
Abstract
Tetragonula gressitti Sakagami is the new species of stingless bee discovered in Nagaland (2020). It was earlier found in Arunachal Pradesh (2013), South West China (2019) and Vietnam in 1978. The nest architecture of this particular species was never studied although there had been regular reports of its presence in forest areas by the local populace and all earlier reports were simply documented on the basis of collected samples from forests. This is the first detailed report on nesting characteristics of the stingless bee, T. gressitti, which were found located in the forest and village areas of Phek district in Nagaland during the early winters of 2019. The natural nests were found in tropical to sub-tropical evergreen forests on tree cavities, commonly at a height of 2.91 m ± 0.21 with a single entrance for each colony. The colonies were naturally active as seen and recorded from the activity of bees at their entrance gates during day time. The nests were extracted carefully with appropriate tools and transported to the experimental site. A total of seven natural nests were located for the study. Out of them, three nests were opened to observe the internal nest structure while the remaining nests were kept intact for further development and pollination studies. The observations revealed that the nest entrance on the external region was free from any wax or cerumen funnel/ tube and was meticulously kept clean by the bees. The nest entrances of all the colonies varied in size and shape. Internally, the entrance was connected to the brood through a waxy passage of 11.42 cm ± 0.73 length till it reached the involucrum. The brood was covered in a partial involucrum of 2.15 cm ± 0.23 thickness. The honey pots and pollen pots were found separated from the brood area and were connected through numerous small openings in the involucrum. The adult bees were observed to be deep black in colour. The required data on size of different cells and castes were also recorded accordingly.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Stingless bees are important pollinators belonging to the order Hymenoptera and family Apidae. These bees are close relatives of honey bees and bumble bees. Unlike honey bees, stingless bees do not possess sting and are quite smaller in size. They are highly developed social insects living in colonies found in all sub-tropical and tropical regions of the globe with more than 500 described species comprising the tribe Meliponini and sub tribe Meliponina (Michener 2013). The presence of nesting sites is essential for survival, maintenance and reproduction of Stingless bees (Hubbell and Johnson 1977; Batista et al. 2003; Eltz et al. 2003; Roubik 2006). Although their nesting habits are variable, most of the stingless bee species build their nests in several substrates ranging from subterranean cavities to arboreal ant nests (Campos 1987; Roubik 2006). The nest entrances of stingless bees vary in shape, size, length, colour etc. and is one of the important characteristic feature among different species (Chauhan et al. 2019). The chief building component of the colony is cerumen, a mixture of bee wax and plant resins (Rasmussen 2013). The common structure of stingless bee nests consists of entrance tunnels, brood cells, food storages (honey and pollen cells), cerumen, bitumen layers and involucrum (Sakagami et al. 1990; Michener 2007). The stingless bees are being reared in Nagaland since time immemorial and the diversity of stingless bees also vary from one district to another. A total of eight species of stingless bees were described from the Indian subcontinent (Rasmussen 2013) and out of them, six species are present in Nagaland (Chauhan and Singh 2020). Among the six species, like other Indian states, two species, viz. T. iridipennis and Lepidotrigona ventralis, are dominant and spread across the map. The nest architecture of these two species are well studied and described (Chauhan et al. 2019). A new species of stingless bee, T. gressitti, was recorded from Nagaland (Chauhan and Singh 2020), which had earlier been found from Southwest China (Pan et al. 2019), Arunachal Pradesh (Rathor et al. 2013) and South Vietnam (Sakagami 1978). However, the nest architecture of this species was never studied due to inability of locating their natural nests. The presence of T. gressitti was recently reported from Nagaland and it was observed that the commercial reproduction of queen and artificial rearing of stingless bee could have immense potential for honey production and pollination of crops grown under protected conditions. In order to commercialize this species of stingless bee, development of scientific hives is required on priority. Keeping in view the essentiality of hive designing for scientific rearing of T. gressitti, the present study was conducted to gather knowledge on the internal natural nest architecture of this species which will definitely help in designing scientific hives for this newly discovered species.
Material and methods
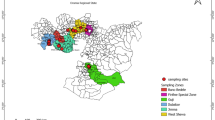
A survey was conducted in the forests and village areas around Phek district of Nagaland during the early winters of the year 2019. In the process, location of feral stingless bee habitat was identified in the forest along with recording of their nesting height preferences and necessary GPS coordinates (Fig. 1). A total of seven nests were identified from different locations. The nests were extracted from whole wooden logs and later, the cylindrical log hives were detached from the trunk and trimmed to about a metre in length, each containing a colony inside. The tedious process required precision and care so that the hives were not disturbed or opened at the ends while cutting them. In rare cases when any end portion is cut open, they are immediately sealed with available mud and leaves. The hives are then transported to the experiment site only in the evening. After bringing the nests to the experimental site, they were placed on tree branches and wooden shelves (Fig. 2). The colonies were allowed to settle down for some weeks and the foraging activity of the bees at the nest entrances were recorded from 0700—1700 h along with weather parameters. The three chosen colonies were then dissected to observe their nest architecture and colony structure. Their external features such as nest entrances were observed and data was recorded by taking their dimensions on shape and size. The internal nest structure of the hives were observed and the measurements were taken for wax tube (entrance), brood cells, honey pots and pollen pots. The size, shape and type of cells (brood, honey and pollen) were measured using vernier calipers along with the involucrum structure of the colony. The different materials used for construction of the colonies were also recorded. The number of brood layers as well as the number of cells per brood layer in the dissected nest along with the activity of bees at the entrance was used to determine the average population in a colony. The average numbers of guard bees were observed visually by noting the data on their presence at the entrance at two hours interval from 0700 – 1600 h.

Map and location of T. gressitti nesting

Extracted nest kept on tree branches
Results
A total of seven colonies of T. gressitti were located during the survey in the hilly regions of Phek district (forest and village areas) at an altitude range of 935-1167 m above mean sea level having GPS coordinates of 25° 37ʹ N and 94° 32ʹ E. All the colonies were found dwelling in tree cavities. The mean space occupied by the observed colonies in the nesting site was 1440 cm3 ± 43. The bees were found to prefer nesting at a mean height of 2.91 m ± 0.21 from the ground and the nesting height ranged between 2.64-3.22 m above from ground (Table 1).
The external features such as type of entrance, shape, material used for the construction and number of guards at the entrance were observed for all explored nests of T. gressitti. None of the nests had entrance funnel neither a deposition of cerumen or wax. The nest entrance were clear and free of any deposition (negligible deposition). Howsoever, the shape of nest entrance vary from round and oval to slit type with an opening diameter of 16.70 mm ± 0.34 in round type entrance while the slit entrance was 4.62 mm ± 0.21 (Fig. 3 a- b). The variation in shape of entrance might be due to the type of place available as choice for nesting naturally. The population of nest vary with the size of cavity. The entrances were guarded by workers (4.44 workers) in round type entrance while in slit type 2.67 workers were present at the entrance.

Nest entrance a Round type b Slit type
Activity of foragers at nest entrance
The activity of bees from the nest of T. gressitti started at 0700 h (4.73 bees/ 5 min) which then increased to maximum at 1100 h (20.15 bees/ 5 min (Table 2). The outgoing activity at the entrance started increasing (5.26, 17.68 and 28.64 outgoing bees/ 5 min) as the day progressed towards noon (0700 h- 1100 h) and eventually decreased towards evening (16.54, 11.67 and 2.33 bees/ 5 min at 1300 h, 1500 h and 1700 h). However, the incoming activity was minimum during early day hours (4.21 and 8.33 incoming bees at 0700 h and 0900 h) which was then found increasing in the early evening hours of the day (14.67 incoming bees/5 min at 1500 h). The maximum activity of bees was recorded at 23 ºC ± 0.54 and 65% ± 1.67 relative humidity.
Internal colony structure
The nest architecture study revealed that the nest entrance in majority of nests was oriented toward south west while in one of the nest it was oriented in south direction. The nest entrance extended inside the nest through a tunnel which was a mixture of wax and cerumen perforated tube, 11.42 cm ± 0.73 length and 1.96 cm ± 0.31 diameter connected to brood cells outside the incomplete involucrum having many openings. The involucrum was perforated 2.15 cm ± 0.23 thick with 3.33 ± 0.19 layers and the bees could go to any part of the nest without hindrance. The brood cells were located in the centre stacked over one another horizontally inside the involucrum and were not visible from outside. The brood layers were supported by pillars and the average number of brood cell layers was 8.33 ± 0.67 in a nest. A mean number of queen cells (3.67 queen cells) were recorded in the brood along with the presence of active queen. The size of T. gressitti queen cell was 8.91 ± 0.21 mm length and 5.16 mm ± 0.11 width while the worker cells were smaller in size (5.85 mm ± 0.20 and 4.29 mm ± 0.19). The honey pots were larger 24.86 × 20.14 mm, in length and width as compared to pollen pots (24.24 × 19.11 mm) and brood cells present in the colony (Fig. 4). The size of different castes was found to be different in T. gressitti, a similar trait as in T. iridipennis. The queen had protruded swollen abdomen, half covered with wings. However, in workers, the wings covered the whole abdomen. The workers were having black antennae with average total body length of 6.10 mm ± 0.06 while the queen was larger in size with average length of 9.89 mm ± 0.11 (Table 3). The pollen pots contained 4.361 g ± 0.43 of pollen/pot and 5.79 g ± 0.62 of honey/honey pot.

Internal nest architecture of T. gressitti showing honey pots and brood cells covered inside involucrum
The nest architecture of stingless bee species, T. gressitti, revealed presence of different types of cells (brood cells and food cells), and their respective positions in the nest. The brood needs to be warm and so an involucrum was present to prevent heat loss. Honey and pollen could ferment at higher temperature and as such, they were stored away from brood involucrum. The nest was lined with cerumen internally thereby insulating the nest from cold weather and free from infection as cerumen also have antimicrobial activity. Most of the nests were located at significant height which is an added protection from other enemies. The nest entrances were open outside but internally they were narrow which ultimately check attacks from enemies or predators. One hive had even closed a portion of the wide entrance and therefore, only a limited number of bees could enter or exit the nest at a time (Fig. 5). All the nests were found to have used the same materials (cerumen, wax, mud and resins) for nest construction.

Reduced nest entrance in T. gressitti
Discussion
Stingless bees have varied nesting sites depending on the species and space availability. The colonies of T. gressitti were found to prefer nesting at a mean height of 2.91 m ± 0.21 from the ground (Table 1). All the colonies were observed to nest in tree cavities. Chauhan et al. (2019) described the nesting preferences of T. iridipennis (2.57 feet) and Lepidotrigona ventralis (5.60 feet) from low hills of Nagaland. Similarly, Nayak et al. (2012) studied the nesting heights of Trigona and reported that stingless bees show distinct preference ranged between 28% – 47% at an elevation range of 0–10 feet and 11–15 feet from the ground. Likewise, the results have shown confirmation with the observations of Suriawanto et al. (2017), wherein they found the nests of Tetragonula spp. at a height ranging between 1.18 cm—321 cm (3.21 m) above the ground. Unlike T. gressitti which nested only on trees the nesting sites of T. fuscobalteata were found in the stone, brick wall, wooden wall, bamboo, and iron cavities, T. biroi in the wooden wall, stone, and brick wall cavities, T. sapiens in stone cavities, while T. laeviceps in wooden walls (Suriawanto et al. 2017).
The nest entrance of T. gressitti were smaller and varied in shape with few workers guarding the nest entrance. Chauhan et al. (2019) reported narrow nest entrances in T. iridipennis and L. ventralis. Similarly, Bui et al. (2020) found funnel shaped nest entrance in L. arcifera with narrow proximal ending. The narrow nest entrance of Melipona and other genera of stingless bees allow the nest to be defended by one or only a few guards positioned at the mouth of the entrance tube and variation of nest entrances related to defence and foraging activities of stingless bees (Biesmeijer et al. 2006). Barbosa et al. (2013) studied the nest architecture of Geniotrigona subterranea and they reported that the stingless bee nest entrance was circular in shape with a diameter of 0.85–1.20 cm. Other researchers like Syafrizal et al. (2014); Kelly et al. (2014) opined that nest entrance of stingless bees varied in shape, length and colour in Heterotrigona itama and G. thoracica.
The incoming activity at the nest of T. gressitti was maximum at evening hours and outgoing was maximum during late morning hours of the day. Similar results were recorded by Chauhan et al. (2019) in T. iridipennis and L. ventralis; Jaapar et al. (2018) while studying the foraging behaviour of Heterotrigona itama at nest entrance. They found that mean outgoing and incoming rate of stingless bee H. itama was maximum during morning hours (0800 -1000 h) and minimum in the evening 1600–1800 h. The bees made their nest in tree cavities and the size of cells (brood and food cells) were larger in T. gressitti. In T. iridipennis and L. ventralis colonies were bigger in size with differentiation in the internal arrangement of brood and food cells (Chauhan et al. 2019). In L. arcifera, the nest architecture was similar to L. ventralis (Bui et al., 2020). Similar observations were recorded by Barbosa et al. (2013) in G. subterranea, where they found that the nest involucrum consisted of many layers covering the brood having a mean area of 11.79 × 9.91 cm. The total population was observed to be in the range of 2726–11,074 bees/ nest in G. subterranea. The size of queen and worker bees are larger as compared to other reported stingless bee species. Similarly, the length of queen (7.46 mm), male (4.02 mm) and worker bees (4.10 mm) was less in T. iridipennis as compared to queen (8.16 mm), male (5.21 mm) and (5.59 mm), length in L.ventralis (Chauhan et al. 2019; Bui et al. 2020). The body size differed considerably among melliponini species and ranged from minute (T. pipoli, 2.5–3.0 mm body length) to medium 9–10 mm body length in P. occidentalis (Michener 2007; Jarau and Barth 2008). The queen bees can be easily separated from workers with large abdomen and short wings as an identifying feature.
There is a limited study on T. gressitti and so very less literature is available on the different aspects of this species. This pioneer study on nest architecture of T. gressitti will help in understanding and designing of new scientific hives for their scientific manipulation in honey production and pollination. Much more studies are required on the pollination potential of T. gressitti. The bigger size of T. gressitti workers may be effective in buzz pollination of crops especially tomato under protected conditions.
Data Availability
Photographs and videos are available if required.
References
Barbosa FM, Alves RM, Souza BA, Carvalo CAL (2013) Nest architecture of the stingless bee, Geotrigona subterranea (Fries, 1901) (Hymenoptera: Apidae: Meliponini). Neotropical Biota 13(1):147–152
Batista MA, Ramalho M, Soares AEE (2003) Nesting sites and abundance of Meliponini (Hymenoptera: Apidae) in heterogeneous habitats of the Atlantic rain forest, Bahia, Brazil. Lundiana 4:19–23
Biesmeijer JC, Roberts SPM, Reemer M (2006) Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313:351–354
Bui M, Singh HK, Alemnila AO, Chauhan A, Behere GT (2020) Diagnistics of wild stingless bees from North East India. Indian Journal of Entomology 82(12):337–342
Campos LAO (1987) Abelhas indigenas sem ferrao: o que sao? Info. Agrop 13 (147)
Chauhan A, Singh HK (2019) Nest architecture studies of stingless bees, Tetragonula iridipennis and Lepidotrigona ventralis in Nagaland. Indian Journal of Entomology 81(1):158–164
Chauhan A, Singh HK (2020) A new record of stingless bee, Tetragonula gressitti sakagami (hymenoptera: apidae: meliponini) from Nagaland. India International Journal of Farm Sciences 10(2):83–87
Eltz T, Bruhl CA, Imiyabir Z, Linsenmair KE (2003) Nesting and nest trees of stingless bees (Apidae: Meliponini) in lowland dipterocarp forests of Sabah, Malaysia, with implications for forest management. For Ecol Manag 172:301–313
Hubbell SP, Johnson LK (1977) Competition and nest spacing in a tropical stingless bee community. Ecology 58:949–963
Jaapar MF, Jajuli R, Mispan R, Ghani, IA (2018) Foraging behaviour of stingless bee Heterotrigona itama (Cockerell, 1918) (Hymenoptera: Apidae: Meliponini). In: AIP Conference proceedings 1940, 020037
Jarau S, Barth FG (2008) Stingless bees of the Golfo Dulce region, Costa Rica (Hymenoptera, Apidae, Apinae, Meliponoini). Stapfia 88:267–276
Kelly N, Farisya MSN, Kumara TK, Marcela P (2014) Species diversity and external nest characteristics of stingless bees in Meliponiculture. Pertanika Journal of Tropical Agricultural Science 37(3):293–298
Michener CD (2013) The Meliponini. In: Vit P, Pedro SRM, Roubik DW (eds) Pot-Honey: A legacy of stingless bees. Springer, New York, pp 3–17
Michener CD (2007) The bees of the world, 2nd edn. Johns Hopkins University Press, Baltimore, p 972p
Nayak PP, Reddy SM, Jayaprakash (2012) Nesting pattern preferences of stingless bee, Trigona iridipennis Smith (Hymenoptera: Apidae) in Jnanabharathi Campus, Karnataka, India. International Research Journal of Biological Sciences 2(2):44–50
Pan P, Wang S, Zhong Y, Xu H, Wang Z (2019) New record of the stingless bee Tetragonula gressitti (Sakagami, 1978) in Southwest China (Hymenoptera: Apidae: Meliponini). J Apic Res https://doi.org/10.1080/00218839.2019.1688494
Rasmussen C (2013) Stingless bees (Hymenoptera: Apidae: Meliponini) of the Indian subcontinent: diversity, taxonomy and current status of knowledge. Zootaxa 3647(3):401–428
Rathor V S, Rasmussen C, Saini MS (2013) New record of the stingless bee Tetragonula gressitti from India (Hymenoptera: Apidae: Meliponini). Journal of Melittology 7:1–5
Roubik DW (2006) Stingless bee nesting biology. Apidologie 37:124–143
Sakagami SF (1978) Tetragonula stingless bees of the continental Asia and Sri Lanka. Journal of the Faculty of Science Hokkaido University series VI Zoology 21(2):165–247
Sakagami SF, Inoue T, Salmah S (1990) Stingless bees of central Sumatra. In: Ohgushi R, Roubik DW (eds) Sakagami SF. Natural History of social wasps and bees in equatorial Sumatra, Hokkaido University Press Sapporo, pp 125–137
Suriwanto N, Atmovidi T, Kahono S (2017) Nesting sites characteristics of stingless bees (Hymenoptera: Apidae) in central Sulawesi. Indonesia Journal of Insect Biodiversity 5(10):1–9
Syafrizal TD, Yusuf R (2014) Biodiversity and habitat of Trigona at secondary tropical rain forest of Lempake education forest, Samarinda. Kalimantan Timur Journal Teknologi Pertanian 9(1):34–38
Acknowledgement
The authors are indebted to Project Coordinator All India Coordinated Research Project on Honey Bees & Pollinators for providing financial support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Consent to participate
We hereby provide the consent to participate.
Consent to publish
We hereby provide the consent to publish our paper in this journal.
Conflict of interest
There is no conflict of interest for publication of manuscript.
Rights and permissions
About this article

Cite this article
Chauhan, A., Singh, H.K. Nest architecture of stingless bee, Tetragonula gressitti Sakagami from Nagaland, India. Int J Trop Insect Sci 41, 3099–3104 (2021). https://doi.org/10.1007/s42690-021-00503-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42690-021-00503-w