
Abstract
Seasonality is a critical driver of resource availability within individual generations and is therefore likely to have exerted selective pressures on hominin evolution. Nonetheless, it has frequently been overlooked in discussions of early technological variability. This paper examines the relationship between seasonality and lithic investment in the Oldowan of eastern Africa, to broaden our understanding of hominin behavioural adaptability. We develop a framework for the identification of seasonal signatures in the archaeological record and examine the nature of lithic variability across 12 Early Pleistocene localities. In combining these insights, we suggest that seasonality played a pivotal role in shaping the pattern of lithic investment in the Oldowan, with peak investment related to greater wet season processing of plant resources and mobility across the landscape. The productivity of cores, retouching of flakes, and use of percussive implements may each play a role in this seasonal patterning of tool use. Our analysis provides further evidence that Oldowan hominins were able to adapt their technological behaviours to changing environmental conditions, even at the scale of intra-seasonal fluctuations in resource availability, and how this interacts with landscape familiarity over longer timescales.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Oldowan, characterised by the production of sharp stone flakes by direct freehand or bipolar reduction (Gallotti, 2018), first appears 2.9–2.5 Ma in eastern Africa, initially identified at the sites of Nyayanga-1 (Kenya), Ledi Geraru BD1, and Gona OGS7, EG10, and EG12, with these latter four sites all in the Afar Depression of northern Ethiopia (Semaw et al., 1997, 2003; Braun et al., 2019; Plummer et al., 2023). Oldowan sites display a perhaps unexpected level of diversity in reduction schemes, material transport, and end products for the earliest widespread technology of the Early Stone Age, with a diversity of hypotheses put forward to explain this variation, including: repeated reinvention (e.g. Snyder et al., 2022), differential raw material use (e.g. Blumenschine et al., 2008; Braun et al., 2008a, b, 2009a, b; Reeves et al., 2021), ecological variability (e.g. Isaac, 1977), site-specific foraging strategies (e.g. Diez-Martín et al., 2010), and/or distinct biological or cultural groupings (e.g. Leakey, 1971; Delagnes & Roche, 2005; Plummer et al., 2023). This latter point has received particular recent attention as a result of the association between Paranthropus remains and the earliest Oldowan at Nyayanga (Plummer et al., 2023), with the broader picture suggesting repeated spatial associations with the Oldowan for both Paranthropus and early Homo (cf. Leakey et al., 1964). These co-associations are the inevitable consequence of substantial range overlap for both taxa (e.g. O’Brien et al., 2023), and thus for the remainder of this paper, we wish to focus on questions of environmental and functional variation.
A number of authors have argued for an increase in the complexity of Oldowan sites later in its range (e.g. Gallotti, 2018; Braun et al., 2019; but see, e.g. Semaw et al., 2009), although both expedient and elaborated toolkits are present throughout the Oldowan range. This co-existence is underscored by the unexpectedly high level of investment at Gona OGS7 ~ 2.6 Ma that stands in stark contrast to the marginally younger sites EG10 and EG12 in the same basin (e.g. Semaw et al., 1997, 2003; Stout et al., 2005, 2010, 2019). A recent analysis suggests that OGS7 clearly falls outside the variation of other early Oldowan sites on a number of technological variables, clustering instead with sites dating to < 2 Ma (Braun et al., 2019). At the same time, the late (~ 1.84 Ma) Oldowan site of FLK-Zinj falls within the range of early variation, suggesting a clear overlap in assemblage composition over time that cannot simply be explained by directional cognitive evolution. In this context, we may consider that the full range of Oldowan knapping abilities and technological solutions were present throughout, but were “more or less fully developed at different sites depending on a variety of factors, including raw material quality or site function” (Barsky et al., 2011).
Seasonality is one such factor that may have been responsible for important changes to site function even within the same year, but its relevance for understanding the nature of Early Stone Age lithic assemblages has so far been largely overlooked. It is known to have a dramatic impact on the foraging strategies of both nonhuman primates and modern human hunter-gatherers in tropical and subtropical latitudes (e.g. Clark & Linares-Matás, 2023; Hawkes et al., 1989; Lee, 2013; Marlowe & Berbesque, 2009; Oelze et al., 2014). For example, ethnographic studies of modern human hunter-gatherers in sub-Saharan Africa strongly emphasise a heavy bias towards acquisition of animal resources in the dry season, when ungulates tend to cluster around permanent water sources (e.g. Ichikawa, 1983; Vincent, 1985; Bunn et al., 1988; O’Connell et al., 1988, 1992, 2002; Hawkes et al., 1991, 2001; Weissner & N!aci, 1998; Marlowe & Berbesque, 2009; Sato et al., 2012; Lee, 2013; Hawkes, 2016). While modern human groups are in no way a model for the behaviour of Early Pleistocene hominins, it is highly likely that seasonality subjected hominins living in similar regions to intra-annual fluctuations in food availability throughout our evolutionary history (e.g. Foley, 1987, 1993; Joannes-Boyau et al., 2019; Linares-Matás & Clark, 2022; Speth, 1987; Sponheimer et al., 2005, 2006). Indeed, yearly seasonality reached levels comparable to the modern day by at least 2 Ma, confirming that Oldowan hominins had to adapt to regular changes to their landscapes, with the extent of this variation fluctuating rather dramatically over relatively short periods of evolutionary time (Blumenthal et al., 2019; Mercader et al., 2021).
We have previously argued that such seasonal variation in resource consumption would have had important implications for Oldowan landscape use, resource processing (Keeley & Toth, 1981; Lemorini et al., 2014, 2019), and investment in stone tool production (Linares-Matás & Clark, 2022). We attempted to test these hypotheses through the lens of Kanjera South (Kenya) and FLK-Zinj (Tanzania), arguing that the season of site occupation can be identified at specific sites in the archaeological record—wet season for the former and dry season for the latter—and that differences in prey selection and the nature of the two lithic assemblages are reflective of seasonal pressures on behavioural investment. We also speculated on the seasonal nature of Olduvai HWK-EE LAS (Pante et al., 2018), arguing it is related to the late dry season, and Koobi Fora FwJj20 (Braun et al., 2010), perhaps spanning a seasonal transition, to begin to explore implications for the wider Oldowan.
In this paper, we build on our previous work by consolidating the framework developed for identifying the season of site occupation at Early Stone Age sites in eastern Africa, before attempting to expand our sample of wet and dry season sites across the Oldowan range, discussing sites from the Olduvai, Fejej, and Gona palaeolandscapes. We then approach the relationship between seasonality and technological variability through a principal component analysis of selected technological features, and use these data to generate post hoc hierarchical clusters. We then discuss the implications for seasonal lithic investment in the wider Oldowan and how this might relate to landscape knowledge and site use.
Towards a Seasonal Framework
Defining the temporality of archaeological site formation at both shorter and longer time scales is crucial to interpreting hominin behaviour. Here, we focus on proxies of yearly seasonality at individual sites, but the severity and duration of the seasons will be inextricably linked to climatic change at the level of the landscape, as well as broader regional factors, and orbital forcing. We hope to relate these different levels of analysis in future work.
The study of the traces of feeding patterns on ungulate teeth represents the main analytical method for assessing seasonality in the Pleistocene, through the identification and quantification of relevant enamel alteration features. In particular, microwear texture analysis (MTA) has had success in better understanding dietary breadth and niche reconstruction (e.g. Domínguez-Rodrigo et al., 2019a; Rivals et al., 2018; Scott et al., 2005; Ungar et al., 2007), but this does not necessarily translate into high success when inferring the specific season(s) of occupation. For example, Domínguez-Rodrigo et al. (2019a) point out that Gurtov’s (2016) actualistic MTA dataset using impala (Aepyceros melampus) from Lake Eyasi was unable to successfully discriminate between wet and dry seasons. As such, while tooth-wear patterns can discriminate time-averaged palimpsests generated over multiple different seasons from assemblages accumulated over shorter periods of time within the same season, we must always use palaeoecological and taphonomic data to contextualise the specific timing of hominin presence at a site. Furthermore, even if the faunal assemblage encompasses multiple episodes of deposition, this does not rule out an increased frequency of hominin visitation at specific times of the year. Dissecting the dynamics of site formation processes at these Early Pleistocene palimpsests (Domínguez-Rodrigo, 2009) requires the integration of multiple sources of evidence.
One of the main aims of our previous work (Linares-Matás & Clark, 2022) was to develop such a framework for identifying the seasonality of site occupation(s) in the archaeological record of eastern Africa. This was largely based on studies of modern animal behaviour and on what the appearance, abundance, and treatment of specific taxa at individual archaeological sites might tell us about the timing of deposition. Application of these insights has been successfully applied to other regions and time periods: in the context of the Late Pleistocene Mediterranean, for example, Munro and Atici (2009) emphasise that seasonality is a critical factor to consider when exploring the accumulation of animal remains at archaeological sites because restricted seasonal use will produce assemblages dominated by few and seasonally-abundant taxa. In contrast, they suggested that multi-seasonal assemblages will display a greater diversity of species. This argument is directly relevant to seasonal contexts in other areas of the world.
While we acknowledge that (a) modern animal behaviour may have changed according to the abundance and behaviour of people alive today, and (b) the frequency of palimpsests adds substantial noise to such assessments, we argued from the records of Kanjera South and FLK-Zinj (Blumenschine et al., 2012; Bunn & Pickering, 2010; Domínguez-Rodrigo et al., 2010; Ferraro et al., 2013; Oliver et al., 2019; Parkinson, 2013, 2018; Parkinson et al., 2022) that not only is it possible to identify signatures of seasonality in the Early Pleistocene, but also that it provides important insights into the nature of hominin behaviour. At FLK-Zinj—where there is an underrepresentation of migratory taxa, an abundance of water-dependent reduncine bovids, a presence of pregnant waterbuck individuals, and meso- and micro-wear traces of a waterbuck mixed-feeding diet—the consistency of palaeoecological and analytical markers makes it likely occupation took place during a single season and perhaps a single year (Domínguez-Rodrigo et al., 2019a). Kanjera, on the other hand, was occupied repeatedly over the course of hundreds or thousands of years (Ditchfield et al., 2019), and yet preserves an abundance of juvenile individuals that is suggestive of repeated visits to the site during at least the early wet season. As such, while it is true that palimpsests are more likely to evidence a diversity of occupation signatures at the same site over time, including in different seasons, seasonal changes in mobility and landscape use may help to maintain the seasonal integrity of certain archaeological assemblages. This should be assessed using as many palaeoecological factors as possible.
At the same time, local palaeoecological factors will interact with properties of the specific ecosystem in which sites are found. For example, the Serengeti migration between Tanzania and southern Kenya impacts the relative abundance of particular taxa in these areas at specific times of the year, with wildebeest found in much greater abundance within the Serengeti environments surrounding the Olduvai palaeolandscape during wetter parts of the year (Domínguez-Rodrigo et al., 2019a). We can also draw particular attention to floodplains which, in semi-arid environments, are quite stable and resilient over time, due to their capacity to retain bioavailable organic carbon reserves when alternating between wet and dry phases, due to repeated flooding, recharging, and recession (Baldwin et al., 2013; Colloff & Baldwin, 2010). As such, these ecosystems experience predictable seasonal changes in the availability and accessibility of resources. With the onset of the rainy season, river channels start to fill, and begin to replenish the grasses and plant resources, turning the basin into a rich ecotone with riparian wetlands and lush grasslands, with very high resource biodiversity (Keddy, 2010) that provides hunter-gatherers with an opportunity to exploit a range of plant and aquatic resources (Dennell, 2007; Nicholas, 1998). However, as the wet season progresses, the river channel reaches capacity and the soils become saturated, making extensive flooding characteristic of the later periods of the wet season and into the early dry. This encourages the migration of non-water dependent terrestrial ungulates away from the proximal floodplain due to the lack of accessible grass (Sheppe & Osborne, 1971). Nonetheless, variation in the geographic extent of flooding may allow for the maintenance of a mosaic of habitats in more peripheral areas of the basin, as seen in the Okavango Delta of Botswana (Bartlam-Brooks et al., 2013a, 2013b; McCarthy et al., 2000).
As the water table slowly recedes, terrestrial animal species return to the ecosystem around the height of the dry season, attracted by renewed opportunities for fresh grazing (Sheppe & Osborne, 1971). As the dry season progresses, however, the carrying capacity of these environments again reduces considerably, since surface water, nutrient production, and high-quality plant biomass progressively decrease. In the Kafue floodplains of Zambia, for example, by the late dry season most grasses have either become unpalatable for ungulates or have already been destroyed by trampling, earlier grazing activities, and/or wildfires (Sheppe & Osborne, 1971). These changes in water and biomass availability have important implications for the seasonal signatures of Oldowan sites found in floodplains, such as at Gona (Quade et al., 2004, 2008).
While assessments of site seasonality in the Early Pleistocene are not abundant, some authors have attempted to define broad palaeoecological variables that could be used. We must stress that many of these variables derive from sites that cover large swathes of time, and thus encompass periods of broader scale climatic change (such as across Olduvai Beds I and II; Uno et al., 2018). Nonetheless, while such climatic fluctuations will have modified the relative length of wet and dry seasons (e.g. deMenocal, 1995, 2004), and produced periods of habitat reconfiguration (e.g. Lupien et al., 2021), many palaeoecological markers of seasonality are likely to have utility across time. For example, Speth and Davis (1976) examined the ratio of chelonids to ungulates and carnivores killed by San groups (G/wi and Ju/’hoansi) throughout the year to show clear seasonal differences in hunting patterns, with chelonids almost exclusively taken during the wet season. The authors then used NISP data to evaluate the faunal assemblages of a number of Olduvai sites in Beds I–II, showing that (a) the Olduvai sites fit well within the distinct clusters of San wet and dry season hunting, and (b) a large proportion of Olduvai sites are likely to have been deposited during the dry season. Four sites deviate from this overall pattern (DK, FLK NN level 3, FLK NN level 1, and MNK Skull Site), instead clustering with the Kalahari wet season data (Speth & Davis, 1976). Interestingly, recent taphonomic assessments suggest that the faunal assemblages at FLK NN 1 and 3 were primarily generated by carnivores rather than archaeological, with a very limited number of stone artefacts and extremely limited engagement with animal tissue (Barba & Domínguez-Rodrigo, 2007; Domínguez-Rodrigo & Barba, 2007a). Juvenile bovids are also well represented in these levels, suggesting faunal accumulations can show a strong wet season signature even in spite of limited hominin presence. Further still, FLK NN1 is a stratigraphic correlate of FLK-Zinj (Barba & Domínguez-Rodrigo, 2007), implying discrete areas of attractiveness for both carnivores and hominins at different points of the year.
The importance of chelonids as a wet season marker is reinforced by the bias towards Pelusios sinuatus at Olduvai (98% of chelonid NISP), a genus that survives today and has been observed to aestivate during the dry season (Auffenberg, 1981; Fritz et al., 2011; Lovich & Gibbons, 2021; Speth & Davis, 1976). This pattern of dry season aestivation and wet season signature of above-ground activity is typical of many other turtles and tortoises from dryland ecosystems, due to their vulnerability to dehydration and overheating (e.g. Chergui et al., 2019; Malonza, 2003; Petrozzi et al., 2020). Tortoises are also very susceptible to the impact of wildfires and/or prolonged drought conditions, with reductions in vegetation cover increasing predation pressures for tortoises (Longshore et al., 2003; Coutourier et al., 2014; Lovich et al., 2017; Chergui et al., 2019).
O’Connell et al., (1988, 1990, 1992) and O’Connell & Hawkes (in prep.) have also pointed out that Hadza ambush hunting exclusively occurs in the second half of the dry season, tends to be associated water sources, and can result in both single-individual and multi-individual, multi-taxon accumulations. In contrast, wet season encounter hunting results in kills being scattered across the landscape, not tied to water sources, and seldom contain body parts of more than one animal. On the other hand, the probability of finding scavengeable carcasses is higher along erosion embankments and the margins of riparian woodlands during the wet season in the Serengeti (Blumenschine, 1987; Hopcraft et al., 2005). The abundance of multi-individual, multi-taxon accumulations of bones near water sources in the archaeological record (mostly at sites representing relatively short periods of time) before 1.4 Ma—suggesting multiple bouts of carcass processing at the same spot—has led O’Connell and Hawkes (in prep.) to suggest that almost all zooarchaeological large bovid accumulations before this time were largely accumulated in the dry season. Contemporary observations of significant seasonal associations of certain gregarious species—such as the occurrence of mixed groups of plains zebra with Cape buffalo, Thompson’s gazelle, or Grant’s gazelle mostly during the wet season at the Serengeti and Tangarire-Manyara ecosystems in Tanzania (Kiffner et al., 2014)—could also prove relevant in elucidating seasonality signatures at Pleistocene sites. Finally, Stewart (1994) used exploitation of catfish (Clarias spp.) in a fluvial context at Olduvai BK (Upper Bed II) to suggest an occupation in the very late dry to early wet season. This is based on the seasonal behaviour of catfish, which migrate upriver en masse to spawn around the onset of the rains (Greenwood, 1955; Stewart, 1994).
Within our two previous case study sites, the intensity of faunal exploitation (and carnivore competition) was much greater at FLK-Zinj, with a much greater number of remains relative to the time represented by deposition, and a much greater emphasis on bone marrow. In contrast, the Kanjera hominins appeared to exploit within-bone nutrients only when the bones were likely to have preserved some fatty tissue during periods of body fat depletion (such as the tibiae or head contents; Ferraro et al., 2013; Parkinson et al., 2022; see Linares-Matás & Clark, 2022 for more extensive discussion of changes to bovid body fat throughout the year). These findings further hint that we can expect taphonomic patterns to differ between seasons, covarying with the extent of ungulate depletion (Bunn & Ezzo, 1993; Speth, 1987). In this context, and as we have discussed previously (Linares-Matás & Clark, 2022), we can also examine the Lower Augitic Sandstone (LAS) unit of HWK-EE, an Oldowan site in Olduvai lower Middle Bed II. Pante et al. (2018) report a pattern of fat-depleted marrow and epiphyseal grease avoidance by both carnivores and hominins at the site, characterised by high preservation of epiphyseal fragments relative to shaft fragments, alongside moderate-to-high tooth (33.7% and 25% for size 1–2 and size 3–4 animals, respectively) counts on midshaft fragments. The proportion of midshaft fragments with percussion marks for size 1–2 animals (6.5%) closely mirrors the 6–9% values reported for Kanjera South and are well below those from FLK-Zinj (Ferraro et al., 2013; Parkinson et al., 2022), while the greater engagement with larger animals (16.7% percussion-marked midshafts) is consistent with their better maintenance of fatty tissue in the face of seasonal reductions in forage quality (Bunn & Ezzo, 1993; Cordain et al., 2000; Linares-Matás & Yravedra, 2021). However, in contrast to Kanjera, engagement with animal resources is somewhat greater, perhaps suggesting any degree of fat depletion was not complete, while the assemblage also includes cut-marked hippopotamid remains, a considerable number of shed crocodile teeth (Pante et al., 2018), and a notable concentration of fish remains with a catastrophic mortality episode (Bibi et al., 2018). Modern observations of such mortality profiles tend to result from fish becoming trapped in pools as sources of water, such as lakes, recede in the (especially late) dry season (Leakey, 1971; Stewart, 1994). Extant crocodiles also tend to aggregate within increasingly isolated pools of water towards the end of the dry season (Njau, 2012), when they also share these localities with hippopotami (Kofron, 1993). This suggests an occupation of LAS at HWK-EE that encompassed the very late dry season, which would be consistent with the patterns of marrow avoidance and the indication of reduced grass availability given by bovid tooth micro- and meso-wear data (Rivals et al., 2018).
Bringing these insights together, we put forward a number of criteria for evaluating seasonality in the archaeological record (Linares-Matás & Clark, 2022). These are summarised in Table 1. Palaeoecological indicators of dry season occupations may include a clustering of water-dependent taxa (particularly reduncine bovids), with the late dry season in particular denoted by large concentrations of fish, shed crocodile teeth, and the co-occurrence of crocodile and hippopotamus remains. In contrast, we may be able to discern a wet season signature from an abundance of chelonid remains, and an early wet season signature from an abundance of juvenile bovids and the presence of catfish in fluvial contexts. Taphonomically, we may expect dry season sites to have a higher density of faunal remains, alongside greater carnivore competition; in the late dry, an increased preference for body parts that retain their fat content throughout the year replaces a high emphasis on meat and especially marrow. This change in preference may also be seen in the early wet season. The wet season more broadly may be associated with lower densities of faunal remains, and relatively limited carnivore competition. Nonetheless, palaeoecological indicators should be more important than taphonomic ones in determining a season of occupation, given they are less prone to (although not free from) biases associated with hominin behaviour.
Seasonal Site Signatures
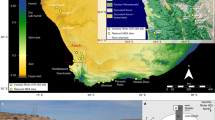
Using the framework outlined above, we now attempt to expand the number of Oldowan sites with seasonal attributions by examining sites from three main palaeolandscapes: Gona, Fejej, and Olduvai (and see Supplementary Information for additional site-level details). In Table 2, selected sites from these landscapes are described according to their palaeoecological and taphonomic signatures, alongside additional Oldowan sites we have discussed above and elsewhere. We also include a map of eastern Africa with the four Early Pleistocene palaeolandscapes discussed in the text (Fig. 1). For more extensive descriptions of seasonality at FLK-Zinj, Kanjera South, and HWK-EE, see Linares-Matás and Clark (2022).

Map of eastern Africa with the four main archaeological areas discussed in the paper
Gona
At Gona (Awash Basin, Ethiopia), multiple localities in the Busidima Formation (notably EG10, EG12, and OGS7) preserve lithic artefact assemblages dated to the period 2.6–2.5 Ma (Levin et al., 2022; Stout et al., 2010). The coeval, nearby sites of EG13 and OGS6 exhibit taphonomic evidence for primary access to carcasses (Cáceres et al., 2015; Domínguez-Rodrigo et al., 2005), but faunal data is otherwise extremely sparse for the two site clusters. Faunal and palynological data from OGS6 and OGS7 highlight an abundance of afromontane taxa and a strong wooded signal in an otherwise mixed environment (López-Sáez & Domínguez-Rodrigo, 2009). EG10 and EG12 are characterised by a predominance of local trachyte and rhyolite raw materials exploited through unifacial flaking, whereas OGS7 shows selective (but still fairly limited) transport of aphanitic and vitreous volcanic materials and an abundance of bi- and multi-facial reduction schemes (Stout et al., 2010). The limited lithic assemblage of OGS6 may be related to OGS7, which lies nearby, by the absence of whole flakes with cortical platforms (Stout et al., 2010).
Stout et al. (2010) suggest possible mechanisms for the distinction between the East Gona and Ounda Gona assemblages, including (a) different cultural variants or (b) a difference in economic strategy between the two clusters in the context of shared technological basis. The presence of EG10 and EG12 in a semi-arid proximal floodplain and of OGS7 in a channel bank or margin context suggests a degree of ecological divergence between the two localities. At the same time, there cannot be a simple correlation between these environments and technological strategies (Stout et al., 2010), given the apparent technological connection between OGS7 and OGS6. OGS6 is located in a proximal floodplain, which allies it more closely with the palaeoenvironments of the East Gona sites, rather than OGS7. Alternatively, the economic differences between the EG and OGS lithic assemblages may have been driven by the markedly seasonal nature of the Gona palaeolandscape (cf. Quade et al., 2004, 2008; Stewart & Murray, 2020), and the resulting variability in the predictability of resource distributions (Clark & Linares-Matás, 2020). While we do not have the depth of data to make a confident seasonal attribution, we can explore the possibilities for seasonal use of the Gona palaeolandscape.
Given the location of EG10 and EG12 in a proximal floodplain, dry season dynamics may have provided hominins with opportunities to maximise animal exploitation, with the second half of the dry season providing a particular opportunity for the interception of herbivores in the process of reaching perennial watercourses (O’Connell et al., 1988, 2002; Hawkes et al., 1991, 2001). EG13 (in the same spatial cluster as EG10 and EG12) and West Gona-9 (~ 500 m west of the EG sites), two sites located on the proximal floodplain of the Kada Gona River, show evidence of early access to carcasses. This includes a cut-mark on a medium-sized bovid rib—consistent with evisceration—at EG13, and meat filleting on a small bovid scapula and the upper limb bone midshaft of a medium-sized bovid at WG-9 (Domínguez-Rodrigo et al., 2005: 116–117).
We can expect Gona hominins to have had an opportunity to exploit more predictably a wider diversity of aquatic and plant resources during the early wet season. In this context, taphonomic data suggest the handling and processing of terrestrial and freshwater carcasses at OGS6, with skinning or disarticulation cut-marks noted on an equid calcaneus (Domínguez-Rodrigo et al., 2005) and on a crocodilian skull (Cáceres et al., 2017). These data may also indicate preferential body part selection, perhaps including for fatty within-bone nutrients at a time of depletion. Bartlam-Brooks et al. (2013a) also point out that equids are known to require a large dietary input to meet their nutritional requirements, and thus prefer areas of seasonal grass abundance (see also Tucker et al., 2020). As such, the presence of anthropogenic traces on the OGS6 equid may imply relatively high local biomass availability in the floodplain during the period of hominin activity.
Fejej
Fejej is located in the eastern Omo-Turkana basin, at the northern limit of the Koobi Fora Formation. Horizon C1 is a short-term archaeological site within a highly localised horizontal distribution of fauna and lithics, with an average vertical thickness of only c.20 cm (de Lumley & Beyene, 2004; Barsky et al., 2011). The faunal and lithic assemblages of this level also appear in spatial association, with taphonomic data suggesting rapid burial and a functional relationship between the two (Echassoux et al., 2004). This is suggestive of high temporal resolution, and of an assemblage that could have formed within a single year or season.
The faunal profile is dominated by impala (Aepyceros shungurae), followed by kudu (Tragelaphus nakuae), in the vicinity of a permanent water source (Barsky et al., 2011). The presence of baboons is also of note, as impala tend to aggregate near baboons throughout the year, save during the birthing (early wet) season, given baboons prey on young impala (Baenninger et al., 1977; Estes, 1991; Kiffner et al., 2014). The wet season markers of catfish and chelonids are completely absent from this unit, despite their presence elsewhere in the Fejej sequence (Barsky et al., 2011).
The impala age profiles comprise one juvenile, one young adult, three prime adults, and one older adult (Echassoux et al., 2004). A considerable number of oblique and spiral fracture planes, some associated with percussion marks, may evidence marrow extraction, but no cut-marks have been reported (Echassoux et al., 2004). Interestingly, no evidence of carnivore activity (tooth marks, furrowing, coprolites) was noted either, even though remains of canids and a leopard-sized felid were found elsewhere in Unit 3 (Echassoux et al., 2004). Thus, while cut- and percussion-marks are perhaps underrepresented, the high bone fragmentation is most consistent with hominin exploitation of non-depleted carcasses, as would be expected for dry season subsistence (Linares-Matás & Clark, 2022). This concords well with the palaeoecological signatures noted above.
The stone tool assemblage from the broader Unit 3 is characterised by raw materials from local alluvial deposits, primarily involving the selection of quartz (91%; de Lumley et al., 2004). On-site stone tool manufacture is illustrated by several instances of refitting, with reduction (occasionally bipolar on anvil) leading to the production of hundreds of small flakes (mostly between 2 and 3.5 cm) through relatively short reduction sequences (Barsky et al., 2011; de Lumley et al., 2004). The site also displays an apparently high proportion of percussive tools, but the true extent is unclear as previous publications have included cobbles without signs of battering in the same category as those that do (Barsky et al., 2011; de Lumley et al., 2004).
Olduvai
DK
DK lay next to a semi-permanent water source and seasonal streams, ~ 5 km east of the main cluster of Upper Bed I sites (including FLK-Zinj, FLK-N, and DS; Domínguez-Rodrigo et al., 2010). The faunal assemblage at DK represents a multi-year palimpsest with minor anthropogenic input, also incorporating felid predation and accumulation, hyaenid secondary ravaging, and likely some natural deaths that both hominins and carnivores were reluctant to engage with (Egeland, 2007a, 2007b). While hominins did occasionally engage with the fauna, the specific duration of this overlap is uncertain. At the same time, the bone assemblage presents consistent markers of wet season deposition, including at times of hominin interaction: alongside the dramatic number of chelonid remains mentioned above (Speth & Davis, 1976), there is also a high proportion of juvenile bovids in DK level 3 for both small- (50%) and medium-sized (61.5%) animals (Egeland, 2007b), which closely mirrors values from Kanjera South (Oliver et al., 2019). Further still, the avoidance of meat from natural deaths is consistent with its reduced attractiveness at specific times of the year, as previously documented for the early wet season of Kanjera South (Linares-Matás & Clark, 2022).
In terms of the lithic assemblage, different authors have varied dramatically in their interpretation, with some emphasising somewhat greater complexity (e.g. Sánchez-Yustos, 2021) than others (e.g. de la Torre & Mora, 2005; see Supplementary Information for more discussion). It is clear that the site also contains an unusually high proportion of lava pieces (~ 83%) within the worked material for an Olduvai site (de la Torre & Mora, 2005; Sánchez-Yustos, 2021). This may suggest either a preference for more durable cutting edges (cf. Key et al., 2020), a focus on tasks that can be performed better with a blunter edge—such as skinning and cutting tubers (Jones, 1994)—or simply a factor of increases in quartz (following Tarriño et al., 2023) raw material transport costs with distance from the likely material source (Blumenschine et al., 2008).
This emphasis on blunter and/or more durable edges may be mirrored in an unusually high proportion of flake retouch for an Oldowan site (Sánchez-Yustos, 2021; but see de la Torre & Mora, 2005), as retouched flakes provide limited advantage in butchery or wood scraping when raw material availability is not a limiting factor (e.g. Clarkson et al., 2015; Galán & Domínguez-Rodrigo, 2014; Jones, 1994). It is notable that transport of lava materials was so limited and yet 16.6% of these flakes are still reported to have been retouched by Sánchez-Yustos (2021), which would suggest that the high representation of durable lavas (cf. Key et al., 2020) and the intensive reduction schemes a greater reflection of the intensity of activity, not simply duration.
FLK-N I-II
FLK-N I-II (1.79 Ma) is the site of freshwater springs present during Upper Bed I times (Ashley et al., 2010), with phytolith analysis suggesting dense woody cover at the site (Barboni et al., 2010). The FLK-N I-II faunal assemblage shows a predominance of the extinct alcelaphine Parmularius altidens and springbok (Antidorcas recki), but is otherwise quite diverse (Domínguez-Rodrigo & Barba, 2007b). Kiffner et al. (2014) note that modern counterparts of several other species found in the assemblage (namely spotted hyaenas, Cape buffalo, hippopotamus, wildebeest and plains zebra) primarily co-occur in mixed species groups across the Serengeti landscape during the wet season. Micro- and meso-wear data on the Parmularius specimens suggest some degree of time-averaging in the accumulation, with carcasses deposited across several seasons (Domínguez-Rodrigo et al., 2019a).
Taphonomic data suggests that the accumulation and modification of the FLK North I–II faunal assemblage is primarily the result of carnivore activity (felid-generated accumulations partially ravaged by hyaenas; Domínguez-Rodrigo & Barba, 2007b). There was only minor hominin interaction with the assemblage, with evidence of hammerstone percussive activity for marrow extraction particularly limited (Domínguez-Rodrigo & Barba, 2007b). At the same time, the lithic assemblage shows evidence of intensive pounding behaviours—which are therefore likely to relate to the exploitation of vegetal resources (Diez-Martín et al., 2022)—alongside elaborate core reduction for an Oldowan site (Diez-Martín et al., 2010; Proffitt, 2018).
DS
At Olduvai DS, the rich faunal assemblage has a clear predominance of waterbuck near a freshwater source, consistent with the dry season behaviour of this species (Ashley et al., 2010; Cobo-Sánchez, 2020; Diez-Martín et al., 2021; Linares-Matás & Clark, 2022). Preliminary micro- and meso-wear of Parmularius altidens teeth suggest that these animals consumed a mixed diet towards the end of their life, suggesting these animals were also deposited during the dry season (Domínguez-Rodrigo et al., 2019a). Notably, the site shows a similar signature of animal acquisition to FLK-Zinj, with a predominance of prime-aged individuals among the medium-sized bovids, high proportions of cut marks (c.20%) and percussion marks (16.5%) on long limb bone specimens with good cortical preservation, and a clear distribution of cut-marks on areas that would be first defleshed by carnivores (Cobo-Sánchez, 2020). DS also displays a clear spatial correlation between bones and lithic remains (Cobo-Sánchez, 2020; Diez-Martín et al., 2021), supporting the functional association between the dry season faunal remains and lithic assemblage. As for the lithic assemblage itself, core reduction at the site was not particularly intensive, with quartz material selectively exploited more intensively (Diez-Martín et al., 2021). This perhaps mirrors FLK-Zinj in a preference for sharper flakes despite a greater abundance of alternative materials (cf. Key et al., 2020). It is worth keeping in mind, however, that basalt flakes are significantly underrepresented at the site compared to the number of basalt flake scars, perhaps suggesting transport of useful (and durable; cf. Key et al., 2020) flakes out of the site (Diez-Martín et al., 2021). This additional desire for blunter edges and/or durability may also be reflected in the relatively high proportion of retouch on basalt, phonolite, and quartz (Diez-Martín et al., 2021).
Oldowan Lithic Variability
Using our assignments of seasonality for Kanjera South and FLK-Zinj, we previously suggested that there may have been seasonal variation in the extent of technological investment during the Oldowan. In particular, we argued the relatively expedient toolkit of FLK-Zinj—including relatively short reduction sequences on local raw materials and a low incidence of retouch (de la Torre & Mora, 2005; Leakey, 1971)—were reflective of a dry season pattern of subsistence in animal tissue was the primary driver of a need for sharp edges (Linares-Matás & Clark, 2022). We suggested that this would lead to an unpredictable and reactive need for core reduction, with flakes produced only when a foraging opportunity became available, on abundant local and/or cached raw materials. In such a scenario, there would also be little benefit to intensive reduction schemes. In contrast, we argued that the increased transport of raw material and reduction intensity at Kanjera South (Braun et al., 2008a, 2009a, 2009b; Lemorini et al., 2014, 2019; Reeves et al., 2021; see also Favreau, 2023) reflect an increased availability of predictable attractive plant resources in the wet season. This would have allowed hominins to better plan their activities throughout the landscape, facilitating greater transport of material and a corresponding intensification of reduction and retouching to maximise the utility of raw material. There will have been deviations from this trend—for example, due to inter-annual changes to the intensity of individual seasons (cf. Hawkes et al., 1991), knowledge of the landscapes hominins were inhabiting (cf. Clark & Linares-Matás, 2020), or temporal changes across the Oldowan range (cf. Gallotti, 2018)—but we expect broad patterns of seasonal changes in complexity to hold over time.
Methods
Here, we attempt to relate our survey of seasonality literature at individual sites to available data on lithic technology to a larger sample of Oldowan localities, both from the sites introduced above and those for which data are limited but we wish to include in the wider discussion. These are shown in Table 2, where we have collated data on five key metric variables to describe the respective lithic assemblages. The variables have largely been chosen to reflect reduction intensity and material use, including the percentage of unifacial and test cores, the ratio of average flake scar count to the log-corrected mean core length, and the ratio between mean flake thickness and mean maximum reported dimension for those over 20–25 mm in size. This latter variable was taken as a marker of flake thinness relative to overall size, and therefore as a measure of investment in maximising raw material utility (Lin et al., 2013; Režek et al., 2018). It would have been better to calculate these values for individual artefacts rather than from assemblage-level averages, but these data are not widely available in existing publications. We have also sourced data on retouched pieces as a proportion of whole flakes and retouched pieces, alongside the proportion of percussive tools in the assemblage, given our previous discussion into the role of seasonality in the extractive foraging of nuts and seeds (Clark & Linares-Matás, 2023; Linares-Matás & Clark, 2022).
Our approach and variables are originally based on a dataset presented by Braun et al. (2019) in their description of Ledi-Geraru—and since reused by Mercader et al. (2021) with reference to Ewass Oldupa—but data are retrieved always from the original sources whenever feasible. This has produced a number of small differences in individual values compared to that dataset. Data are derived from Braun et al. (2019) if no primary source of data could be accessed. Braun et al.’s (2019) k-means cluster analysis highlighted the technological consistency of Oldowan assemblages in relation to the material from Lomekwi-3 (Harmand et al., 2015), flaked tools produced by Capuchin monkeys (Proffitt et al., 2016), or from the early Acheulean. It also highlighted interesting dimensions of variability within the Oldowan beyond directional chronological trends, which we wish to explore further here.
Following Braun et al. (2019) and Mercader et al. (2021), we conducted a principal component analysis (PCA) on our data, alongside post hoc hierarchical clustering analyses with the site-level PC values. All data analyses were carried out in RStudio (Posit Team, 2022) running R version 4.2.2 (R Core Team, 2022). Our code and principal component data used as the input for the hierarchical clustering are available as Supplementary Information. Where multiple estimates have been presented for a variable at any given site, we took the mean value for use in our analyses. While not ideal, this allowed us to perform our analyses without having to arbitrarily select the value of one author(s) over another. All variables were standardised to mean zero and standard deviation one before analysis was carried out. To account for the missing retouch values for Kanjera South, OGS7, and Nyayanga-1, we ran the analyses twice: once without retouch as a variable, and once with the missing values estimated by a mean from all other sites, the simplest estimation approach (cf. Dray & Josse, 2015). This approach almost certainly overestimates retouch values for OGS7 (and likely Nyayanga), as the true value is likely to be low (Semaw, 2006), but as the results below suggest, its relative PCA placement was not substantially altered whether retouch was included or not.
For the cluster analyses, we tested the agglomerative coefficients (AC) of each possible method, with Ward’s method—which aims to minimise within-cluster variance—outperforming the others (AC with retouch = 0.849 and AC without retouch = 0.858), namely average distances between each point in a cluster (0.670 and 0.683, respectively), the closest distances between single points (0.449 and 0.413, respectively), and “complete”, i.e. the closest distances between furthest-separated points between each cluster (0.797 and 0.805, respectively). We subsequently carried out an entanglement analysis between the two dendrograms, giving a coefficient between 0 and 1 describing their level of alignment. Values closer to zero indicate fewer branching differences and those closer to one entail greater distinction.
Results
Figures 2 and 3 display the results of the PCA on PC1-3 for each site, with retouch included and not included in the analysis, respectively. When retouch is included, PC1 (30.9% of variation) largely reflects the proportion of flakes retouched, the proportion of unifacial cores, and the average number of flake scars for cores of a given log-standardised length, while PC2 (25.1% of variation) displays a greater influence of mean flake thickness/mean flake maximum dimension, alongside the size-controlled scar average. PC3 (20.3% of variation) reflects the proportion of percussive material, and to a lesser extent the proportion of unifacial cores. When retouch is excluded, PC1 (33.2% of variation) is associated with mean flake thickness/mean flake maximum dimension, as well as the percentage of unifacial cores. PC2 (26.6%) values are associated with the proportion of percussive elements, as well as the average number of flake scars for log-standardised core length and the proportion of unifacial cores, while PC3 (24.3%) relates to the size-controlled flake scar count, and to the proportion of percussive elements.

Results of PCA for the sites in Table 3, with retouch included in the analysis. Site values for PC1 and PC2 are shown in A, the contributions of the top four variables to PC1 and PC2 in B, site values for PC1 and PC3 are shown in C, and the contributions of the top four variables to PC1 and PC3 are shown in D. Key: pc.unifacial = percentage of unifacial and test cores, fsc.lmmd = mean core flake scar count / log mean length, mft.mmd = mean flake thickness / mean flake length, pc.retouch = retouched pieces as a percentage of whole flakes + retouched pieces, and pc.percussive = proportion of the assemblage relating to percussive activities

Results of PCA for the sites in Table 3, with retouch not included in the analysis. Site values for PC1 and PC2 are shown in A, the contributions of the top four variables to PC1 and PC2 in B, site values for PC1 and PC3 are shown in C, and the contributions of the top four variables to PC1 and PC3 are shown in D. Key: pc.unifacial = percentage of unifacial and test cores, fsc.lmmd = mean core flake scar count / log mean length, mft.mmd = mean flake thickness / mean flake length, pc.retouch = retouched pieces as a percentage of whole flakes + retouched pieces, and pc.percussive = proportion of the assemblage relating to percussive activities
In both PCAs, Kanjera South and OGS-7 cluster tightly, suggesting that the use of a mean retouch value is not resulting in an overly biased output. Indeed, Kanjera and OGS7 sit together in every analysis run, as do EG10 and EG12. HWK-EE LAS and DS may show a similar association. DK lies with Kanjera and OGS7 on PC1 and 2 of the analysis which includes retouch, and on PC1 and 3 of the analysis which excludes it. The two dendrograms relating to the hierarchical cluster analysis are shown in Fig. 4. They confirm the close relationships between Kanjera South and OGS7 on one hand, and the East Gona sites on another. They also highlight a consistent technological association between DS and HWK-EE LAS, between Fejej, FLK-Zinj, and Nyayanga-1, and between DK and FLKN to the exclusion of the other Olduvai sites (perhaps excluding Ewass Oldupa when retouch is excluded). The entanglement analysis, shown in Fig. 5, provides a value of 0.575 for the two models, suggesting moderate agreement. DS, HWK-EE LAS, HWK-EE Lemuta, Fejej, FLK-Zinj, and Nyayanga form a consistent cluster in each dendrogram, with the main differences coming in the placement of Ewass Oldupa and East Gona: either forming an outgroup to all other sites, or forming a cluster with FLK-N and DK.

Dendrograms showing the results of a post hoc hierarchical clustering analysis of the principal component values. Analysis including retouch is shown in A and excluding retouch in B

Entanglement analysis of the two dendrograms presented in Fig. 4, with retouch included on the left and not included on the right
These findings reveal interesting commonalities with the analysis of Braun et al. (2019), most notably in the clustering of OGS7 with later Oldowan sites, especially Kanjera, and the distinction between DK and FLK-Zinj. Braun et al. (2019) interpreted their clusters as the result of temporal differences throughout the Oldowan, with OGS7, Fejej, and FLK-Zinj forming individual exceptions to a broad shift in Oldowan knapping skill that distinguished sites at the end of its range from those at the beginning. However, our addition of the late assemblages from HWK-EE and DS—not included by Braun et al. (2019)—add other possible exceptions to this overarching pattern, given they consistently cluster closer to FLK-Zinj than to Kanjera or DK.
We argue that seasonality may, instead, be driving the key markers of Oldowan inter-assemblage variability reflected in these distinct clusters. In this context, the clusters shown in Fig. 4 are extremely promising, with Kanjera and FLK-Zinj located at either end of our hierarchical clustering analysis in Fig. 4A. In the next section, we contextualise these results with site-specific data to explore the relationships between the lithic data from these assemblages and seasonality.
Discussion
Seasonality and the Early Oldowan
We previously used the records of Kanjera South and FLK-Zinj to suggest that sites with a wet season signature would have seen greater raw material transport distances as hominins increased their planned mobility across the landscape, producing a corresponding increase in reduction intensity, a greater selection of durable materials, and an increase in flake retouch to maximise the utility of transported material and/or to carry out more abrasive cutting tasks (Linares-Matás & Clark, 2022). The data summarised in Table 3 show quantitatively that unifacial reduction methods are indeed much less common at Kanjera South—where they are exceedingly rare—than at FLK-Zinj, with slightly higher core productivity also seen in the Kanjera assemblage. The Kanjera flakes are also comparatively thinner for their length than those from FLK-Zinj, similarly implying greater raw material utility (cf. Lin et al., 2013; Režek et al., 2018). No data are available on the frequency of retouch at Kanjera South, making it difficult to quantitatively compare, but FLK-Zinj displays between 3.91% (Proffitt, 2018) and 10.7% (de la Torre & Mora, 2005) of whole flakes retouched. It is worth noting, however, that the 10.7% is an overinflation of the retouch signature identified by de la Torre and Mora (2005), as 73.3% of retouched specimens were made on flake fragments, with fragments not included in the unretouched component of the calculation. The four retouched specimens on whole flakes would correspond to a value of 3.1% of whole flakes, while including flake fragments in the original calculation would lower it further to 1.5%.
The principal component and hierarchical clustering analyses presented here suggest a clear structure to the nature of lithic variability in the wider Oldowan. From the individual faunal assessments of site seasonality at localities such as Kanjera South, FLK-Zinj, HWK-EE (see Linares-Matás & Clark, 2022), DK, FLK-N, DS, and Fejej (see below), we argue that seasonality plays a critical role in patterning this variability across the Oldowan range. An obvious question that arises from these potential differences in technological investment between seasons in the Oldowan is how far back such a pattern may have gone in time; a question that is heavily dependent on the function(s) for which Oldowan tools were being deployed. The focus on flaked tools that emerges in the Oldowan is likely an initial investment in the processing of animal and/or plant tissue with a greater efficiency (e.g. Clark & Linares-Matás, 2023; Domínguez-Rodrigo et al., 2005; Hovers, 2012; Shea, 2017). Indeed, use-wear analysis from the site of Nyayanga-1 suggests the very early Oldowan is associated with deployment of flaked tools in a diversity of both plant and animal processing tasks, alongside limited evidence of anthropogenic modification of animal, including megafaunal, remains (1.03% NISP; Plummer et al., 2023). The lithic data available for the site are broadly in keeping with other early Oldowan sites—especially in the relatively high percentage of unifacial cores (50.79%) and a relatively low number of flake scars for cores of their log-controlled size—but the notable bifacial component is in keeping with expert knowledge of fracture mechanics and how best to produce a consistent series of stone flakes (Plummer et al., 2023). Nonetheless, Nyayanga-1 still lies closest to the next oldest Oldowan site of Ledi-Geraru—the most distinctive of any Oldowan site in each of the first principal components of Braun et al. (2019), Mercader et al. (2021), and Plummer et al. (2023)—on Plummer et al.’s (2023) PC1. The distinctiveness of these two assemblages on one or more variables may be expected for sites so close to the beginning of the Oldowan range.
We may even consider the nature of lithic investment at Nyayanga as the outcome of a much longer technological learning trajectory and seasonal resource use, perhaps beginning at least as early as the Lomekwian 3.3 Ma (Flicker & Key, 2023). Indeed, that Nyayanga shows some similarities with Lomewki-3 can be seen in their distinctly high proportion of cores relative to the rest of the assemblage, as well as the elevated percentage of cores with evidence of percussion (Harmand et al., 2015; Plummer et al., 2023). Given the diversity of resources being processed at Nyayanga (Plummer et al., 2023), it seems likely that such a process of learning how and when to deploy sharp flakes must have involved adaptation to changes in resource availability across the year. How exactly this relationship manifested, however, is obscured by the great difficulty in identification of a prevailing seasonal signature at either of these sites, especially at Lomekwi-3, where only 33 faunal fragments were reported from the site itself, with none displaying evidence of anthropogenic modification (Harmand et al., 2015). At Nyayanga-1, faunal preservation is a great deal better (Plummer et al., 2023), but signatures of seasonality are conflicting: the presence of reduncine bovids near a water source (river channel) and the co-occurrence of hippopotamids and crocodiles may indicate a dry season input, while the presence of chelonids may reflect a wet season input. We do note, however, that the assemblage clustered firmly with sites attributed to the dry season in our PCA and cluster analysis (Figs. 2, 3, and 4).
The discussion of technological distinction between the East Gona and Ounda Gona South sites is also interesting in this regard, as we may hypothesise that seasonal differences were driving the lithic variation we see in the Gona palaeolandscape at 2.6–2.5 Ma. There is clearly a substantial difference in investment between EG10 and EG12 on one hand and OGS7 on the other, with unifacial reduction so much more infrequent at the latter, which occurred alongside increased productivity of cores, thinner flakes, and a greater transport of raw material (Stout et al., 2005, 2010, 2019; Table 3). The unexpected level of complexity witnessed for the OGS7 assemblage also confirms a high level of familiarity with stone tool manufacture early in the Oldowan’s temporal range (Stout et al., 2010). As highlighted above, these differences cannot be readily explained by developments in knapping skill or investment over time, as OGS7 is somewhat older than the East Gona sites, nor can it be related to differences in habitat use, given the limited assemblage of OGS6 may be technologically linked to OGS7 and yet shares with the East Gona sites a floodplain depositional context (Stout et al., 2010). While data for the sites are far too scarce to make confident assignments of seasonality, modern ecosystem dynamics in analogous basins make it likely that distinct economic pressures were acting on hominins at different times of the year.
Indeed, our results, alongside those of Braun et al. (2019), emphasise that Gona displays a strong distinction between the OGS7 and EG10-12 assemblages, with OGS7 allying with sites with high levels of investment dated to after 2 Ma. This leads us to consider if Oldowan tool production could have been biased towards the dry season in the earlier parts of its range, in an analogous process to chimpanzee use of individual tools—frequently directed at resources available at particular times of the year (see Clark & Linares-Matás, 2023 for a review). Given the temporal association between the emergence of Oldowan tool use and evidence of hominin processing of animal carcasses (Domínguez-Rodrigo et al., 2005; Plummer et al., 2023), the consistency of sharp-edged tools in the Oldowan may reflect longer-term (pre-Pleistocene) interactions with novel prey types and an accumulation of landscape knowledge pertaining to their exploitation (Clark & Linares-Matás, 2023; Thompson et al., 2019). At such a point when hominins would have been able to gain semi-regular access to animal remains, they may then have been able to seasonally invest in flaked stone technologies upon acquisition of carcasses, because the returns would predictably exceed the costs of tool manufacture. We would expect this use would to be highly expedient and on immediately available raw materials (cf. Linares-Matás & Clark, 2022).
The existence of such knowledge surrounding tool use could then be directed fairly quickly towards the use of similar resources at other times of year where opportunistically available, but also in experimentation with novel resources (Clark & Linares-Matás, 2023). Such a process has been repeatedly observed in ape populations. For example, Gruber et al. (2009) report that chimpanzees from Kanyawara and Sonso, Uganda, each used pre-existing knowledge of tool use when engaging with a novel food acquisition task. When presented with honey within holes drilled into logs, the Kanyawara individuals (who occasionally use sticks to extract the waxy honey of carpenter bees) fashioned sticks that could be used to scrape out the honey, whereas the Sonso chimpanzees used their fingers or attempted to use leaf sponges that they use to obtain water from hollows in trees. These findings highlight the value of redeploying existing technologies in adapting to novel situations and gaining access to resources that are otherwise opaque or embedded (Clark & Linares-Matás, 2023).
Wet season deployment of early Oldowan tools may have initially involved an elaboration of production sequences that were originally shorter and biased towards other parts of the year, when returns from individual foraging bouts where high and success was predictable. This may be observed in the context of OGS7. However, there may also have been constraints against such increased investment in the early Oldowan, including reduced mobility compared to later phases of the Early Pleistocene (Linares-Matás & Yravedra, 2021), reduced amount of time in specific landscapes (Clark & Linares-Matás, 2020), or a patchy distribution of plant resources whose consumption could have been substantially aided with sharp-edged stone. These factors may restrict the need or ability to carry material between resources—and for a corresponding investment in its utility (Linares-Matás & Clark, 2022)—or remove the need for their deployment in the first place. This may explain why only later do we see substantial increases in material transport (particularly at Kanjera South and Ewass Oldupa), and an inferred repeated wet season deployment of lithic material.
Seasonal Differences in Lithic Investment
We have much clearer faunal data on Oldowan seasonality later on in its range, especially from 2 Ma, that allow us to make clearer links to the variability seen in the lithic record. Following the information presented above, Olduvai DK—based on the high proportion of chelonid and juvenile bovid remains, as well as taphonomic data from level 3—may have been occupied during the wet season, and DS—based on the concentration of waterbuck individuals and micro-wear suggestive of a mixed feeding diet at the time of death—is likely to have seen greatest hominin activity during the dry season. We also suspect a strong wet season influence is present at FLK-N I-II, and a dry season input at Fejej FJ-1a, but fewer data are available to test these hypotheses. These attributions are nonetheless important as they provide a broader platform for inferences about seasonal influences on hominin behaviour, beyond the two sites we initially used to generate our hypotheses (Linares-Matás & Clark, 2022). In particular, we have used these additional sites to examine our predictions, based on the Kanjera South and FLK-Zinj lithics, that we should expect to see greater lithic investment in the wet season compared to the dry season due to differences in tool-mediated resource exploitation.
As discussed above and in the Supplementary Information, the nature of the DK assemblage is subject to different interpretations between authors, but a number of metrics agree that the intensity of reduction was high for an Oldowan site, with among the highest average number of flakes released from cores when controlling for size, and a relatively low number of unifacial cores identified by two out of three authors (Table 3). This places it alongside Kanjera and OGS7, and possibly FLK-N—to which it displays a number of similarities—and accounts for the close hierarchical clustering where retouch is included. The potential relationship between the four sites highlighted in Fig. 4A is particularly interesting, as a wet season attribution for each site would match our predictions for an increase in investment during this time of the year. However, the association between Kanjera and OGS7 on one hand and DK and FLK-N on the other was impacted by the inclusion of retouch in the analysis, with this figure presenting difficulty for all four sites; having been not reported for Kanjera and OGS7, and having a range of values at DK and FLK-N.
Sánchez-Yustos (2021) suggests DK has an extremely high proportion of retouch for an Oldowan site at 19.87% of whole flakes (but see de la Torre & Mora, 2005), a figure that would be rivalled only by the 20.81% of retouched flakes at Garba IVE (Gallotti, 2018; Gallotti & Mussi, 2015) and the 14.34% reported for DS (Diez-Martín et al., 2021). This DK retouch figure increases to 50% for quartz, suggesting a dramatic attempt to maximise the utility of this raw material, something also noted in the intensity of its reduction (Reti, 2016; Sánchez-Yustos, 2021). Nonetheless, the potential 16.6% of retouched lava flakes suggest that the overwhelming presence of local volcanic materials in the assemblage is not simply an artefact of prohibitive transport costs (Blumenschine et al., 2008), but perhaps an intentional selection of durable materials and an attempt to maintain edges where they blunted; a process that would happen quicker for quartz (cf. Key et al., 2020). Assuming Sánchez-Yustos (2021) is correct in his emphasis on retouch, the only area in which DK fails to meet our wet season predictions is for its limited transport distances, which may simply have been due to the abundance of appropriate material in the immediate vicinity of the site, making the need for transport of material into it unnecessary. Flake production at FLK-N differs from DK in its increased use of quartz material that is not immediately local, a pattern shared with FLK-Zinj and DS. This may be due to differences in activity with DK that emphasised a sharper edge (cf. Key et al., 2020)—although this almost certainly was not animal exploitation (Domínguez-Rodrigo & Barba, 2007b)—or the success of caching quartz material in this area of the gorge (cf. Potts, 1984, 1991).
Even leaving aside retouch, Fejej and DS clearly present a very different pattern of reduction to DK and FLK-N I-II, in that there is likely a much higher percentage of unifacial cores at each, alongside limited core productivity (Barsky et al., 2011; de Lumley et al., 2004; Diez-Martín et al., 2021). This allies them closely with FLK-Zinj and the two main units of HWK-EE (Fig. 4). These findings are highly consistent with our predictions for dry season sites and are backed up by palaeoecological data at each. DS also shares with FLK-Zinj clear evidence for primary access to animal tissue (Cobo-Sánchez, 2020), alongside PTK, another locality on the same palaeosurface (Diez-Martín et al., 2021, 2022; Domínguez-Rodrigo et al., 2019a; Organista et al., 2023). Phytolith analysis suggesting that all three sites possessed woodland cover at the time of hominin activity (Arráiz et al., 2017), with pollen retrieved from the sediments of Fejej FJ-1a similarly indicating the presence of wooded areas in an otherwise open, grassy landscape (Barsky et al., 2011). Such cover may have made the sites attractive ambush spots for Oldowan hominins (Bunn & Gurtov, 2014; Bunn & Pickering, 2010; Cobo-Sánchez, 2020), with lithic production at these locations servicing the acquisition of carcasses.
The importance of habitat type across the landscape does raise the question of alternative variables that may explain the seasonal patterning of Oldowan lithic investment. In particular, the Olduvai sites we have discussed here cover a relatively large geographical area with a mosaic ecosystem (Uribelarrea et al., 2014), and span swathes of time, from Lower Bed I to Lower Bed II, which incorporates at least one period of major landscape reconfiguration alongside technological change (Bibi et al., 2018; Diez-Martín et al., 2015; Uribelarrea et al., 2017). While we discussed the potential role of woodland environments in dry season animal exploitation, the evidence of animal exploitation at HWK-EE is associated with greater grassland availability (Bibi et al., 2018), although there is still evidence of at least some tree cover in the vicinity of the site (de la Torre et al., 2018; Prassack et al., 2018). We would emphasise that the extent of hominin interaction with bone is much greater at FLK-Zinj and DS than either unit at HWK-EE, but that the existence of shared behaviours between the sites in spite of differences in time and palaeoenvironment underline the importance of seasonality in structuring hominin behaviour.
Similarly, FLK-N is likely to have been located within a forested area (Barboni et al., 2010; Blumenschine et al., 2012), despite its qualitative lithic similarity to Kanjera South, which represents a more open environment (Ferraro et al., 2013; Oliver et al., 2019), but a shared wet season reconstruction. At the same time, the two sites are distinguished by their frequency of percussive behaviours (cf. Diez-Martín et al., 2010), suggesting a different focus on certain subsistence strategies, and perhaps an extent of intra-seasonal variation in habitat usage—tree cover may have been important for access to seeds and nuts. In sum, we would argue that preferred habitat use varied in relation to predictable fluctuations in the spatiotemporal distribution of resources throughout the year, and therefore the spatial selection of habitat types is itself intrinsically linked to the role of seasonality (cf. Linares-Matás & Clark, 2022; Pennec et al., 2020). These findings fit well with the growing number of authors who are recognising the importance of seasonality in hominin behaviour (e.g. Rivals et al., 2009, 2018; Hosfield, 2020; Linares-Matás & Clark, 2022; Descamps et al., 2022). Future research may be able to better integrate these findings with cultural and biological affinities in the palaeoanthropological record, with regard to the identity (or identities) of the makers of the Oldowan (cf. Plummer et al., 2023).
Intra-seasonal Variation in Resource Availability
As highlighted by the discussion of habitat usage throughout the year, it is important to note that we should not expect resource consumption to be completely homogenous within individual seasons, and therefore that stone tool investment may also vary on smaller temporal scales; most of which we are unlikely to have the resolution to identify in the archaeological record. This can be underlined by the fact that the relative availability and therefore importance of berries, tubers, and honey to the Hadza vary quite drastically within the wet season itself, even resulting in a frequent movement of basecamps (Hawkes et al., 1997). Furthermore, while most of the dry season sites discussed in this paper have so far been united by evidence of early access to animal remains, this is unlikely to have been the preferred subsistence strategy for hominins throughout the dry season. This is due to a number of reasons, including the greater ease of access to meat in the second half of the dry season (e.g. Hawkes et al., 1991; O’Connell et al., 1988, 2002), changes in the quality and desirability of animal tissue throughout the season (Sinclar, 1975; Speth, 1987; Bunn & Ezzo, 1993; Cordain et al., 2000), and inter-annual fluctuations in both of these factors (e.g. Hawkes et al., 2001).
Among the sites discussed here, DS does appear to have a much higher proportion of retouched flakes compared to FLK-Zinj (Diez-Martín et al., 2021) that may hint at some economic differences between the sites. This may also be identified in the differential use of raw material, in which quartz flakes were preferentially transported into the site or quartz cores out of it, while basalt and phonolite flakes were transported out of the site after knapping, with only 13.5% of the volcanic flakes expected from flake scars actually recovered (Diez-Martín et al., 2021). This suggests selection of durable (cf. Key et al., 2020) material to use in activities elsewhere in the landscape, and plays into a wider picture in which lava materials played a greater role in the formation of the DS assemblage compared to FLK-Zinj (Diez-Martín et al., 2021). Further still, DS displays a relatively high proportion of material that can be related to percussive activities (4.93%), in clear distinction to the 1.2% at FLK-Zinj (Table 3), and the distribution of percussive (and bipolar) materials at the site is spatially distinct to that related to handheld knapping (Diez-Martín et al., 2021). By extension, this makes the spatial distribution of percussive activities distinct to the distribution of animal remains (Cobo-Sánchez, 2020). It is unclear if these distinct spatial distributions are the result of a segregation of contemporaneous activities within the same site, or temporally-distinct activities in a relatively short timespan. However, it is notable that FLK-Zinj and PTK both show single-cluster accumulations of lithics and faunal remains (Domínguez-Rodrigo & Cobo-Sánchez, 2017), in contrast to the three identified at DS (Cobo-Sánchez, 2020; Diez-Martín et al., 2021).
The combined emphasis on retouch and focus on percussive activities, alongside the underrepresentation of lava flakes, nonetheless allies DS with HWK-EE LAS (Arroyo & de la Torre, 2018; de la Torre & Mora, 2018; Diez-Martín et al., 2021). As such, it is no surprise they formed a distinct clade in each of our cluster analyses, with and without retouch (Fig. 4). If HWK-EE LAS is indeed an occupation during the late dry season (Bibi et al., 2018; Linares-Matás & Clark, 2022; Pante et al., 2018; Rivals et al., 2018), we may thus consider the possibility that the occupation of DS extended similarly towards the end of the dry season. This would hint at intra-seasonal variation in behavioural activities when compared to FLK-Zinj, which may have been abandoned earlier in the season. Domínguez-Rodrigo et al. (2019b) estimate that FLK-Zinj may have been occupied for 2–4 months, leaving scope for such an abandonment of the site before the season had concluded. We do not currently have the data to test the potential for an occupation of DS that continued into the late dry season, with crocodile and fish remains not reported thus far, for example, while the limited number of hippopotamid remains is perhaps unrelated to hominin activity (Cobo-Sánchez, 2020). Future data may allow this hypothesis to be tested empirically.
Intra-seasonal variation in hominin behaviour may also be displayed between the two units of HWK-EE. At the site, both the Lemuta and LAS units preserve the presence of reduncine bovids and the co-occurrence of crocodile and hippopotamids, although the number of shed crocodile teeth and the concentration of fish remains is greater in LAS (Bibi et al., 2018; de la Torre et al., 2018; Pante et al., 2018; Rivals et al., 2018). This is what allowed the inference of a late dry season occupation for LAS (Linares-Matás & Clark, 2022; Pante et al., 2018; Rivals et al., 2018). Furthermore, Lemuta shows a reduced browsing signature in the tooth-wear of Antidorcas recki and Alcelaphini (Bibi et al., 2018; de la Torre et al., 2018; Pante et al., 2018; Rivals et al., 2018), which suggests a greater availability of grasses in the Lemuta member. This pattern may be expected for a wet season occupation of Lemuta (Rivals et al., 2018: 211), but can also be explained by differences in timing of occupation within the dry season, and/or a reduced availability of dicots, which would force these bovids to increase their reliance on grazing (Rivals et al., 2018: 211–212). Indeed, the relationship between the standard deviation and coefficient of variation for the number of micro-wear scratches fails to separate LAS from Lemuta, with individual taxa from the different units instead clustering together (Rivals et al., 2018). This suggests that both units are palimpsestic (although perhaps moreso in the Lemuta; Hay, 1976), with different taxa entering the record at different times and for different durations, with a much shorter duration (perhaps a single season) for equids. Furthermore, there is also a greater incidence of both carnivore tooth marks and hominin percussion marks on midshaft fragments of all animal sizes in Lemuta, 37.4% and 45.7% of size 1–2 and size 3–4 bones, respectively—with 12.6% and 23.5% percussion marks—compared to the respective values of 33.7%, 25%, 6.5%, and 16.7% in LAS (Pante et al., 2018). While the degree of bone marrow and grease avoidance in LAS points to its occupation in the late dry (Pante et al., 2018), these figures highlight intense carnivore activity in LAS, alongside indications that animal tissue was an important component of the hominin diet during this period at the site.
As such, the existing data do not rule out a predominantly dry season occupation for both stratigraphic units, and are equally consistent with occupations during the same season but with (a) inter-annual fluctuations in grass availability, and/or (b) different (but perhaps overlapping) sub-periods of hominin presence. Indeed, such a conclusion may explain the overall similarities in the lithic assemblages, if animal tissue was the driving factor behind the need for sharp edges throughout the dry season, even when it was at reduced attractiveness towards the end. The occupation of LAS also took place in the context of increased forest cover compared to Lemuta (de la Torre et al., 2018), with phytoliths indicating the presence of species including the wild date palm, Phoenix reclinata (Albert et al., 2009). This species produces attractive fruits that are consumed by chimpanzees at a number of sites, including at Mahale, Tanzania, where they also use additional parts of the trees to produce probes for ant fishing (Shimada, 2018). At Bulindi, Uganda, they are available almost year-round and are eagerly consumed when available (McLennan, 2013), representing a core preferred resource (sensu Clark & Linares-Matás, 2023). This is just one such resource which may have been available to hominins towards the end of the dry season to compensate for reduced animal attractiveness. The wide array of pounding tools in HWK-EE LAS, in the context of reduced access to animal tissue compared to Lemuta, may therefore be one mechanism by which hominins compensated for the declining fat content of animal resources at the end of the dry season (Arroyo & de la Torre, 2018; Pante et al., 2018). As at FLK-N, these tools are likely to have been used to process resources other than animal tissue, such as seeds and nuts (Linares-Matás & Clark, 2022), and indeed palm trees were also located at this site (Barboni et al., 2010). As we have pointed out elsewhere, seasonal transitions may have been particularly important in percussive behaviours, as seed and nut availability is assumed to be highest during these periods as fruits ripen and rot (Eaton et al., 1997; Hitchcock et al., 1996; Lee, 2013; Linares-Matás & Clark, 2022; Lombard & Kyriacou, 2018; Testart, 1982).
Outside the role of percussive technologies, the lithic assemblage is relatively consistent between the two units, with the predominance of low-productivity cores giving an overall impression of expediency (de la Torre & Mora, 2018). This accounts for their relatively close association in both cluster analyses (Fig. 4). As outlined in Table 3, there is, however, a non-negligible reduction in the proportion of unifacial cores in the Lemuta member, hinting at greater core productivity and investment in flake production when compared to LAS. This is also suggested by the reduced thickness of flakes compared to their maximum dimension, and the increased number of flake scars per core of the same size. This is perhaps related to the increased use of materials not directly available in the vicinity of the site (de la Torre & Mora, 2018). As discussed with reference to DS, LAS clearly showed a greater need for retouched edges than the Lemuta member, which may hint at a later dry season role for retouched flakes. Given the data discussed here, it is plausible that at least some of the inter-assemblage variance at HWK-EE can be accounted for by intra-seasonal differences in subsistence, from a shared technological competence. It also highlights the importance of retouched flakes in analyses of Oldowan variability, and justifies their inclusion in our analysis.
Seasonal Mobility and Landscape Knowledge
While our conclusions point to different patterns of seasonality at individual Olduvai sites, they may also hint at a wider trend of Oldowan landscape use, with the older DK and FLK-N giving a greater wet season signature than the younger, likely dry season, sites of FLK-Zinj, DS, HWK-EE Lemuta, and HWK-EE LAS. In order to examine if this pattern may be a reflection of hominin behaviour, we can turn to Ewass Oldupa, which presents evidence of hominin presence in the basin as far back as 2.036 Ma (Mercader et al., 2021). The site was occupied sequentially over 235,000 years, in the context of changing environmental conditions, by hominins with a relatively stable toolkit (Cueva-Temprana et al., 2022; Mercader et al., 2021). The lithic assemblage from the site includes an intermediate proportion of unifacial cores and average number of flake scars per core, controlling for log corrected size, although flakes are the thickest, on average, relative to maximum dimension of any site summarised in Table 3, and retouch is uncommon at 2.18–2.5%. These data do not fit comfortably into our predictions for lithic investment in either the wet or dry seasons, which explains why in our cluster analysis, it aligned most closely once with DK and once with the East Gona sites, as an outgroup to all other assemblages. Nonetheless, it is particularly interesting that some of the quartz present at the site was transported around 12 km, suggesting an unusual level of mobility for Olduvai hominins (Mercader et al., 2021), that instead allies the site with transport at Kanjera South (Braun et al., 2008a, 2008b). Patalano et al., (2021: 9–10) argue that the nature of Oldowan tool use at sites such as Ewass Oldupa is suggestive of:
“technological adaptation to major geomorphic and ecological transitions, whereby stone tool use permitted provisioning across ecologically diverse and complex environments over time and space. Tool use allowed for a more generalist strategy in acquiring plant food resources within a rapidly changing plant landscape that ranged from fern meadows to woodland mosaics, naturally burned landscapes, lakeside woodland/palm groves, and hyper-xeric steppes.”
From the perspective of our seasonality model, we would expect such mobility and stone tool-mediated exploitation of diverse plant resources to be predominantly related to repeated visits to the site in the context of wet season foraging strategies, and the ability to plan the deployment of stone tools throughout the landscape. Indeed, as predicted in Table 1, evidence for animal exploitation at the site is extremely limited, with just two cut-marked specimens (including the ventral side of a rib, consistent with evisceration; Nilssen, 2000) and twelve percussion marks (0.3% of the assemblage; Akuku et al., 2022). Alongside this limited anthropogenic interaction is restricted carnivore access to the remains, with just 2% of the assemblage exhibiting tooth marks. Some extent of hominin-carnivore competition is nonetheless confirmed by the presence of a single bone with adjacent cut marks and tooth scoring (Akuku et al., 2022). Beyond this, the palimpsestic nature of the faunal assemblage and the lack of evidence for a coeval hominin presence at the site mean we can only speculate at palaeoecological signatures of seasonality. For example, the presence of a single reduncine bovid bone, alongside both hippopotamid (albeit extremely rare) and crocodile, leaves open the possibility for dry season input, while an extent of wet season accumulation is suggested by the presence of chelonids at the site (Akuku et al., 2022; Mercader et al., 2021).
If hominin visits to the site were biased towards the wet season it may explain the unexpectedly high transport distances and overall consistency in stone tool use over time, but it does not explain the intermediate level of investment in reduction (although with a heavy emphasis on multifacial schemes; Cueva-Temprana et al., 2022). We speculate that the answer to such a discrepancy may be found in the interplay between factors governing investment at distinct temporal scales. As outlined for animal resources, technological investment is intimately tied to knowledge of the landscape(s) that populations are inhabiting (including seasonal changes in resource availability), and that this only accumulates over much longer swathes of time (Clark & Linares-Matás, 2020, 2023). If a population is new to a landscape, their knowledge of the distribution and predictability of resources will be restricted (Rockman & Steele, 2003), constraining the ability to plan the spatial and temporal deployment of stone tools across the landscape and limiting returns on investment. It is only through longer-term interaction with a landscape that returns on investment become more predictable, and when we would therefore expect predicted patterns of wet season lithic investment to fully emerge.
Nonetheless, it makes sense that movement into novel environments would have taken place during the wet season, as this is when modern human hunter-gatherer groups (a) display greater mobility between base camps and resource patches (Hawkes et al., 1997; Lee, 2013) and (b) have greater food security (as a direct result of its predictability), which is associated with a greater likelihood of experimentation with new resources (Lee, 2013). This makes the potential pattern of early Olduvai sites (DK and FLK-N, and potentially Ewass Oldupa) being occupied during the wet season particularly notable, as it would suggest gradual adaptation to predictable (and low competition) plant resources before accumulation of sufficient knowledge (particularly of animal exploitation) to survive in the much riskier dry seasons at FLK-Zinj, DS, and HWK-EE. At a more general level, the limited number of Oldowan sites with clear primary access to animal remains (most of which are at Olduvai) may suggest that carcass acquisition was often constrained to very specific points in the landscape that provided unusually high returns (Clark & Linares-Matás, 2023). Such a process would require intense familiarity with the landscape and the behaviour of herbivores and carnivores within it.
For the inhabitants of Ewass Oldupa, we can expect early knowledge of the Olduvai landscape to have initially been low, with the ability to invest in the toolkit relatively limited due to the uncertainty of return. In such contexts, generalist toolkits would have been particularly important in mediating often dramatic habitat and micro-habitat variation (Patalano et al., 2021). It is only when landscape knowledge became more consolidated for Olduvai that we would predict greater investment in stone tool reduction. The stone tool sample may be too small to identify changes over time between the Ewass Oldupa strata, but it is clear that hominins were experimenting with novel and more complex ways of manufacturing stone tools by the time of DK (Sánchez-Yustos, 2021). This may even include important transitional steps towards the later development of Acheulean technologies, including large flake production (with some of > 10 cm) and their subsequent secondary flaking into cores and large tools. In this context, it is also worth noting that the second analysis of Oldowan and early Acheulean sites by Braun et al. (2019) found that DK fell alongside early Acheulean sites, rather than the earlier or later Oldowan clusters. This may be suggestive of its transitional character, and opens up the potential for wet season drivers of lithic investment to be involved in the innovation of early Acheulean artefacts, in the context of longer-term landscape habituation (cf. Davis & Ashton, 2019).
Conclusions
With the literature presented here, we have attempted to consolidate our framework for identifying the season of site occupation in the Early Pleistocene archaeological record of eastern Africa (Linares-Matás & Clark, 2022). Our main aim was to develop a predictive characterisation of technological investment in relation to seasonal ecological signatures, which would enable us to develop a more comprehensive and attribute-based seasonality framework for Oldowan lithic assemblages. In particular, we emphasise and expand upon previous scholarship suggesting that visits to Olduvai DK were biased towards the wet season (Speth & Davies, 1976), perhaps alongside FLK-N I-II, while DS is likely to have been occupied during the dry season (Cobo-Sánchez, 2020; Diez-Martín et al., 2021; Domínguez-Rodrigo et al., 2019a). Fejej Fj-1a may similarly encompass dry season activities.
We argue that the lithic assemblages from these sites fit into our predictions derived from Kanjera South and FLK-Zinj (Linares-Matás & Clark, 2022) that sites with hominin presence in the wet season should show a greater degree of technological investment when compared to those deriving from dry season activity. In our analyses, the inclusion of retouch produced two distinct technological clusters that would map directly onto our predictions for wet and dry season investment. Indeed, DS and HWK-EE LAS highlight the importance of retouch for understanding inter-assemblage variability, and thus it should be considered an integral component of comparisons between Oldowan sites. The 2.6–2.5 Ma sites of the Gona palaeolandscape may also fit into our broader framework, with OGS7 clustering with predicted wet season assemblages, but data to test these hypotheses directly remain limited.
Based on our findings, we strongly argue that yearly seasonality constitutes a relevant factor to consider when evaluating variation in archaeological sites in the Early Stone Age. This emphasis does not detract from other drivers of variation, but rather highlights that seasonality interacts with a number of other more proximate pressures, such as ecological context, raw material use, and foraging strategies. These pressures can be considered alongside environmental constraints at broader scales, from inter-annual changes to resource availability (Hawkes et al., 1991; Speth et al., 1991) up to longer-term climatic variability (Potts, 1996a,b, 1998; 2012; Potts & Faith, 2015). Indeed, the character of Olduvai DK as transitional to the Acheulean (Sánchez-Yustos, 2021) highlights the role of seasonal investment within longer term trends of innovation and change in the archaeological record.
We also wish to emphasise that seasonality is crucial to consider when studying hominin adaptation to specific landscapes (Clark & Linares-Matás, 2020, 2023), as we have discussed in relation to Ewass Oldupa. We predict that initial movements into novel landscapes are most likely to take place during periods of high mobility and high biomass availability across the landscape, both of which are more limited during the dry season when plant and animal resources are concentrated around permanent water sources and camps become larger and occupied during several months at a time (e.g. Bartram et al., 1991; Domínguez-Rodrigo et al., 2019b; Hawkes et al., 1991, 1997; Lee, 2013). The occupation of specific landscapes throughout the year was dependent on hominins developing a substantial awareness of spatio-temporal variation in resource availability and carnivore competition. As such, we argue that movement into novel landscapes should initially be associated with reduced investment as hominins employed generalist toolkits (cf. Patalano et al., 2021; Shea, 2010) to adapt to novel resources of which they only possessed limited knowledge. The consolidation of landscape knowledge would have ensured more predictable returns on seasonally mediated technological investment (Clark & Linares-Matás, 2020).
Data Availability
The authors confirm that all data generated or analysed during this study are included in this published article or supplementary information.
References
Akuku, P., Saladié, P., Mohamed, A., Mwambwiga, A., Bushozi, P., & Mercader, J. (2022). Faunal assemblages from Lower Bed I (Oldupai Gorge, Tanzania). Frontiers in Ecology and Evolution, 10, 895305. https://doi.org/10.3389/fevo.2022.895305
Albert, R. M., Bamford, M. K., & Cabanes, D. (2009). Palaeoecological significance of palms at Olduvai Gorge, Tanzania, based on phytolith remains. Quaternary International, 193(1–2), 41–48. https://doi.org/10.1016/j.quaint.2007.06.008
Arráiz, H., Barboni, D., Ashley, G. M., Mabulla, A., Baquedano, E., & Domínguez-Rodrigo, M. (2017). The FLK Zinj paleolandscape: Reconstruction of a 1.84Ma wooded habitat in the FLK Zinj-AMK-PTK-DS archaeological complex, Middle Bed I (Olduvai Gorge, Tanzania). Palaeogeography, Palaeoclimatology, Palaeoecology, 488, 9–20. https://doi.org/10.1016/j.palaeo.2017.04.025
Arroyo, A., & de la Torre, I. (2018). Pounding tools in HWK EE and EF-HR (Olduvai Gorge, Tanzania): Percussive activities in the Oldowan-Acheulean transition. Journal of Human Evolution, 120, 402–421. https://doi.org/10.1016/j.jhevol.2017.10.005
Ashley, G. M., Barboni, D., Dominguez-Rodrigo, M., Bunn, H. T., Mabulla, A. Z. P., Diez-Martin, F., Barba, R., & Baquedano, E. (2010). A spring and wooded habitat at FLK Zinj and their relevance to origins of human behavior. Quaternary Research, 74(3), 304–314. https://doi.org/10.1016/j.yqres.2010.07.015
Auffenberg, W. (1981). The fossil turtles of Olduvai Gorge, Tanzania. Africa. Copeia, 1981(3), 509–522. https://doi.org/10.2307/1444555
Baenninger, R., Estes, R. D., & Baldwin, S. (1977). Anti-predator behaviour of baboons and impalas toward a cheetah. African Journal of Ecology, 15, 327–329.
Baldwin, D. S., Rees, G. N., Wilson, J. S., Colloff, M. J., Whitworth, K. L., Pitman, T. L., & Wallace, T. A. (2013). Provisioning of bioavailable carbon between the wet and dry phases in a semi-arid floodplain. Oecologia, 172(2), 539–550. https://doi.org/10.1007/s00442-012-2512-8
Barba, R., & Domínguez-Rodrigo, M. (2007). FLK North North 1: “Living floor” or natural accumulation? In M. Domínguez-Rodrigo, R. Barba, & C. P. Egeland (Eds.), Deconstructing Olduvai: A taphonomic study of the Bed I sites (pp. 217–228). Springer.
Barboni, D., Ashley, G. M., Dominguez-Rodrigo, M., Bunn, H. T., Mabulla, A. Z. P., & Baquedano, E. (2010). Phytoliths infer locally dense and heterogeneous paleovegetation at FLK North and surrounding localities during upper Bed I time, Olduvai Gorge. Tanzania. Quaternary Research, 74(3), 344–354. https://doi.org/10.1016/j.yqres.2010.09.005
Barsky, D., Chapon-Sao, C., Bahain, J.-J., Beyene, Y., Cauche, D., Celiberti, V., Desclaux, E., de Lumley, H., de Lumley, M.-A., Marchal, F., Moullé, P.-E., & Pleurdeau, D. (2011). The Early Oldowan stone-tool assemblage from Fejej FJ-1A, Ethiopia. Journal of African Archaeology, 9, 207–224. https://doi.org/10.3213/2191-5784-10196
Bartlam-Brooks, H. L. A., Beck, P. S. A., Bohrer, G., & Harris, S. (2013a). In search of greener pastures: Using satellite images to predict the effects of environmental change on zebra migration. Journal of Geophysical Research: Biogeosciences, 118(4), 1427–1437. https://doi.org/10.1002/jgrg.20096
Bartlam-Brooks, H. L. A., Bonyongo, M. C., & Harris, S. (2013b). How landscape scale changes affect ecological processes in conservation areas: External factors influence land use by zebra (Equus burchelli) in the Okavango Delta. Ecology and Evolution, 3(9), 2795–2805. https://doi.org/10.1002/ece3.676
Bartram, L. E., Kroll, E. M., & Bunn, H. T. (1991). Variability in camp structure and bone food refuse patterning at Kua San Hunter-Gatherer Camps. In E. M. Kroll & T. D. Price (Eds.), The interpretation of archaeological spatial patterning (pp. 77–148). Plenum Press (NY).
Bibi, F., Pante, M., Souron, A., Stewart, K., Varela, S., Werdelin, L., Boisserie, J.-R., Fortelius, M., Hlusko, L., Njau, J., & de la Torre, I. (2018). Paleoecology of the Serengeti during the Oldowan-Acheulean transition at Olduvai Gorge, Tanzania: The mammal and fish evidence. Journal of Human Evolution, 120, 48–75. https://doi.org/10.1016/j.jhevol.2017.10.009
Blumenschine, R. J. (1987). Characteristics of an early hominid scavenging niche. Current Anthropology, 28(4), 383–407.
Blumenschine, R. J., Masao, F. T., Tactikos, J. C., & Ebert, J. I. (2008). Effects of distance from stone source on landscape-scale variation in Oldowan artifact assemblages in the Paleo-Olduvai Basin Tanzania. Journal of Archaeological Science, 35(1), 76–86. https://doi.org/10.1016/j.jas.2007.02.009
Blumenschine, R. J., Stanistreet, I. G., Njau, J. K., Bamford, M. K., Masao, F. T., Albert, R. M., Stollhofen, H., Andrews, P., Prassack, K. A., McHenry, L. J., Fernández-Jalvo, Y., Camilli, E. L., & Ebert, J. I. (2012). Environments and hominin activities across the FLK Peninsula during Zinjanthropus times (1.84 Ma), Olduvai Gorge Tanzania. Journal of Human Evolution, 63(2), 364–383. https://doi.org/10.1016/j.jhevol.2011.10.001
Blumenthal, S. A., Cerling, T. E., Smiley, T. M., Badgley, C. E., & Plummer, T. W. (2019). Isotopic records of climate seasonality in equid teeth. Geochimica Et Cosmochimica Acta, 260, 329–348. https://doi.org/10.1016/j.gca.2019.06.037
Braun, D. R., Plummer, T. W., Ditchfield, P., Ferraro, J. V., Maina, D., Bishop, L. C., & Potts, R. (2008a). Oldowan behavior and raw material transport: Perspectives from the Kanjera Formation. Journal of Archaeological Science, 35(8), 2329–2345. https://doi.org/10.1016/j.jas.2008.03.004
Braun, D. R., Rogers, M. J., Harris, J. W. K., & Walker, S. J. (2008b). Landscape-scale variation in hominin tool use: Evidence from the developed Oldowan. Journal of Human Evolution, 55(6), 1053–1063. https://doi.org/10.1016/j.jhevol.2008.05.020
Braun, D. R., Plummer, T., Ferraro, J. V., Ditchfield, P., & Bishop, L. C. (2009b). Raw material quality and Oldowan hominin toolstone preferences: Evidence from Kanjera South, Kenya. Journal of Archaeological Science, 36(7), 1605–1614. https://doi.org/10.1016/j.jas.2009.03.025
Braun, D. R., Harris, J. W. K., Levin, N. E., McCoy, J. T., Herries, A. I. R., Bamford, M. K., Bishop, L. C., Richmond, B. G., & Kibunjia, M. (2010). Early hominin diet included diverse terrestrial and aquatic animals 1.95 Ma in East Turkana, Kenya. Proceedings of the National Academy of Sciences, 107(22), 10002–10007. https://doi.org/10.1073/pnas.1002181107
Braun, D. R., Plummer, T. W., Ditchfield, P. W., Bishop, L. C., & Ferraro, J. V. (2009a). Oldowan technology and raw material variability at Kanjera South. In E. Hovers, & D. R. Braun (Eds.), Interdisciplinary approaches to the Oldowan (pp. 99–110). Springer Netherlands. https://doi.org/10.1007/978-1-4020-9060-8_9
Braun, D. R., Aldeias, V., Archer, W., Arrowsmith, J. R., Baraki, N., Campisano, C. J., Deino, A. L., DiMaggio, E. N., Dupont-Nivet, G., Engda, B., Feary, D. A., Garello, D. I., Kerfelew, Z., McPherron, S. P., Patterson, D. B., Reeves, J. S., Thompson, J. C., & Reed, K. E. (2019). Earliest known Oldowan artifacts at >2.58 Ma from Ledi-Geraru, Ethiopia, highlight early technological diversity. Proceedings of the National Acadedmy of Sciences, 116, 11712–11717. https://doi.org/10.1073/pnas.1820177116
Bunn, H. T., & Ezzo, J. A. (1993). Hunting and scavenging by Plio-Pleistocene hominids: Nutritional constraints, archaeological patterns, and behavioural implications. Journal of Archaeological Science, 20(4), 365–398. https://doi.org/10.1006/jasc.1993.1023
Bunn, H. T., & Gurtov, A. N. (2014). Prey mortality profiles indicate that Early Pleistocene Homo at Olduvai was an ambush predator. Quaternary International, 322–323, 44–53. https://doi.org/10.1016/j.quaint.2013.11.002
Bunn, H. T., & Pickering, T. R. (2010). Bovid mortality profiles in paleoecological context falsify hypotheses of endurance running–hunting and passive scavenging by early Pleistocene hominins. Quaternary Research, 74(3), 395–404. https://doi.org/10.1016/j.yqres.2010.07.012
Bunn, H. T., Bartram, L. E., & Kroll, E. M. (1988). Variability in bone assemblage formation from Hadza hunting, scavenging, and carcass processing. Journal of Anthropological Archaeology, 7(4), 412–457. https://doi.org/10.1016/0278-4165(88)90004-9
Cáceres, I., Rogers, M.J., Semaw, S. & Leiss, A.C. (2015). Hominin exploitation of animal resources at the Gona Plio-Pleistocene archeological sites (Afar, Ethiopia). II Jornadas de Prehistoria Africana, 2015, 27. http://cir.cenieh.es/handle/20.500.12136/1329. Accessed 31 Jan 2023.
Cáceres, I., Rogers, M. J., Semaw, S., & Leiss, A. C. (2017). Crocodiles in the diet of the first Oldowan toolmakers: evidence from OGS-6 at Gona (Afar, Ethiopia). III Jornadas de Prehistoria Africana, 2017, 20. https://cir.cenieh.es/handle/20.500.12136/944
Chergui, B., Rodríguez-Caro, R. C., Graciá, E., Fahd, S., & Santos, X. (2019). Population density of the spur-thighed tortoise Testudo graeca declines after fire in north-western Africa. Plos One, 14, e0220969. https://doi.org/10.1371/journal.pone.0220969
Clark, J., & Linares-Matás, G. (2023). Seasonal resource categorisation and behavioral adaptation among chimpanzees: Implications for early hominin carnivory. Journal of Anthropological Sciences, 101, 1–35. https://doi.org/10.4436/JASS.10006
Clark, J., & Linares-Matás, G. (2020). The role of landscape knowledge networks in the Early Pleistocene technological variability of East Africa. Archaeological Review from Cambridge, 35(2), 25–44. https://doi.org/10.17863/CAM.71847
Clarkson, C., Haslam, M., & Harris, C. (2015). When to retouch, haft, or discard? Modeling optimal use/maintenance schedules in lithic tool use. In N. Goodale & W. Andrefsky Jr. (Eds.), Lithic Technological Systems and Evolutionary Theory (pp. 117–138). Cambridge University Press. https://doi.org/10.1017/CBO9781139207775.011
Colloff, M. J., & Baldwin, D. S. (2010). Resilience of floodplain ecosystems in a semi-arid environment. The Rangeland Journal, 32(3), 305. https://doi.org/10.1071/RJ10015
Cobo-Sánchez, L. (2020). Taphonomic and spatial study of the archaeological site DS from Bed I in Olduvai Gorge (Tanzania). Unpublished PhD Dissertation, Universidad Complutense de Madrid, Madrid.
Cordain, L., Miller, J. B., Eaton, S. B., Mann, N., Holt, S. H., & Speth, J. D. (2000). Plant-animal subsistence ratios and macronutrient energy estimations in worldwide hunter-gatherer diets. The American Journal of Clinical Nutrition, 71(3), 682–692. https://doi.org/10.1093/ajcn/71.3.682
Couturier, T., Besnard, A., Bertolero, A., Bosc, V., Astruc, G., & Cheylan, M. (2014). Factors determining the abundance and occurrence of Hermann’s tortoise Testudo hermanni in France and Spain: Fire regime and landscape changes as the main drivers. Biological Conservation, 170, 177–187. https://doi.org/10.1016/j.biocon.2013.12.028
Cueva-Temprana, A., Lombao, D., Soto, M., Itambu, M., Bushozi, P., Boivin, N., Petraglia, M., & Mercader, J. (2022). Oldowan technology amid shifting environments ∼2.03–1.83 million years ago. Frontiers in Ecology and Evolution, 10, 788101. https://doi.org/10.3389/fevo.2022.788101
Davis, R., & Ashton, N. (2019). Landscapes, environments and societies: The development of culture in Lower Palaeolithic Europe. Journal of Anthropological Archaeology, 56, 101107. https://doi.org/10.1016/j.jaa.2019.101107
de la Torre, I., & Mora, R. (2005). Technological strategies in the Lower Pleistocene at Olduvai Beds I & II. Universite de Liége.
de la Torre, I., & Mora, R. (2018). Oldowan technological behaviour at HWK EE (Olduvai Gorge, Tanzania). Journal of Human Evolution, 120, 236–273. https://doi.org/10.1016/j.jhevol.2018.04.001
de la Torre, I., Albert, R. M., Arroyo, A., Macphail, R., McHenry, L. J., Mora, R., Njau, J. K., Pante, M. C., Rivera-Rondón, C. A., Rodríguez-Cintas, Á., Stanistreet, I. G., Stollhofen, H., & Wehr, K. (2018). New excavations at the HWK EE site: Archaeology, paleoenvironment and site formation processes during late Oldowan times at Olduvai Gorge, Tanzania. Journal of Human Evolution, 120, 140–202. https://doi.org/10.1016/j.jhevol.2017.07.018
de Lumley, H., Barsky, D., Moncel, M. H., Carbonell, E., Cauche, D., Celiberti, V., Notter, O., Pleurdeau, D., Hong, M.-Y., Rogers, M. J., & Semaw, S. (2018). The first technical sequences in human evolution from East Gona, Afar region Ethiopia. Antiquity, 92(365), 1151–1164. https://doi.org/10.15184/aqy.2018.169
de Lumley, H., & Beyene, Y. (Eds.). (2004). Les sites préhistoriques de la région de Fejej, Sud-Omo, Ethiopie, dans leur contexte stratigraphique et paléontologique. Paris: Association pour la diffusion de la pensée française (ADPF), Editions Recherche sur les civilisations.
de Lumley, H., Beyene, Y., Barsky, D., Byrne, L., Camara, A., Cauche, D., Celiberti, V., Fournier, A., & Pleurdeau, D. (2004). L'industrie lithique préoldowayenne du site de Fejej FJ-1, Ethiopie. In de Lumley, H. & Beyene, Y. (Eds.), Les sites préhistoriques de la région de Fejej, Sud-Omo, Ethiopie, dans leur contexte stratigraphique et paléontologique (pp. 391–564). Paris: Association pour la diffusion de la pensée française (ADPF), Editions Recherche sur les civilisations
Delagnes, A., & Roche, H. (2005). Late Pliocene hominid knapping skills: The case of Lokalalei 2C, West Turkana Kenya. Journal of Human Evolution, 48(5), 435–472. https://doi.org/10.1016/j.jhevol.2004.12.005
deMenocal, P. B. (1995). Plio-Pleistocene African climate. Science, 270, 53–59. https://doi.org/10.1126/science.270.5233.53
deMenocal, P. B. (2004). African climate change and faunal evolution during the Pliocene-Pleistocene. Earth and Planetary Science Letters, 220, 3–24. https://doi.org/10.1016/S0012-821X(04)00003-2
Dennell, R. (2007). “Resource-rich, stone-poor”: Early hominin land use in large river systems of northern India and Pakistan. In M. D. Petraglia & B. Allchin (Eds.), The evolution and history of human populations in south Asia (pp. 41–68). Dordrecht: Springer Netherlands. https://doi.org/10.1007/1-4020-5562-5_3
Deschamps, M., Martín-Lerma, I., Linares-Matás, G., & Zilhão, J. (2022). Organization of residential space, site function variability, and seasonality of activities among MIS 5 Iberian Neandertals. Scientific Reports, 12, 20221. https://doi.org/10.1038/s41598-022-24430-z
Diez-Martin, F., Sanchez Yustos, P., Domínguez-Rodrigo, M., Mabulla, A. Z. P., Bunn, H. T., Ashley, G. M., Barba, R., & Baquedano, E. (2010). New insights into hominin lithic activities at FLK North Bed I, Olduvai Gorge Tanzania. Quaternary Research, 74(3), 376–387. https://doi.org/10.1016/j.yqres.2010.07.019
Diez-Martín, F., Sánchez Yustos, P., Uribelarrea, D., Baquedano, E., Mark, D. F., Mabulla, A., et al. (2015). The origin of the Acheulean: The 1.7 million-year-old site of FLK West, Olduvai Gorge (Tanzania). Scientific Reports, 5, 17839. https://doi.org/10.1038/srep17839
Diez-Martín, F., Cobo-Sánchez, L., Baddeley, A., Uribelarrea, D., Mabulla, A., Baquedano, E., & Domínguez-Rodrigo, M. (2021). Tracing the spatial imprint of Oldowan technological behaviors: A view from DS (Bed I, Olduvai Gorge, Tanzania). Plos One, 16(7), e0254603. https://doi.org/10.1371/journal.pone.0254603
Diez-Martín, F., Panera, J., Maíllo-Fernández, J. M., Santonja, M., Sánchez-Yustos, P., Pérez-González, A., Duque, J., Rubio, S., Marín, J., Fraile, C., Mabulla, A., Baquedano, E., & Domínguez-Rodrigo, M. (2022). The evolution of stone tool technology at Olduvai Gorge (Tanzania): Contributions from the Olduvai Paleoanthropology and Paleoecology Project. L’anthropologie, 126(1), 103000. https://doi.org/10.1016/j.anthro.2022.103000
Ditchfield, P. W., Whitfield, E., Vincent, T., Plummer, T., Braun, D., Deino, A., Hertel, F., Oliver, J. S., Louys, J., & Bishop, L. C. (2019). Geochronology and physical context of Oldowan site formation at Kanjera South Kenya. Geological Magazine, 156(07), 1190–1200. https://doi.org/10.1017/S0016756818000602
Domínguez-Rodrigo, M., & Barba, R. (2007a). Reanalysis of FLK North North 3: Yet another case of a palimpsest? In M. Domínguez-Rodrigo, R. Barba, & C. P. Egeland (Eds.), Deconstructing Olduvai: A taphonomic study of the bed I sites (pp. 239–252). Dordrecht: Springer Netherlands.
Domínguez-Rodrigo, M., & Cobo-Sánchez, L. (2017). A spatial analysis of stone tools and fossil bones at FLK Zinj 22 and PTK I (Bed I, Olduvai Gorge, Tanzania) and its bearing on the social organization of early humans. Palaeogeography, Palaeoclimatology, Palaeoecology, 488, 21–34. https://doi.org/10.1016/j.palaeo.2017.04.010
Domínguez-Rodrigo, M., Rayne Pickering, T., Semaw, S., & Rogers, M. J. (2005). Cutmarked bones from Pliocene archaeological sites at Gona, Afar, Ethiopia: Implications for the function of the world’s oldest stone tools. Journal of Human Evolution, 48(2), 109–121. https://doi.org/10.1016/j.jhevol.2004.09.004
Domínguez-Rodrigo, M., Bunn, H. T., Mabulla, A. Z. P., Ashley, G. M., Diez-Martin, F., Barboni, D., Prendergast, M. E., Yravedra, J., Barba, R., Sánchez, A., Baquedano, E., & Pickering, T. R. (2010). New excavations at the FLK Zinjanthropus site and its surrounding landscape and their behavioral implications. Quaternary Research, 74(3), 315–332. https://doi.org/10.1016/j.yqres.2010.07.003
Domínguez-Rodrigo, M., Cobo-Sánchez, L., Aramendi, J., & Gidna, A. (2019a). The meta-group social network of early humans: A temporal–spatial assessment of group size at FLK Zinj (Olduvai Gorge, Tanzania). Journal of Human Evolution, 127, 54–66. https://doi.org/10.1016/j.jhevol.2018.11.001
Domínguez-Rodrigo, M., Sánchez-Flores, A. J., Baquedano, E., Carmen Arriaza, M., Aramendi, J., Cobo-Sánchez, L., Organista, E., & Barba, R. (2019b). Constraining time and ecology on the Zinj paleolandscape: Microwear and mesowear analyses of the archaeofaunal remains of FLK Zinj and DS (Bed I), compared to FLK North (Bed I) and BK (Bed II) at Olduvai Gorge (Tanzania). Quaternary International, 526, 4–14. https://doi.org/10.1016/j.quaint.2019.05.041
Domínguez-Rodrigo, M., & Barba., R. (2007b). A palimpsest at FLK North 1–2: Independent carnivore- and hominid-made bone accumulations. In M. Domínguez-Rodrigo, R. Barba, & C. P. Egeland (Eds.), Deconstructing Olduvai: A taphonomic study of the bed I sites (pp. 127–163). Dordrecht: Springer Netherlands.
Domínguez-Rodrigo, M. (2009). Are all Oldowan sites palimpsests? If so, what can they tell us about hominid carnivory? In E. Hovers & D. R. Braun (Eds.), Interdisciplinary approaches to the Oldowan (pp. 129–147). https://doi.org/10.1007/978-1-4020-9060-8_11
Dray, S., & Josse, J. (2015). Principal component analysis with missing values: A comparative survey of methods. Plant Ecology, 216, 657–667. https://doi.org/10.1007/s11258-014-0406-z
Eaton, S., Eaton, S., III., & Konner, M. (1997). Paleolithic nutrition revisited: A twelve-year retrospective on its nature and implications. European Journal of Clinical Nutrition, 51(4), 207–216. https://doi.org/10.1038/sj.ejcn.1600389
Echassoux, A., Moullé, P. E., Desclaux, E., & Alemseged, Z. (2004). Les faunes plio-pléistocènes du site de Fejej FJ-1. In H. de Lumley & Y. Beyene (Eds.), Les sites préhistoriques de la région de Fejej, Sud-Omo, Ethiopie, dans leur contexte stratigraphique et paléontologique (pp. 203–340). Paris: Association pour la diffusion de la pensée française (ADPF), Editions Recherche sur les civilisations.
Egeland, C. P. (2007a). Zooarchaeology and taphonomy of the DK site. In M. Domínguez-Rodrigo, R. B. Egido, & C. P. Egeland (Eds.), Deconstructing Olduvai: A taphonomic study of the bed I sites (pp. 253–268). Dordrecht: Springer Netherlands. https://doi.org/10.1007/978-1-4020-6152-3_15
Egeland, C. P. (2007b). Zooarchaeological and taphonomic perspectives on hominid and carnivore interactions at Olduvai Gorge, Tanzania. Unpublished PhD Dissertation. Bloomington, IN: Indiana University.
Estes, R. D. (1991). The behavior guide to African mammals. University of California Press.
Favreau, J. (2023). Sourcing Oldowan and Acheulean stone tools in Eastern Africa: Aims, methods, challenges, and state of knowledge. Quaternary Science Advances, 9, 100068. https://doi.org/10.1016/j.qsa.2022.100068
Ferraro, J. V., Plummer, T. W., Pobiner, B. L., Oliver, J. S., Bishop, L. C., Braun, D. R., Ditchfield, P. W., Seaman, J. W., III, Binetti, K. M., Seaman, J. W., Jr., Hertel, F., & Potts, R. (2013). Earliest archaeological evidence of persistent hominin carnivory. PLOS One, 8(4), e62174. https://doi.org/10.1371/journal.pone.0062174
Flicker, D., & Key, A. (2023). Statistical assessment of the temporal and cultural relationship between the Lomekwian and Oldowan. Journal of Archaeological Science: Reports, 48, 103834. https://doi.org/10.1016/j.jasrep.2023.103834
Foley, R. A. (1987). Another Unique Species: Patterns in Human Evolutionary Ecology. Harlow: Longman.
Foley, R. A. (1993). The influence of seasonality on hominid evolution. In S. J. Ulijaszek & S. S. Strickland (Eds.), Seasonality and human ecology (pp. 17–37). Cambridge University Press. https://doi.org/10.1017/CBO9780511600517.003
Fritz, U., Branch, W. R., Hofmeyr, M. D., Maran, J., Prokop, H., Schleicher, A., Široký, P., Stuckas, H., Vargas-Ramírez, M., Vences, M., & Hundsdörfer, A. K. (2011). Molecular phylogeny of African hinged and helmeted terrapins (Testudines: Pelomedusidae: Pelusios and Pelomedusa). Zoologica Scripta, 40(2), 115–125. https://doi.org/10.1111/j.1463-6409.2010.00464.x
Galán, A. B., & Domínguez-Rodrigo, M. (2014). Testing the efficiency of simple flakes, retouched flakes and small handaxes during butchery: The efficiency of flakes and small handaxes during butchery. Archaeometry, 56(6), 1054–1074. https://doi.org/10.1111/arcm.12064
Gallotti, R., & Mussi, M. (2015). The unknown Oldowan: ~1.7-million-year-old standardized obsidian small tools from Garba IV, Melka Kunture Ethiopia. Plos One, 10(12), e0145101. https://doi.org/10.1371/journal.pone.0145101
Gallotti, R. (2018). Before the Acheulean in East Africa: An overview of the Oldowan lithic assemblages. In R. Gallotti & M. Mussi (Eds.), The emergence of the Acheulean in East Africa and beyond (pp. 13–32). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-319-75985-2_2
Greenwood, P. H. (1955). Reproduction in the cat-fish, Clarias mossambicus peters. Nature, 176(4480), 516–518. https://doi.org/10.1038/176516b0
Gruber, T., Muller, M. N., Strimling, P., Wrangham, R., & Zuberbühler, K. (2009). Wild chimpanzees rely on cultural knowledge to solve an experimental honey acquisition task. Current Biology, 19(21), 1806–1810. https://doi.org/10.1016/j.cub.2009.08.060
Gurtov, A. N. (2016). Dental microwear texture analysis of early pleistocene hominin prey: A test of foraging seasonality. Unpublished PhD Dissertation. Madison, WI: University of Wisconsin-Madison.
Harmand, S., Lewis, J. E., Feibel, C. S., Lepre, C. J., Prat, S., Lenoble, A., Boës, X., Quinn, R. L., Brenet, M., Arroyo, A., Taylor, N., Clément, S., Daver, G., Brugal, J.-P., Leakey, L., Mortlock, R. A., Wright, J. D., Lokorodi, S., Kirwa, C., et al. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521(7552), 310–315. https://doi.org/10.1038/nature14464
Hawkes, K. (2016). Ethnoarchaeology and Plio-Pleistocene sites: Some lessons from the Hadza. Journal of Anthropological Archaeology, 44, 158–165. https://doi.org/10.1016/j.jaa.2016.07.005
Hawkes, K., O’Connell, J. F., & Blurton-Jones, N. G. (1989). Hardworking Hadza grandmothers. In V. Standen & R. A. Foley (Eds.), Comparative socioecology: The behavioural ecology of humans and other mammals (pp. 341–366). Blackwell Scientific.
Hawkes, K., O’Connell, J. F., Blurton Jones, N. G., Oftedal, O. T., Blumenschine, R. J., Widdowson, E. M., Whiten, A., & Bone, Q. (1991). Hunting income patterns among the Hadza: Big game, common goods, foraging goals and the evolution of the human diet. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 334(1270), 243–251. https://doi.org/10.1098/rstb.1991.0113
Hawkes, K., O’Connell, J. F., & Blurton Jones, N. G. (1997). Hadza women’s time allocation, offspring provisioning, and the evolution of long postmenopausal life spans. Current Anthropology, 38(4), 551–577. https://doi.org/10.1086/204646
Hawkes, K., O’Connell, J. F., & Blurton Jones, N. G. (2001). Hadza meat sharing. Evolution and Human Behavior, 22(2), 113–142. https://doi.org/10.1016/S1090-5138(00)00066-0
Hay, R. L. (1976). Geology of the Olduvai Gorge: A study of sedimentation in a semiarid basin. University of California Press.
Hitchcock, R. K., Yellen, J. E., Gelburd, D. J., Osborn, A. J., & Crowell, A. L. (1996). Subsistence hunting and resource management among the Ju/’Hoansi of Northwestern Botswana. African Study Monographs, 17(4), 153–220. https://doi.org/10.14989/68150
Hopcraft, J. G. C., Sinclair, A. R. E., & Packer, C. (2005). Planning for success: Serengeti lions seek prey accessibility rather than abundance: Prey accessibility in Serengeti lions. Journal of Animal Ecology, 74(3), 559–566. https://doi.org/10.1111/j.1365-2656.2005.00955.x
Hosfield, R. (2020). The earliest Europeans – A year in the life: Seasonal survival strategies in the Lower Palaeolithic. Oxbow Books.
Hovers, E. (2012). Invention, reinvention and innovation: The makings of Oldowan lithic technology. In S. Elias (Ed.), Origins of human innovation and creativity (pp. 51–68). Amsterdam: Elsevier B.V. https://doi.org/10.1016/B978-0-444-53821-5.00005-1
Ichikawa, M. (1983). An examination of the hunting-dependent life of the Mbuti Pygmies, eastern Zaire. African Study Monographs, 4, 55–76. https://doi.org/10.14989/68001
Isaac, G. L. (1977). Olorgesailie: Archaeological studies of a Middle Pleistocene lake basin in Kenya. University of Chicago Press.
Joannes-Boyau, R., Adams, J. W., Austin, C., Arora, M., Moffat, I., Herries, A. I. R., Tonge, M. P., Benazzi, S., Evans, A. R., Kullmer, O., Wroe, S., Dosseto, A., & Fiorenza, L. (2019). Elemental signatures of Australopithecus africanus teeth reveal seasonal dietary stress. Nature, 572(7767), 7767. https://doi.org/10.1038/s41586-019-1370-5
Jones, P. R. (1994). Results of experimental work in relation to the stone industries of Olduvai Gorge. In M. D. Leakey & D. A. Roe (Eds.), Olduvai Gorge volume V: Excavations in Beds III, IV, and the Masek beds 1968–1971 (pp. 254–298). Cambridge University Press.
Keddy, P. A. (2010). Wetland ecology: Principles and conservation (2nd ed.). Cambridge University Press.
Keeley, L. H., & Toth, N. (1981). Microwear polishes on early stone tools from Koobi Fora Kenya. Nature, 293(5832), 464–465. https://doi.org/10.1038/293464a0
Key, A., Proffitt, T., & de la Torre, I. (2020). Raw material optimization and stone tool engineering in the Early Stone Age of Olduvai Gorge (Tanzania). Journal of the Royal Society Interface, 17(162), 20190377. https://doi.org/10.1098/rsif.2019.0377
Kiffner, C., Kioko, J., Leweri, C., & Krause, S. (2014). Seasonal patterns of mixed species groups in large East African mammals. Plos One, 9(12), e113446.
Kofron, C. P. (1993). Behavior of Nile crocodiles in a seasonal river in Zimbabwe. Copeia, 1993(2), 463. https://doi.org/10.2307/1447146
Leakey, M. D. (1971). Olduvai Gorge: Volume 3, excavations in beds I and II, 1960-1963. Cambridge University Press.
Leakey, L. S. B., Tobias, P. V., & Napier, J. R. (1964). A New Species of The Genus Homo From Olduvai Gorge. Nature, 202, 7–9. https://doi.org/10.1038/202007a0
Lee, R. B. (2013). The Dobe Ju/’hoansi (4th ed.). Belmont, CA: Wadsworth.
Lemorini, C., Plummer, T. W., Braun, D. R., Crittenden, A. N., Ditchfield, P. W., Bishop, L. C., Hertel, F., Oliver, J. S., Marlowe, F. W., Schoeninger, M. J., & Potts, R. (2014). Old stones’ song: Use-wear experiments and analysis of the Oldowan quartz and quartzite assemblage from Kanjera South (Kenya). Journal of Human Evolution, 72, 10–25. https://doi.org/10.1016/j.jhevol.2014.03.002
Lemorini, C., Bishop, L. C., Plummer, T. W., Braun, D. R., Ditchfield, P. W., & Oliver, J. S. (2019). Old stones’ song—Second verse: Use-wear analysis of rhyolite and fenetized andesite artifacts from the Oldowan lithic industry of Kanjera South Kenya. Archaeological and Anthropological Sciences, 11(9), 4729–4754. https://doi.org/10.1007/s12520-019-00800-z
Levin, N. E., Simpson, S. W., Quade, J., Everett, M. E., Frost, S. R., Rogers, M. J., & Semaw, S. (2022). The 6-million-year record of ecological and environmental change at Gona, Afar Region, Ethiopia. In S. C. Reynolds, & R. Bobe (Eds.), African paleoecology and human evolution. Cambridge: Cambridge University Press.
Lin, S. C., Režek, Z., Braun, D., & Dibble, H. L. (2013). On the utility and economization of unretouched flakes: The effects of exterior platform angle and platform depth. American Antiquity, 78(4), 724–745. https://doi.org/10.2307/43184970
Linares Matás, G. J., & Yravedra, J. (2021). ‘We hunt to share’: Social dynamics and very large mammal butchery during the Oldowan-Acheulean transition. World Archaeology, 53(2), 224–254. https://doi.org/10.1080/00438243.2022.2030793
Linares-Matás, G. J., & Clark, J. (2022). Seasonality and Oldowan behavioral variability in East Africa. Journal of Human Evolution, 164, 103070. https://doi.org/10.1016/j.jhevol.2021.103070
Lombard, M., & Kyriacou, K. (2018). Hunter-gatherer women. In I. M. Lombard & K. Kyriacou (Eds.), Oxford research encyclopedia of anthropology. https://doi.org/10.1093/acrefore/9780190854584.013.105
Longshore, K. M., Jaeger, J. R., & Sappington, J. M. (2003). Desert tortoise (Gopherus agassizii) survival at two eastern Mojave desert sites: Death by short-term drought? Journal of Herpetology, 37, 169–177.
López-Sáez, J.-A., & Domínguez-Rodrigo, M. (2009). Palynology of OGS-6a and OGS-7, two new 2.6 Ma archaeological sites from Gona, Afar, Ethiopia: Insights on aspects of Late Pliocene habitats and the beginnings of stone-tool use. Geobios, 42(4), 503–511. https://doi.org/10.1016/j.geobios.2008.12.002
Lovich, J. E., Quillman, M., Zitt, B., Schroeder, A., Green, D. E., Yackulic, C., Gibbons, P., & Goode, E. (2017). The effects of drought and fire in the extirpation of an abundant semi-aquatic turtle from a lacustrine environment in the southwestern USA. Knowledge & Management of Aquatic Ecosystems, 418, 1–11. https://doi.org/10.1051/kmae/2017008
Lovich, J. E., & Gibbons, W. (2021). Turtles of the world: A guide to every family. Princeton University Press.
Lupien, R. L., Russell, J. M., Subramanian, A., Kinyanjui, R., Beverly, E. J., Uno, K. T., et al. (2021). Eastern African environmental variation and its role in the evolution and cultural change of Homo over the last 1 million years. Journal of Human Evolution, 157, 103028. https://doi.org/10.1016/j.jhevol.2021.103028
Malonza, P. K. (2003). Ecology and distribution of the pancake tortoise, Malacochersus tornieri in Kenya. Journal of East African Natural History, 92, 81–96.
Marlowe, F. W., & Berbesque, J. C. (2009). Tubers as fallback foods and their impact on Hadza hunter-gatherers. American Journal of Physical Anthropology, 140(4), 751–758. https://doi.org/10.1002/ajpa.21040
McCarthy, T. S., Cooper, G. R. J., Tyson, P. D., & Ellery, W. N. (2000). Seasonal flooding in the Okavango Delta, Botswana—Recent history and future prospects. South African Journal of Science, 96(1), 25–33. https://doi.org/10.10520/AJA00382353_7717
McLennan, M. R. (2013). Diet and feeding ecology of chimpanzees (Pan troglodytes) in Bulindi, Uganda: Foraging strategies at the forest–farm interface. International Journal of Primatology, 34(3), 585–614. https://doi.org/10.1007/s10764-013-9683-y
Mercader, J., Akuku, P., Boivin, N., Bugumba, R., Bushozi, P., Camacho, A., Carter, T., Clarke, S., Cueva-Temprana, A., Durkin, P., Favreau, J., Fella, K., Haberle, S., Hubbard, S., Inwood, J., Itambu, M., Koromo, S., Lee, P., Mohammed, A., et al. (2021). Earliest Olduvai hominins exploited unstable environments ~ 2 million years ago. Nature Communications, 12(1), 3. https://doi.org/10.1038/s41467-020-20176-2
Munro, N. D., & Atici, L. (2009). Human subsistence change in the Late Pleistocene Mediterranean Basin: the statusof research on faunal intensification, diversification & specialisation. Before Farming, 2009, 1–6.
Nicholas, G. P. (1998). Wetlands and hunter-gatherers: A global perspective. Current Anthropology, 39(5), 720–731. https://doi.org/10.1086/204795
Nilssen, P. J. (2000). An actualistic butchery study in south Africa and its implications for reconstructing hominid strategies of carcass acquisition and butchery in the upper Pleistocene and Plio-Pleistocene. Unpublished PhD Dissertation. Cape Town: University of Cape Town, Cape Town.
Njau, J.K., 2012. Crocodile predation and hominin evolution: Landscape paleoanthropology at Olduvai Gorge. LAP GmbH and Co. KG.
O’Brien, K., Hebdon, N., & Faith, J. T. (2023). Paleoecological evidence for environmental specialization in Paranthropus boisei compared to early Homo. Journal of Human Evolution, 177, 103325. https://doi.org/10.1016/j.jhevol.2023.103325
O’Connell, J. F., Hawkes, K., & Blurton Jones, N. (1988). Hadza hunting, butchering, and bone transport and their archaeological implications. Journal of Anthropological Research, 44(2), 113–161. https://doi.org/10.1086/jar.44.2.3630053
O’Connell, J. F., Hawkes, K., & Blurton Jones, N. (1990). Reanalysis of large mammal body part transport among the Hadza. Journal of Archaeological Science, 17(3), 301–316. https://doi.org/10.1016/0305-4403(90)90025-Z
O’Connell, J. F., Hawkes, K., & Blurton-Jones, N. G. (1992). Patterns in the distribution, site structure and assemblage composition of Hadza kill-butchering sites. Journal of Archaeological Science, 19(3), 319–345. https://doi.org/10.1016/0305-4403(92)90020-4
O’Connell, J. F., Hawkes, K., Lupo, K. D., & Blurton Jones, N. G. (2002). Male strategies and Plio-Pleistocene archaeology. Journal of Human Evolution, 43(6), 831–872. https://doi.org/10.1006/jhev.2002.0604
O’Connell, J.F., & Hawkes, K., (in prep). Evolution of human life history: Subsistence, mating sex ratios, carnivore competition.
Oelze, V. M., Head, J. S., Robbins, M. M., Richards, M., & Boesch, C. (2014). Niche differentiation and dietary seasonality among sympatric gorillas and chimpanzees in Loango National Park (Gabon) revealed by stable isotope analysis. Journal of Human Evolution, 66, 95–106. https://doi.org/10.1016/j.jhevol.2013.10.003
Oliver, J. S., Plummer, T. W., Hertel, F., & Bishop, L. C. (2019). Bovid mortality patterns from Kanjera South, Homa Peninsula, Kenya and FLK-Zinj, Olduvai Gorge, Tanzania: Evidence for habitat mediated variability in Oldowan hominin hunting and scavenging behavior. Journal of Human Evolution, 131, 61–75. https://doi.org/10.1016/j.jhevol.2019.03.009
Organista, E., Moclán, A., Aramendi, J., Cobo-Sánchez, L., Egeland, C. P., Uribelarrea, D., Martín-Perea, D., Vegara-Riquelme, M., Hernández-Vivanco, L., Gidna, A., Mabula, A., Baquedano, E., & Domínguez-Rodrigo, M. (2023). A taphonomic analysis of PTK (Bed I, Olduvai Gorge) and its bearing on the interpretation of the dietary and eco-spatial behaviors of early humans. Quaternary Science Reviews, 300, 107913. https://doi.org/10.1016/j.quascirev.2022.107913
Pante, M. C., Njau, J. K., Hensley-Marschand, B., Keevil, T. L., Martín-Ramos, C., Peters, R. F., & de la Torre, I. (2018). The carnivorous feeding behavior of early Homo at HWK EE, Bed II, Olduvai Gorge, Tanzania. Journal of Human Evolution, 120, 215–235. https://doi.org/10.1016/j.jhevol.2017.06.005
Parkinson, J. A. (2018). Revisiting the hunting-versus-scavenging debate at FLK Zinj: A GIS spatial analysis of bone surface modifications produced by hominins and carnivores in the FLK 22 assemblage, Olduvai Gorge, Tanzania. Palaeogeography, Palaeoclimatology, Palaeoecology, 511, 29–51. https://doi.org/10.1016/j.palaeo.2018.06.044
Parkinson, J. A., Plummer, T. W., Oliver, J. S., & Bishop, L. C. (2022). Meat on the menu: GIS spatial distribution analysis of bone surface damage indicates that Oldowan hominins at Kanjera South, Kenya had early access to carcasses. Quaternary Science Reviews, 277, 107314. https://doi.org/10.1016/j.quascirev.2021.107314
Parkinson, J. A. (2013). A GIS image analysis approach to documenting Oldowan hominin carcass acquisition: Evidence from Kanjera South, FLK Zinj, and neotaphonomic models of carnivore bone destruction. Unpublished PhD Dissertation. New York, NY: City University of New York.
Patalano, R., Hamilton, R., Finestone, E., Amano, N., Heddell-Stevens, P., Itambu, M., Petraglia, M., & Roberts, P. (2021). Microhabitat variability in human evolution. Frontiers in Earth Science, 9, 787669. https://doi.org/10.3389/feart.2021.787669
Pennec, F., Gérard, C., Meterreau, L., Monghiemo, C., Ngawolo, J.-C.B., Laurent, R., & Narat, V. (2020). Spatiotemporal variation in bonobo (Pan paniscus) habitat use in a forest–savanna mosaic. International Journal of Primatology, 41, 775–799. https://doi.org/10.1007/s10764-020-00180-5
Petrozzi, F., Hema, E. M., Sirima, D., Segniagbeto, G. H., Akani, G. C., Eniang, E. A., Dendi, D., Fa, J. E., & Luiselli, L. (2020). Tortoise ecology in the West African savannah: Multi-scale habitat selection and activity patterns of a threatened giant species, and its ecological relationships with a smaller-sized species. Acta Oecologica, 105, 103572. https://doi.org/10.1016/j.actao.2020.103572
Plummer, T. W., Oliver, J. S., Finestone, E. M., Ditchfield, P. W., Bishop, L. C., Blumenthal, S. A., Lemorini, C., Caricola, I., Bailey, S. E., Herries, A. I. R., Parkinson, J. A., Whitfield, E., Hertel, F., Kinyanjui, R. N., Vincent, T. H., Li, Y., Louys, J., Frost, S. R., Braun, D. R., … Potts, R. (2023). Expandedgeographic distribution and dietary strategies of the earliest Oldowan hominins and Paranthropus. Science, 379, 561–566. https://doi.org/10.1126/science.abo7452
Posit Team. (2022). RStudio: Integrated development environment for R. In Posit software. Boston, MA: PBC. http://www.posit.co/
Potts, R. (1984). Home bases and early hominids: Reevaluation of the fossil record at Olduvai Gorge suggests that the concentrations of bones and stone tools do not represent fully formed campsites but an antecedent to them. American Scientist, 72(4), 338–347.
Potts, R. (1991). Why the Oldowan? Plio-Pleistocene toolmaking and the transport of resources. Journal of Anthropological Research, 47(2), 153–176. https://doi.org/10.1086/jar.47.2.3630323
Potts, R. (1996a). Evolution and climate variability. Science, 273(5277), 922–923. https://doi.org/10.1126/science.273.5277.922
Potts, R. (1996). Humanity’s descent: The consequences of ecological instability. William Morrow & Co.
Potts, R. (1998). Variability selection in hominid evolution. Evolutionary Anthropology, 7(3), 81–96. https://doi.org/10.1002/(SICI)1520-6505(1998)7:3%3c81::AID-EVAN3%3e3.0.CO;2-A
Potts, R. (2012). Environmental and behavioral evidence pertaining to the evolution of early Homo. Current Anthropology, 53(S6), S299–S317. https://doi.org/10.1086/667704
Potts, R., & Faith, J. T. (2015). Alternating high and low climate variability: The context of natural selection and speciation in Plio-Pleistocene hominin evolution. Journal of Human Evolution, 87, 5–20. https://doi.org/10.1016/j.jhevol.2015.06.014
Prassack, K. A., Pante, M. C., Njau, J. K., & De La Torre, I. (2018). The paleoecology of Pleistocene birds from Middle Bed II, at Olduvai Gorge, Tanzania, and the environmental context of the Oldowan-Acheulean transition. Journal of Human Evolution, 120, 32–47. https://doi.org/10.1016/j.jhevol.2017.11.003
Proffitt, T. (2018). Is there a developed Oldowan A at Olduvai Gorge? A diachronic analysis of the Oldowan in Bed I and Lower-Middle Bed II at Olduvai Gorge, Tanzania. Journal of Human Evolution, 120, 92–113. https://doi.org/10.1016/j.jhevol.2018.01.006
Proffitt, T., Luncz, L. V., Falótico, T., Ottoni, E. B., de la Torre, I., & Haslam, M. (2016). Wild monkeys flake stone tools. Nature, 539(7627), 85–88. https://doi.org/10.1038/nature20112
Quade, J., Levin, N., Semaw, S., Stout, D., Renne, P., Rogers, M., & Simpson, S. (2004). Paleoenvironments of the earliest stone toolmakers, Gona Ethiopia. GSA Bulletin, 116(11–12), 1529–1544. https://doi.org/10.1130/B25358.1
Quade, J., Levin, N. E., Simpson, S. W., Butler, R., McIntosh, W. C., Semaw, S., et al. (2008). The geology of Gona, Afar, Ethiopia. Geological Society of America Special Papers, 446, 1–31.
R Core Team. (2022). R: A language and environment for statistical computing. Vienna: Foundation for Statistical Computing https://www.R-project.org/
Reeves, J. S., Braun, D. R., Finestone, E. M., & Plummer, T. W. (2021). Ecological perspectives on technological diversity at Kanjera South. Journal of Human Evolution, 158, 103029. https://doi.org/10.1016/j.jhevol.2021.103029
Reti, J. S. (2016). Quantifying Oldowan stone tool production at Olduvai Gorge Tanzania. Plos One, 11(1), e0147352. https://doi.org/10.1371/journal.pone.0147352
Režek, Ž, Dibble, H. L., McPherron, S. P., Braun, D. R., & Lin, S. C. (2018). Two million years of flaking stone and the evolutionary efficiency of stone tool technology. Nature Ecology & Evolution, 2(4), 628–633. https://doi.org/10.1038/s41559-018-0488-4
Rivals, F., Moncel, M.-H., & Patou-Mathis, M. (2009). Seasonality and intra-site variation of Neanderthal occupations in the Middle Palaeolithic locality of Payre (Ardèche, France) using dental wear analyses. Journal of Archaeological Science, 36, 1070–1078. https://doi.org/10.1016/j.jas.2008.12.009
Rivals, F., Uno, K. T., Bibi, F., Pante, M. C., Njau, J., & de la Torre, I. (2018). Dietary traits of the ungulates from the HWK EE site at Olduvai Gorge (Tanzania): Diachronic changes and seasonality. Journal of Human Evolution, 120, 203–214. https://doi.org/10.1016/j.jhevol.2017.08.011
Rockman, M., & Steele, J. (Eds.). (2003). Colonization of unfamiliar landscapes: The archaeology of adaptation. London: Routledge.
Sánchez-Yustos, P. (2021). Knocking on Acheulean’s door. DK revisited (Bed I, Olduvai, Tanzania). Journal of Archaeological Science: Reports, 35, 102763. https://doi.org/10.1016/j.jasrep.2020.102763
Sato, H., Kawamura, K., Hayashi, K., Inai, H., & Yamauchi, T. (2012). Addressing the wild yam question: How Baka hunter-gatherers acted and lived during two controlled foraging trips in the tropical rainforest of southeastern Cameroon. Anthropological Science, 120(2), 129–149. https://doi.org/10.1537/ase.110913
Scott, R. S., Ungar, P. S., Bergstrom, T. S., Brown, C. A., Grine, F. E., Teaford, M. F., & Walker, A. (2005). Dental microwear texture analysis shows within-species diet variability in fossil hominins. Nature, 436, 693–695. https://doi.org/10.1038/nature03822
Semaw, S., Renne, P., Harris, J. W. K., Feibel, C. S., Bernor, R. L., Fesseha, N., & Mowbray, K. (1997). 2.5-Million-year-old stone tools from Gona Ethiopia. Nature, 385(6614), 6614. https://doi.org/10.1038/385333a0
Semaw, S., Rogers, M. J., Quade, J., Renne, P. R., Butler, R. F., Dominguez-Rodrigo, M., Stout, D., Hart, W. S., Pickering, T., & Simpson, S. W. (2003). 2.6-Million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar Ethiopia. Journal of Human Evolution, 45(2), 169–177. https://doi.org/10.1016/S0047-2484(03)00093-9
Semaw, S., Rogers, M., & Stout, D. (2009). The Oldowan-Acheulian transition: Is there a “developed Oldowan” artifact tradition? In M. Camps & P. Chauhan (Eds.), Sourcebook of paleolithic transitions: Methods, theories, and interpretations (pp. 173–193). New York, NY: Springer. https://doi.org/10.1007/978-0-387-76487-0_10
Semaw, S. (2006). The oldest stone artifacts from Gona (2.6-2.5 Ma), Afar, Ethiopia: Implications for understanding the earliest stages of stone knapping. In N. Toth & K. Schick (Eds.), The Oldowan: Case studies into the earliest stone age. Ann Arbor, MI: Stone Age Institute Press, University of Michigan.
Shea, J. J. (2017). Occasional, obligatory, and habitual stone tool use in hominin evolution. Evolutionary Anthropology, 26(5), 200–217. https://doi.org/10.1002/evan.21547
Shea, J. J. (2010). Stone age visiting cards revisited: A strategic perspective on the lithic technology of early hominin dispersal. In j. g. fleagle, j. j. shea, F. E. Grine, A. L. Baden, & R. E. Leakey (Eds.), Out of Africa I: The first hominin colonization of Eurasia (pp. 47–64). Dordrecht: Springer Netherlands. https://doi.org/10.1007/978-90-481-9036-2_4
Sheppe, W., & Osborne, T. (1971). Patterns of use of a flood plain by Zambian mammals. Ecological Monographs, 41(3), 179–205. https://doi.org/10.2307/1942365
Shimada, M. (2018). Use of wild date palm (Phoenix reclinata) by Mahale chimpanzees: A likely case of social learning via direct observation. Pan Africa News, 25(2), 19–21.
Sinclair, A. R. E. (1975). The resource limitation of trophic levels in tropical grassland ecosystems. The Journal of Animal Ecology, 44(2), 497. https://doi.org/10.2307/3608
Snyder, W. D., Reeves, J. S., & Tennie, C. (2022). Early knapping techniques do not necessitate cultural transmission. Science Advances, 8(27), eabo2894. https://doi.org/10.1126/sciadv.abo2894
Speth, J. D. (1987). Early hominid subsistence strategies in seasonal habitats. Journal of Archaeological Science, 14(1), 13–29. https://doi.org/10.1016/S0305-4403(87)80003-1
Speth, J. D., & Davis, D. D. (1976). Seasonal variability in early hominid predation: Most archeological sites in Olduvai Gorge beds I and II may be dry season occupations. Science, 192(4238), 441–445. https://doi.org/10.1126/science.192.4238.441
Speth, J. D., Widdowson, E. M., Oftedal, O. T., Foley, R. A., Van Soest, P., Widdowson, E. M., Whiten, A., & Bone, Q. (1991). Protein selection and avoidance strategies of contemporary and ancestral foragers: Unresolved issues. Philosophical Transactions of the Royal Society of London Series B: Biological Sciences, 334(1270), 265–270. https://doi.org/10.1098/rstb.1991.0115
Sponheimer, M., Lee-Thorp, J., de Ruiter, D., Codron, D., Codron, J., Baugh, A. T., & Thackeray, F. (2005). Hominins, sedges, and termites: New carbon isotope data from the Sterkfontein valley and Kruger National Park. Journal of Human Evolution, 48(3), 301–312. https://doi.org/10.1016/j.jhevol.2004.11.008
Sponheimer, M., Passey, B. H., de Ruiter, D. J., Guatelli-Steinberg, D., Cerling, T. E., & Lee-Thorp, J. A. (2006). Isotopic evidence for dietary variability in the early hominin Paranthropus robustus. Science, 314(5801), 980–982. https://doi.org/10.1126/science.1133827
Stewart, K. M. (1994). Early hominid utilisation of fish resources and implications for seasonality and behaviour. Journal of Human Evolution, 27(1), 229–245. https://doi.org/10.1006/jhev.1994.1044
Stewart, K. M., & Murray, A. M. (2020). Pliocene and Pleistocene fishes from Gona, Ethiopia: Inferences for reconstructing freshwater paleoecology. Journal of Vertebrate Paleontology, 40(5), e1819302. https://doi.org/10.1080/02724634.2020.1819302
Stout, D., Quade, J., Semaw, S., Rogers, M. J., & Levin, N. E. (2005). Raw material selectivity of the earliest stone toolmakers at Gona, Afar Ethiopia. Journal of Human Evolution, 48(4), 365–380. https://doi.org/10.1016/j.jhevol.2004.10.006
Stout, D., Semaw, S., Rogers, M. J., & Cauche, D. (2010). Technological variation in the earliest Oldowan from Gona, Afar Ethiopia. Journal of Human Evolution, 58(6), 474–491. https://doi.org/10.1016/j.jhevol.2010.02.005
Stout, D., Rogers, M. J., Jaeggi, A. V., & Semaw, S. (2019). Archaeology and the origins of human cumulative culture: A case study from the earliest Oldowan at Gona Ethiopia. Current Anthropology, 60(3), 309–340. https://doi.org/10.1086/703173
Tarriño, A., Ábalos, B., Puelles, P., Eguiluz, L., & Díez-Martín, F. (2023). The crystalline quartz-rich raw material from Olduvai Gorge (Tanzania): Why is it called quartzite when it should be called quartz? Archaeological and Anthropological Sciences, 15, 78. https://doi.org/10.1007/s12520-023-01774-9
Testart, A. (1982). The significance of food storage among hunter-gatherers: Residence patterns, population densities, and social inequalities. Current Anthropology, 23(5), 523–537.
Thompson, J. C., Carvalho, S., Marean, C. W., & Alemseged, Z. (2019). Origins of the human predatory pattern: The transition to large-animal exploitation by early hominins. Current Anthropology, 60(1), 1–23. https://doi.org/10.1086/701477
Ungar, P. S., Merceron, G., & Scott, R. S. (2007). Dental microwear texture analysis of Varswater bovids and Early Pliocene paleoenvironments of Langebaanweg, Western Cape Province, South Africa. Journal of Mammal Evolution, 14, 163–181. https://doi.org/10.1007/s10914-007-9050-x
Uno, K. T., Rivals, F., Bibi, F., Pante, M., Njau, J., & de la Torre, I. (2018). Large mammal diets and paleoecology across the Oldowan-Acheulean transition at Olduvai Gorge, Tanzania from stable isotope and tooth wear analyses. Journal of Human Evolution, 120, 76–91. https://doi.org/10.1016/j.jhevol.2018.01.002
Uribelarrea, D., Domínguez-Rodrigo, M., Pérez-González, A., Vegas Salamanca, J., Baquedano, E., Mabulla, A., et al. (2014). Geo-archaeological and geometrically corrected reconstruction of the 1.84 Ma FLK Zinj paleolandscape at Olduvai Gorge Tanzania. Quaternary International, 322–323, 7–31. https://doi.org/10.1016/j.quaint.2013.12.023
Uribelarrea, D., Martín-Perea, D., Díez-Martín, F., Sánchez-Yustos, P., Domínguez-Rodrigo, M., Baquedano, E., et al. (2017). A reconstruction of the paleolandscape during the earliest Acheulian of FLK West: The co-existence of Oldowan and Acheulian industries during lowermost Bed II (Olduvai Gorge, Tanzania). Palaeogeography, Palaeoclimatology, Palaeoecology, 488, 50–58. https://doi.org/10.1016/j.palaeo.2017.04.014
Vincent, A. S. (1985). Plant foods in savanna environments: A preliminary report of tubers eaten by the Hadza of northern Tanzania. World Archaeology, 17(2), 131–148. https://doi.org/10.1080/00438243.1985.9979958
Weissner, P., & N!aici, A. (1998). Population, subsistence, and social relations in the Nyae Nyae area: Three decades of change. Unpublished Report. Windhoek: Nyae Nyae Foundation.
Acknowledgements
The research of J.C. was supported by the Arts and Humanities Research Council (grant number AH/R012709/1) through a St. John’s College Match-funded Studentship for his Ph.D. in Biological Anthropology at St. John’s College, University of Cambridge (UK). The work of GLM was supported by a Research Fellowship at Emmanuel College, University of Cambridge, as well as an Arts and Humanities Research Council (grant number AH/R012709/1) Baillie Gifford AHRC Scholarship (OOC-DTP program) for his Ph.D. in Archaeology at St. Hugh’s College, University of Oxford (UK). We thank the Editor, Dr. Jessica Thompson, and two anonymous reviewers for their helpful comments and suggestions which improved the final manuscript.
Funding
No specific additional funding was received for the preparation of this manuscript. The research of J.C. is supported by the Arts and Humanities Research Council (grant number AH/R012709/1). The work of GLM is supported by a Research Fellowship at Emmanuel College, University of Cambridge, as well as the Arts and Humanities Research Council (grant number AH/R012709/1).
Author information
Authors and Affiliations
Contributions
Both JC and GLM contributed equally to the analysis, as well as the writing and reviewing of the manuscript.
Corresponding author
Ethics declarations
Ethical Approval
As the research involved no human or animal involvement, ethical approval was not required for the present investigation.
Competing Interests
The authors have no competing interests, financial or non-financial.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
41982_2023_165_MOESM1_ESM.docx
Supplementary file1 (DOCX 31.1 KB)
41982_2023_165_MOESM3_ESM.xlsx
Supplementary file3 (XLSX 12 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Clark, J., Linares-Matás, G.J. Seasonality and Lithic Investment in the Oldowan. J Paleo Arch 6, 38 (2023). https://doi.org/10.1007/s41982-023-00165-9
Accepted:
Published:
DOI: https://doi.org/10.1007/s41982-023-00165-9