
Abstract
The objective of this study is intended to explore the effect of maintain high efficiency water and fertilizer supply for improving the tomato production and soil bacterial communities by negative pressure irrigation (NPI) in greenhouse. The pot culture experiments consisted of three irrigation treatments: conventional irrigation (CI), drip irrigation (DI) and NPI, which have been conducted in greenhouse for two consecutive years. The study found that NPI reduced water consumption remained stable with slight variations on dynamic soil water content, as well as increased the water use efficiencies compared with CI and DI, respectively. The NPI reduced water consumption by 20% and 18% compared to CI in 2016 and 2017, respectively. Under NPI treatment, the variations in soil water content were stabilized, within the range of 12.3–14.7% in the two years, which was smaller than that of CI and DI treatments of 8.6–15.6%. Moreover, the tomato yields, fruit quality, uptake quantitates of nutrition and soil rhizosphere microbial diversity were increased in the NPI treatment comparison with the CI and DI treatments. With NPI, the yield of tomato was significantly increased by 26% and 8, 41 and 17% compared to that under CI and DI treatments in 2016 and 2017, respectively. NPI was also considered to benefit plant growth by increasing the abundances of Bacillus, Streptomyces and Pseudolabrys at the genus level. Therefore, NPI should be utilized as an advanced technique for saving water and producing high yields in future sustainable agriculture.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Tomato (Solanum lycopersicum L.) is a popular commercial vegetable that is widely cultivated in China, and it is rich in carotene, lycopene, vitamin C and B [1, 2]. Tomato plants require extensive amounts of nutrients and water, and water and fertilizer utilization need to be improved in areas with limited water and mineral resources [3, 4]. However, excess fertilizer input will cause environmental pollution and reduce the quality of fruit [5, 6]. Therefore, an agricultural irrigation measure that could increase water and nutrient efficiencies needs to be explored. Drip irrigation (DI) is better than conventional irrigation (CI) from the aspect of being more effective at saving water and fertilizer [4, 7, 8]. DI has matured and is a widely used irrigation technology in agricultural production [7, 8]. Maximum yields and water use efficiencies can be obtained from higher utilization of water and fertilizer, and of 25% of those can be saved by drip irrigation [9]. However, DI shortcomings still exist; for example, water can easily transpire from the soil surface, and water cannot be supplied to meet the needs of plants. Thus, advanced technologies are required to solve this shortcoming.
Negative pressure irrigation (NPI) is a new technology used to save water [8, 10]. NPI was initially used in greenhouses and supplies water depending on the physiological characteristics of plants and the soil tension [11, 12]. Consequently, NPI can consistently acquire and supply water for plants. Under the NPI water supply conditions, the unsaturated soil drives the gravitational movement of water into the soil [8, 10]. As a result, soil leakage and evaporation losses are reduced to achieve the effect of saving water [13]. Moreover, it was found that under high temperature and high sunlight conditions, NPI could reduce the water supply by 45% compared to that required by a sprinkler irrigation system [14, 15]. However, planting tomatoes improves the integration of water and fertilizer in an NPI system, but few studies have concentrated on the mechanisms, especially in the nutrient content and microbial diversity of the rhizosphere.
Rhizosphere microorganisms play an important role in a soil ecosystem [16, 17]. The variations in soil microbial community structure reflect the productivity and stability characteristics of soil, which are pivotal to the growth of crops [16, 17]. The lack or excess of water will affect the regular physiological processes of microorganisms [18]. Several studies have found that improvements in the diversity of microorganisms were related to the growth of crops [17, 18]. Microorganism diversity can promote the populations of Actinomycetes and other bacteria that are beneficial to soil and reduce the number of fungi [19, 20]. However, few studies have focused on the detailed taxonomic changes in the microbial communities in the rhizosphere soil under different irrigation system conditions.
Therefore, we tested the hypothesis that effective NPI is linked to stable supply and save water, improve tomato production and diversity of rhizosphere soil. The objectives of this study are to: (1) determine the response of water utilization to different irrigation practices and (2) measure the effects of tomato production and bacterial communities of the rhizosphere soil under different irrigation treatments.
2 Materials and methods
2.1 Site description
Pot experiments were conducted in the greenhouse of the Chinese Academy of Agricultural Sciences (116.3°E, 39.9°N) in Beijing, China, during 2016 and 2017. The basic soil physical and chemical characteristics were as follows: bulk density, 1.41 g/cm3; pH, 7.53; organic matter, 1.32%; total N, 1.63 g/kg; Olsen P, 13.21 mg/kg; and available potassium, 96.32 mg/kg.
2.2 Experimental design
The soil pot (42 × 26 × 30 cm) experiment was conducted in the greenhouse, and the FH9 tomato genotype was employed as plant material. Each pot contained 30 kg of loam. There were three treatments including CI, DI and NPI. There were three replicates for each treatment in this study, of which three pots as one replication.
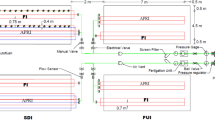
The NPI system was constructed using a negative pressure device that provides continuous water and fertilizer according to the crop needs (Invention Patents, China, ZL201110093923.2 and ZL201310554433.7). The NPI system consists of four parts: a liquid storage tank, voltage generator, clay pipe and planting pot (Fig. S1). The irrigation emitter (clay pipe) was formed by a porous ceramic pipe 20 cm in length that was inserted to a depth of 15 cm in the soil, and it easily established contact with the tomato roots when the entire NPI system is in a sealed state. When the NPI system is running, the irrigation emitter supplies water to the plant, and the amount of liquid at the surface of the liquid storage tank will decline. The amount of water consumption can be obtained by recording the amount of water in the storage tank every day. According to the previous NPI experiments, the pressure value of the voltage generator is − 5 kpa [10, 12]. In the NPI treatments, the plants absorbed a nutrient solution every day. The CI and DI treatments were set to consistent irrigation quantities and ensure the proper growth of tomato depending on the maintain field capacity, and the total amounts of irrigation were 53.6 and 59.2 L in the years of 2016 and 2017, respectively. The CI treatment provided water to the plants once each week, and approximately, 2 L of water and fertilizer were dissolved together and then added to the pot at the same time. The flow rate of the DI treatment was 2 L per hour, and the pressure of the DI was 0.3 Mpa. The same amounts of water and fertilizer were used in the CI and DI treatments each time.
The standard concentration was used for the tomato nutrient solution formula in this experiment [4, 8]. The total nutrient solution was: 354 mg L−1, Ca(NO3)2·4H2O; 404 mg L−1 KNO3; 77 mg L−1 NH4H2PO4; 246 mg L−1 MgSO4·7H2O and micronutrients.
2.3 Measurements
The soil water content was measured gravimetrically in the 0–30 cm soil profile by the drying (105 °C for 24 h) method, and the soil water content was sampled every 2 weeks after the tomato seedlings were transplanted. Soil water content (%) = wet soil weight—dry soil weight/dry soil weight × 100%. Water use efficiency (WUE, g m−3) = yield/water consumption × 100%, where yield is the fruit weight (g pot−1). The rhizosphere soil was sampled at harvest time in 2017. For rhizosphere soil collection, we first removed the plants from the pot and shook off the soil around the root system and then washed the soil attached to the root system with sterile water. The rhizosphere soil was collected by centrifugation at 10,000 rpm for 10 min. These soils were used for the determination of soil nutrients and bacteria diversity.
The fruit yield and quality (vitamin C, soluble sugar and nitrate) and the plant N, P and K contents were measured after ripening of the fruit. Plants were dried at 105 °C for 30 min and then kept at 75 °C until completely dry to determine the dry weight biomass. The total N, P and K contents of the plant and soil were measured using the semimicro Kjeldahl procedure combined with a N analyzer (Kjeldahl 2300; FOSS, Hoganas, Sweden), a phosphorus–molybdate blue color reaction, and flame photometer and 1 mol L−1 NH4AC extraction method, respectively [10, 21]. The yield of each plant was measured after harvest, and the fruit samples were taken back to the laboratory to determine the nitrate content by salicylic acid colorimetry, soluble sugar by anthrone colorimetry and vitamin C by 2,6-chlorophenol spectrophotometry [10].
Determination of soil bacterial diversity: The total DNA in the soil was extracted by the PowerSoil DNA Isolation Kit, and 30 ng DNA samples were amplified by PCR and sequenced by BGI Tech in Shenzhen. The main measurement indexes and calculation methods are as follows: Shannon index, \(H = - \sum {(P_{i} )(\ln P_{i} )}\), where Pi expresses the proportional number in a specific group relative to the total number [22]. The unique sequences were aligned with the Silva 106 database. The operational taxonomic units (OTUs) that were analyzed were all classified at a dissimilarity of 0.03 degrees. Sequences that could not be assigned to any division were labeled unclassified. Richness (Ace and Chao1) and Shannon diversity indexes were generated.
2.4 Statistical analysis
All data in the experiments were statistically analyzed by ANOVA using Excel 2016 software and SAS 9.1 (SAS Inc., Cary, NC, USA).
3 Results
3.1 Yield and quality of tomato fruit
The yield and quality of the tomato fruit were significantly increased by the NPI system (Table 1). The tomato yield in the NPI treatment was considerably higher than that of the CI and DI treatments, which was 26% and 8% higher in 2016, and 41% and 17% higher in 2017, respectively. The shoot biomass in the NPI treatment was substantially higher compared to the CI and DI treatments in the 2 years.
The NPI treatment improved the quality of the tomato fruit mainly via the improvement of the vitamin C and soluble sugar contents and decreases in the nitrate content (Table 1). The NPI treatment had the highest vitamin C concentration, and it was increased by 9% and 14%, and 16% and 6% compared to the concentrations in the CI and DI treatments in 2016 and 2017, respectively. NPI also increases the soluble sugar content, which was increased by 13% and 21% compared to the CI treatment in the two years, respectively. Meanwhile, the nitrate content of tomato fruit was the lowest in the NPI treatment. The results showed that NPI could significantly promote the growth and quality of tomatoes in a greenhouse.
3.2 Utilization of water
The NPI system could reduce water consumption and enhance soil water content and WUE compared to the CI and DI treatments (Table 2; Fig. 1). For example, NPI reduced water consumption by 10.5 L and 10.9 L compared with the CI and DI treatments throughout the tomato growth period, which resulted in water savings of 20% and 18% in 2016 and 2017, respectively. Additionally, the WUE was improved by the NPI treatment with 56% and 73% increases compared with CI in the 2 years, respectively. The results indicated that the NPI system could reduce water consumption and improve water use efficiency.

Effects of different irrigation treatments on soil water content of tomato. CI represents conventional irrigation, DI represents drip irrigation, and NPI represents negative pressure irrigation
The soil water content remained more stable in the NPI treatment compared to that in the CI and DI treatments throughout the tomato growth stages in 2016 and 2017 (Fig. 1). For the NPI treatment, the variations in soil water content were stabilized, with ranges of 12.3–14.7% in the two years. Meanwhile, the ranges in the CI and DI treatments exhibited much larger fluctuations than observed in the NPI treatment, and the variations were 8.6–15.6% and 10.9–14.8%, respectively. This result proves that NPI can maintain soil water stability.
3.3 Analysis of nutrient contents in the rhizosphere soil and plants
NPI decreased the available potassium content in the rhizosphere soil of tomato, as well as the nitrogen and phosphorus contents (Table 3). Table 3 suggests that NPI significantly decreased the available potassium content in the rhizosphere soil with 18% and 14% compared to the contents in the CI treatments in the years of 2016 and 2017, respectively. NPI had a tendency to reduce the available nitrogen content by 11% and 20% in comparison with the CI treatment in 2016 and 2017, respectively. However, little change occurred in the available P content between the different irrigation treatments. The results indicated that NPI could result in more nutrient consumption in the rhizosphere soil.
NPI significantly increased the nutrient contents of the tomato plants (Table 3). In comparison with the CI and DI treatments, the nitrogen contents of the plants were noticeably increased by 38% and 17, 53 and 23% in 2016 and 2017, respectively. Similarly, the P and K contents of the plants were also significantly improved.
3.4 Bacterial diversity in rhizosphere soil
NPI significantly increased the bacterial community in the rhizosphere soil, as indicated by the highest Chao1, ACE and Shannon index values determined by Illumina genome analysis (Table 4). For example, in the rhizosphere soil, the Chao1 values were increased by 59% and 8%, and the ACE indexes were increased by 59% and 5% in the NPI treatments compared to those in the CI and DI treatments, respectively. Similarly, the Shannon indexes of the rhizosphere soil showed higher diversity in the NPI treatments.
The Venn diagram shows the numbers of unique OTUs (operational taxonomic units) in the different soil samples as well as the overlaps of common OTUs, and the samples are displayed in different colors (Fig. 2). The abundance OTUs indicated the degrees of species abundance in the samples. As shown in Fig. 2, the highest number of OTUs was in the NPI treatment (green circle), and there were 2143 OTUs with 51% and 3% increases in comparison with the yellow circle (CI) and purple circle (DI), respectively.

Venn diagram of OTUs in rhizosphere soil under different irrigation conditions. CI is yellow, DI is purple, and NPI is green. OTUs are operational taxonomic units
The differences in the relative abundance of the bacterial community at the genus level in the three irrigation treatments are summarized in Table 5. The most common bacteria at genus level were Sphingomonas, which accounted for 4.96% in the CI treatment, 5.12% in the DI treatment and 6.04% in the NPI treatment. The results indicated that reads of Bacillus, Streptomyces and Pseudolabrys were more common in the NPI treatment than in the CI and DI treatments. For example, the percentages of Pseudolabrys were 0.26, 0.45 and 0.67% in the CI, DI and NPI treatments, respectively. The results indicated that irrigation treatment could significantly change the abundance of bacterial communities at the genus level.
4 Discussion
It has been well documented that NPI can yield benefits and save water in most crops [8, 10, 12]. The results of this study in two consecutive years also agree that NPI can increase the tomato yield by 8–41% and save an average of 18–20% of water compared with CI and DI (Tables 1, 2). Meanwhile, it is widely noted that NPI systems control the negative pressure values of a water feeder, which can provide persistent water and automatically supply water to the soil in the root layer according to the crop needs [8, 10,11,12], and this ensures that the soil water content during the crop growth period remains relatively stable (Fig. 1); therefore, NPI also contributes to improvements in the yield and WUE (Table 1 and 2). In the current study, we found that NPI could continue to provide water to keep the soil water content within a more stable range than in the CI and DI treatments (Fig. 1). The soil water content in the NPI treatment was maintained at approximately 12.3–14.7% with smaller variations than in the other treatments, which was probably because the water feeder at a depth of 15 cm in the soil is in direct contact with tomato roots and can take full advantage of the rhizosphere soil and nutrients. Furthermore, the soil moisture of the root zone at 0–20 cm had a greater effect on crop growth than other factors [8, 12]. In comparison, CI and DI mainly supply water to the soil surface, which implies that a large proportion of irrigated water and fertilizer would easily transpire and percolate outside of the root zone; therefore, water and fertilizer use efficiency will be reduced, and groundwater contamination would be aggravated.
The NPI system can improve the yields and qualities of the crops, which is mainly caused by the direct supply of water and fertilizer to the root zone [8, 10, 12]. The present study was in agreement with the previous studies, and improved yields and qualities of tomato were found in the greenhouse experiments in the NPI treatments compared with the CI and DI treatments (Tables 1, 3). This result suggested that the irrigation mode can affect the yields and qualities of tomato because CI and DI systems cannot directly and precisely contact the rhizosphere zone; thus, it is easy to cause the fixation and loss of nutrients, which will reduce the uptake nutrients from the roots and increase the accumulation of dry matter and reduce the yields and qualities of the crop. Meanwhile, the macroelements of nitrogen (N), phosphorus (P) and potassium (K) play an important role in the yields and qualities of plants [23,24,25,26]. We found that the NPK contents of the plants were significantly higher in the NPI treatment than in the CI and DI treatments (Table 3), which indicates that the higher frequency of NPI over a long period can maintain consistent nutrient concentrations in the root zone, promote the development of the root system, enhance the photosynthetic rate and chlorophyll content to promote photosynthesis and nutrient uptake and accumulation and finally increase the crop yield and quality [8, 10]. Simultaneously, deficits and excesses of water can regulate the quality of tomato during the fruit setting period of tomato plants [24]. Badr et al. [27] found that water is involved in the regulation of fruit penetration, and it can increase sugar concentrations through the phloem into the fruit. However, excessive water supplies can reduce the qualities of VC and soluble sugar [4, 24]. In this study, NPI improved tomato quality, which might due to the steady water supply and the avoidance of interference from excessively wet and dry soil moisture conditions. In addition, the application of potassium fertilizer could increase the soluble sugar and VC contents in tomato fruit, and potassium ions can affect the uptake of nitrate and promote nitrogen metabolism in crops [28], which partly proves that increases in the uptake of potassium can improve the quality of tomato (Table 3).
High throughput sequencing technology has greatly expanded our exploration of the relationship between microorganisms and plants [19]. The microbial communities in the rhizosphere are influenced by plant species, soil type and water content [29, 30], and high-quality soils exhibit higher bacterial diversity [20]. In the current study, based on three different soil water content treatments, we found that NPI results in the highest microbial diversity (Fig. 2; Table 4). This result indicated that a steady water supply results in higher diversity than alternating dry and wet soil, which is probably because appropriate water supply can dissolve minerals in the rhizosphere soil, thereby promoting the absorption of nutrients by microorganisms, while water affects soil aeration and regulates microbial communities [31, 32]. Due to the steady water emissions in the NPI system, air permeability is maintained in the tomato rhizosphere, which is beneficial for nutrient absorption by aerobic bacteria; however, when the roots of tomatoes are under drought or flooded conditions, the growth of microorganisms is prevented [4, 11]. Simultaneously, the NPI treatment had a significant effect on the taxonomy of bacteria at the genus level (Table 5), and several probiotics potentially involved in the promotion of tomato growth were found. For example, Bacillus, Pseudolabrys and Streptomyces, which have the ability to promote crop growth [33,34,35], were detected as dominant genera in this study, and the abundances of these genera were highest in the NPI treatment (Table 5). In addition, Bacillus species can be considered aerobes or facultative anaerobes [36], which also proved that the relative contents of aerobe bacteria can be improved by good aeration conditions in the rhizosphere under NPI. Meanwhile, Streptomyces, which is the largest genus of Actinobacteria, was distributed in the soil with suitable water contents and established aerobic life, and most of the antibiotics were produced by this genus, which is also considered as a probiotic bacterium [37,38,39,40]. It was proved that the relative abundances of Bacillus and Streptomyces can be increased under the better-aerated NPI treatment, thus promoting the growth of tomato. Since rhizosphere of the root zone has a high number of bacteria, a more thorough search for probiotics should be explored in the future. Therefore, this research contributed to the understanding of how the NPI system can improve the number of beneficial microorganisms, optimize the soil ecological environment and improve crop yield and quality.
The results indicate that NPI provides suitable and steady soil water content for tomato production while reducing water consumption and increasing the WUE in a greenhouse. The yields and qualities of tomato plants were strengthened in the NPI treatment compared to the CI and DI treatments, and the uptakes of N, P and K by the plants were significantly improved. Meanwhile, NPI makes it more possible for the bacterial diversity of the rhizosphere soil as well as the abundances of Bacillus, Streptomyces and Pseudolabrys at the genus level to increase. NPI can be used as a new irrigation method, and this study provides the scientific and theoretical basis for the sustainable development of agriculture.
References
Ren T et al (2010) Root zone soil nitrogen management to maintain high tomato yields and minimum nitrogen losses to the environment. Sci Hortic 125:25–33
He X et al (2016) Environmental impact assessment of organic and conventional tomato production in urban greenhouses of Beijing city, China. J Clean Prod 134:251–258
Ismail SM et al (2008) Influence of single and multiple water application timings on yield and water use efficiency in tomato (var. First power). Agric Water Manage 95:116–122
Wang C et al (2015) Assessing the response of yield and comprehensive fruit quality of tomato grown in greenhouse to deficit irrigation and nitrogen application strategies. Agric Water Manag 161:9–19
Fan Z et al (2014) Conventional flooding irrigation causes an overuse of nitrogen fertilizer and low nitrogen use efficiency in intensively used solar greenhouse vegetable production. Agric Water Manag 144:11–19
Cao J et al (2015) Changes in potential denitrification-derived N2O emissions following conversion of grain to greenhouse vegetable cropping systems. Eur J Soils Biol 68:94–100
Liu H et al (2013) Drip irrigation scheduling for tomato grown in solar greenhouse based on pan evaporation in North China Plain. J Integr Agric 12:520–531
Li YK et al (2017) Comparison of drip fertigation and negative pressure fertigation on soil water dynamics and water use efficiency of greenhouse tomato grown in the North China Plain. Agric Water Manag 184:1–8
Marino S et al (2014) Use of soil and vegetation spectroradiometry to investigate crop water use efficiency of a drip irrigated tomato. Eur J Agron 59:67–77
Zhao XJ et al (2017) Effect of different potassium levels on the growth of bok choy under negative pressure. Sci Agric Sin 50:689–697 (in Chinese)
Wang J et al (2016) Water and salt movement in different soil textures under various negative irrigation pressures. J Integr Agric 15:1874–1882
Long H et al (2018) Design and experiment of heavy-type negative pressure value used for negative pressure irrigation. Trans CSAE 34:85–92 (in Chinese)
Amtmann A et al (2008) The effect of potassium nutrition on pest and disease resistance in plants. Physiol Plant 133:682–691
Farneslli M et al (2015) High fertigation frequency improves nitrogen uptake and crop performance in processing tomato grown with high nitrogen and water supply. Agric Water Manag 154:52–58
Nalliah V et al (2009) Evaluation of a plant-controlled subsurface drip irrigation system. Biosyst Eng 102:313–320
Yuan S et al (2014) Evaluation of bacillus-fortified organic fertilizer for controlling tobacco bacterial wilt in greenhouse and field experiments. Appl Soil Ecol 75:86–94
Wu K et al (2014) Effects of bioorganic fertilizer plus soil amendment on the control of tobacco bacterial wilt and composition of soil bacterial communities. Biol Fert Soils 50:961–971
Bernard E et al (2012) Compost, rapeseed rotation, and biocontrol agents significantly impact soil microbial communities in organic and conventional potato production systems. Appl Soil Ecol 52:29–41
Borrero C et al (2006) Tomato Fusarium wilt suppressiveness. The relationship between the organic plant growth media and their microbial communities as characterized by Biolog. Soil Biol Biochem 38:1631–1637
Bottomley P et al (2006) Responses of soil bacterial and fungal communities to reciprocal transfers of soil between adjacent coniferous forest and meadow vegetation in the Cascade Mountains of Oregon. Plant Soil 289:35–45
Gao X et al (2014) Root interactions in a maize/soybean intercropping system control soybean soil-borne disease, red crown rot. PLoS ONE 9:e95031
Schloss PD et al (2009) Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75:7537–7541
Chen Q et al (2004) Evaluation of current fertilizer practice and soil fertility in vegetable production in the Beijing region. Nutr Cycl Agroecosyst 69:51–58
Patane C et al (2010) Effects of soil water on yield and quality of processing tomato under a Mediterranean climate conditions. Agric Water Manage 97:131–138
Christian Z et al (2014) Potassium in agriculture–status and perspectives. J Plant Physiol 171:656–669
Gao X et al (2018) Potassium-induced plant resistance against soybean cyst nematode via root exudation of phenolic acids and plant pathogen-related genes. PLoS ONE 13:0200903
Bakr A et al (1997) Trials to reduce nitrate and oxalate content in some leafy vegetables. J Sci Food Agric 73:169–178
Chen J et al (2013) Quantitative response of greenhouse tomato yield and quality to water deficit at different growth stages. Agric Water Manage 129:152–162
Berg G (2009) Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol 68:1–13
Li X et al (2014) Dynamics of the bacterial community structure in the rhizosphere of a maize cultivar. Soil Biol Biochem 68:392–401
Chaparro JM et al (2012) Manipulating the soil microbiome to increase soil health and plant fertility. Biol Fertil Soils 48:489–499
Berendsen R et al (2012) The rhizosphere microbiome and plant health. Trends Plant Sci 17:478–486
Tian B et al (2007) Bacteria used in the biological control of plant parasitic nematodes: populations, mechanisms of action, and future prospects. FEMS Microbiol Ecol 61:197–213
Aliye N et al (2008) Evaluation of rhizosphere bacterial antagonists for their potential to bioprotect potato (Solanum tuberosum) against bacterial wilt (Ralstonia solanacearum). Biol Control 47:282–288
Algam S et al (2010) Effects of Paenibacillus strains and chitosan on plant growth promotion and control of Ralstonia wilt in tomato. J Plant Pathol 92:593–600
Turnbull P (1996) Bacillus. In: Baron S (ed) Barron’s medical microbiology, chapter 15, 4th edn. University of Texas Medical Branch at Galveston, Galveston
Huang MY (2011) Identification and fermentation of antagonistic bacterium against Ralstonia solanacearum. Microbiol China 38:214–220
Kämpfer P (2006) The family Streptomycetaceae, part I: taxonomy. In: Dworkin M, Falkow S et al (eds) The prokaryotes. Springer, New York, pp 538–604
Wei Z et al (2011) Efficacy of bacillus-fortified organic fertiliser in controlling bacterial wilt of tomato in the field. Appl Soil Ecol 48:152–159
Hu W et al (2017) Microbial communities in the cysts of soybean cyst nematode affected by tillage and biocide in a suppressive soil. Appl Soil Ecol 119:396–406
Acknowledgements
This research was financially supported by the project for Modulation technology for water and nutrients in the soil environment for plant growth (2013AA102901) and the Special Fund for Agro-scientific Research in the Public Interest of China (201503120).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Gao, X., Zhang, S., Zhao, X. et al. Stable water and fertilizer supply by negative pressure irrigation improve tomato production and soil bacterial communities. SN Appl. Sci. 1, 718 (2019). https://doi.org/10.1007/s42452-019-0719-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42452-019-0719-6