
Abstract
The beneficial microbes colonizing the plant root, designated as plant growth promoting rhizobacteria (PGPR), increase the crop productivity and present an attractive and promising way to substitute chemical fertilizers, pesticides, and other harmful supplements. Hence, the present study is envisaged to identify, screen and characterize PGPR from rhizospheric soil of Brassica juncea (Indian mustard). Three sites out of major mustard producing areas under Allahabad in Eastern Uttar Pradesh, India were targeted. Total 20 bacterial isolates were identified and purified from these regions based on occurrence percentage and they were further tested in vitro for plant growth promoting (PGP) traits. Among 20 isolates, only two solubilized phosphate, three isolates produced hydrogen cyanide, 14 assimilated symbiotic nitrogen, two showed siderophore production and nine produced indole-3-acetic acid. Based on PGP tests, three potential isolates were selected for plant growth promotion. Further, this study also investigates the potential use of agricultural wastes as carrier materials, viz. charcoal, sawdust, wheat bran and rice husk, for preparation of bioformulations (or biofertilizers) using three PGPR isolates for viability assessment.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Microorganisms have been employed as an important practice in agriculture aimed to improve availability of nutrients for plants (Freitas et al. 2007; Vishwakarma et al. 2016). For sustainable agriculture practices, it is necessary to ensure integration of environmental health and socio-economic feasibility in order to have better production of food crops (Yadav et al. 2017; Verma et al. 2018). Plant growth promoting rhizobacteria (PGPR) are the potential tools that have been extensively used to meet this need worldwide (Ahemad and Kibret 2014). Soil is considered dynamically active and highly diverse habitat for microorganisms. Further in soil, rhizosphere is the most vital location vigorously involved in both organic and inorganic nutritional biogeochemical cycling affecting the growth of plants (McNear 2013). This region is the key point for isolation of PGPR that possess various properties directly and indirectly related to growth and yield of plants (Sharma et al. 2003; Sharma 2003). The beneficial association occurring between plant and microbes in rhizosphere determines the health of plant and fertility of soil (Jeffries et al. 2003; Vishwakarma et al. 2017a, b). Several studies have reported substantial enhancement in growth of agriculturally important crops when inoculated with PGPR (Shukla et al. 2012; Das et al. 2013). They can become effective alternatives to chemical based fertilizers, pesticides or genetically modified plants (Rajkumar et al. 2010; Vilchez et al. 2016).
PGPR’s colonization and diversity depends on the type of plant and physicochemical properties of soil and even vary with season, crop age and agriculture practices employed. The mechanism of their action involves phytostimulation (promoting plant growth by producing phytohormones), biofertilization (making nutrients available to plants) and biocontrol (limiting pathogens and diseases by producing antibiotics and inducing defense system of plants) (Ahemad and Kibret 2014). Plant growth promoting potential of any bacterial species can be examined by numerous tests such as phosphate solubilization, indole-3-acetic acid production, free nitrogen assimilation, siderophore production, hydrogen cyanide production and biocontrol (Idris et al. 2007; Zaidi et al. 2009; Ahemad and Kibret 2014; Shameer and Prasad 2018). They also stimulate the chemotactic response by showing attraction towards chemical compounds present in root exudates and facilitate root colonization (Podile et al. 2013). Numerous studies have reported that PGPR specifically colonize the host plant roots thereby indicating plant and microbe coexistence (Shukla et al. 2012; Santoyo et al. 2016). Their effective application in agriculture can prove to reduce the use of harmful chemicals, agricultural cost, as well as minimize soil degradation. Further, their ability to tolerate a broad range of pollutants make them a suitable candidate that can be employed as bioinoculant along with appropriate carrier material in order to increase the shelf-life of bioformulation (Tripti et al. 2017). Several agricultural waste materials can be employed as carrier support; for example, wheat bran, charcoal, saw dust, rice husk, rice bran etc. due to their organic behavior and vermiculite, perlite, silicates etc. due to their inert characteristics as they ensure the maximum viability of bacterial cells (Malusá et al. 2012; Sangeetha 2012).
Brassica juncea, also known as Indian Mustard, is considered to be an essential oilseed crop that is majorly grown in North India and utilized as a common ingredient in every Indian household as mustard oil. The mustard straw is being consumed in brick kiln as fuel source. It also has multiple characteristics such as phytoremediation of soil contaminated with heavy metals such as cadmium (Belimov et al. 2005) and arsenic (Srivastava et al. 2013). Number of PGPR have been identified in the rhizosphere of B. juncea that are capable of increasing plant growth and suppressing pathogens in surrounding area. These include Pseudomonas spp., Rhodococcus sp. (Belimov et al. 2001), Variovorax paradoxus, Flavobacterium sp. (Belimov et al. 2005), Staphylococcus arlettae (Srivastava et al. 2013), Bacillus cereus (Anwar et al. 2014), Achromobacter sp., Streptococcus sp., Pseudomonas sp. and Bacillus sp. (Sinha and Jee 2018).
With this background, the current study was envisaged to isolate bacteria from rhizosphere of B. juncea grown in Allahabad and characterize them for plant growth promoting potential. Further, waste carriers such as charcoal, sawdust, wheat bran and rice husk were checked for their ability to support survival of PGPR for long time so as to develop multipurpose bioinoculant.
Materials and methods
Site description
Allahabad city in Uttar Pradesh, India was selected for this study. The regions taken under Allahabad include Narayani Ashram, Teliyarganj (Site 1); Malak Herher, Bhadri (Site 2); and Hathga, Champatpur (Site 3) located around 15 km from each other. This region experiences humid to sub humid monsoonal climate and receives moderate to high rainfall ranges between 1000 and 1500 mm. They are major producing areas of Indian mustard (B. juncea) which is an oilseed crop widely used as vegetable and oil in Indian foods. B. juncea is a Rabi crop grown from November to March each year. Global Positioning System (GPS) was used to locate the sites. The details of three locations are mentioned in Table 1.
Collection of soil samples
The soil samples were taken from the established fields of B. juncea. Soil attached to the plant roots (i.e. rhizospheric soil) was collected from the aforementioned regions. Random selection of three locations were made from each site and collection of samples was carried out using coring method (Dinesh et al. 2015; Lamizadeh et al. 2016). With this, there were three replicates (triplicates) from every site. Opaque low density polyethylene (having thickness of about 75 micron) bags were used to place soil immediately after sampling. These samples were taken to laboratory, dried (at 45 °C) in hot air oven and sieved (through < 2 mm mesh) for further processing. The soil samples were tested for available phosphorous (P), potassium (K), organic carbon (OC), pH and electrical conductivity (EC) at Krishi Vigyan Kendra, Naini Allahabad.
Isolation and purification of microorganisms
Serial dilution procedure was utilized to isolate the culturable bacterial population in rhizospheric soil. For serial dilution, 1 g soil was taken in a test tube and mixed with 9 ml de-ionized water followed by vortexing. Appropriate dilutions were made thereafter (up to 10−5) (Johnson and Curl 1972). 100 µl sample from each dilution was spread plated on Nutrient Agar (HiMedia®) petri plates and incubated at 30 °C for 24 h. Colonies were further assessed based on color, texture, shape etc. for occurrence percentage and colony forming unit (CFU) calculation.
Further, purification of isolates was carried out by streaking the loop full of culture over nutrient agar plates and well-isolated colony was re-streaked to obtain single colony type culture (Zhou 1987).
Morphological and biochemical characterization
Isolated cultures were examined for morphological characteristics via Gram staining, endospore and capsule staining (McClelland 2001). The biochemical tests including sugar utilization, amino acids and starch hydrolysis were performed according to Bergey’s Manual to Determinative Bacteriology (Brown 1939).
Plant growth-promoting attributes
The cultures were further screened in vitro to examine their plant growth promoting potential. These included phosphate (P) solubilization, indole-3-acetic acid (IAA) production, siderophore production, free nitrogen assimilation and hydrogen cyanide (HCN) production.
Phosphate solubilization
The isolates were examined for their capacity to solubilize tricalcium phosphate (TCP) as per the method of Wahyudi et al. (2011). Briefly, spot inoculation of cultures was done on plates of Pikovskaya Agar (HiMedia®) followed by incubation at 30 °C for 5 days. Clear halo zone on the plates was observed for P-solubilization activity of isolates.
Phosphate solubilization was quantitatively estimated for efficient isolates in 250 ml Erlenmeyer flask having 100 ml Pikovskaya broth with TCP (Wahyudi et al. 2011). To each flask, 0.5 g of TCP was added separately as insoluble phosphate and the contents were sterilized. The initial pH of the medium was 6.0. The flasks were cooled and inoculated with selected isolates followed by incubation at 27 °C for 10 days. The growth medium was taken out from each flask daily in duplicates and subjected to filtration through Whatman number 42 filter papers. The chlorostanous reduced molybdo-phosphoric acid blue methodology was employed to test the filtrates for P2O5 content. The absorbance was taken at 700 nm by using double beam UV–Vis spectrophotometer (Eppendorf BioSpectrometer).
IAA production
IAA production was estimated by Salkowski reagent according to the methodology described by Loper et al. (1982). In this method, 50 ml nutrient broth was supplemented with 0.1% tryptophan and 500 µl freshly grown bacterial culture was added to it followed by incubation at 30 °C for 48 h in dark under shaking conditions. Un-inoculated nutrient broth (NB) tube with tryptophan was kept as control. The bacterial cultures were harvested by centrifugation and estimation of IAA was facilitated by Salkowski’s reagent (perchloric acid and 0.5 M FeCl3 solution) using colorimetric assay. Appearance of pink color indicated production of IAA in test tubes.
Quantitative measurement of IAA was carried out colorimetrically using Salkowski reagent according to the methodology described by Gordon and Weber (1951).
Siderophore production
The glasswares used for siderophore detection were firstly dipped in 20% HCl to ensure the absence of any iron followed by rinsing with double distilled water. Isolates were tested for siderophore production by spot inoculation on blue agar plate in the presence of chrome-azurol sulfonate (CAS) dye according to the procedure given by Schwyn and Neilands (1987). After inoculation, the plates were allowed to incubate at 30 °C for 10–15 days. The formation of yellowish orange zones around the bacterial colonies indicated the production of siderophore.
CAS-shuttle assay was utilized for estimating siderophores quantitatively (Payne 1994). CAS reagent was added to the 0.5 ml of the culture supernatant and its optical density was recorded at 630 nm. Uninoculated broth along with CAS reagent was taken as reference. Siderophore content was estimated by using the following formula:
where Ar = optical density of reference and As = optical density of the sample
Chemical nature of siderophores was analyzed by preparing ferric siderophore complex with the addition of 3 ml of 2% ferric chloride to 1 ml culture supernatant. Absorbance spectra was scanned from 200 to 1100 nm range using UV/Visible Spectrophotometer (UV-2450, Shimadzu) taking uninoculated broth as blank.
HCN production
All the isolates were tested for hydrogen cyanide gas production by adapting the methodology of Castric (1975). Nutrient agar medium was prepared and 4.4 g/L glycine was amended to it followed by streaking of 24 h old cultures to the plates. Further, Whatman paper was soaked in 0.5% picric acid solution made in 2% Na2CO3 and placed on the upper lid of the petri dishes. Control plate is the one without streaking of culture. Parafilm was used to seal all the plates and kept at 28 °C for 5–6 days for incubation. Positive HCN production was observed with the development of brown color on filter paper.
Free nitrogen assimilation
Free nitrogen assimilation by bacterial isolates were assessed by the method described by Jensen (1955). In this, Jensen’s medium was prepared and poured into the petri plates. All the isolates were streaked on the plates and incubated for 24 h at 30 °C. The isolates were then observed for their growth on streaked plates.
Molecular characterization
The isolates were further identified by molecular characterization using 16SrRNA sequencing. Isolation of DNA from bacterial samples was performed by phenol–chloroform method according to the protocol of Mamiatis et al. (1985). Quality of DNA was tested qualitatively on 1% agarose gel electrophoresis. After visualization of band on gel, DNA was amplified with the help of primers specific to 16s rRNA amplification i.e. 8F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′ AAGGAGGTGATCCAGCCGCA-3′) using Prima-96™ (HiMedia®). Once the sequence was obtained, it was compared to the similar sequences at GenBank database using Basic Local Alignment Search Tool (BLAST) at NCBI database. Further, neighbor-joining tree was constructed to analyze the evolutionary history of the isolates using MEGA7 (Saitou and Nei 1987; Tamura et al. 2004). The bootstrap consensus tree inferred from 500 replicates is taken to represent the evolutionary history of the taxa analyzed (Felsenstein 1985). The sequences so obtained were submitted to GenBank Database NCBI to obtain accession numbers.
Carrier studies
Four different carrier materials viz. charcoal, sawdust (from salwood i.e. Shorea robusta), wheat bran and rice husk were obtained from local supplier in Allahabad and were utilized in the study. The carrier materials were subjected to grinding followed by drying. The physico-chemical properties such as inherent moisture content, water holding capacity and pH of carrier materials were estimated as per the methodologies described by Aeron et al. (2011). 40 g of these materials were weighed and packed in thick, low-density polythene bags. The bags were sealed after leaving two-third vacant space for proper circulation of air. These bags with carrier materials were then double autoclaved.
The three potential isolates were cultured in nutrient broth at 150 rpm for 24 h. Cells at 0.1 optical density (108 cells/ml at 595 nm) were taken for inoculation. Cultures were used in combination (KVS20 + KVS25, KVS20 + KVS27, KVS25 + KVS27 and KVS20 + KVS25 + KVS27) in equal amounts. Aseptic inoculation of 1 ml of bacterial suspension (having 108 cells/ml) was carried out using hollow needle into the bags and the holes were immediately shielded with the adhesive tape. The content in bag was mixed properly and stored at room temperature in dark. Each treatment was placed in three replicates.
To study the survival of bacterial isolates in carrier material, samples from bags were collected at different time intervals under sterile conditions for up to 150 days. The samples were serially diluted and plated on nutrient agar plates to determine the number of bacterial cells by viable count method.
Pot culture experiment for testing bacterial isolates for plant growth promotion
For pot culture study, B. juncea seeds were surface sterilized using 1% sodium hypochlorite solution followed by washing with deionized water. Next, seeds were treated with bacterial suspension (as prepared in Sect. 2.7) by soaking seeds in suspension for 30 min (Vidhyasekaran et al. 1997). Soil collected previously was autoclaved at 15 psi and 121 °C for 15 min twice. For experimentation, pots were also surface sterilized with 70% ethanol and filled with sterilized soils. Thereafter, various treatments (control, KVS20, KVS25, KVS27, KVS20 + KVS25, KVS20 + KVS27, KVS25 + KVS27 and KVS20 + KVS25 + KVS27) were formulated in triplicates and kept in plant growth chamber with 12 h/12 h light/dark photoperiod cycle at 16–25 °C for 30 days. The bacterized seeds were then sowed in each pot according to the treatment. The various treatments were T1-control; T2-KVS20; T3-KVS25; T4-KVS27; T5-KVS20 + KVS25; T6-KVS25 + KVS27; T7-KVS20 + KVS27; T8-KVS20 + KVS25 + KVS27.
Results
Properties of soil sample and isolation of bacteria
The locations selected for conducting the study were spotted on map and with the help of global positioning system (GPS), their coordinates were noted (Table 1). Also, the properties of soil samples collected from three different locations are summarized in Table 1.
Serial dilution method was used to isolate bacteria from soil samples and an average of 16 × 106 CFU/g soil sample was obtained. They were further screened on the basis of their appearance such as shape, texture, color, size etc. and occurrence percentage was calculated as shown in Fig. 1. Isolates having abundance more than 5% were taken for further studies. The calculation resulted into selection of eight isolates from Site 1, nine isolates from Site 2 and three from Site 3. Purified colonies of these 20 isolates were obtained by streaking (Fig. 2a). The isolates selected on the basis of abundance were KVS1, KVS2, KVS5, KVS6, KVS7, KVS9, KVS12, KVS13, KVS16, KVS17, KVS18, KVS19, KVS20, KVS22, KVS23, KVS24, KVS25, KVS27, KVS28 and KVS29.

Graph depicting the occurrence % of bacterial colonies in soil sample from a Site 1 (Narayani Ashram), b Site 2 (Bhadri) and c Site 3 (Hathga)

a Isolates showing halo zones on Pikovskaya plates, b Quantitative estimation of phosphate concentration on each sampling day, and c Scatter graph indicating the change in pH of samples over a span of 10 days
Morphological characteristics
Isolates were subjected to different staining procedures and the results are summarized in Table S1. It was observed that out of 20 isolates, 50% were Gram-positive and rest 50% were Gram-negative (Fig S1). Only two isolates showed endospore formation whereas none of them was positive for capsule formation.
Plant growth-promoting characteristics
Phosphate solubilization
Phosphate solubilization was exhibited by only two isolates (KVS25 and KVS 27) out of 20 (Fig. 2a). When spot inoculated on Pikovskaya plates, zone of clearance was observed to be 11 and 13 mm for KVS25 and KVS27, respectively. The amount of phosphate solubilized was estimated quantitatively by supplementing 100 ml Pikovskaya broth with 0.5 g tricalcium phosphate and inoculation with these two isolates. It was observed that KVS27 solubilized 57.6 mg/l phosphate after 8 days whereas KVS 25 showed maximum of 51.09 mg/l concentration of phosphate at day 6th as compared to control (Fig. 2b). Simultaneously, the pH of the medium was also analyzed and it was shown to decrease from pH 5 (control and initial pH) to pH 3.6 for KVS25 and 3.8 for KVS27 (Fig. 2c).
IAA production
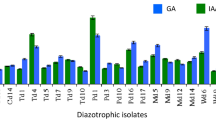
A total of 11 isolates out of 20 showed the production of phytohormone IAA. The range of IAA produced by these isolates was between 0.027 mg/ml and 3.17 mg/ml leading to the average IAA production of 0.63 mg/ml. Out of 11 isolates, KVS29 was observed to be the highest producer of IAA (3.177 mg/ml) followed by KVS23, KVS27, KVS25 and KVS9 (Fig. 3).

Bar graph representing the concentration of IAA produced by various isolates. The standard equation used for calculating the IAA concentration is, y = 0.1827x + 0.075 (R2 = 0.9952)
Siderophore production
Siderophore production was found positive for only two isolates viz. KVS18 and KVS20. Orange colored zone was prominently seen around these isolates. Further, siderophore units were also calculated for KVS18 and KVS20 and it was observed to be 5% and 14% respectively. The absorbance spectrum was at 404 nm for KVS18 and 406 nm for KVS20 indicating their chemical nature as hydroxamate (Fig S2a, b).
HCN production and free nitrogen assimilation
Three isolates have shown the production of HCN which was indicated by the change in color of filter paper in plates from yellow to brown (Fig S2c). Free nitrogen was assimilated by total 14 isolates out of 20 that showed the growth on streaked Jensen’s plates (Fig S2d).
On the basis of PGP characteristics displayed by different isolates (Table 2), three potential isolates namely KVS20, KVS25 and KVS27 were selected.
Biochemical characterization
The three isolates selected after screening for their PGP attributes were biochemically characterized for their ability to utilize various sugars, amino acids, salts, organic acids and dyes. The results for substrate utilization are presented in Table 3.
Molecular characterization
Based on 16s rRNA sequencing, the isolates KVS20 showed maximum similarity with Pseudomonas sp., while KVS25 and KVS27 showed maximum similarity with Bacillus sp. The neighbor joining phylogenetic tree was constructed using 16S rRNA gene sequence with other sequences of similar species and it was revealed that the isolates KVS20, KVS25 and KVS27 form a close cluster with strains Pseudomonas sp. VET-5 (Fig. 4a), Bacillus thuringiensis (Fig. 4b) and Bacillus paramycoids, (Fig. 4c) respectively. The accession number of submitted sequences for species Pseudomonas sp. KVS20, Bacillus thuringiensis KVS25 and Bacillus paramycoids KVS27 are MH780493, MH780494 and MH780495 respectively.


Phylogenetic tree based on 16S ribosomal RNA gene sequences showing relationships between a KVS20 and other Pseudomonas sp., b KVS25 and other Bacillus sp., and c KVS27 and other Bacillus sp.
Viability of potential isolates in carrier materials
The characteristics of four different carrier materials are listed in Table 4. Viability studies of bacterial cultures in combinations on different carrier materials i.e. charcoal, sawdust, wheat bran and rice husk was analyzed by plating the cultures on nutrient agar plates after serial dilution and calculating the population dynamics at interval of 30 days for about 150 days. It was found that the two combinations (Pseudomonas sp. KVS20 + B. thuringiensis KVS25 and Pseudomonas sp. KVS20 + B. thuringiensis KVS25 + B. paramycoides KVS27) showed higher bacterial count in wheat bran carrier (Table 5a, d) whereas the other two (Pseudomonas sp. KVS20 + B. paramycoides KVS27 and B. thuringiensis KVS25 + B. paramycoides KVS27) showed high count in sawdust (Table 5b, c). From initial day of study, bacterial count consistently increased till 120th day followed by a drop till 150th day in all the cases.
Pot culture study for analysis of plant growth promotion
The plants were harvested after 30 days for analysis of growth parameters such as root and shoot length and fresh mass as well as soil properties including available phosphate, potassium and organic carbon. The results of changes in aforesaid parameters are listed in Table 6. It was observed that treatment T5 was most effective as it increased the shoot length by 28.7%, root length by 36.1% and fresh weight by 47.3% as compared to control. However, the phosphate content in soil was increased greatly by T8. The trend of organic carbon as well as potassium was found enhanced in T5 treatment.
Discussion
PGPR colonize the roots of plant and exhibit beneficial impact on growth and development of plants by diverse mode of actions. The mechanisms by which growth of plants is enhanced by PGPRs are not precisely recognized, however, involvement of several methods including phosphate solubilization, phytohormone production, solubilization of nutrients, and suppression of various pathogens in soil have been observed (Glick 1995; Bowen and Rovira 1999; Shukla et al. 2012; Singh and Lal 2016).
The current study was conducted in order to identify the rhizobacterial isolates on the basis of morphological, biochemical properties from the rhizosphere of B. juncea and further screened for their ability to enhance growth of plant by assessing various traits like production of phytohormone IAA, making nutrients available to plants and production of hydrogen cyanide to suppress pathogens. These isolates were subjected to in vitro screening for aforementioned characteristics.
Total 20 isolates were screened in the study based on their occurrence percentage and abundance in the mustard rhizosphere. The selected three regions come under the major mustard producing areas in Allahabad. Bacterial isolates were microscopically assessed for Gram staining, endospore and capsule staining which showed that ten isolates were Gram-positive and remaining ten were Gram-negative. All the isolates were rod shaped. Further, isolates KVS25 and KVS29 showed positive result for endospore formation. After the morphological examination of isolates, they were examined for plant growth promotion activity by conducting various tests like phosphate solubilization, IAA production, siderophore production, free nitrogen assimilation and HCN production.
Phosphate solubilizing bacteria present in the rhizosphere secrete organic acids and phosphatase enzyme that convert the insoluble complex of phosphorous to the form that is easily available to plants (Kim et al. 1997; Pradhan and Sukla 2006). Only two isolates, namely KVS25 and KVS27, solubilized phosphate in our study. It can be inferred that a greater portion of insoluble P in soil that cannot be utilized directly by the plants is made available to the plants by these isolates. Our study is in accordance with the study carried out by Sinha and Jee (2018) that B. juncea rhizosphere inhabits the phosphate solubilizing bacteria Bacillus sp. These isolates were also found to produce IAA. IAA is an auxin phytohormone that initiates development of roots, facilitates cell division and enlargement (Salisbury 1994). PGPR are very commonly found to produce this phytohormone (Barazani and Friedman 1999). Several researchers have reported that indigenous soil microbiota produces a range of phytohormones like auxins (IAA), cytokines and gibberellins (Ahemad and Khan 2012). In this study, total 11 isolates were observed to produce IAA; however, the isolate KVS29 produced highest amount of IAA i.e. 3.177 mg/ml. It indicates that KVS29 may have the capacity to enhance the root growth and root length resulting in higher surface area to enable more access of soil nutrients to plants. The genera of PGPR that exhibit this property include Pseudomonas, Bacillus, Rhizobium, Alcaligenes (Ahmad et al. 2008) and they may vary depending upon species and growth stages of plant.
Iron is made available to plants by siderophores that are very essential as they act as iron chelator and also compete for iron with other microorganisms in the rhizosphere. The most commonly utilized form of iron by plants is ferrous ion (Fe2+); however, the well-aerated soil environment is dominated with ferric ion (Fe3+) that is further oxidized to form iron-oxides (Salisbury and Ross 1992). Isolates KVS18 and KVS20 exhibited production of siderophores, out of which more siderophore units were produced by KVS20. In a study by Siddiqui et al. (2003), isolates of Pseudomonas and Flavobacterium were found to be efficient siderophore producers. The various categories in which siderophores fall are hydroxamates (400–450 nm), catecholates (320 nm, 495 nm) and carboxylates (190–280 nm) (Neilands 1974; Carson et al. 2000). In this work, both the isolates were categorized as producers of hydroxamates (KVS18-404 nm and KVS20-406 nm). Furthermore, the ability for symbiotic nitrogen fixation was also tested and majority of isolates showed ability of symbiotic nitrogen fixation. Rhizobia are the most researched and exploited PGPR which includes various genera like Azorhizobium, Bradyrhizobium, Allorhizobium, Mesorhizobium, Sinorhizobium, and Rhizobium and found active in nitrogen fixation in leguminous plants. In addition, PGPR are also involved in eradicating the deleterious effects of variety of plant pathogens indirectly by producing inhibitory compounds (Nehl et al. 1996; Persello-Cartieaux et al. 2003). One such mechanism is production of hydrogen cyanide for biological control of pathogens. HCN production was detected to be highest by the isolate KVS27 followed by isolates KVS25 and KVS20. Role of HCN in disease suppression was earlier studied and Pseudomonas species are well known in suppressing phytopathogens by production of HCN (Haas et al. 1991; Geetha et al. 2014; Modi et al. 2017).
When the isolates were screened in vitro for plant growth promoting potential, three isolates possessed majority of PGP traits and hence, they were further tested and identified. Since, morphological and biochemical characterization cannot guarantee the identification of isolates, therefore, 16s rRNA sequencing is necessary for clear identification. It was observed that KVS20 belonged to Pseudomonas sp. and KVS25 and KVS27 belonged to Bacillus sp. When the sequences were aligned and assessed for evolutionary history using MEGA7 software, the strains were confirmed for the three isolates along with their accession numbers namely Pseudomonas sp. KVS20 (MH780493), Bacillus thuringiensis KVS25 (MH780494) and Bacillus paramycoides. KVS27 (MH780495). The sequencing results are in support to the previously obtained results for biochemical and morphological tests and that also show the characteristics of plant growth promotion (Raddadi et al. 2008; Ahmad et al. 2008).
Number of studies were carried out to assess the viability of bacteria in carrier materials. Wheat bran and rice husk are the lignocellulosic agriculture by product of wheat and rice crops which are also utilized as animal feed. While other carriers like saw dust and charcoal are cost effective and easily available in the market. In the present study, carrier based bioformulation of potential PGP isolates (in consortium) were prepared in different carrier materials i.e. saw dust, wheat bran, rice husk and charcoal and studied for 150 days of storage. Similarly, researchers have studied the survival of efficient PGPR isolates in different solid based carriers for 6 months and determined population dynamics through viable count method (Sivasakthivelan and Saranraj 2013; Sangeetha 2012). The viability of promising treatments in different carrier based formulations is presented in Table 4. The results clearly indicate that the highest survival of consortia is observed in sawdust followed by wheat bran, charcoal and rice husk. The combinations of bacterial isolates were further tested for their plant growth promotion efficacy in pot experiments and it was noted that T5 treatment i.e. KVS20 and KVS25 that belong to species Pseudomonas and Bacillus respectively, enhanced the growth of mustard plant as compared to control. The result indicated the coexistence of both bacteria and their synergistic effect towards plant. The result is in accordance with the previous studies showing the role of Pseudomonas sp. in plant growth promotion and disease suppression (Aeron et al. 2011) as well as Bacillus sp in alleviating the heavy metal stress in coalmine regions (Upadhyay et al. 2017). Such researches indicate the intricate role of bacterial species in plant growth promotion along with stress amelioration.
Conclusion
From rhizospheric soil samples of Indian mustard (B. juncea), bacteria were isolated that showed different PGP activities such as IAA production, phosphate solubilization, siderophore production, symbiotic N2 fixation, and HCN production. Based on these tests, three potential isolates were selected and after 16s rRNA sequencing, they were identified as Pseudomonas sp. KVS20, Bacillus thuringiensis KVS25 and Bacillus paramycoides. KVS27 that are previously known for their plant growth promotion activities. They were further formulated as consortia on different carriers in which wheat bran and sawdust showed higher viability of isolates. Also, Pseudomonas sp and B. thuringiensis enhanced the growth of mustard plant in present study. Hence, their formulation as consortia will help in development and application as effective biofertilizer to overcome the cost and hazards associated with chemical fertilizer. This will lead to improvement in health of soil and maintenance of soil fertility for sustainable agriculture.
References
Aeron A, Dubey RC, Maheshwari DK, Pandey P, Bajpai VK, Kang SC (2011) Multifarious activity of bioformulated Pseudomonas fluorescens PS1 and biocontrol of Sclerotinia sclerotiorum in Indian rapeseed (Brassica campestris L.). Eur J Plant Pathol 131:81–93
Ahemad M, Khan MS (2012) Effect of fungicides on plant growth promoting activities of phosphate solubilizing Pseudomonas putida isolated from mustard (Brassica campestris) rhizosphere. Chemosphere 86:945–950
Ahemad M, Kibret M (2014) Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J King Saud Univ Sci 26:1–20
Ahmad F, Ahmad I, Khan M (2008) Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol Res 163:173–181
Anwar MS, Siddique MT, Verma A, Rao YR, Nailwal T, Ansari M, Pande V (2014) Multitrait plant growth promoting (PGP) rhizobacterial isolates from Brassica juncea rhizosphere: keratin degradation and growth promotion. Commun Integr Biol 7(1):e27683
Barazani O, Friedman J (1999) Is IAA the major root growth factor secreted from plant-growth-mediating bacteria? J Chem Ecol 25:2397–2406
Belimov AA, Safronova VI, Sergeyeva TA, Egorova TN, Matveyeva VA, Tsyganov VE, Borisov AY, Tikhonovich IA, Kluge C, Preisfeld A, Dietz KJ (2001) Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can J Microbiol 47(7):642–652
Belimov AA, Hontzeas N, Safronova VI, Demchinskaya SV, Piluzza G, Bullitta S, Glick BR (2005) Cadmium-tolerant plant growth-promoting bacteria associated with the roots of Indian mustard (Brassica juncea L. Czern.). Soil Biol Biochem 37(2):241–250
Bowen GD, Rovira AD (1999) The rhizosphere and its management to improve plant growth. Adv Agron 66:1–102
Brown JH (1939) Bergey’s manual of determinative bacteriology. Am J Public Health Nations Health 29(4):404–405
Carson KC, Meyer JM, Dilworth MJ (2000) Hydroxamate siderophore of root nodule bacteria. Soil Biol Biochem 32:11–21
Castric PA (1975) Hydrogen cyanide, a secondary metabolite of Pseudomonas aeruginosa. Can J Microbiol 21:613–618
Das AJ, Kumar M, Kumar R (2013) Plant growth promoting rhizobacteria (PGPR): an alternative of chemical fertilizer for sustainable environment friendly agriculture. Res J Agric For Sci 1:21–23
Dinesh R, Anandaraj M, Kumar A, Bini YK, Subila KP, Aravind R (2015) Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol Res 173:34–43
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Freitas ADS, Vieira CL, Santos CERS, Stamford NP, Lyra MCCP (2007) Caracterizacao de rizobios isolados de Jacatupe cultivado em solo salino no Estado de Pernanbuco. Brasil Bragantia 66:497–504
Geetha K, Venkatesham E, Hindumathi A, Bhadraiah B (2014) Isolation, screening and characterization of plant growth promoting bacteria and their effect on Vigna Radiata (L.) R. Wilczek. Int J Curr Microbiol Appl Sci 3:799–809
Glick B (1995) The enhancement of plant growth by free living bacteria. Microbiol 41:109–117
Gordon SA, Weber RP (1951) Colorimetric estimation of indole acetic acid. Plant Physiol 26:192–195
Haas D, Keel C, Laville J, Maurhofer M, Oberhansli T, Schnider U, Voisard C, Wuthrich B, Defago G (1991) Secondary metabolites of Pseudomonas fluorescens strain CHAO involved in the suppression of root diseases. In: Hennecke H, Verma DPS (eds) Advances in molecular genetics of plant–microbe interactions, vol 11. Kluwer Academic Publishers, Dordrecht, pp 450–456
Idris EE, Iglesias DJ, Talon M, Borriss R (2007) Tryptophan-dependent production of indole-3-acetic acid (IAA) affects level of plant growth promotion by Bacillus amyloliquefaciens FZB42. Mol Plant Microbe Interact 20(6):619–626
Jeffries S, Gianinazzi S, Perotto S, Turnau K, Barea JM (2003) The contribution of Arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol Fertil Soils 37:1–16
Jensen HL (1955) Non-symbiotic nitrogen fixation. In: Barthdlomew WV, Clark FE (eds) Soil nitrogen, vol 10. American Soc Agron Inc Madison, pp 436–480
Johnson LF, Curl EA (1972) Methods for research on the ecology of soil-borne plant pathogens. In: Methods for research on the ecology of soil-borne plant pathogens
Kim KY, Jordan D, Krishnan HB (1997) Rahnella aquatilis, bacterium isolated from soybean rhizosphere, can solubilize hydroxyapatite. FEMS Microbiol Lett 153:273–277
Lamizadeh E, Enayatizamir N, Motamedi H (2016) Isolation and identification of plant growth-promoting rhizobacteria (PGPR) from the rhizosphere of sugarcane in saline and non-saline soil. Int J Curr Microbiol App Sci 5(10):1072–1083
Loper JE, Schroth MN, Panopoulos N (1982) Influence of bacterial sources of indole-3-acetic-acid (IAA) on root elongation of sugar-beet. Phytopathol 72:997
Malusá E, Sas-Paszt L, Ciesielska J (2012) Technologies for beneficial microorganisms inocula used as biofertilizers. Sci World J 2012:491206
Mamiatis T, Fritsch EF, Sambrook J, Engel J (1985) Molecular cloning—a laboratory manual. Cold Spring Harbor Laboratory 1982, New York, p 545
McClelland R (2001) Gram’s stain: the key to microbiology. MLO 33(4):20–22
McNear DH Jr (2013) The rhizosphere—roots, soil and everything in between. Nat Educ Knowl 4(3):1
Modi K, Patel P, Parmar K (2017) Isolation, screening and characterization of PGPR from rhizosphere of rice. Int J Pure Appl Biosci 5:264–270
Nehl DB, Allen SJ, Brown JF (1996) Deleterious rhizosphere bacteria: an integrating perspective. Appl Soil Ecol 5:1–20
Neilands JB (1974) Microbial iron metabolism. Academic Press, Inc., USA, p 26723
Payne SM (1994) Detection, isolation, and characterization of siderophores. Methods Enzymol 235:329
Persello-Cartieaux F, Nussaume L, Robaglia C (2003) Tales from the underground: molecular plant-rhizobacteria interactions. Plant Cell Environ 26:189–199
Podile AR, Vukanti RV, Sravani A, Kalam S, Dutta S, Durgeshwar P, Rao VP (2013) Root colonization and quorum sensing are the driving forces of plant growth promoting rhizobacteria (PGPR) for growth promotion. Proc Natl Acad Sci India Sect B 80:407–413
Pradhan N, Sukla LB (2006) Solubilization of inorganic phosphates by fungi isolated from agriculture soil. Afr J Biotechnol 5:850–854
Raddadi N, Cherif A, Boudabous A, Daffonchio D (2008) Screening of plant growth promoting traits of Bacillus thuringiensis. Ann Microbiol 58(1):47–52
Rajkumar M, Ae N, Prasad MNV, Freitas H (2010) Potential of siderophore producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol 28:142–149
Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Salisbury FB (1994) Role of plant hormones. In: Wilkinson RE (ed) Plant environment interactions. Marcel Dekker, New York, pp 39–81
Salisbury FB, Ross CW (1992) Plant physiology. Wadsworth Pub. Co. Belmont, p 682
Sangeetha D (2012) Survival of plant growth promoting inoculants in different carriers. Int J Pharm Biol Arch 3:231–239
Santoyo G, Moreno-Hagelsieb G, del Carmen Orozco-Mosqueda M, Glick BR (2016) Plant growth-promoting bacterial endophytes. Microbiol Res 183:92–99
Schwyn B, Neilands JB (1987) Universal chemical assay for the detection and determination of siderophore. Anal Biochem 160:47–56
Shameer S, Prasad TNVKV (2018) Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant Growth Regul 84(3):1–13
Sharma S (2003) Response of various isolates of Bradyrhizobium inoculation on protein content and its yield attributes of green gram [Vigna radiata (L.) Wilczek]. Legume Res Int J 26:28–31
Sharma S, Upadhyay RG, Sharma CR (2003) Response of various levels of nitrogen and phosphorus application on growth, physiological parameters and yield of Vigna radiata (L.) wilczek under rainfed and mid-hill conditions of Himachal Pradesh. Indian J Agric Res 37:52–55
Shukla KP, Sharma S, Singh NK, Singh V (2012) Deciphering rhizosphere soil system for strains having plant growth promoting and bioremediation traits. Agric Res 1:251–257
Siddiqui IA, Shaukat SS, Khan GH, Ali NI (2003) Suppression of Meloidogyne javanica by Pseudomonas aeruginosa IE-6S + in tomato: the influence of NaCl, oxygen and iron level. Soil Biol Biochem 35:1625–1634
Singh Y, Lal N (2016) Isolation and characterization of PGPR from wheat (Triticum aestivum) rhizosphere and their plant growth promoting traits in vitro. Indian J Biol 3:139–144
Sinha T, Jee C (2018) Isolation and characterization of Rhizobacteria from Brassica juncea L. plant. IJCRT 6(1):1200–1207
Sivasakthivelan P, Saranraj P (2013) Azospirillum and its formulations: a review. Int J Microbiol Res 4:275–287
Srivastava S, Verma PC, Chaudhry V, Singh N, Abhilash PC, Kumar KV, Sharma N, Singh N (2013) Influence of inoculation of arsenic-resistant Staphylococcus arlettae on growth and arsenic uptake in Brassica juncea (L.) Czern. Var. R-46. J Hazard Mater 262:1039–1047
Tamura K, Nei M, Kumar S (2004) Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci USA 101:11030–11035
Tripti Kumar A, Usmani Z, Kumar V (2017) Biochar and flyash inoculated with plant growth promoting rhizobacteria act as potential biofertilizer for luxuriant growth and yield of tomato plant. J Environ Manage 190:20–27
Upadhyay N, Vishwakarma K, Singh J, Mishra M, Kumar V, Rani R, Mishra RK, Chauhan DK, Tripathi DK, Sharma S (2017) Tolerance and reduction of chromium (VI) by Bacillus sp. MNU16 isolated from contaminated coal mining soil. Front Plant Sci 8:778
Verma RK, Sachan M, Vishwakarma K, Upadhyay N, Mishra RK, Tripathi DK, Sharma S (2018) Role of PGPR in sustainable agriculture: molecular approach toward disease suppression and growth promotion. In: Meena VS (ed) Role of rhizospheric microbes in soil. Springer, Singapore, pp 259–290
Vidhyasekaran P, Rabindran R, Muthamilan M, Nayar K, Rajappan K, Subramanian N, Vasumathi K (1997) Development of a powder formulation of Pseudomonas fluorescens for control of rice blast. Plant Pathol 46:291–297
Vilchez JI, Navas A, Gonzalez-Lopez J, Arcos SC, Manzanera M (2016) Biosafety test for plant growth-promoting bacteria: proposed environmental and human safety index (EHSI). Protoc Front Microbiol 6:1514
Vishwakarma K, Sharma S, Kumar N, Upadhyay N, Devi S, Tiwari A (2016) Contribution of microbial inoculants to soil carbon sequestration and sustainable agriculture. In: Singh DP, Singh HB, Ratna P (eds) Microbial inoculants in sustainable agricultural productivity, vol 2. Springer, New Delhi, pp 101–113
Vishwakarma K, Mishra M, Jain S, Singh J, Upadhyay N, Verma RK, Verma P, Tripathi DK, Kumar V, Mishra R, Sharma S (2017a) Exploring the role of plant-microbe interactions in improving soil structure and function through root exudation: a key to sustainable agriculture. In: Singh DP, Singh HB, Ratna P (eds) Plant–microbe interactions in agro-ecological perspectives, vol 1. Springer, Singapore, pp 467–487
Vishwakarma K, Sharma S, Kumar V, Upadhyay N, Kumar N, Mishra R, Yadav G, Verma RK, Tripathi DK (2017b) Current scenario of root exudate-mediated plant-microbe interaction and promotion of plant growth. In: Kumar V, Kumar M, Sharma S, Prasad R (eds) Probiotics in agroecosystem. Springer, Singapore, pp 349–369
Wahyudi AT, Astuti RI (2011) Screening of Pseudomonas sp. isolated from rhizosphere of soybean plant as plant growth promoter and biocontrol agent. Am J Agric Biol Sci 6:134–141
Yadav G, Vishwakarma K, Sharma S, Kumar V, Upadhyay N, Kumar N, Verma RK, Mishra R, Tripathi DK, Upadhyay RG (2017) Emerging significance of rhizospheric probiotics and its impact on plant health: current perspective towards sustainable agriculture. In: Kumar V, Kumar M, Sharma S, Prasad R (eds) Probiotics and plant health. Springer, Singapore, pp 233–251
Zaidi A, Khan MS, Ahemad M, Oves M (2009) Plant growth promotion by phosphate solubilizing bacteria. Acta Microbiol Immunol Hung 56:263–284
Zhou LK (1987) The science of soil enzyme. The Science Press, Beijing, pp 267–270
Acknowledgements
Authors are thankful to Director MNNIT Allahabad for providing necessary facilities. The services and facilities provided by Central Instrumentation Facility under Department of Biotechnology, MNNIT Allahabad are also acknowledged.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Vishwakarma, K., Kumar, V., Tripathi, D.K. et al. Characterization of rhizobacterial isolates from Brassica juncea for multitrait plant growth promotion and their viability studies on carriers. Environmental Sustainability 1, 253–265 (2018). https://doi.org/10.1007/s42398-018-0026-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42398-018-0026-y