined as per the formula:
where P = number of collections containing the species and N = total number of collections.
The species were grouped into the following constancy categories: constant, present in 50% or more collections; accessory, observed in >25% and < 50% of the collections; and accidental, detected in < 25% of the collections [68].
Dissimilarity in the species composition between communities was compared via the multivariate statistical method principal coordinate analysis (PCoA) [69], based on the Bray–Curtis dissimilarity matrix [70]. The one-way permutational multivariate analysis of variance (PERMANOVA) was used to assess whether the difference/dissimilarity between communities was significant [71]. The similarity percentage analysis (SIMPER) was used to determine the individual contribution of each species to the dissimilarities found between communities [72].
Analyses were performed using the PAST (version 4.03) [73] and Biodiversity Pro 2 [74] software.
Results
A total of 135 species of saprotrophic filamentous fungi distributed in 85 genera were identified in decaying needles and twigs of A. angustifolia in the two areas investigated (Table 1). Among the collected fungi, only two species (1.5%) belonged to the phylum Basidiomycota, Jaculispora submersa and Peyronelina glomerulata, and were classified into distinct families, orders, classes, and subphyla. The other 133 species (98.5%) belonged to the phylum Ascomycota; they were classified into 83 genera (97.6%) distributed in 38 families, 22 orders, seven classes, and one subphylum (Fig. 2). Among these, 22 genera (25.8%) and 38 species (28.1%) were classified as incertae sedis since they do not have a higher classification (Table S2).

Number of taxonomic categories per phylum of filamentous fungi isolated from A. angustifolia
The order with the largest number of families was Pleosporales (10), followed by Helotiales (3) and Hypocreales (3) (Fig. 3). Among families, Chaetosphaeriaceae showed the greatest richness (10), followed by Dictyosporiaceae (6) (Table S1). The genera with the largest number of species isolated were Chalara (7), Dactylaria (5), and Helicosporium (5) (Table S2).

Numbers of families (blue), genera (orange), and species (gray) classified by order of filamentous fungi associated with A. angustifolia
Considering the investigated substrates, the twigs showed the highest species richness (S = 104) and the lowest abundance (N = 1635); conversely, the needles showed the lowest richness (S = 92) and the highest abundance (N = 1980). The FLONA-SFP study area presented higher species richness (S = 116) and abundance (N = 1885) than PARNA-SJ (S = 78; N = 1730). Furthermore, the following results were observed with respect to the richness estimator Chao1: needles (99), twigs (162.5), FLONA-SFP, (140.6), and PARNA-SJ (93.3).
The fungi community associated with needles showed a lower species dominance (D = 0.04), greater evenness (J = 0.78), and, consequently, a higher Simpson diversity index value (C = 0.95) than that associated with twigs (D = 0.06; J = 0.72; C = 0.93). Moreover, FLONA-SFP contained lower species dominance (D = 0.04) and higher values of evenness (J = 0.77) and diversity index (C = 0.95), while PARNA-SJ showed higher dominance (D = 0.06) and lower evenness (J = 0.74) and diversity index (C = 0.93). These differences found in the values of the different indices calculated between the substrates and areas were significant based on the 95% confidence interval (Table 1).
The distribution of species per constancy category revealed the predominance of accidental species, followed by accessory and constant species for the needles and twigs (Fig. 4). Parasympodiella laxa and Helicosporium ramosum were the most constant species from needles and twigs, respectively, with both occurring in 100% of the collections. The dominance–diversity curve further supported these findings (Fig. 5). The fungal communities from both substrates presented few species with high abundance (dominant) and a predominance of species with one or two occurrences (rare) (Fig. 5). The most abundant species on needles were Anungitea fragilis (152), Chalara microspora (112), Helicosporium ramosum (118), Parasympodiella laxa (210), and Zygosporium gibbum (145). The most abundant species on twigs were Clonostachys rosea (130), Helicosporium guianense (266), H. ramosum (143), and P. laxa (194) (Table S1). Interestingly, 31 (22.9%) and 43 species (31.8%) were detected exclusively on needles and twigs, respectively.

Number of species isolated from needles (blue) and twigs (orange), considering the constancy categories of fungi isolated from A. angustifolia

Dominance–diversity curve of filamentous fungi isolated from needles and twigs of A. angustifolia
Sixty-one species were shared between the two substrates, resulting in an overlap of 45.2%. Among species shared by needles and twigs, A. fragilis (192), Ch. cylindrosperma (142), Ch. microspora (175), Cl. rosea (228), Dactylellina lysipaga (119), H. guianense (285), H. ramosum (261), P. laxa (404), Polyscytalum gracilisporum (127), Veronaea botryosa (102), and Z. gibbum (171) were the most frequent in this study. Conversely, among the rare species, 18 (13.3%) and 16 (11.8%) were detected only once and twice, respectively, on needles. On twigs, 35 (25.9%) and 10 species (7.4%) were detected only once and twice, respectively. In the study areas, 19 species (14.0%) were detected exclusively in PARNA-SJ and 57 species (42.2%) exclusively in FLONA-SFP, and 59 species (43.7%) were shared between the two study areas (Table S1).
On the biodiversity of filamentous fungi associated with A. angustifolia, a new genus Arthromoniliphora araucariae [40] and five new species, Chaetochalara mutabilis [36], Cryptocoryneum parvulum [37], Dictyosporium araucariae [39], and Trichoconis foliicola [38], were previously published; additionally, another new species of Endophragmiella was discovered (unpublished data) (Table 1). Among these new taxa, three occurred only on needles: C. parvulum was registered on 15 needles and present in 80% of collections; D. araucariae was found on 30 needles and present in 40% of collections; and T. foliicola was observed on 52 needles and present in 50% of collections.
Interestingly, PCoA revealed a tendency of separation of filamentous fungal communities from the two substrates and from the two areas studied (Fig. 6). The first principal coordinate represents the variability of filamentous fungi communities from needles and twigs (21.27%). while the second principal coordinate represents the variability of filamentous fungi communities detected in the different study areas (FLONA-SFP and PARNA-SJ; 18.43%). This ordering obtained by PCoA was confirmed by PERMANOVA, which revealed the differences in the fungi communities between the substrates and areas studied were statistically significant (F = 2.885; p = 0.0001). Moreover, the SIMPER analysis indicated a considerable dissimilarity between the fungal communities from the two areas (FLONA-SFP and PARNA-SJ), with an overall average of 54.52%. Three species, P. laxa, Z. gibbum, and A. fragilis, were the major contributors to this result (Table 2). With respect to the substrates, the SIMPER analysis revealed an average of 49.32% dissimilarity, with the species H. guianense, Z. gibbum, and A. fragilis contributing the most to this result (Table 2).

Ordering in two dimensions using PCoA for communities of filamentous fungi isolated from needles (N) and twigs (T) of A. angustifolia, in PARNA-SJ (P) (blue) and FLONA-SFP (F) (orange). Collection expeditions were identified with numbers 1 to 5, next to the samples
Discussion
Species composition
In this study, the decaying needles and twigs of A. angustifolia showed a wide variety of saprotrophic filamentous fungi. All of them represent fungi asexual forms, which is not unexpected since asexual reproduction is more common than sexual reproduction in the natural environment [75]. Of note, for most asexual fungi, no sexual counterparts were identified. Previously, the connection between sexual and asexual reproduction forms was only possible via the study of fungi developed as pure cultures under aseptic conditions in the laboratory. Only in the last decade has the connection between sexual and asexual reproduction been widely observed, following the popularization of complex molecular techniques. Using these new techniques combined with phylogenetic studies, a more accurate classification was possible, allowing the inclusion of asexual species (with unknown sexual morphological characteristics) in some of the higher hierarchical categories such as classes, orders, and families [75]. However, even considering these indisputable advances, hundreds to thousands of species have still not been subjected to molecular studies and, consequently, are not associated with any class, order, or family. These species are included in incertae sedis, representing a position not yet known within the formal higher classification.
The fungal community associated with A. angustifolia was mainly represented by the phylum Ascomycota (133 species), followed by Basidiomycota (two species). The low number of asexual Basidiomycota was expected, as there are fewer asexual fungi known within this phylum than those within Ascomycota [61, 76]. Among the 83 ascomycetes genera isolated in this study, 22 genera (26.5%) were considered incertae sedis, indicating that more effort is needed to better elucidate the higher classification of the group. The other 61 genera (73.4%) were classified into higher taxonomic categories and all belonged to the subphylum Pezizomycotina, the largest subphylum of Ascomycota [77].
Sordariomycetes and Dothideomycetes are the largest classes within the phylum Ascomycota. Because of their ability to degrade the lignocellulosic matrix of plant substrates, fungi from these classes are abundant in litter [78]. Not unexpectedly, our study, as well as other studies on saprobic microfungi [79,80,81], highlighted these classes as the most representative among wood, twigs, leaves, herbaceous plants, and soil decomposers.
In this study, the most representative order was Pleosporales, the largest within the Dothideomycetes class, and its representatives are found in the most diverse substrates and environments. The number of asexual fungi within this order is quite broad [82], which contributes to its large occurrence in this work. The next relevant order was Helotiales (Leotiomycetes class). Helotiales include many well-described species of readily cultured saprotrophic soil fungi, with many important species interacting with humans, animals such as bats (Geomyces ssp.) [83], and plants (Sclerotium ssp.) [84]. The Hypocreales order, also relevant within the fungi isolated here, belongs to Sordariomycetes class. This order is composed of ecologically diverse saprobic and pathogenic species found in a wide variety of substrates [85, 86].
The most representative families within Sordariales were Chaetosphaeriaceae and Dictyosporiaceae. Members of these families are found widely on decaying wood and plant debris and are important lignocellulose degraders [61, 87, 88]. Importantly, these families also stood out considering the number of saprobic asexual fungi species associated with palm trees in the Brazilian Amazon Forest [81]. The genera Chalara, Dactylaria, and Helicosporium, also quite relevant in our study (as the number of species identified), have a worldwide distribution [61] and are commonly found in other species of gymnosperms [27, 89, 90].
Diversity of species by substrate
The studied A. angustifolia substrates have a saprotrophic filamentous fungal community with a high species richness and diversity, as observed for other gymnosperms [24,25,26,27]. The presence of a few dominant species and a high proportion of rare species, as per the dominance–diversity curve (Fig. 5), is an expected pattern as shown by various ecological studies on diverse organism communities [91, 92], including microfungi [79, 93,94,95,96,97]. Among the fungi species, the following were the most abundant: A. fragilis (occurring in 233 needles and 41 twigs), Ch. microspora (147 needles and 63 twigs), Cl. Rosea (104 needles and 119 twigs), H. guianense (19 needles and 258 twigs), H. ramosum (144 needles and 126 twigs), and P. laxa (221 needles and 194 twigs); and they colonized up to 70% of the needles’ and twigs’ surface area. This observation suggests a fundamental role of these species in the decomposition of the substrates studied [79] and possibly a strategy of differentiated growing in relation to other species, especially those that develop more slowly. It is important to note, however, that rare species also play important ecological roles, as the decomposition process is carried out by several species, each probably having a specific role [12, 98].
Are substrates a determining factor of the composition and diversity of filamentous fungal communities?
In the present study, a significant difference was found between filamentous fungal communities isolated from the different substrates analyzed (Table 2). These differences may be associated with substrate species specificity [5, 12]. In fact, other studies reported this as an important factor affecting the constitution of fungal communities [5, 79, 81, 99]. Overall, the different tissues that make up plant parts (leaves, petioles, trunks, barks, roots, twigs, and branches) contribute to the recurrence of certain species of fungi and the consequent formation of different communities. In other words, different substrates support different fungal communities [99,100,101].
Saprobic/plant affinity ratio
Most species isolated in the present study have been associated with other plants, within both gymnosperms (46 species) [27, 46, 47] and angiosperms (118 species) [10, 46, 102, 103]. For instance, Cai et al. investigated bamboo-associated fungi and observed that most isolated species had already been reported in other plants, suggesting a low specificity [79]. This may be owing to the high ability of fungi to colonize different plants [104].
On the contrary, of the six new species found in A. angustifolia, only Chaetochalara mutabilis was registered in another plant species (Calophyllum brasiliense Cambess) [36]. The other five new species may represent a new saprobic/plant affinity relationship, as Paulus et al. suggested [14]. However, one cannot rule out the possibility that these species may be found in other plants or unexplored niches in the future.
Interestingly, among these new species, C. parvulum, D. araucariae, and T. foliicola occurred abundantly and recurrently only on needles. These data allow us to infer that these species have an affinity relationship with this part of the plant (needles), which may be related to its chemical composition and plant tissue. Another substrate specificity observed was the exclusive occurrence of C. parvulum on the stomata, suggesting the affinity of this fungal species to this group of leaf epidermis cells. Many phytopathogenic fungi use this characteristic route of infection (via the stomata) [105]. However, there is no evidence of this type of interaction for C. parvulum because living leaves were not analyzed.
Pseudaegerita conifera is another example of a fungal species with affinity to needles. It was previously recorded only on Pinus halepensis Mill needles [106], and now on A. angustifolia needles. This species was originally identified to be associated with submerged needles [106]. Pseudaegerita is characterized as aero-aquatic because of the presence of clathrate conidia [107]. All species of this genus are commonly found in aquatic environments or near riverbanks and forest streams [106]. However, in the present study, Ps. conifera was isolated from a terrestrial environment. This may be related to the possible proximity of the collection sites with streams or with the high humidity of the studied areas during the collection expeditions.
The affinity relationship between saprotrophic fungi and specific parts of plants may be associated with the nutritional conditions provided. With respect to needles, they have xeromorphic characteristics, with lignified hypodermis, stomata containing guard cells with extremely lignified periclinal walls, pectin secretion in the spongy parenchyma, and secretory channels of phenolic compounds between vascular bundles [108]. An important factor essential for the establishment of such relationships is the presence of an enzymatic framework in fungal species capable of decomposing such tissue and leaf structures [8].
Among the species associated with A. angustifolia twigs, the fungus Sterigmatobotrys macrocarpus stood out in the present study. In the literature, this species has always been associated with coniferous wood (Abies Mill., Agathis Salisb., Dacrydium Torell, Picea A. Dietr., Podocarpus Pers., and Taxus L.) both in terrestrial and aquatic environments [46, 109] and can, thus, be considered a fungus with an affinity for coniferous wood. This is probably owing to the common characteristics of coniferous wood (which differs from that of angiosperms): presence of long fibers, predominance of a single cell type—longitudinal tracheids [110], higher concentration of lignin [111], and structurally different lignin [112].
Chalara stipitata has previously been recorded on the leaves of Agathis australis (D. Don) Loudon and Podocarpus hallii Kirk [113, 114]. In the present work, Ch. stipitata was registered on nine needles and eight twigs of A. angustifolia, developing well in both substrates. With this similar abundance in needles and twigs, we cannot infer that there is an affinity for a specific gymnosperm part, even if we consider the studies reporting the presence of this fungal species only in leaves of this group of vascular plants. However, we cannot exclude an affinity for gymnosperms, since, almost half a century after the discovery of this saprobic fungus, no reports in the context of angiosperms are available. This affinity may be related to the nutritional characteristics of particular plant structures already mentioned. This said, further taxonomic/ecological investigations on the mycobiota associated with different parts of gymnosperms must be carried out to better understand these complex relationships.
Diversity and composition of filamentous fungi species in the study areas
The analysis of the composition of filamentous fungi species comparing the two studied areas showed that the “area factor” is also important for the determining of fungal communities. A high dissimilarity between the areas was demonstrated via SIMPER analysis.
Although the Araucaria forest areas studied have the same climate and similar altitudes, they also present important environmental differences: FLONA-SFP is an area that is composed mostly of dense vegetation of native Araucaria forest, while PARNA-SJ is composed of Araucaria forests discontinued by grass fields. This environmental heterogeneity can influence the composition of the fungal communities [115] and contribute to the differences in diversity and composition of microfungi observed in this study.
The PCoA analysis corroborated the abovementioned results. Remarkably, this observation was also reported in the context of composition studies of palm-associated fungal communities. Distinct collection sites significantly interfered with the fungal occurrence, with more similar compositions reported in palm trees from the same versus distinct sites [116, 117]. Additionally, differences in fungal communities in bamboos from two different sites were also observed [118]. Considering endophytic fungi and their characteristic fungus/plant interactions, site-specific/environmental factors were also shown to exert a great influence on the fungal communities. In a study on fungi associated with Pinus radiata D. Don, the fungi communities varied from one site to another [119].
Conclusions
The study of the mycobiota associated with A. angustifolia decomposing needles and twigs revealed that this plant species is a reservoir of great filamentous fungi biodiversity, with different communities associated with its different substrates. A high species richness was observed, including rare and new species. Saprobic/plant affinity ecological relationships were also noted. These factors should be considered for the conservation of this conifer and the consequent conservation of its mycobiota, which plays a fundamental role in the process of decomposition and the consequent maintenance of ecosystems.
The fungi associated with this critically endangered conifer are also at risk of co-extinction in future. Understanding whether the fungi considered closely associated with A. angustifolia can colonize other conifers (or any other plant) is, therefore, extremely important. Further studies are required to ensure the preservation of these fungal species in nature. In contrast, the axenic culture of these fungi is a real possibility, although the genetic variability of the population in this context will probably be reduced.
Our data showed that both substrates and study areas are determining factors for the composition of fungal communities. Thus, future research in other niches of this plant, such as reproductive structures and bark, as well as other areas of occurrence of A. angustifolia, could enrich the knowledge on the associated mycobiota. Such studies may even allow the identification of new fungal species.
There is still an absence of multidisciplinary studies, in particular ecological ones, on critically endangered plants (including Araucaria angustifolia). Our data clearly shows that these plants can contribute to the generation of new and extremely valuable data. However, numerous sites, substrates, and niches are still untapped and these should be considered in the context of global fungal biodiversity estimates.
Data availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Code availability
Not applicable.
References
Meguro M, Vinueza GN, Delitti WBC (1980) Ciclagem de nutrientes minerais na Mata Mesófila Secundária, São Paulo, III - Decomposição do material foliar e liberação dos nutrientes minerais. Boletim de Botânica 8:7–20. https://doi.org/10.11606/issn.2316-9052.v8i0p7-20
Fernandes AV, Backes A (1998) Produtividade primária em floresta com Araucaria angustifolia no Rio Grande do Sul. Iheringia, Série Botânica 5(11):63–78
Hättenschwiler S, Fromin N, Barantal S (2011) Functional diversity of terrestrial microbial decomposers and their substrates. C R Biologies 334:393–402. https://doi.org/10.1016/j.crvi.2011.03.001
Berg B, Berg MP, Box E, Bottner P, Breymeyer A, Calvo de Anta R, Couteaux MM, Gallardo A, Escudero A, Kratz W, Madeira M, Malkonen E, McClaugherty C, Meentemeyer M, Muñoz F, Piussi P, Remacle J, Virzo de Santo A (1993) Litter mass loss rates in pine forests of Europe and Eastern United States: some relationships with climate and litter quality. Biogeochemistry 20:127–159. https://doi.org/10.1007/BF00000785
Allegrucci N, Bucsinszkya AM, Arturib M, Cabello MN (2015) Communities of anamorphic fungi on green leaves and leaf litter of native forests of Scutia buxifolia and Celtis tala: Composition, diversity, seasonality and substrate specificity. Rev Iberoam Micol 32(2):71–78. https://doi.org/10.1016/j.riam.2013.11.002
Osono T (2020) Functional diversity of ligninolytic fungi associated with leaf litter decomposition. Ecol Res 35:30–43. https://doi.org/10.1111/1440-1703.12063
Sharma RK, Arora DS (2013) Fungal degradation of lignocellulosic residues: an aspect of improved nutritive quality. Crit Rev Microbiol 41(1):52–60. https://doi.org/10.3109/1040841X.2013.791247
Barlocher F, Kendrick B (1974) Dynamics of the fungal population on leaves in a stream. J Ecol 62:761–791. https://doi.org/10.2307/2258954
Jordan CF, Herrera R (1981) Tropical rain forests: are nutrients really critical? Am Nat 117(2):167–180
Shanthi S, Vittal BPR (2010) Fungi associated with decomposing leaf litter of cashew (Anacardium occidentale). Mycology 1(2):121–129. https://doi.org/10.1080/21501201003743154
Santana MS, Lodge DJ, Lebow P (2005) Relationship of host recurrence in fungi to rates of tropical leaf decomposition. Pedobiol 49:549–564. https://doi.org/10.1016/j.pedobi.2005.06.009
McGuire KL, Bent E, Borneman J, Majumder A, Allison SD, Treseder KK (2010) Functional diversity in resource use by fungi. Ecology 91:2324–2332. https://doi.org/10.1890/09-0654.1
Hättenschwiler S, Tiunov AV, Scheu S (2005) Biodiversity and litter decomposition in terrestrial ecosystems. Annu Rev Ecol Evol Syst 36:191–218. https://doi.org/10.1146/annurev.ecolsys.36.112904.151932
Paulus BC, Kanowski J, Gadek PA, Hyde KD (2006) Diversity and distribution of saprobic microfungi in leaf litter of an Australian tropical rainforest. Mycol Res 110:1441–1454. https://doi.org/10.1016/j.mycres.2006.09.002
Costa LA, Gusmão LFP (2017) Communities of saprobic fungi on leaf litter of Vismia guianensis in remnants of the Brazilian Atlantic Forest. J For Res 28(1):163–172. https://doi.org/10.1007/s11676-016-0268-4
Santa Izabel TS, Gusmão LFP (2018) Richness and diversity of conidial fungi associated with plant debris in three enclaves of Atlantic Forest in the Caatinga biome of Brazil. Plant Ecol Evol 151(1):35–47. https://doi.org/10.5091/plecevo.2018.1332
Kodsueb R, Lumyong S (2019) Diversity of saprobic fungi on Magnolia garrettii: do collecting sites and seasons affect the fungal community? Sains Malaysiana 48(11):2437–2449. https://doi.org/10.17576/jsm-2019-4811-14
Lacerda LT, Gusmão LFP, Rodrigues A (2019) Fungal communities in different aged leaves of Eucalyptus microcorys F. Muell Braz J Bot 42(3):499–508. https://doi.org/10.1007/s40415-019-00557-8
Hollins TW, Jellis GJ, Scott PR (1983) Infection of potato and wheat by isolates of Rhizoctonia solani and Rhizoctonia cerealis. Plant Pathol 32:303–310. https://doi.org/10.1111/j.1365-3059.1983.tb02838.x
Zhou D, Hyde KD (2001) Host-specificity, host-exclusivity, and host-recurrence in saprobic fungi. Mycol Res 105(12):1449–1457. https://doi.org/10.1017/S0953756201004713
Hyde KD (2001) Where are the missing fungi? Does Hong Kong have any answers? Mycol Res 105:1514–1518. https://doi.org/10.1017/S0953756201004889
Hyde KD, Bussaban B, Paulus B, Crous PW, Lee S, Mckenzie EHC, Photita W, Lumyong S (2007) Diversity of saprobic microfungi. Biodivers Conserv 16:7–35. https://doi.org/10.1007/s10531-006-9119-5
Stevenson JA (1975) Fungi of Puerto Rico and the American Virgin Islands. Contr Reed Herb, USA
Crous PW, Seifert KA, Castañeda Ruiz RF (1996) Microfungi associated with Podocarpus leaf litter in South Africa. S Afr J Bot 62(2):89–98. https://doi.org/10.1016/S0254-6299(15)30597-4
Minter DW, Holubová-Jechová V (1981) New or interesting Hyphomycetes on decaying pine litter from Czechoslovakia. Folia Geobot Phytotax Praha 16:195–217. https://doi.org/10.1007/BF02851863
Tokumasu S (1998) Fungal successions on pine needles fallen at different seasons: the succession of surface colonizers. Mycoscience 39(4):417–423. https://doi.org/10.1007/bf02460902
Tokumasu S, Aoiki T (2002) A new approach to studying microfungal succession on decaying pine needles in an oceanic subtropical region in Japan. In: K.D. Hyde and E.B.G. Jones (eds) Fungal succession. Fungal Diversity 10:167–183
Bensch K, Braun U, Groenewald JZ, Crous PW (2012) The genus Cladosporium. Stud Mycol 72:1–401. https://doi.org/10.3114/sim0003
Thomas P (2013) Araucaria angustifolia. The IUCN Red List of Threatened Species https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T32975A2829141.en Accessed 24 April 2020
Shimizu JY, Oliveira YMM (1981) Distribuição da variação e usos de recursos genéticos de araucária no Sul do Brasil. Embrapa-URPFCS, Curitiba
Souza VC (2015) Araucariaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB33971 Accessed 31 May 2020
Cozzo D (1980) Distribución fitogeográfica en la Argentina de Araucaria araucana y A. angustifolia. In: IUFRO meeting on forestry problems of the genus Araucaria. FUPEF, Curitiba, pp. 1–3
Lopez JA, Little Junior EL, Ritz GF, Rombold JS, Hahn WJ (1987) Arboles comunes del Paraguay: nande yvyra mata kuera. Cuerpo de Paz, Colección e Intercambio de Información, Washington
Sutton BC, Hodges CSJr (1977) Harknessia araucariae from Brazil. Mycologia 69:829–831. https://doi.org/10.2307/3758872
Kirk PM, Sutton BCA (1985) Reassessment of the anamorph genus Chaetopsina (hyphomycetes). Trans Br Mycol Soc 85(4):709–718. https://doi.org/10.1016/S0007-1536(85)80267-9
Silva SS, Silva CR, Gusmão LFP, Castañeda-Ruiz RF (2015) A new species of Chaetochalara on decaying leaves from Brazil. Mycotaxon 130:505–509. https://doi.org/10.5248/130.505
Silva SS, Gusmão LFP, Castañeda-Ruiz RF (2015) Cryptocoryneum parvulum, a new species on Araucaria angustifolia (Brazilian pine). Mycotaxon 130:465–469. https://doi.org/10.5248/130.465
Silva SS, Gusmão LFP, Castañeda-Ruiz RF (2015) Conidial fungi on Araucaria angustifolia: Trichoconis foliicola sp. nov. and two new records from Brazil. Mycotaxon 130:1051–1059. https://doi.org/10.5248/130.1051
Silva SS, Castañeda-Ruiz RF, Gusmão LFP (2015) New species and records of Dictyosporium on Araucaria angustifolia (Brazilian pine). Nova Hedwigia 102:523–530. https://doi.org/10.1127/nova_hedwigia/2015/0325
Silva SS, Gusmão LFP, Castañeda-Ruiz RF (2016) Arthromoniliphora araucariae gen. & sp. nov. from Brazilian pine. Mycotaxon 131:821–826. https://doi.org/10.5248/131.821
Silva SS, Gusmão LFP (2017) New records of rare dematiaceous conidial fungi on Araucaria angustifolia from Brazil. Nova Hedwigia 104(4):529–538. https://doi.org/10.1127/nova_hedwigia/2016/0390
Hodges CS, May LC (1972) A root disease of pine, Araucaria and Eucalyptus in Brazil caused by a new species of Cylindrocladium. Phytopathology 62:898–901. https://doi.org/10.1094/Phyto-62-898
Butin H, Peredo HL (1986) Hongos parasitos em coniferas de America Del Sur, con especial referencia a Chile. Biblioth Mycol 101:1–100
Auer CG, Grigoletti A Jr (1997) Doenças registradas em Araucaria angustifolia e Pinus spp. nos estados do Paraná e de Santa Catarina. Colombo EMBRAPA-CNPF 31:1–3
Crous PW (2002) Taxonomy and pathology of Cylindrocladium (Calonectria) and allied genera. American Phytopathological Society, St. Paul
Farr DF, Rossman AY (2020) Fungal Databases, Systematic Mycology and Microbiology Laboratory, ARS, USDA. http://nt.ars-grin.gov/fungaldatabases/ Accessed 7 July 2020
Mendes MAS, Urben AF (2020) Fungos relatados em plantas no Brasil, Laboratório de Quarentena Vegetal. Brasília, DF: Embrapa Recursos Genéticos e Biotecnologia. http://pragawall.cenargen.embrapa.br/aiqweb/michtml/fgbanco01.asp Accessed 25 June 2020
Moreira M, Trufem SFB, Gomes-Da-Costa SM, Cardoso EJBN (2003) Arbuscular mycorrhizal fungi associated with Araucaria angustifolia (Bert.) O. Ktze Mycorrhiza 13:211–215. https://doi.org/10.1007/s00572-003-0221-1
Moreira M, Baretta D, Tsai SM, Gomes-da-Costa SM, Cardoso EJBN (2007) Biodiversity and distribution of arbuscular mycorrhizal fungi in Araucaria angustifolia forest. Scientia agricola 64(4):393–399. https://doi.org/10.1590/S0103-90162007000400010
Moreira M, Baretta D, Cardoso EJBN (2012) Doses de fósforo determinam a prevalência de fungos micorrízicos arbusculares em Araucaria angustifolia. Ciência Florestal 22(4):813–820. https://doi.org/10.5902/198050987562
Silva RF, Antoniolli ZI, Leal L, Silva AS (2009) Ocorrência de fungos micorrízicos em espécies florestais na região central do estado do Rio Grande do Sul. R Bras Agrociência 15(1–4):65–70
Zandavalli RB, Stürmer SL, Dillenburg RL (2008) Species richness of arbuscular mycorrhizal fungi in forests with Araucaria in Southern Brazil. Hoehnea 35(1):63–68. https://doi.org/10.1590/S2236-89062008000100003
Vilcatoma-Medina C, Kaschuk G, Zanette F (2018) Colonization and spore richness of arbuscular mycorrhizal fungi in Araucaria nursery seedlings in Curitiba, Brazil. Hindawi Int J Agron 2018:1–6. https://doi.org/10.1155/2018/5294295
Nimer E (1979) Climatologia do Brasil. Recursos naturais e meio ambiente, Instituto Brasileiro de Geografia e Estatística (IBGE). Superintendência de Recursos Naturais e Meio Ambiente (SUPREN), Rio de Janeiro
ICMBio (2020) Plano de Manejo da Floresta Nacional de São Francisco de Paula. http://www.icmbio.gov.br/portal/flona-de-sao-francisco-de-paula?highlight=WyJmbG9uYSJd Accessed 26 June 2020
Sonego RC, Backes A, Souza AF (2007) Descrição da estrutura de uma Floresta Ombrófila Mista, RS, Brasil, utilizando estimadores não-paramétricos de riqueza e rarefação de amostras. Acta bot Bras 21(4):943–955. https://doi.org/10.1590/S0102-33062007000400019
Ferreira LM, Menezes EO, Silva PSC, Omena MTRN, Zanchetti F (2018) Plano de manejo do Parque Nacional de São Joaquim. ICMBio, Brasília
MMA- Ministério do Meio Ambiente (2020) Parque Nacional de São Joaquim. Cadastro Nacional de Unidades de Conservação. http://sistemas.mma.gov.br/cnuc/index.php?ido=relatorioparametrizado.exibeRelatorio&relatorioPadrao=true&idUc=165 Accessed 26 June 2020
Castañeda Ruiz RF, Heredia G, Gusmão LFP, LI D-W (2016) Fungal diversity of Central and South America. In: LI D-W (ed) Biology of Microfungi, Fungal Biology. Springer, Cham, pp. 197–217. https://doi.org/10.1007/978-3-319-29137-6_9
Trappe JM, Schenck NC (1982) Taxonomy of the fungi forming endomycorrhizae. In: Schenck NC (ed) Methods and principles of Mycorrhizal Research. The American Phytopathological Society, St. Paul, pp 1–9
Seifert K, Morgan-Jones G, Gams W, Kendrick B (2011) The genera of hyphomycetes. CBS Biodiversity Series no.9. CBS-KNAW Fungal Biodiversity Centre, Utrecht
Brower JE, Zar JH, Von Ende CN (1998) Field and laboratory methods for general ecology. Wm. C. Brown Publishers, Iowa
Chao A (1984) Non-parametric estimation of the number of classes in a population. Scand J Statist 11:265–270
Magurran AE (2004) Measuring biological diversity. Blackwell Science Ltd, Oxford. https://doi.org/10.2989/16085910409503825
Magurran AE (1988) Ecological diversity and its measurement. Princeton University Press, Princeton
Presenti N, Quatto P, Ripamonti E (2017) Bootstrap confidence intervals for biodiversity measures based on Gini index and entropy. Qual Quant 51:847–858. https://doi.org/10.1007/s11135-016-0443-x
Wilson JB (1991) Methods for fitting dominance/diversity curves. J Veg Sci 2:35–46. https://doi.org/10.2307/3235896
Dajoz R (1983) Ecologia Geral. Vozes, Petrópolis
Gower JC (2005) Principal coordinates analysis In: Armitage P, Colton T (eds) Encyclopedia of biostatistics. John Wiley & Sons Ltd, Chichester, pp 1–6. https://doi.org/10.1002/0470011815.b2a13070
Bray JR, Curtis JT (1957) An ordination of the upland forest communities of southern Wisconsin. Ecol Monogr 27:325–349. https://doi.org/10.2307/1942268
Anderson MJ (2017) Permutational multivariate analysis of variance (PERMANOVA). Wiley Stats Refs: Statistics Reference Online https://doi.org/10.1002/9781118445112.stat07841
Clarke KR (1993) Non-parametric multivariate analysis of changes in community structure. Aust J Ecol 18:117–143. https://doi.org/10.1111/j.1442-9993.1993.tb00438.x
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1):1–9. http://palaeo-electronica.org/2001_1/past/issue1_01.htm. Accessed 23 May 2020
McAleece N (1997) Biodiversity Professional Beta I. The Natural History Museum & The Scottish Association for Marine Science, London
Wallen RM, Perlin MH (2018) An overview of the function and maintenance of sexual reproduction in dikaryotic fungi. Front Microbiol 9:1–24. https://doi.org/10.3389/fmicb.2018.00503
Hyde KD, McKenzie EHC, KoKo TW (2011) Towards incorporating anamorphic fungi in a natural classification – checklist and notes for 2010. Mycosphere 2(1):1–88
Spatafora JW, Sung G-H, Johnson D, Hesse CN, O’Rourke B, Serdani M, Spotts R, Lutzoni F, Hofstetter V, Miadlikowska J, Reeb V, Gueidan C, Fraker E, Lumbsch T, Lücking R, Schmitt I, Hosaka K, Aptroot A, Roux C, Miller A, Geiser D, Hafellner J, Hestmark G, Arnold A, Büdel B, Rauhut A, Hewitt D, Untereiner W, Cole M, Scheidegger C, Schultz M, Sipman H, Schoch C (2006) A five-gene phylogeny of Pezizomycotina. Mycologia 98(6):1018–1028. https://doi.org/10.3852/mycologia.98.6.1018
Melo M, Araujo ACV, Chogi MAN, Duarte ICS (2018) Cellulolytic and lipolytic fungi isolated from soil and leaf litter samples from the Cerrado (Brazilian Savanna). Rev Biol Trop 66(1):237–245. https://doi.org/10.15517/rbt.v66i1.27768
Cai LK, Frang JI, Hyde KD (2006) Variation between freshwater and terrestrial fungal communities on decaying bamboo culms. Anton Leeuw 89:293–301. https://doi.org/10.1007/s10482-005-9030-1
Hernández-Restrepo M, Gené J, Castañeda-Ruiz RF, Mena-Portales J, Crous PW, Guarro J (2017) Phylogeny of saprobic microfungi from Southern Europe. Stud Mycol 86:53–97. https://doi.org/10.1016/j.simyco.2017.05.002
Monteiro JS, Sarmento PSM, Sotão HMP (2019) Saprobic conidial fungi associated with palm leaf litter in eastern Amazon. Brazil An Acad Bras Cienc 91(3):e20180545. https://doi.org/10.1590/0001-3675201920180545
Zhang Y, Schoch CL, Fournier J, Crous PW, de Gruyter J, Woudenberg JHC, Hirayama K, Tanaka K, Pointing SB, Spatafora JW, Hyde KD (2009) Multi-locus phylogeny of Pleosporales: a taxonomic, ecological and evolutionary re-evaluation. Stud Mycol 64:85–102. https://doi.org/10.3114/sim.2009.64.04
Boddy L (2016) Interactions with humans and other animals. In: Watkinson SC, Boddy L, Money NP (eds) The fungi, Academic Press, pp 293–336. https://doi.org/10.1016/B978-0-12-382034-1.00009-8
Punja ZK (1985) The biology, ecology, and control of Sclerotium rolfsii. Ann Rev Phytopathol 23:97–127. https://doi.org/10.1146/annurev.py.23.090185.000525
Maharachchikumbura SSN, Hyde KD, Gareth JEB, McKenzie EHC, Huang S-K, Abdel-Wahab MA, Daranagama DA, Dayarathne M, D’souza MJ, Goonasekara ID, Hongsanan S, Jayawardena RS, Kirk PM, Konta S, Liu J-K, Liu Z-Y, Norphanphoun C, Pang K-L, Perera RH, Senanayake IC, Shang Q, Shenoy BD, Xiao Y, Bahkali AH, Kang J, Somrothipol S, Suetrong S, Wen T, Xu J (2015) Towards a natural classification and backbone tree for Sordariomycetes. Fungal Diversity 72:199–301. https://doi.org/10.1007/s13225-015-0331-z
Ekanayaka AH, Hyde KD, Gentekaki E, McKenzie EHC, Zhao Q, Bulgakov TS, Camporesi E (2019) Preliminary classification of Leotiomycetes. Mycosphere 10(1):310–489. https://doi.org/10.5943/mycosphere/10/1/7
Boonmee S, D’souza MJ, Luo Z, Pinruan U, Tanaka K, Su H, Bhat DJ, McKenzie EHC, Jones EBG, Taylor JE, Phillips AJL, Hirayama K, Eungwanichayapant PD, Hyde HD (2016) Dictyosporiaceae fam. nov. Fungal Diversity 80:457–482. https://doi.org/10.1007/s13225-016-0363-z
Couturier M, Navarro D, Favel A, Haon M, Lechat C, Lesage-Meessen L, Chevret D, Lombard V, Henrissat B, Berrin JG (2016) Fungal secretomics of ascomycete fungi for biotechnological applications. Mycosphere 7(10):1546–1553. https://doi.org/10.5943/mycosphere/si/3b/6
McKenzie EHC, Buchanan PK, Johnston PR (2002) Checklist of fungi on kauri (Agathis australis) in New Zealand. New Zeal J Bot 40(2):269–296. https://doi.org/10.1080/0028825X.2002.9512788
Tokumasu S, Aoki TA, Oberwinkler F (1994) Fungal succession on pine needles in Germany. Mycoscience 35:29–37. https://doi.org/10.1007/BF02268525
Magurran AE, Henderson PA (2003) Explaining the excess of rare species in natural species abundance distributions. Nature 422:714–718. https://doi.org/10.1038/nature01547
McGill B, Etienne R, Gray J, Alonso D, Anderson M, Benecha H, Dornelas M, Enquist B, Green J, He F, Hurlbert A, Magurran AE, Marquet P, Maurer B, Ostling A, Soykan C, Ugland K, White E (2007) Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework. Ecol Lett 10:995–1015. https://doi.org/10.1111/j.1461-0248.2007.01094.x
Bills GF, Polishook JD (1994) Abundance and diversity of microfungi in leaf litter of a lowland rain forest in Costa Rica. Mycologia 86(2):187–198. https://doi.org/10.1080/00275514.1994.12026393
Lodge DJ, Cantrell S (1995) Fungal communities in wet tropical forests: variation in time and space. Can J Bot 73(S1):1391–1398. https://doi.org/10.1139/b95-402
Tsui CKM, Hyde KD, Hodgkiss IJ (2000) Biodiversity of fungi on submerged wood in Hong Kong streams. Aquat Microb Ecol 21:289–298
Gazis R, Chaverri P (2010) Diversity of fungal endophytes in leaves and stems of wild rubber trees (Hevea brasiliensis) in Peru. Fungal Ecol 3:240–254. https://doi.org/10.1016/j.funeco.2009.12.001
Prakash CP, Thirumalai E, Govinda Rajulu MB, Thirunavukkarasu N, Suryanarayanan TS (2015) Ecology and diversity of leaf litter fungi during early-stage decomposition in a seasonally dry tropical forest. Fungal Ecol 17:103–113. https://doi.org/10.1016/j.funeco.2015.05.004
Yuen TK, Hyde KD, Hodgkiss IJ (1999) Wood-degrading capabilities of tropical freshwater fungi. Material and Organismen 33:37–48
Yanna WHH, Hyde KD (2001) Fungal communities on decaying palm fronds in Australia, Brunei, and Hong Kong. Mycol Res 105(12):458–1471. https://doi.org/10.1017/S0953756201005214
Hyde KD, Chalermpongse A, Boonthavikoon T (1990) Ecology of intertidal fungi at Ranong mangrove, Thailand. Trans Mycol Soc Japan 31:17–28
Pinruan U, Hyde KD, Lumyong S, McKenzie EHC, Jones EBG (2007) Occurrence of fungi on tissues of the peat swamp palm Licuala longicalycata. Fungal Diversity 25:157–173
Lee S, Mel’nik V, Taylor JE, Crous PW (2004) Diversity of saprobic hyphomycetes on Proteaceae and Restionaceae from South Africa. Fungal Diversity 17:91–114
Barbosa FR, Maia LC, Gusmão LFP (2009) Fungos conidiais associados ao folhedo de Clusia melchiorii Gleason e C. nemorosa G. Mey. (Clusiaceae) em fragmento de Mata Atlântica, Bahia. Brasil Acta Bot Bras 23:79–84. https://doi.org/10.1590/S0102-33062009000100010
Thomas K, Chilvers GA, Norris RH (1989) Seasonal occurrence of conidia of aquatic hyphomycetes (fungi) in Lees Creek, Australian Capital Territory. Aust J Mar Freshw Res 40:11–23. https://doi.org/10.1071/MF9890011
Agrios GN (2004) Plant pathology. Academic Press, San Diego
Abdullah SK, Gené J, Guarro J (2005) A synopsis of the aero-aquatic genus Pseudaegerita and description of two new species. Mycol Res 109:590–594. https://doi.org/10.1017/S0953756205002819
Abdullah SK, Webster J (1983) The aero-aquatic genus Pseudaegerita. Trans Br Mycol Soc 80:247–254. https://doi.org/10.1016/S0007-1536(83)80007-2
Mastrobert AA, Mariath JEA (2003) Leaf anatomy of Araucaria angustifolia (Bertol.) Kuntze (Araucariaceae). Revista Brasil Bot. 26(3):343–353. https://doi.org/10.1590/S0100-84042003000300007
Reblová M, Seifert KA (2011) Discovery of the teleomorph of the hyphomycetes, Sterigmatobotrys macrocarpa and epitypification of the genus to holomorphic status. Stud Mycol 68:193–202. https://doi.org/10.3114/sim.2011.68.08
Butterfield BG (2003) Wood anatomy in relation to wood quality. In: Barnett JR, Jeronimidis G (eds) Wood quality and its biological basis. Blackwell Publishing Ltd, Oxford, pp 30–52
Minami E, Saka S (2003) Comparison of the decomposition behaviors of hardwood and softwood in supercritical methanol. J Wood Sci 49:73–78. https://doi.org/10.1007/s100860300012
Campbell MM, Sederoff RR (1996) Variation in lignin content and composition (mechanism of control and implications for the genetic improvement of plants). Plant Physiol 110:3–13. https://doi.org/10.1104/pp.110.1.3
Nag Raj TR, Kendrick B (1975) A monograph of Chalara and allied genera. Wilfrid Laurier University Press, Ontario
McKenzie EHC, O’Sullivan PJ, Wilkie JP (1992) A list of type specimens of New Zealand fungi held in DSIR Plant Protection Herbarium (PDD). Mycotaxon 43:77–156
Feinstein LM, Blackwood CB (2013) The spatial scaling of saprotrophic fungal beta diversity in decomposing leaves. Mol Ecol 22:1171–1184. https://doi.org/10.1111/mec.12160
Taylor JE, Hyde KD, Jones EBG (2000) The biogeographical distribution of microfungi associated with three palm species from tropical and temperate habitats. J Biogeography 27:297–310. https://doi.org/10.1046/j.1365-2699.2000.00385.x
Yanna WHH, Hyde KD, Goh TK (2001) Occurrence of fungi on tissues of Livistona chinensis. Fungal Diversity 6:167–180
Hyde KD, Ho WH, McKenzie EHC, Dalisay T (2001) Saprobic fungi on bamboo culms. Fungal Diversity 7:35–48
Prihatini I, Glen M, Wardlaw TJ, Ratkowsky DA, Mohammed CL (2015) Needle fungi in young Tasmanian Pinus radiata plantations in relation to elevation and rainfall. N Z J For Sci 45:25. https://doi.org/10.1186/s40490-015-0055-6
Acknowledgements
The authors would like to thank the Postgraduate Program in Botany (PPGBot/UEFS) and ICMBIO for permission to collect samples in the “Floresta Nacional de São Francisco de Paula” and the “Parque Nacional de São Joaquim” (Proc.: 42334-1).
Funding
The authors SSS and LFPG are grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for financial support (Proc.: 141475/2013–7 and 303062/2014–2, respectively).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Responsible Editor: LUCY SELDIN
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
da Silva, S.S., Costa, L.A. & Gusmão, L.F.P. Diversity of saprotrophic filamentous fungi on Araucaria angustifolia (Bertol.) Kuntze (Brazilian pine). Braz J Microbiol 52, 1489–1501 (2021). https://doi.org/10.1007/s42770-021-00531-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42770-021-00531-1