
Abstract
Purpose and methods
Since the second half of the 19th century, the Arcillas de Morella Formation (late Barremian) has yielded abundant vertebrate fossil material from several outcrops of the Morella region (Maestrat Basin, Castellón, eastern Spain). Several historical specimens of fossil reptiles, so far unpublished or not studied in detail, are housed in the Museo Nacional de Ciencias Naturales in Madrid (Spain). In fact, many of the first dinosaur specimens discovered in Spain, from Morella, are part of the vertebrate palaeontology collection of that institution. Herein, this sauropod material is described and discussed in order to study the diversity of the sauropod fauna during the late Barremian on the Maestrat Basin.
Results
The specimens include both axial and appendicular elements. The systematic study of this material suggests the presence of indeterminate titanosauriforms, some of which have somphospondylan and ‘laurasiform’ affinities. Comparative analysis of two isolated humeri from Morella (MNCN 59703 and MNCN 68484) and a humerus subsequently found in this area, indicates the presence of three titanosauriform taxa in the upper Barremian of the Maestrat Basin, two of which have somphospondylan affinities.
Conclusions
The sauropod diversity of the Arcillas de Morella Formation recognized herein, in particularly considering the titanosauriforms, is greater than that previously considered, at least three taxa.
Resumen
Objetivos y métodos
Numerosos especímenes de vertebrados fósiles han sido identificados, desde la segunda mitad del siglo XIX, en varios yacimientos de la Formación Arcillas de Morella (Barremiense superior) situados en la localidad de Morella (Cuenca del Maestrazgo, Castellón, este de España). Varios ejemplares clásicos, correspondientes a fósiles de reptiles procedentes de Morella, pero que permanecían hasta ahora inéditos y que no habían sido detalladamente analizados, están depositados en el Museo Nacional de Ciencias Naturales (Madrid). De hecho, varios de los primeros especímenes de dinosaurios descubiertos en España, procedentes de Morella, forman parte de la colección de paleontología de vertebrados de dicha institución. El material de saurópodos es descrito y discutido en este trabajo, con el objetivo de analizar la diversidad de este grupo de dinosaurios registrada en el Barremiense superior de la Cuenca del Maestrazgo.
Resultados
Los ejemplares analizados incluyen tanto elementos axiales como apendiculares. El estudio sistemático de este material permite reconocer la presencia de titanosauriformes indeterminados, identificándose formas afines a Somphospondyli y ‘Laurasiformes’. El análisis comparativo de dos húmeros clásicos aislados de Morella (MNCN 59703 y MNCN 68484), y de un tercer húmero hallado con posterioridad en esta región, permite reconocer la presencia de tres miembros de Titanosauriformes en el Barremiense superior de la Cuenca del Maestrazgo, dos de ellos compatibles con Somphospondyli.
Conclusiones
La diversidad de saurópodos de la Formación Arcillas de Morella reconocida aquí, compuesta por miembros de Titanosauriformes, es mayor que aquella hasta ahora notificada, estando integrada por, al menos, tres taxones.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The Barremian-Aptian (Lower Cretaceous) sedimentary rocks of the Iberian Peninsula are rich in dinosaur fossil remains. Several discoveries have been made in Portugal (e.g., Lapparent and Zbyszewski 1957; Antunes and Mateus 2003; Mateus et al. 2011a; Figueiredo et al. 2015) and, especially, in Spain (e.g., Lapparent 1966; Sanz et al. 1982, 1988, 1996; Sanz and Buscalioni 1992; Pérez-Moreno et al. 1994; Pereda Suberbiola et al. 2003, 2012; Ruiz Omeñaca and Canudo 2003; Ortega et al. 2006, 2010; Canudo et al. 2008; Gasulla et al. 2011a, b, 2012, 2014, 2015; Ruiz-Omeñaca 2011; McDonald et al. 2012; Royo-Torres et al. 2012; Kirkland et al. 2013; Verdú et al. 2015). Although sauropods are a relatively common group of dinosaurs, the first partial skeletons from the Iberian Peninsula were not found until the end of the twentieth century (e.g., Sanz et al. 1982; Pereda Suberbiola et al. 2003; Fuentes Vidarte et al. 2005; Canudo et al. 2008; Gasulla et al. 2008, 2010; Royo-Torres et al. 2012; Fernández-Baldor et al. 2013).
Only two Early Cretaceous sauropods from the Iberian Peninsula have been described to date: Tastavinsaurus sanzi Canudo et al. 2008 and Demandasaurus darwini Torcida Fernández-Baldor et al. 2011. Tastavinsaurus sanzi is a somphospondylan titanosauriform first found in Peñarroya de Tastavins (Teruel; Xert Formation, uppermost Barremian) and described by Canudo et al. (2008). These authors considered it as the sister taxon of Venenosaurus dicrocei Tidwell et al. 2001, a titanosauriform from the Lower Cretaceous of Utah (USA). Different phylogenetic positions were proposed for Tastavinsaurus on the basis of this specimen (e.g., Royo-Torres 2009; Carballido et al. 2011b; D’Emic 2012). Royo-Torres et al. (2012) described a new specimen referred to T. sanzi from El Castellar (Teruel; Forcall Fm., lower Aptian), this taxon being considered as a basal somphospondylan (following the definition of Wilson and Sereno 1998), belonging to a new clade defined as Laurasiformes (Royo-Torres 2009; Royo-Torres et al. 2012). Similar results were obtained by Royo-Torres et al. (2014). Carballido and Sander (2014) recovered Tastavinsaurus as a non-titanosauriform macronarian. Mannion et al. (2013) and Poropat et al. (2015, 2016) resolved Tastavinsaurus as a non-titanosaurian, non-euhelopodid somphospondylan. Upchurch et al. (2015) resolved it as either a non-titanosauriform macronarian or a non-euhelopodid, non-titanosaurian somphospondylan. Pereda Suberbiola et al. (2003) published a partial skeleton found near Salas de los Infantes (Castrillo de la Reina Fm., upper Barremian-lower Aptian). This specimen was later attributed to a new taxon, the rebbachisaurid Demandasaurus darwini (Torcida Fernández-Baldor et al. 2011), and considered as a member of Nigersaurinae (Whitlock 2011; Carballido et al. 2012). Wilson and Allain (2015) renamed this clade as Rebbachisaurinae. Furthermore, an undescribed titanosauriform specimen from El Oterillo II (Castrillo de la Reina Fm. in Salas de los Infantes, Burgos) has been mentioned (Torcida Fernández-Baldor et al. 2013), as has another, which is based on several skeletons from Sant Antoni de la Vespa (Arcillas de Morella Fm., Morella) (Mocho et al. 2016a).
The Arcillas de Morella Formation (upper Barremian) is rich in vertebrate fossil sites, many of which are notable for their dinosaur fauna (e.g. Sanz et al. 1982; Ortega et al. 2006; Santos-Cubedo et al. 2009; Gasulla et al. 2011b, 2012, 2015). Several sauropod specimens have been identified from this Formation, including some found prior to 1980s (e.g., Sanz et al. 1982, Sanz 1984, 1996; Pereda Suberbiola and Ruiz Omeñaca 1999; Ruiz Omeñaca et al. Ruiz-Omeñaca et al. 2003) and several specimens that have been found since then, some of which comprise partial skeletons (Sanz et al. 1982; Yaguë et al. 2001; Ortega et al. 2006; Gasulla et al. 2008, 2011a, 2012; Santos-Cubedo et al. 2010; Mocho et al. 2016a). All authors agree that Titanosauriformes are present there, with some specimens said to have affinities with Brachiosauridae (Sanz et al. 1982; Yaguë et al. 2001; Ortega et al. 2006; Gasulla et al. 2008) and others with Titanosauria (Santos-Cubedo et al. 2010). More recently, Mocho et al. (2016a) proposed a preliminary phylogenetic approach for the sauropod taxon found in Sant Antoni de la Vespa (Morella). Remains of at least three sauropod individuals were collected from this locality, which yields the most complete sauropod specimens known from this Formation. The Sant Antoni de la Vespa taxon was recovered as a member of the Titanosauriformes clade, closely related to Tastavinsaurus sanzi (late Barremian-early Aptian) (Mocho et al. 2016a). Sanz et al. (1982) identified the remains of a sauropod in El Canteret (Morella); a systematic re-evaluation of this specimen might alter our views on the diversity of this clade in the Early Cretaceous of Spain.
The present study is focused on the detailed description and discussion of several sauropod specimens housed in the Museo Nacional de Ciencias Naturales (Madrid, Spain), among which are some of the first dinosaur specimens found in Spain (see Pérez-García et al. 2009 and references therein). All studied specimens were found in the Arcillas de Morella Formation cropping out in Morella (Maestrat Basin) (Fig. 1). This upper Barremian unit was deposited in muddy and fluvial plains with some marine and tidal influence, especially towards the top of the Formation (Gàmez et al. 2003; Bover-Arnal et al. 2016). The systematic study of these specimens is necessary if we try to understand the composition of the sauropod faunas during the Early Cretaceous of the Maestrat Basin, and their relationship with several subsequent discoveries such as those made at El Canteret and Sant Antoni de la Vespa (e.g., Ortega et al. 2006). In this paper, the presence of three different sauropod taxa in the Arcillas de Morella Fm. is determined for the first time.

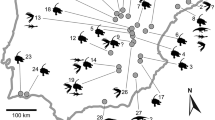
Geographical and stratigraphic locations of the Morella dinosaur sites (Castellón, Spain) which have yielded the sauropod remains mentioned in this paper. a Geographic location of Morella. b Chronostratigraphic chart of the Barremian-Early Aptian of the Maestrat Basin, showing the position of the Arcillas de Morella Formation, from which the remains of sauropods studied in this paper are derived. Modified from Fig. 3 of Bover-Arnal et al. (2016). c Geographical situation of the outcrops or fossiliferous areas of Morella and Cinctorres that produced the sauropod remains cited in this paper: 1 El Beltrán, Morella; 2 outcrop next to the bridge on the Bergantes River, on the road that connects Morella with Cinctorres, Morella; 3 El Canteret, Morella; 4 Sant Antoni de la Vespa, Morella; 5 Ana, Cinctorres
Anatomical abbreviations prdl, prezygodiapophyseal lamina; spof, spinopostzygapophyseal lamina; sprl, spinoprezygapophyseal lamina.
Institutional abbreviations MNCN, Museo Nacional de Ciencias Naturales, Madrid, Spain.
2 Historical context
Systematic studies of Spanish dinosaurs developed much more slowly than in other European countries. In fact, the modern development of this discipline in Spain began in the early 1980s, specifically from the paper of Sanz et al. (1982) in which the dinosaur fauna of the upper Barremian Arcillas de Morella Formation was analysed. In addition, the record from this formation is relevant from a historical point of view, including both: some of the earliest Spanish dinosaur discoveries, presented by Juan Vilanova y Piera in the second half of the nineteenth century (Vilanova y Piera 1872, 1873); the initiation of systematic prospecting and study of these reptiles was performed by José Royo y Gómez and his collaborators in the first decades of the twentieth century (e.g. Royo y Gómez 1926a, b, 1927a, b, see also Pérez-García et al. 2009). The Spanish Civil War (1936–1939) ended the Royo y Gómez research on these faunas. Most of the specimens collected by him were not figured and remain currently undescribed or have not been studied in detail, and have not been reviewed in light of the information now available on both taxa from Morella and other European sauropods.
In 1872, Vilanova y Piera reported the presence of several remains of large reptiles in the area of El Beltrán (Morella) (Vilanova y Piera 1872; Gasulla et al. 2009). The specimen MNCN 59506, studied here, corresponds to one of these fossils. These specimens were discovered in 1868 by Nicolás Ferrer Julve (Segura Barreda 1868; Gasulla Asensio 2005; Pereda Suberbiola and Ruiz-Omeñaca 2005; Pérez-García et al. 2009). In 1873, Vilanova y Piera attributed these remains to Iguanodon, without justifying this identification (Ruiz-Omeñaca et al. 2003; Pérez-García et al., 2009). Ruiz-Omeñaca et al. (2003), Pereda Suberbiola and Ruiz-Omeñaca (2005) and Pereda Suberbiola et al. (2006) indicated that none of the specimens of Morella from the Vilanova y Piera collection can be attributed to Iguanodon. MNCN 59506 was recognized as a neural spine of a diplodocoid sauropod by Pereda Suberbiola and Ruiz-Omeñaca (2005).
In 1914, José Royo y Gómez made his first dinosaur discovery, in the historical locality of El Beltrán (Royo y Gómez 1926a, b). He and his collaborators collected remains of dinosaurs and other Mesozoic reptiles from several outcrops of the Arcillas de Morella Formation for more than two decades.
In the summer of 1922, a bridge over the Bergantes River (in Morella) was built using material extracted from a nearby outcrop. This material included abundant fossil remains, among them appendicular bones and vertebrae of large dinosaurs (Royo y Gómez 1926a, b). The Piarists members of a Catholic religious institute who were based in Morella collected some of these remains and reported their discoveries to Royo y Gómez, who also collected other bones and prospected the area. Thus, in March 1923, he discovered and extracted several specimens, among them MNCN 59697, MNCN 59691 and MNCN 9363 (Pérez-García et al. 2009). Other specimens are also known to have been found in El Beltrán at that time. It is probable, for example, that MNCN 68484 is one of the bones found by the Piarists in that outcrop, having being donated to the MNCN in 1924 (Pérez-García et al. 2009). MNCN 31050, MNCN 50414 and MNCN 68484 come from that outcrop or from El Beltrán (Pérez-García et al. 2009). Accurate information on the date of collection and on the outcrop from where MNCN 59703 derives is not known, but it was also probably found in one of these locations and in that period (see descriptions and discussions of all these specimens in the following sections).
Between November 1924 and February 1925, Royo y Gómez carried out comparative studies between the Spanish material and specimens deposited in several paleontological collections in France, Switzerland, Germany and Belgium (Royo y Gómez 1925; Pérez-García et al. 2009). As a result, he concluded that a sauropod that could belong to the genus Cetiosaurus was represented in the Mesozoic of eastern Spain (both in the Villar del Arzobispo Formation, in Benagéber, and the Arcillas de Morella Fm., in Morella) (Royo y Gómez 1925). However, as in his other works on these reptiles, no sauropod specimen was figured or described, nor the characters of the specimen discussed or the attribution to Sauropoda justified.
Royo y Gómez visited the British Museum (Natural History) (London) in May 1927 in order to conduct additional comparative studies. This allowed him to recognize that the taxon from Benagéber (Valencia) and that from Morella represented two different forms, both of which he identified as cetiosaurids. Royo y Gómez recognized that several of the bones from Morella were attributable to Sauropoda, including a femur (MNCN 9363) and the aforementioned vertebrae (MNCN 59691, 59697), as well as other vertebrae (MNCN 59509, 59510, 59694 and 59695), which were also collected in the previous years. However, several vertebrae now recognized as pertaining to ornithopod dinosaurs were erroneously attributed to Sauropoda by him (MNCN 59693, 59696, 59698 and 59699). In addition, he thought that MNCN 68484, here interpreted as a sauropod humerus, was a stegosaurid femur (see Pérez-García et al. 2009). Other bones attributed to Stegosauria by Royo y Gómez are now known belong to other taxa, as is the case of MNCN 68485, which is an ornithopod vertebra (see Pérez-García et al. 2009). The interpretation by Royo y Gómez that only a single taxon, represented by multiple specimens (which we now realize pertain to multiple taxa) was present in Morella led him to suggest that they represented a new species of Cetiosaurus (Royo y Gómez 1927a, b).
Sanz et al. (1982) and Sanz (1984, 1996) noted that the identification of Cetiosaurus in Morella was unlikely and suggested instead that some of the material pertained to a brachiosaur sauropod. Ruiz-Omeñaca and Pereda-Suberbiola (1999) assigned some sauropod caudal vertebrae from Morella (and more specifically the Royo y Gómez collection) to primitive titanosauriforms. They suggested that the bone MNCN 68484, previously assigned by Royo y Gomez to the femur of a stegosaurid, was a very graceful humerus of a brachiosaur sauropod. Furthermore, Ruiz-Omeñaca et al. (2003) noted the presence of Sauropoda indet. and Titanosauriformes indet. Nevertheless, none of these historical specimens have been analysed in detail. Rectifying and studying these sauropod specimens are the motivations for this paper.
3 Systematic paleontology
All the specimens herein described were found in Morella, in levels of the Arcillas de Morella Formation (late Barremian sensu Bover-Arnal et al. 2016), Maestrat Basin. More precise locality details for each specimen are given in their respective historical context sections).
Dinosauria Owen, 1842
Saurischia Seeley, 1887
Sauropodomorpha Huene, 1932
Sauropoda Marsh, 1878
Sauropoda indet.
(Figure 2a–e)

Historical sauropod specimens from the late Barremian of the Arcillas de Morella Formation (Morella, Castellón, Spain). a–e, Sauropoda indet., right ungual I (MNCN 50414) in medial (a), proximal (b), lateral (c), ventral (d) and dorsal views. f–h Eusauropoda indet., middle or posterior dorsal neural spine (MNCN 59506) in anterior (f), left (g) and posterior (h) view. Scale bars 50 mm. gr groove, spdl spinodiapophyseal lamina, spol spinopostzygapophyseal lamina, sprl spinoprezygapophyseal lamina, tap triangular aliform process
3.1 MNCN 59706
Material Left anterior-middle dorsal rib (MNCN 59706).
Description MNCN 59706 corresponds to a partial anterior or middle left dorsal rib lacking the proximal and distal ends. This rib has an L-shaped cross-section and becomes transversely compressed distally. This element does not preserve evidences of pneumaticity.
Discussion: This rib is referred to Sauropoda indet. because it is L-shaped in cross-section as occur in the middle section of sauropod dorsal ribs (e.g., Janensch 1950).
3.2 MNCN 50414
Material Right pedal ungual I (MNCN 50414) (Fig. 2a–e).
Description MNCN 50414 is interpreted as an ungual I of a right pes (Fig. 2a–e). This bone bears a dorsoventrally convex medial surface and a dorsoventrally flat lateral one; the latter is marked by a longitudinal groove. The distal tip is ventrolaterally deflected. The proximal surface extends to the ventral surface of the ungual. The proximal surface is sub-oval in proximal view and bears ventral and dorsal protuberances. The dorsal protuberance is situated near the dorsal margin and is smaller than the ventral one. These protuberances are separated by a mediolateral groove.
Discussion The sickle-shaped form of MNCN 50414, and the fact that it is much deeper dorsoventrally than broad transversely, gives it a similar appearance to the first pedal unguals of many sauropods. This feature was considered as a synapomorphy of Sauropoda by Wilson (2002).
Eusauropoda Upchurch, 1995
Eusauropoda indet.

Historical sauropod specimens from the late Barremian of the Arcillas de Morella Formation (Morella, Castellón, Spain). Eusauropoda indet., a–f, middle-posterior caudal vertebra (MNCN 59695) in posterior (a), anterior (b), right (c), left (d), dorsal (e) and ventral (f) views; g, l middle-posterior caudal vertebra (MNCN 59509) in right (g) and dorsal (l) view; h–k middle-posterior caudal vertebra (MNCN 59694) in anterior (h), right (i), left (j) and dorsal (k) views; m–q, posterior caudal vertebra (MNCN 59510) in posterior (m), anterior (n), left (o), dorsal (p) and ventral (q) views. Scale bars 50 mm
3.3 MNCN 59506
Material Middle to posterior dorsal (?) neural spine (Fig. 2f–h).
Description This robust neural spine is poorly preserved, with only its left side intact (Fig. 2f–h). Pereda Suberbiola and Ruiz-Omeñaca (2005) also described this element. However, the area recognized here as the lateral surface of the neural spine was interpreted by those authors as anterior. The preserved sector of the spine has a slightly triangular aliform process, laterally projected and well visible in lateral view. A lamina herein interpreted as a spinodiapophyseal lamina (spdl) extends from this process. The other two preserved laminae are identified as the spinoprezygapophyseal (sprl) and spinopostzygapophyseal (spol) laminae. This interpretation suggests that the triangular process is slightly anteriorly displaced from the posterior face of the spine. The spol does not contact the spdl.
Discussion: Our interpretation of this element differs from that proposed by Pereda Suberbiola and Ruiz-Omeñaca (2005). The presence of triangular aliform processes on the middle-posterior dorsal neural spines was considered as synapomorphy of Eusauropoda by Wilson (2002) and of Macronaria by Wilson and Sereno (1998), Upchurch et al. (2004) and Carballido et al. (2011b). However, Carballido and Sander (2014) scored several basal eusauropods (including Shunosaurus, Mamenchisaurus and Patagosaurus) as having aliform processes on their middle-posterior dorsal neural spines, suggesting that this feature is a synapomorphy of Eusauropoda. Furthermore, members of the non-neosauropod group Turiasauria also possess these processes (Casanovas et al. 2001; Royo-Torres et al. 2006). Pereda Suberbiola and Ruiz-Omeñaca (2005) suggested that this neural spine was paddle-shaped. Our reinterpretation of this specimen invalidates this interpretation. Consequently, the assignation of MNCN 59506 to Diplodocoidea is not confirmed. No synapomorphies of Diplodocoidea or more exclusive groups (e.g., Wilson 2002; Upchurch et al. 2004; Whitlock 2011; Mannion et al. 2012; Tschopp et al. 2015) can be identified in MNCN 59506.
3.4 MNCN 59695
Material Middle-posterior caudal vertebra (MNCN 59695) (Fig. 3a–f).
Description MNCN 59695 corresponds to a posterior caudal vertebra lacking the neural arch (Fig. 3a–f). The neural arch is centred anteriorly to the mid-point of the centrum. The posterior and anterior articulation faces are both concave and transversely compressed. The lateral surface is dorsoventrally convex and lacks pneumatic fossae or foramina. A longitudinal crest is present near the dorsal margin of the lateral face. A rounded boss is present on the lateral face of the caudal vertebra, near the base of the neural arch. The ventral surface of this bone is transversely convex. The margins of the articular faces are poorly preserved, and no chevrons facets are present.
Discussion: MNCN 59695 centrum is transversely compressed, distinguishing it from the caudal vertebrae of several basal titanosauriforms (Upchurch et al. 2004) such as Giraffatitan (Janensch 1950), Cedarosaurus (Tidwell et al. 1999), Venenosaurus (Tidwell et al. 2001), Tastavinsaurus (Canudo et al. 2008; Royo-Torres, 2009), Lusotitan atalaiensis (Mannion et al. 2013; Mocho et al. 2016b) and Galveosaurus (Barco 2009). However, some titanosauriforms have transversely compressed middle and posterior centra (Csiki et al. 2010; Mannion and Calvo 2011). The neural arch is markedly displaced anteriorly as in Titanosauriformes (Salgado et al. 1997; D’Emic 2012; Mannion et al. 2013) and the non-neosauropod eusauropod Cetiosaurus oxoniensis (Upchurch and Martin 2003). Despite the absence of non-neosauropod material from the Iberian Early Cretaceous, the presence of non-neosauropod eusauropods in the Early Cretaceous of Europe, North America and Africa has recently been suggested (Sereno et al. 1999; Mocho et al. 2016c; Royo-Torres et al. 2016). Accordingly, this middle-posterior caudal vertebra is interpreted herein as pertaining to an indeterminate eusauropod.
3.5 MNCN 59694
Material Middle-posterior caudal vertebra (MNCN 59694) (Fig. 3h–k).
Description MNCN 59694 is a middle-posterior caudal vertebra lacking the neural arch (Fig. 3h–k). The centrum has concave anterior and posterior articulation faces. Both faces bear small pits in their center. The neural arch is marked displaced anteriorly. A small bulge is present on the lateral face of the vertebra, at the transition between the neural arch and the centrum. A longitudinal crest is present ventral to this bulge on the lateral face, above the midheight of the centrum. The margins of the articular surfaces are poorly preserved and the chevron articulations could not be described. The ventral face is transversely convex.
Discussion The presence of an anteriorly-displaced neural arch suggests that this vertebra should be referred to an indeterminate eusauropod (see the discussion for MNCN 59695).
3.6 MNCN 59510
Material Posterior caudal vertebra (MNCN 59510) (Fig. 3m–q).
Description MNCN 59510 corresponds to a posterior caudal vertebra (more posterior than MNCN 59695 and 59694) lacking the neural arch (Fig. 3m–q). As in MNCN 59695 and 59694, the neural arch is anteriorly located. The centrum is dorsoventrally compressed. The posterior and anterior articular faces are concave. The lateral surface is dorsoventrally convex and lacks pneumatic fossae and foramina. A longitudinal crest, interrupted at midpoint, is present near the ventral margin of the lateral face. A rounded boss is present in the lateral surface of the centrum, near the base of the neural arch. The ventral surface is slightly transversely concave. The articulations for the chevrons are poorly developed.
Discussion MNCN 59510 centrum is dorsoventrally compressed, as is the case in the posterior caudal vertebrae of several titanosauriforms (e.g., Janensch 1950; Tidwell et al. 1999, 2001; Upchurch et al. 2004; Canudo et al. 2008; Barco 2009; Royo-Torres 2009; Mannion et al. 2013; Mocho et al. 2016b), and in some non-neosauropod eusauropods such as Cetiosaurus oxoniensis Phillips 1871 (Upchurch and Martin 2003). As in MNCN 59695 and MNCN 59694, this centrum is characterized by an anterior displacement of the neural arch, shared with Titanosauriformes (Salgado et al. 1997; D’Emic 2012; Mannion et al. 2013) and the non-neosauropod eusauropod Cetiosaurus oxoniensis (Upchurch and Martin 2003). The presence of anterior displacement of the neural arch and the dorsoventrally compressed centrum suggest that this vertebra belongs to an indeterminate eusauropod.
3.7 MNCN 59509
Material Middle-posterior caudal vertebra (MNCN 59509) (Fig. 3g, l).
Description MNCN 59509 is interpreted as the centrum of a middle-posterior caudal vertebra lacking the neural arch (Fig. 3g, l). This centrum is markedly dorsoventrally compressed, but this condition might simply be a consequence of taphonomic deformation. The centrum is relatively long and amphicoelous. The neural arch is clearly anteriorly displaced. The ventral surface is transversely wide and flat. The lateral face of the right side is dorsoventrally concave. The margins of the posterior and anterior articular surfaces are poorly preserved.
Discussion: The centrum of MNCN 59509 is dorsoventrally compressed and has a pronounced anteriorly displaced neural arch. As occur in MNCN 59510, these features are common in several titanosauriforms (e.g., Janensch 1950; Tidwell et al. 1999, 2001; Upchurch et al. 2004; Canudo et al. 2008; Barco 2009; Royo-Torres 2009; Mannion et al. 2013; Mocho et al. 2016b), and in the non-neosauropod Cetiosaurus oxoniensis (Upchurch and Martin 2003). This combination of features is only present within Eusauropoda. Therefore, MNCN 59509 is here referred to Eusauropoda indet.
Neosauropoda Bonaparte, 1986
Neosauropoda indet.
(Figure 4f)

Historical sauropod specimens from the late Barremian of the Arcillas de Morella Formation (Morella, Castellón, Spain). a–e Titanosauriformes indet., metacarpal II or IV (MNCN 31050) in dorsal (a), lateral/medial? (b), posterior (c), lateral/medial? (d) and distal (e) views. Macronaria indet., left femur (MNCN 9363) in anterior view (f). Scale bars 50 mm. icg intercondylar groove, lb lateral bulge
3.8 MNCN 9363
Material Proximal end of a left femur (MNCN 9363) (Fig. 4f).
Description Few details of the partial femur MNCN 9363 can be described as a consequence of its poor state of preservation. A lateral bulge is present as in Titanosauriformes (e.g. Salgado et al. 1997). Unfortunately, it is not possible to determine the level of development of the trochanteric shelf. The femoral head is elliptical in proximal view, being anteroposteriorly compressed.
Discussion The lateral bulge identified on this femur is a characteristic widely considered as a synapomorphy of Titanosauriformes (e.g., Salgado et al. 1997; Wilson 2002; Upchurch et al. 2004; D’Emic 2012) or of a more inclusive group within Macronaria (Carballido et al. 2011b; Carballido and Sander 2014). The femora of some derived diplodocids also developed a lateral bulge (e.g., Hatcher 1901; McIntosh 2005); however, diplodocids have not yet been recognized in the European Cretaceous record (e.g., Royo-Torres 2009; Torcida Fernández-Baldor et al. 2011; Mannion et al. 2012; Royo-Torres et al. 2012; Tschopp et al. 2015). The presence of a lateral bulge suggests that MNCN 9363 pertains to a neosauropod form, and possibly to an indeterminate macronarian.
Macronaria Wilson and Sereno, 1998
Titanosauriformes Salgado et al. 1997
Titanosauriformes indet.

Historical sauropod specimens from the late Barremian of the Arcillas de Morella Formation (Morella, Castellón, Spain). a–d Titanosauriformes indet., left humerus (MNCN 59703) in anterior (a), lateral (b), posterior (c) and medial (d) views. e–i Somphospondyli indet., right humerus (MNCN 68484) in anterior (e), lateral (f), posterior (g), medial (h) and distal (i) views. Scale bars 100 mm. ac accessory condyle, af anconeal fossa, dpc deltopectoral crest, rc radial condyle, ulnar condyle

Historical sauropod specimens from the late Barremian of the Arcillas de Morella Formation (Morella, Castellón, Spain). Titanosauriformes indet., a–f, anterior caudal vertebra (MNCN 59691) in posterior (a), anterior (b), right (c), left (d), ventral (e) and dorsal (f) views; g–l anterior caudal vertebra (MNCN 59697) in posterior (g), anterior (h), right (i), left (j), ventral (k) and dorsal (l) views. Scale bar 100 mm. aca anterior chevron articulation, cr caudal rib, pca posterior chevron articulation
3.9 MNCN 31050
Material Distal end of a metacarpal II or IV (MNCN 31050) (Fig. 4a–e).
Description MNCN 31050 appears to be the distal end of a metacarpal II or IV (Fig. 4a–e). The dorsal surface is flat and the ventral one is transversely concave. The ventral surface is composed of two slightly individualized condyles (dorsoventrally oriented) separated by an intercondylar groove. The distal face is distally restricted.
Discussion MNCN 31050 shares with Titanosauriformes the presence of a reduced metacarpal distal articular facet, which was considered a synapomorphy of Titanosauriformes by D’Emic (2012).
3.10 MNCN 59703
Material Left humerus lacking the distal and proximal end (MNCN 59703) (Fig. 5a–d).
Description: The distal tip of the deltopectoral crest of the partial humerus MNCN 59703 is directed anteriorly and shows neither medial displacement nor distal expansion. The proximal tip of the deltopectoral crest is not preserved. Transversely, the anterior face of the distal end is slightly concave. This concavity is bordered by two proximodistal crests, which probably connected to the radial and ulnar condyles. The humeral diaphysis is markedly anteroposteriorly compressed, with an elliptical cross-section. No posterior bulge is present behind the deltopectoral crest.
Discussion The presence of a reduced humeral deltopectoral crest, present in MNCN 59703, was considered a synapomorphy of Sauropoda by Wilson and Sereno (1998) and Wilson (2002). The humeral shaft eccentricity (mediolateral to anteroposterior width ratio at midshaft) of MNCN 59703 is 2.2. Such high values were recorded for some titanosauriforms, such as Galveosaurus and Angolatitan (Mateus et al., 2011b). Thus, MNCN 59703 is referred as Titanosauriformes indet.
3.11 MNCN 59691
Material Anterior caudal vertebra (MNCN 59691) (Fig. 6a–f).
Description MNCN 59691 is an anterior caudal vertebra with a concave anterior face and flat-to-slightly concave posterior one (Fig. 6a–f). It is very similar in size and morphology to MNCN 59697. The centrum is dorsoventrally compressed (albeit not as markedly as in MNCN 59697) and the neural arch is anteriorly displaced. The caudal ribs are reduced (more developed than in MNCN 59697), posterolaterally oriented and extend to the lateral face of the neural arch. No prezygodiapophyseal lamina (prdl) is present. The ventral face is transversely constricted and bears smooth longitudinal crests projecting from the posterior chevron articulations. Between these smooth crests, the ventral face is slightly concave. As in MNCN 59697, the ventral border of the anterior articulation is poorly preserved. The posterior articulations for the chevrons are well-developed and semicircular. The base of the neural spine is present, and shows a slight anterodorsal orientation. The posterior face of the neural spine is poorly preserved. Consequently, the presence of a hyposphenic crest cannot be excluded.
Discussion The presence of dorsoventrally compressed anterior caudal centra is a common feature among non-titanosaur titanosauriforms, such as Giraffatitan (Janensch 1950), Lusotitan (Mocho et al. 2016b), Astrophocaudia (D’Emic 2013), Tastavinsaurus (Royo-Torres 2009), Galveosaurus (Barco 2009), Chubutisaurus (Carballido et al. 2011a), Sauroposeidon (Rose 2007; D’Emic and Foreman 2012) and in a few titanosaurs (e.g., Powell 1992; Kellner and Azevedo 1999). However, this feature is also present in the non-neosauropod eusauropod Cetiosaurus oxoniensis (Upchurch and Martin 2003). The posterior orientation of the caudal ribs in the last anterior caudal vertebrae are common in Titanosauriformes (Mannion et al. 2013) and considered a synapomorphy of the group by D’Emic (2012). In some titanosauriforms, these ribs reach or surpass the posterior articulation, including Lusotitan (Mannion et al. 2013; Mocho et al. 2016b), Sonorasaurus (Ratkevich 1998) and Tastavinsaurus (Royo-Torres 2009). For the presence of this feature to be accurately determined, is necessary to access to well-preserved anterior caudal series. Thus, the fact that this caudal vertebra is isolated means that it is impossible to confirm the absence of this feature in the caudal series of the sauropod to which MNCN 59691 belonged. Some non-titanosauriforms also possess posteriorly projecting caudal ribs in the anterior caudal vertebrae, such as Spinophorosaurus (pers. obs., PM), Omeisaurus (Mannion et al. 2013) and Jobaria (D’Emic 2012). The presence of flat posterior articulations in the anterior caudal vertebrae is a common feature in Macronaria (Mocho et al. 2014). In some sauropods, a flat posterior surface marks the transition between procoelous and amphicoelous condition; this generally occurs in the anterior-middle section of the series (e.g., Casanovas et al. 2001; McIntosh 2005).
One of the most important features of this caudal vertebra is the orientation of the neural spine. Anteriorly directed neural spines in anterior and middle caudal vertebrae are relatively uncommon in sauropods, with a few titanosauriforms having developed this feature, such as Cedarosaurus (Tidwell et al. 1999), Venenosaurus (Tidwell et al. 2001), Tastavinsaurus (Royo-Torres 2009), Malarguesaurus (González Riga et al. 2009), Gondwanatitan (Kellner and Azevedo 1999), Aeolosaurus rionegrinus Powell 1987 and Aeolosaurus maximus Santucci and Arruda-Campos, 2011. This feature is also interpreted for MNCN 59697. The slight anterodorsal projection of the neural spine was considered as a putative synapomorphy of the ‘Laurasiformes’, which includes Venenosaurus, Tastavinsaurus and Cedarosaurus (Royo-Torres et al. 2012). MNCN 59691 is easily distinguished from the titanosaurs Gondwanatitan, Aeolosaurus rionegrinus and Aeolosaurus maximus, since it does not share with them several other features such as the presence of anterodorsally-projecting anterior corporeal articular surfaces, postzygapophyses located on the anterior end of the centrum, dorsally and ventrally expanded prezygapophyses and markedly convex posterior corporeal articular surfaces. Malarguesaurus, from the Upper Cretaceous of Mendoza (Argentina), was originally considered a non-titanosaur titanosauriform (González Riga et al. 2009), but recent studies have suggested other phylogenetic positions within Titanosauriformes (Mannion et al. 2013; Carballido and Sander 2014; Poropat et al. 2015; Gorscak and O’Connor 2016). The uncertain phylogenetic position is probably related to the fact that Malarguesaurus was established on a rather incomplete specimen (González Riga et al. 2009). The presence of anterior caudal centra with flat posterior articulations, posteriorly-projecting caudal ribs, dorsoventrally-compressed anterior centra and anterodorsally-projecting neural spines is a combination present in only some Titanosauriformes. The anterodorsally-projected neural spines and the overall morphology of this vertebra allies it with the putative members of ‘Laurasiformes’ clade, Cedarosaurus and Venenosaurus, also proposed as members of Brachiosauridae (Mannion et al. 2013; Carballido and Sander 2014). ‘Laurasiformes’ have been recovered by some authors (Royo-Torres 2009; Royo-Torres et al. 2012; Mocho et al. 2014; Royo-Torres et al. 2014); however, recent phylogenetic analyses do not support the monophyly of this clade (D’Emic 2012; Mannion et al. 2013; Carballido and Sander 2014; Poropat et al. 2015). In conclusion, MNCN 59691 is considered as Titanosauriformes indet.
3.12 MNCN 59697
Material Anterior caudal vertebra (MNCN 59697) (Fig. 6g–l).
Description MNCN 59697 is an amphicoelous anterior caudal vertebra (Fig. 6g–l), situated farther posteriorly in the tail than MNCN 59691. The centrum is dorsoventrally compressed and the neural arch is anteriorly displaced. The caudal ribs are reduced but extend to the lateral surface of the neural arch. No prdl is present. The ventral surface is flat, transversally constricted and bears smooth longitudinal crests projecting from the posterior articulations for the chevrons. The ventral border of the anterior articular facet is poorly preserved, making it difficult to determine the presence/absence of the anterior chevron articulations. Only the base of the neural spine is preserved, and a slight anterodorsal orientation of the neural spine is interpreted.
Discussion Flat posterior articular surface, posteriorly-projecting caudal ribs, dorsoventrally-compressed centrum and anterodorsally-projecting neural spines are a combination present in only some Titanosauriformes. The anterodorsally-projected neural spines and the overall morphology are shared with the putative members of ‘Laurasiformes’ (see discussion of MNCN 59691).
Somphospondyli Wilson and Sereno (1998)
Somphospondyli indet.
(Figure 5e–i)
3.13 MNCN 68484
Material Right humerus, lacking the proximal end (MNCN 68484) (Fig. 5e–i).
Description MNCN 68484 is a very slender humerus, only slightly expanded in the distal end (Fig. 5e–i). The diaphysis has a subcircular cross-section. The deltopectoral crest is relatively pronounced. It has a roughened surface and projects anteromedially. The base of the deltopectoral crest is also medially displaced. This crest dissipates ventrally, giving rise to a proximodistal crest on the anterior face of the diaphysis. A bulge on the posterior face, near the lateral margin of the humerus, is present behind the deltopectoral crest. In anterior view, the lateral margin of the humerus is straight, resulting in an asymmetrical transverse expansion of the proximal end. The distal end is roughened and transversely flat, as occur in eusauropods (excluding Saltasauridae; Wilson 2002) and the ulnar and radial condyles are medially bevelled. The anteroventral margin of the distal end preserves two accessory condyles near the medial margin. The anterior surface of the distal end is flat, whereas the posterior one is transversely and deeply concave (the anconeal fossa). The distal condyles slightly extend to the anterior and posterior surface of the distal end. An accessory condyle is present in the center of the anterodistal edge of the humerus.
Discussion This humerus has several important features. Despite the absence of the proximal end, it is clear that this element was overall extremely slender (length/midshaft width higher than 7.5), probably more slender than the humeri of brachiosaurids (Janensch 1961; Tidwell et al. 1999) and other derived titanosauriforms such as Ligabuesaurus (Bonaparte et al. 2006). The presence of a gracile humerus was considered a synapomorphy of Giraffatitan + (Brachiosaurus + (Abydosaurus, Cedarosaurus, Venenosaurus)) by D’Emic (2012). Deltopectoral crests that extend medially across the anterior face of the humerus are common in Titanosauriformes, such as Cedarosaurus (Tidwell et al. 1999), Angolatitan (Mateus et al. 2011b), Dreadnoughtus (Lacovara et al. 2014) and Opisthocoelicaudia (Borsuk-Bialynicka 1977). This feature was recovered as a synapomorphy of Titanosauria (based on LSDM matrix) or Lithostrotia (based on LCDM matrix) by Mannion et al. (2013). MNCN 68484 also has a strong bulge (attachment site for M. latissimus dorsi) close to the lateral margin of the posterior surface, behind the deltopectoral crest. This bulge is present in some saltasaurids such as Saltasaurus (Powell 1992), Neuquensaurus (Otero 2010) and Opisthocoelicaudia (Borsuk-Bialynicka 1977) but also in Cedarosaurus (Tidwell et al. 1999), Angolatitan (Mateus et al. 2011b), Epachthosaurus (Martínez et al. 2004), Lirainosaurus (Díez Díaz et al. 2013), Jainosaurus (Wilson et al. 2011) and Zby (Mateus et al. 2014).
The diaphysis is subcircular in cross-section, differing from the condition present in Cedarosaurus (Tidwell et al. 1999), Brachiosaurus (Riggs 1903), Lusotitan (Mannion et al. 2013) and Giraffatitan (Janensch 1961), all of which possess anteroposteriorly-compressed humeral diaphyses. Humerus shaft eccentricity (mediolateral to anteroposterior width ratio at midshaft) is 1 for MNCN 68484, being less eccentric than that of other sauropods as Giraffatitan, Lapparentosaurus, Ligabuesaurus, Diplodocus, Nigersaurus or Europasaurus (see Mannion et al. 2013). The presence of a circular cross-section of the humeral diaphysis was considered diagnostic of Rebbachisauridae (Wilson 2002). However, rebbachisaurid humeri are much more robust (e.g., Wilson and Allain 2015) than MNCN 68484, and do not have anteromedially projected deltopectoral crests or deep anconeal fossae.
The presence of a deep anconeal fossa has been identified by several authors as an important feature in derived titanosauriforms (e.g., Borsuk-Bialynicka 1977; Mateus et al. 2011b; Poropat et al. 2015), and considered as a synapomorphy of titanosaurs by Upchurch et al. (2004) and of Somphospondyli/Titanosauria by Mannion et al. (2013). This would imply that non-neosauropod eusauropods like Zby and Turiasaurus acquired their deep anconeal fossae convergently (Royo-Torres et al. 2006; Mateus et al. 2014). The presence of a straight lateral margin in anterior view is common in several titanosauriforms, such as Cedarosaurus (Tidwell et al. 1999), a putative titanosauriform from the Lower Cretaceous of the Spanish Province of Soria (Fuentes Vidarte et al. 2005), Chubutisaurus (Carballido et al. 2011a), Ligabuesaurus (Bonaparte et al. 2006), Rukwatitan (Gorscak et al. 2014), Muyelensaurus (Calvo et al. 2007), Malawisaurus (Gomani 2005) and Opisthocoelicaudia (Borsuk-Bialynicka 1977), as well as in the rebbachisaurid Comahuesaurus (Carballido et al. 2012). This combination of characters (i.e., deep anconeal fossa, a bulge in the posterior surface and behind the deltopectoral crest, medially-deflected deltopectoral crest and straight lateral margin of the humerus) was only reported in somphospondylan sauropods, suggesting that this humerus might pertain to a indeterminate somphospondylan.
4 Sauropods of the upper Barremian Arcillas de Morella Formation
Several sauropod specimens were recovered in Arcillas de Morella Formation across several sites in Morella (Castellón, Spain) (e.g., Segura Barreda 1868; Vilanova y Piera 1872; Royo y Gómez 1925, 1926a, b; Sanz et al. 1982; Sanz 1984, 1996; Ruiz-Omeñaca and Pereda-Suberbiola 1999; Yaguë et al. 2001; Gasulla Asensio 2005; Pereda Suberbiola and Ruiz-Omeñaca 2005; Ortega et al. 2006; Gasulla et al. 2008, 2011a, 2015; Pérez-García et al. 2009; Santos-Cubedo et al. 2010; Mocho et al. 2016a). The Arcillas de Morella Formation of the Maestrat Basin, from which all of the sauropod specimens described in this paper come, has recently been considered to be late Barremian in age. It directly underlies the Xert Formation, which is in turn overlain by the Forcall Formation (Villanueva-Amadoz et al. 2014; Bover-Arnal et al. 2016); sauropod remains have also been reported from these formations (Canudo et al. 2008; Royo-Torres 2009; Royo-Torres et al. 2012).
Although sauropod remains from the Arcillas de Morella Formation were reported in the second half of the nineteenth century and in the first decades of the twentieth century, the first partial sauropod skeletons from Morella were only identified during the final decades of the twentieth century (Sanz et al. 1982; Yagüe et al. 2001; Gasulla et al. 2008, 2011a, 2012; Ortega et al. 2006). These recent finds have provided a set of specimens, which preserve valuable information about the phylogenetic affinities of the sauropods to which they pertained and the relationship of these taxa with the others from the Maestrat Basin, especially with those from the Xert and Forcall Formations. Our study, based on the sauropod specimens deposited in the collections of the Museo Nacional de Ciencias Naturales (Madrid), has demonstrated the presence of material bearing shared features with Titanosauriformes (MNCN 31050, 59703, 68484, 59691 and 59697). This agrees with the most recent systematic approaches, some based on new material found in the Arcillas de Morella Formation (Sanz et al. 1982; Yagüe et al. 2001; Ortega et al. 2006; Gasulla et al. 2008, 2011a, 2012). However, the presence of brachiosaurid sauropods, an interpretation widely applied to some of the specimens recovered from the Arcillas de Morella Fm.—including some of the specimens studied in the present study (e.g., Sanz et al. 1982; Ruiz-Omeñaca and Pereda-Suberbiola 1999; Yagüe et al. 2001; Ortega et al. 2006; Gasulla et al. 2008) cannot not be confirmed. The discovery and description of more complete specimens might eventually lead to the recognition of brachiosaurids in this Formation.
The systematic proposal of Ruiz-Omeñaca and Pereda Suberbiola (1999) considering MNCN 59697 and MNCN 59691 as a basal titanosauriform is supported here, and the detailed study performed herein identifies the presence of features shared with Tastavinsaurus, Cedarosaurus and Venenosaurus, considered by some authors as members of the ‘Laurasiformes’ clade. Thus, these specimens are herein identified as possible evidence of ‘laurasiforms’ in the Arcillas de Morella Formation. This group has already been identified in the Xert and Forcal Formations (Royo-Torres et al. 2012). Our research indicates that the stratigraphic distribution of this clade in the Maestrat Basin was quite broad. These caudal vertebrae share the same kind of morphology with the sauropods found in Sant Antoni de la Vespa (Morella, Arcillas de Morella Fm.) (Ortega et al. 2006; pers. observ., PM), which appear to pertain to a form closely related to Tastavinsaurus sanzi (Mocho et al. 2016a).
The humeri described in this paper have proven to be an important source of information about the diversity of sauropods present in Arcillas de Morella Formation. MNCN 59703 and 68484 are referred to Titanosauriformes, and the latter might pertain to a somphospondylan. The presence of somphospondylan sauropods in the Barremian-Aptian of the Iberian Peninsula is not surprising, since some phylogenetic analyses have placed Tastavinsaurus within this clade (D’Emic 2012; Mocho et al. 2016a), and also because the presence of other possible derived somphospondylan material in the Iberian Barremian-Aptian (Sánchez-Hernández et al. 2007; Santos-Cubedo et al. 2010). The presence of titanosaurian forms has been discussed in some European Barremian-Aptian localities (e.g., Le Loeuff 1993; D’Emic 2012; Dal Sasso et al. 2016).
The description of MNCN 59703 and 68484 provide evidence of two different humeral morphotypes in Morella. One is characterized by a slender diaphysis with a circular cross-section, a straight lateral edge, a deep anconeal fossa and a medially-displaced and anteromedially-projected deltopectoral crest (MNCN 68484). The other morphotype is characterized by a robust and anteroposteriorly-compressed diaphysis, and an anteriorly-projected deltopectoral crest (MNCN 59703). On the basis of the morphological variation present in other macronarians (e.g., Camarasaurus, Giraffatitan, Lirainosaurus), it is probable that this marked morphological variability does not reflect intraspecific variability. This suggests the presence of at least two titanosauriform taxa in the sauropod collections from the Arcillas de Morella Formation of the MNCN, one of which appears to have somphospondylan affinities. When both humeral morphotypes are compared with the El Cantaret sauropod, which also derives from this Formation (see Sanz et al. 1982), a third humeral morphotype can be recognized. This is characterized by a slender and slightly anteroposteriorly-compressed diaphysis, an anteroposteriorly-projected deltopectoral crest and a deep anconeal fossa, and this morphotype is interpreted herein to represent an indeterminate somphospondylan (see plate III, fig. 1a–b in Sanz et al. 1982). This humerus differs from MNCN 68484 by the absence of an anteromedially-projected deltopectoral crest, a diaphysis with a circular cross-section and straight lateral margin. On the other hand, this humerus is much more slender than MNCN 59703, which has a marked anteroposteriorly-compressed diaphysis. In conclusion, the three humeral morphotypes recognized in the Arcillas de Morella Fm. could correspond to three distinct titanosauriform taxa, indicating a high diversity of sauropods in this area during the Early Cretaceous. Unfortunately, only the El Canteret humerus is associated with a partial skeleton (see Sanz et al. 1982). This specimen is clearly distinct from the Sant Antoni de la Vespa sauropod (which does not preserve any humerus), this last form being considered as possible more closely related to the ‘laurasiform’ Tastavinsaurus. Furthermore, the systematic revision of MNCN 59506 refutes the presence of any recognized diplodocoid form in the Arcillas de Morella Formation, contra Pereda Suberbiola and Ruiz-Omeñaca (2005).
This comparative study of the known sauropod remains from the Arcillas de Morella Formation has allowed us to determine that the sauropod diversity of this formation was rather high, with three titanosauriforms (recognized herein based on three different humeral morphotypes), including a possible ‘laurasiform’ (MNCN 59697 and MNCN 59691 and the Sant Antoni de la Vespa sauropod) and two other somphospondylans (El Canteret sauropod and MNCN 68484). MNCN 59703 is a humerus referred to an indeterminate titanosauriform, which might correspond to a non-somphoposdylan or a somphospondylan sauropod. The incorporation of the ‘Laurasiformes’ within Somphospondyli as well as its monophyly is under discussion (e.g., Royo-Torres 2009; Royo-Torres et al. 2012, 2014; Carballido et al. 2011b; D’Emic 2012; Mannion et al. 2013; Carballido and Sander 2014; Mocho et al. 2014; Upchurch et al. 2015). The presence of a titanosaurian sauropod in this Formation, as suggested by Santos-Cubedo et al. (2010), has not yet been ruled out, pending a detailed study of the sauropod femur collected in Ana (Cinctorres) and the discovery of new material. Nevertheless, some of the features used to support this taxonomic assessment present a wider distribution within Titanosauriformes (see D’Emic 2012; Royo-Torres et al. 2012; Mannion et al. 2013; Poropat et al. 2015).
The sauropod faunas in the Xert (uppermost Barremian) and Forcall (early Aptian) Formations, also deposited in the Maestrat Basin and above the Arcillas de Morella Formation, are represented by the titanosauriform Tastavinsaurus sanzi (Canudo et al. 2008; Royo-Torres 2009, Royo-Torres et al. 2012). However, new discoveries will be necessary to understand the evolutionary history of the sauropod faunas throughout the Barremian-Aptian sequence in the Maestrat Basin. Finally, several sauropod occurrences have also been reported from the upper Barremian-lower Aptian Castrillo de la Reina Formation, deposited in the Cameros Basin, north of the Maestrat Basin. This sauropod fauna at present comprises the rebbachisaurid Demandasaurus darwini and an undescribed titanosauriform—with somphospondylan affinities—which is distinct from Tastavinsaurus sanzi (Torcida Fernández-Baldor et al. 2013). The relationship between the sauropods, in particularly titanosauriforms, of the Maestrat and Cameros basins still remains poorly known. The systematic study of the Sant Antoni de la Vespa (Arcillas de Morella Formation, Maestrat Basin) and El Oterillo II (Castrillo de la Reina Formation, Cameros Basin) sauropods and the reassessment of El Canteret specimen (Arcillas de Morella Formation, Maestrat Basin) is necessary in order to better understand the evolutionary history of the titanosauriforms in the Early Cretaceous of the Iberian Peninsula.
5 Conclusions
Several specimens pertaining to Sauropoda, housed in the Museo Nacional de Ciencias Naturales (Madrid) and collected in the second half of the nineteenth century and in the first decades of the twentieth century, are described and discussed here in detail - some of them for the first time. These specimens, which are some of the first dinosaur specimens ever found in Spain, were collected from several outcrops in Morella (Castellón, Spain), in levels of the Arcillas de Morella Formation (upper Barremian, Maestrat Basin). Systematic study of this material has allowed us to identify indeterminate titanosauriforms, some of which have somphospondylan and ‘laurasiform’ affinities. However the monophyly and the phylogenetic position of ‘Laurasiformes’ within Titanosauriformes remains uncertain. Comparative analysis of two isolated sauropod humeri from Morella (MNCN 59703 and 68484) and another found in the second half on the twentieth century, indicates the possible presence of three titanosauriform taxa during the upper Barremian of the Maestrat Basin, two of which have somphospondylan affinities. The sauropod diversity in this Formation, particularly for the titanosauriforms, is higher than was previously known. However, the study of several new occurrences from the Arcillas de Morella Formation will be necessary in order to understand the evolution of this group during the Barremian-Aptian lapse in the Maestrat Basin, and the relationship of the Morella taxa with other contemporaneous sauropod faunas from both the Iberian Peninsula and the rest of the European territory.
References
Antunes, M. T., & Mateus, O. (2003). Dinosaurs of Portugal. Comptes Rendus Palevol, 2, 77–95. doi:10.1016/S1631-0683(03)00003-4.
Barco, J.L. (2009). Sistemática e implicaciones filogenéticas y paleobiogeográficas del saurópodo Galvesaurus herreroi (Formación Villar del Arzobispo, Galve, España). Unpublished PhD Thesis, Universidad de Zaragoza, Zaragoza, Spain, 389.
Bonaparte, J. F. (1986). Les dinosaures (Carnosaures, Allosaurides, Sauropodes, Cetiosaurides) du Jurassique Moyen de Cerro Cóndor (Chubut, Argentine). Annales de Paléontologie, 72, 247–386.
Bonaparte, J. F., González Riga, B. J., & Apesteguía, S. (2006). Ligabuesaurus leanzai gen. et sp. nov. (Dinosauria, Sauropoda), a new titanosaur from the Lohan Cura Formation (Aptian, Lower Cretaceous) of Neuquén, Patagonia, Argentina. Cretaceous Research, 27, 364–376. doi:10.1016/j.cretres.2005.07.004.
Borsuk-Bialynicka, M. (1977). A new camarasaurid sauropod Opisthocoelicaudia skarzynskii gen. n., sp. n. from the Upper Cretaceous of Mongolia. Palaeontologica Polonica., 37, 5–63.
Bover-Arnal, T., Moreno-Bedmar, J. A., Frijia, G., Pascual-Cebrian, E., & Salas, R. (2016). Chronostratigraphy of the Barremian-Early Albian of the Maestrat Basin (E Iberian Peninsula): integrating strontium-isotope stratigraphy and ammonoid biostratigraphy. Newsletters on Stratigraphy, 49, 41–68.
Calvo, J. O., González-Riga, B. J., & Porfiri, J. D. (2007). A new titanosaur sauropod from the Late Cretaceous of Neuquén, Patagonia, Argentina. Arquivos do Museu Nacional, 65, 485–504.
Canudo, J. I., Royo-Torres, R., & Cuenca-Bescós, G. (2008). A new sauropod: Tastavinsaurus sanzi gen. et sp. nov. from the Early Cretaceous (Aptian) of Spain. Journal of Vertebrate Paleontology, 28, 712–731. doi:10.1671/0272-4634(2008)28[712:ANSTSG]2.0.CO;2.
Carballido, J. L., Pol, D., Cerda, I., & Salgado, L. (2011a). The osteology of Chubutisaurus insignis del Corro, 1975 (Dinosauria: Neosauropoda) from the ‘middle’ Cretaceous of central Patagonia, Argentina. Journal of Vertebrate Paleontology, 31, 93–110. doi:10.1080/02724634.2011.539651.
Carballido, J. L., Rauhut, O. W. M., Pol, D., & Salgado, L. (2011b). Osteology and phylogeny relationships of Tehuelchesaurus benitezii (Dinosauria, Sauropoda) from the Upper Jurassic of Patagonia. Zoological Journal of the Linnean Society, 163, 605–662. doi:10.1111/j.1096-3642.2011.00723.x.
Carballido, J. L., Salgado, L., Pol, D., Canudo, J. I., & Garrido, A. (2012). A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Basin; evolution and biogeography of the group. Historical Biology, 24, 631–654. doi:10.1080/08912963.2012.672416.
Carballido, J. L., & Sander, P. M. (2014). Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria. Journal of Systematic Paleontology, 12(3), 335–387. doi:10.1080/14772019.2013.764935.
Casanovas, M. L., Santafé, J. V., & Sanz, J. L. (2001). Losillasaurus giganteus, un nuevo saurópodo del tránsito Jurásico-Cretácico de la cuenca de « Los Serranos » (Valencia, España). Paleontologia i Evolución, 32–33, 99–122.
Csiki, Z., Codrea, V., Jipa-Murzea, C., & Godefroit, P. (2010). A partial titanosaur (Sauropoda, Dinosauria) skeleton from the Maastrichtian of Nalat-Vad, Hateg Basin. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 258, 297–324. doi:10.1127/0077-7749/2010/0098.
D’Emic, M. D. (2012). The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society, 166(3), 624–671. doi:10.1111/j.1096-3642.2012.00853.x.
D’Emic, M. D. (2013). Revision of the sauropod dinosaurs of the Early Cretaceous Trinity Group, southern USA, with the description of a new genus. Journal of Systematic Palaeontology, 11, 707–726. doi:10.1080/14772019.2012.667446.
D’Emic, M. D., & Foreman, B. Z. (2012). The beginning of the sauropod dinosaur hiatus in North America: insights from the Lower Cretaceous Cloverly Formation of Wyoming. Journal of Vertebrate Paleontology, 32, 883–902. doi:10.1080/02724634.2012.671204.
Dal Sasso, C., Pierangelini, G., Famiani, F., Cau, A., & Nicosia, U. (2016). First sauropod bones from Italy offer new insights on the radiation of Titanosauria between Africa and Europe. Cretaceous Research, 64, 88–109. doi:10.1016/j.cretres.2016.03.008.
Díez Díaz, V., Suberbiola, X. P., & Sanz, J. L. (2013). Appendicular skeleton and dermal armour of the Late Cretaceous titanosaur Lirainosaurus astibiae (Dinosauria:Sauropoda) from Spain. Palaeontologia Electronica, 16.2.19A, 18.
Figueiredo, S., Rosina, P., & Figuti, L. (2015). Dinosaurs and other vertebrates from the Papo-Seco Formation (Lower Cretaceous) of southern Portugal. Journal of Iberian Geology, 41, 301–314. doi:10.5209/rev_JIGE.2015.v41.n3.47828.
Fuentes Vidarte, C., Meijide Calvo, M., Meijide Fuentes, F., & Meijide Fuentes, M. (2005). Fauna de vertebrados del Cretácico Inferior del Yacimiento de “Zorralbo” en Golmayo (Soria, España) (pp. 83–92). Número Extraordinario X: Revista Española de Paleontología.
Gàmez, D., Paciotti, P., Colombo, F., & Salas, R. (2003). La Formación Arcillas de Morella (Aptiense inferior), Cadena Ibérica oriental (España): caracterización sedimentológica. Geogaceta, 34, 191–194.
Gasulla Asensio, J. M. (2005). Los dinosaurios de Morella (Castellón, España): historia de su investigación. Revista española de Paleontología, N.E., 10, 29–38.
Gasulla, J. M., Escaso, F., Narváez, I., Ortega, F., & Sanz, J. L. (2015). A new sail-backed styracosternan (Dinosauria: Ornithopoda) from the Early Cretaceous of Morella, Spain. PLoS One, 10, e0144167. doi:10.1371/journal.pone.0144167.
Gasulla, J. M., Escaso, F., Ortega, F., & Sanz, J. L. (2014). New hadrosauriform cranial remains from the Arcillas de Morella Formation (lower Aptian) of Morella, Spain. Cretaceous Research, 47, 19–24. doi:10.1016/j.cretres.2013.10.004.
Gasulla, J.M, Ortega, F., Escaso, F., & Pérez-García, A. (2011a). Los yacimientos de vertebrados de la Formación Arcillas de Morella (Aptiense inferior). In A. Pérez-García, F. Gascó, J.M. Gasulla, F. Escaso (Eds.), Viajando a mundos pretéritos, Morella pp. 157–171.
Gasulla, J. M., Ortega, F., Pereda Suberbiola, X., Escaso, F., & Sanz, J. L. (2011b). Elementos de la armadura dérmica del dinosaurio anquilosaurio Polacanthus Owen, 1865, en el Cretácico Inferior de Morella (Castellón, España). Ameghiniana, 48, 508–519.
Gasulla, J. M., Ortega, F., Pérez García, A., & Sanz, J. L. (2009). Actividad paleontológica en la comarca de Els Ports a finales del siglo XIX y principios del XX [what does this XX mean?]. In B. Poza, A. Santos-Cubedo, B. Vila, & M. Suñer (Eds.), Dinosaurios lagartos terriblemente grandes (pp. 39–46). Fundación Blasco de Alagón: Un paseo por la exposición.
Gasulla, J. M., Ortega, F., Sanz, J. L., Escaso, F., Yagüe, P., & Pérez-García, A. (2012). Els vertebrats de la Formació Argiles de Morella (Aptià inferior, Cretaci Inferior). Nemus, 2, 15–27.
Gasulla, J. M., Sanz, J. L., Escaso, F., & P. Yagüe. (2008). Elementos de la cintura pélvica de dinosaurios saurópodos Titanosauriformes del Cretácico Inferior (Aptiense inferior) de Morella (Castellón). In J. I. Ruiz-Omeñaca, L. Piñuela, J. C. García-Ramos (Eds.), Libro de resúmenes de las 24ª Jornadas de la Sociedad Española de Paleontología (Colunga) (pp. 129–130).
Gomani, E. M. (2005). Sauropod dinosaurs from the Early Cretaceous of Malawi. Palaeontologia Electronica, 8, 1–37.
González Riga, B. J., Previtera, E., & Pirrone, C. A. (2009). Malarguesaurus florenciae gen. et sp. nov., a new titanosauriform (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Cretaceous Research, 30, 135–148. doi:10.1016/j.cretres.2008.06.006.
Gorscak, E., & O‘Connor, P. M. (2016). Time-calibrated models support congruency between Cretaceous continental rifting and titanosaurian evolutionary history. Biology Letters, 12, 20151047. doi:10.1098/rsbl.2015.1047.
Gorscak, E., O’Connor, P. M., Stevens, N. J., & Roberts, E. R. (2014). The basal titanosaurian Rukwatitan bisepultus (Dinosauria, Sauropoda) from the middle Cretaceous Galula Formation, Rukwa Rift Basin, southwestern Tanzania. Journal of Vertebrate Paleontology, 34, 1133–1154. doi:10.1080/02724634.2014.845568.
Hatcher, J.B. (1901). Diplodocus (Marsh): its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum, 1, 1–63.
Huene, F. von (1932). Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte. Monograph Geologie und Palaeontologie, 4, 1–361.
Janensch, W. (1950). Die wirbelsaule von Brachiosaurus brancai. Palaeontographica, Supplementbände, 7(3), 27–93.
Janensch, W. (1961). Die Gliedmaszen und Gliedmaszengürtel der Sauropoden der Tendaguru-Schichten. Palaeontographica, Supplementbände., 7(3), 177–235.
Kellner, A.W., & de Azevedo, S.A. (1999). A new sauropod dinosaur (Titanosauria) from the Late Cretaceous of Brazil. In Y. Tomida, T.H. Rich, P. Vickers-Rich (eds.), Proceedings of the Second Gondwanan Dinosaur Symposium. Toyko National Science Museum Monographs, Number 15. pp. 111–142
Kirkland, J. I., Alcalá, L., Loewen, M. A., Espílez, E., Mampel, L., & Wiersma, J. P. (2013). The basal Nodosaurid Ankylosaur Europelta carbonensis n. gen., n. sp. from the Lower Cretaceous (Lower Albian) Escucha Formation of Northeastern Spain. PLoS One, 8(12), e80405. doi:10.1371/journal.pone.0080405.
Lacovara, K. J., Lamanna, M. C., Ibiricu, L. M., Poole, J. C., Schroeter, E. R., Ullmann, P. V., et al. (2014). A gigantic, exceptionally complete titanosaurian sauropod dinosaur from southern Patagonia, Argentina. Scientific Reports, 4, 1–9. doi:10.1038/srep06196.
Lapparent, A. F. (1966). Nouveux gisements de reptiles Mesozoiques en Espagne. Notas y Comunicaciones del Instituto Geológico y Minero de España, 84, 103–110.
Lapparent, A. F., & Zbyszewski, G. (1957). Les dinosauriens du Portugal. Mémoires des Services Géologiques du Portugal, Nouvelle Série, 2, 1–63.
Le Loeuff, J. (1993). European titanosaurs. Revue de Paléobiologie, 7, 105–117.
Mannion, P. D., & Calvo, J. O. (2011). Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian-early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: implications for titanosaur systematics. Zoological Journal of the Linnean Society, 163, 155–181. doi:10.1111/j.1096-3642.2011.00699.x.
Mannion, P., Upchurch, P., Barnes, R. N., & Mateus, O. (2013). Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal Titanosauriformes. Zoological Journal of the Linnean Society, 168(1), 98–206. doi:10.1111/zoj.12029.
Mannion, P., Upchurch, P., Mateus, O., Barnes, R. N., & Jones, M. E. H. (2012). New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology, 10, 521–551. doi:10.1080/14772019.2011.595432.
Marsh, O. C. (1878). Principal characters of American Jurassic dinosaurs. Part I. American Journal of Science, 16, 411–416.
Martínez, R. D., Giménez, O., Rodríguez, J., Luna, M., & Lamanna, M. C. (2004). An articulated specimen of the basal titanosaurian (Dinosauria: Sauropoda) Epachthosaurus sciuttoi from the early Late Cretaceous Bajo Barreal Formation of Chubut province, Argentina. Journal of Vertebrate Paleontology, 24, 107–120. doi:10.1671/9.1.
Mateus, O., Araújo, R., Natário, C., & Castanhinha, R. (2011a). A new specimen of the theropod dinosaur Baryonyx from the early Cretaceous of Portugal and taxonomic validity of Suchosaurus. Zootaxa, 2827, 54–68.
Mateus, O., Jacobs, L. L., Schulp, A. S., Polcyn, M. J., Tavares, T. S., Neto, A. B., et al. (2011b). Angolatitan adamastor, a new sauropod dinosaur and the first record from Angola. Anais da Academia Brasileira de Ciências, 83, 1–13.
Mateus, O., Mannion, P. D., & Upchurch, P. (2014). Zby atlanticus, a new turiasaurian sauropod Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology, 34(3), 618–634. doi:10.1080/02724634.2013.822875.
McDonald, A. T., Espílez, E., Mampel, L., Kirkland, J. I., & Alcalá, L. (2012). An unusual new basal iguanodont (Dinosauria: Ornithopoda) from the Lower Cretaceous of Teruel, Spain. Zootaxa, 3595, 61–76.
McIntosh, J. S. (2005). The genus Barosaurus Marsh (Sauropoda, Diplodocidae). In T. Tidwell & K. Carpenter (Eds.), Thunder-Lizards: the sauropodomorph dinosaurs (pp. 38–77). Bloomington: Indiana University Press.
Mocho, P., Escaso, F., Gasulla, J.M., Galobart, A., Poza, B., Santos-Cubedo, A., Sanz, J.L., & Ortega, F. (2016a). A new titanosauriform sauropod from the Arcillas de Morella Formation (Early Cretaeous). In F. Torcida Fernández-Baldor, J.I. Canudo, P. Huerta, X. Pereda (eds.), Abstract book of the VII International Symposium about Dinosaurs Palaeontology and their Environment. Colectivo Arqueológico y Paleontológico de Salas, Salas de los Infantes, pp.97–98
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., Silva, B., & Ortega, F. (2016b). Turiasauria-like teeth from the Upper Jurassic of the Lusitanian Basin, Portugal. Historical Biology, 28, 861–880. doi:10.1080/08912963.2015.1049948.
Mocho, P., Royo-Torres, R., & Ortega, F. (2016c). New data of the Portuguese brachiosaurid Lusotitan atalaiensis (Sobral Formation, Upper Jurassic). Historical Biology. doi:10.1080/08912963.2016.1247447.
Mocho, P., Royo-Torres, R., & Ortega, F. (2014). Phylogenetic reassessment of Lourinhasaurus alenquerensis, a basal Macronaria (Sauropoda) from the Upper Jurassic of Portugal. Zoological Journal of the Linnean Society, 170, 875–916. doi:10.1111/zoj.12113.
Néraudeau, D., Allain, R., Ballèvre, M., Batten, D. J., Buffetaut, E., Colin, J. P., et al. (2012). The Hauterivian-Barremian lignitic bone bed of Angeac (Charente, south-west France): stratigraphical, palaeobiological and palaeogeographical implications. Cretaceous Research, 35, 1–14. doi:10.1016/j.cretres.2012.01.006.
Ortega, F., Escaso, F., Gasulla, J. M., Dantas, P., & Sanz, J. L. (2006). Los Dinosaurios de la Península Ibérica. Estudios Geológicos, 62(1–2), 219–240.
Ortega, F., Escaso, F., & Sanz, J. L. (2010). A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain. Nature, 467, 203–206. doi:10.1038/nature09181.
Otero, A. (2010). The appendicular skeleton of Neuquensaurus, a Late Cretaceous saltasaurine sauropod from Patagonia, Argentina. Acta Palaeontologica Polonica, 55, 399–426. doi:10.4202/app.2009.0099.
Owen, R. (1842). Report on British fossil reptiles. Reports of the British Association for the Advancement of Science, 11, 60–204.
Pereda Suberbiola, X., & Ruiz-Omeñaca, J. I. (2005). Los primeros descubrimientos de dinosaurios en España. Revista Española de Paleontología, 10, 15–28.
Pereda Suberbiola, X., Ruiz-Omeñaca, J. I., & Canudo, J. I. (2006). Los primeros descubrimientos de dinosaurios ibéricos. Naturaleza Aragonesa, 17, 4–10.
Pereda Suberbiola, X., Torcida, F., Izquierdo, L. A., Huerta, P., Montero, D., & Pérez, G. (2003). First rebbachisaurid dinosaur (Sauropoda, Diplodocoidea) from the early Cretaceous of Spain: palaeobiogeographical implications. Bulletin de la Société Géologique de France, 174, 471–479. doi:10.2113/174.5.471.
Pereda-Suberbiola, X., Ruiz-Omeñaca, J. I., Canudo, J. I., Torcida, F., & Sanz, J. L. (2012). Dinosaur faunas from the Early Cretaceous of Spain. In P. Godefroit (Ed.), Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems (pp. 378–407). Bloomington: Indiana University Press.
Pérez-García, A., Sánchez Chillón, B., & Ortega, F. (2009). Aportaciones de José Royo y Gómez al conocimiento sobre los dinosaurios de España. In: A. Pérez-García, B.C. Silva, E. Malafaia, F. Escaso (eds.), Paleolusitana. Torres Vedras, pp. 339–364.
Pérez-Moreno, B. P., Sanz, J. L., Buscalioni, A. D., Moratalla, J. J., Ortega, F., & Raskin-Gutman, D. (1994). A unique multitoothed ornithomimosaur from the Lower Cretaceous of Spain. Nature, 30, 363–367. doi:10.1038/370363a0.
Phillips, J. (1871). Geology of Oxford and the Valley of the Thames (p. 529). Oxford: Clarendon Press.
Poropat, S. F., Mannion, P. D., Upchurch, P., Hocknull, S. A., Kear, B. K., Kundrát, M., et al. (2016). New Australian sauropods shed light on Cretaceous dinosaur palaeobiogeography. Scientific Reports, 6, 34467. doi:10.1038/srep34467.
Poropat, S. F., Upchurch, P., Mannion, P. D., Hocknull, S. A., Kear, B. P., Sloan, T., et al. (2015). Revision of the sauropod dinosaur Diamantinasaurus matildae Hocknull et al. 2009 from the mid-Cretaceous of Australia: implications for Gondwanan titanosauriform dispersal. Gondwana Research, 27, 995–1033. doi:10.1016/j.gr.2014.03.014.
Powell, J. E. (1987). The Late Cretaceous fauna of Los Alamitos, Patagonia, Argentina Part VI. The titanosaurids. Revista del Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia’, 3, 147–153.
Powell, J. E. (1992). Osteologia de Saltasaurus loricatus (Sauropoda-Titanosauridae) del Cretacico Superior del Noroeste argentino. In J. L. Sanz & A. D. Buscalioni (Eds.), Los dinosaurios y su entorno biotico (pp. 165–230). Cuenca: Instituto Juan de Valdes, Serie Actas Academicas.
Ratkevich, R. (1998). New Cretaceous brachiosaurid dinosaur, Sonorasaurus thompsoni gen. et sp. nov., from Arizona. Journal of the Arizona-Nevada Academy of Sciences, 31, 71–82.
Riggs, E. S. (1903). Brachiosaurus altithorax, the largest known dinosaur. American Journal of Science, 15, 299–306.
Rose, P.J. (2007). A new titanosauriform sauropod (Dinosauria: Saurischia) from the Early Cretaceous of central Texas and its phylogenetic relationships. Palaeontologica Electronica, 10, 1–65.
Royo y Gómez, J. (1925). Resultados científicos obtenidos en su viaje por Francia; Suiza, Alemania y Bélgica. Boletín de la Real Sociedad Española de Historia Natural, 25, 114–117.
Royo y Gómez, J. (1926a). Los vertebrados del Cretácico español de facis weáldica. Boletín del Instituto Geológico y Minero de España, 47((7 de la 3ª serie)), 171–176.
Royo y Gómez, J. (1926b). Los descubrimientos de reptiles gigantescos en Levante. Boletín de la Sociedad Castellonense de Cultura, 7, 147–162.
Royo y Gómez, J. (1927a). Comunicación sobre el viaje realizado a Francia e Inglaterra, pensionado por la Junta de Ampliación de Estudios. Boletín de la Real Sociedad Española de Historia Natural, 27, 307–309.
Royo y Gómez, J. (1927b). Sur le faciès Wealdien d’Espagne. Comptes Rendus Sommaire des Séances de la Société Géologique de France, 11, 125–128.
Royo-Torres, R. (2009). El saurópodo de Peñarroya de Tastavins. Monograpfías Turonlenses, 6, 1–548.
Royo-Torres, R., Alcalá, L., & Cobos, A. (2012). A new specimen of the Cretaceous sauropod Tastavinsaurus sanzi from El Castellar (Teruel, Spain), and a phylogenetic analysis of the Laurasiformes. Cretaceous Research, 34, 61–83.
Royo-Torres, R., Cobos, A., & Alcalá, L. (2006). A giant european dinosaur and a new sauropod clade. Science, 314, 1925–1927. doi:10.1126/science.1132885.
Royo-Torres, R., Upchurch, P., Kirkland, J. I., Deblieux, D., Foster, J., Cobos, A., Rey, G., & Alcalá, L. (2016). Systematics of a new sauropod from the basal Yellow Cat Member (Early Cretaceous) from Doelling’s Bowl (Eastern, Utah, USA). In A. Farke, A MacKenzie, J. Miller-Camp (Eds.), Society of Vertebrate Paleontology, Octobre 2016, Abstracts of Papers, 76th Annual Meeting, Grand America Hotel, Salt Lake City Utah, Octobre 26 (pp. 215–215).
Royo-Torres, R., Upchurch, P., Mannion, P. D., Mas, R., Cobos, A., Gascó, F., et al. (2014). The anatomy, phylogenetic relationships and stratigraphic position of the Tithonian-Berriasian Spanish sauropod dinosaur Aragosaurus ischiaticus. Zoological Journal of the Linnean Society, 171, 623–655. doi:10.1111/zoj.12144.
Ruiz-Omeñaca, J. I. (2011). Delapparentia turolensis nov. gen. et sp., un nuevo dinosaurio iguanodontoideo (Ornithischia: Ornithopoda) en el Cretácico Inferior de Galve. Estudios Geológicos, 67, 83–110.
Ruiz-Omeñaca, J. I., & Canudo, J. I. (2003). Dinosaurios (Saurischia, Ornithischia) en el Barremiense (Cretácico inferior) de la Península Ibérica. Ciencias de la Tierra, 26, 269–312.
Ruiz-Omeñaca, J.I., Pereda Suberbiola, X., & Company, J. (2003). Juan Vilanova, José Royo y Gómez y los dinosaurios de Morella en las colecciones del Museo Nacional de Ciencias Naturales de Madrid. In XIX Jornadas de Paleontología, Morella, pp. 150–151.
Ruiz-Omeñaca, J. I., & Pereda-Suberbiola, X. (1999). Un documento inédito de Royo y Gómez sobre los dinosaurios del Levante. Temas Geológico-Mineros, 26, 111–112.
Salgado, L., Coria, R. A., & Calvo, J. O. (1997). Evolution of titanosaurid sauropods. I: phylogenetic analysis based on the postcranial evidence. Ameghiniana, 34, 332.
Sánchez-Hernández, B., Benton, M. J., & Naish, D. (2007). Dinosaurs and other fossil vertebrates from the Late Jurassic and Early Cretaceous of the Galve area, NE Spain. Palaegeography, Palaeoclimatology, Palaeoecology, 249, 180–215. doi:10.1016/j.palaeo.2007.01.009.
Santos-Cubedo, A., Santisteban, C. de, & Galobart, A. (2009). New dinosaur findings from Arcillas de Morella Formation (Spain). In P. Godefroit, O. Lambert (eds.), Tribute to Charles Darwin and Bernissart Iguanodons: New Perspectives on Vertebrate Evolution and Early Cretaceous Ecosystems (p. 87) Brussels.
Santos-Cubedo, A., Poza, B., Suñer, M., & de Santisteban, C. (2010). New remains of a titanosaur (Dinosauria: Sauropoda) from the Early Cretaceous of Spain. Journal of Vertebrate Paleontology Supplement to Number, 30, 157A.
Santucci, R. M., & Arruda-Campos, A. C. de (2011). A new sauropod (Macronaria, Titanosauria) from the Adamantina Formation, Bauru Group, Upper Cretaceous of Brazil and the phylogenetic relationships of Aeolosaurini. Zootaxa, 3085, 1–33.
Sanz, J.L. (1984). Las faunas españolas de dinosaurios. In I Congreso Español de Geología (pp. 497–506) Segovia.
Sanz, J. L. (1996). José Royo y Gómez y los dinosaurios españoles. Geogaceta, 19, 167–168.
Sanz, J. L., Bonaparte, J. F., & Lacasa, A. (1988). Unusual Lower Cretaceous birds from Spain. Nature, 331, 433–435.
Sanz, J. L., & Buscalioni, A. D. (1992). A new bird from the Early Cretaceous of Las Hoyas, Spain, and the early radiation of birds. Palaeontology, 35, 829–845.
Sanz, J.L., Casanovas, M.L., & Santafé, J.V. (1982). Paleontología. In J.V. Santafé, M.L. Casanovas, J.L. Sanz, S. Calzada (coord.), Geología y Paleontología (Dinosaurios) de las Capas rojas de Morella (Castellón, España). Diputación Provincial de Castellón y Diputación de Barcelona (pp. 71–169) Spain.
Sanz, J. L., Chiappe, L. M., Pérez-Moreno, B. P., Buscalioni, A. D., Moratalla, J. J., Ortega, F., et al. (1996). An Early Cretaceous bird from Spain and its implications for the evolution of avian flight. Nature, 382, 442–445. doi:10.1038/382442a0.
Seeley, H. G. (1887). On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London, 43, 165–171.
Segura Barreda, J. (1868). Morella y sus aldeas. Tomo 2. Imprenta de Javier Soto, F. 490 p.
Sereno, P. C., Beck, A. L., Dutheil, D. B., Larssen, H. C. E., Lyon, G. H., Moussa, B., Sadleir, R. W., Sidor, C. A., Varricchio, D. J., Wilson, G. P., & Wilson, J. A. (1999). Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs. Science, 286, 1342–1347. doi:10.1126/science.286.5443.1342.
Tidwell, V., Carpenter, K., & Brooks, B. (1999). New sauropod from the Lower Cretaceous of Utah. Oryctos, 2, 21–37.
Tidwell, V., Carpenter, K., & Meyer, S. (2001). New titanosauriform (Sauropoda) from the Poison Strip Member of Cedar Mountain Formation (Lower Cretaceous), Utah. In D. Tanke & D. K. Carpenter (Eds.), Mesozoic Vertebrate Life (pp. 139–165). Bloomington: Indiana University Press.
Torcida Fernández-Baldor, F.T., Canudo, J.I., Contreras, R., Huerta, P., Montero, D., & Urién, V. (2013). Description of a dorsal vertebra from the sauropod from El Oterillo II (Burgos, Spain). In: F. Torcida Fernández-Baldor, P. Huerta (eds.), Abstract book of the VI International Symposium about Dinosaurs Palaeontology and their Environment. Colectivo Arqueológico y Paleontológico de Salas (pp. 130–131) Salas de los Infantes.
Torcida Fernández-Baldor, F. T., Canudo, J. I., Huerta, P., Montero, D., Pereda Suberbiola, X., & Salgado, L. (2011). Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica, 56, 535–552. doi:10.4202/app.2010.0003.
Tschopp, E., Mateus, O., & Benson, R. (2015). A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ, 3, e857. doi:10.7717/peerj.857.
Upchurch, P. (1995). Evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London. Series B, 349, 365–390. doi:10.1098/rstb.1995.0125.
Upchurch, P., Barrett, P. M., & Dodson, P. (2004). Sauropoda. In D. B. Weishampel, P. Dodson, P. Dodson, & H. Osmólska (Eds.), The Dinosauria II (pp. 259–322). Berkeley: University of California.
Upchurch, P., Mannion, P. D., & Taylor, M. P. (2015). The Anatomy and phylogenetic relationships of “Pelorosaurus” becklesii (Neosauropoda, Macronaria) from the Early Cretaceous of England. PLoS One, 10, e0125819. doi:10.1371/journal.pone.0125819.
Upchurch, P., & Martin, J. (2003). The anatomy and taxonomy of Cetiosaurus (Saurischia, Sauropoda) from the Middle Jurassic of England. Journal of Vertebrate Paleontology, 23, 208–231. doi:10.1671/0272-4634(2003)23[208:TAATOC]2.0.CO;2.
Verdú, F. J., Royo-Torres, R., Cobos, A., & Alcalá, L. (2015). Perinates of a new species of (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain). Cretaceous Research, 56, 250–264. doi:10.1016/j.cretres.2015.05.010.
Vilanova y Piera, J. (1872). Compendio de Geología (p. 588). Madrid: Imprenta de Alejandro Gómez Fuentenebro.
Vilanova y Piera, J. (1873). Participa a la Sociedad hallarse hace algún tiempo en posesión de los únicos restos hasta ahora conocidos, en España, de Iguanodon. Anales de la Sociedad Española de Historia Natural, Actas, 2, 8.
Villanueva-Amadoz, U., Santisteban, C., & Santos-Cubedo, A. (2014). Age determination of the Arcillas de Morella Formation (Maestrat Basin, Spain). Historical Biology, 27, 389–397. doi:10.1080/08912963.2013.874422.
Whitlock, J. A. (2011). A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society, 161, 872–915. doi:10.1111/j.1096-3642.2010.00665.x.
Wilson, J. A. (2002). Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society, 136, 217–276. doi:10.1046/j.1096-3642.2002.00029.x.
Wilson, J. A., & Allain, R. (2015). Osteology of Rebbachisaurus garasbae Lavocat, 1954, a diplodocoid (Dinosauria, Sauropoda) from the early Late Cretaceous–aged Kem Kem beds of southeastern Morocco. Journal of Vertebrate Paleontology, 35, e1000701. doi:10.1080/02724634.2014.1000701.
Wilson, J.A., Barrett, P.M., & Carrano, M.T. (2011). An associated partial skeleton of Jainosaurus cf. septentrionalis (Dinosauria: Sauropoda) from the Late Cretaceous of Chhota Simla, Central India. Palaeontology, 54, 981–998. doi:10.1111/j.1475-4983.2011.01087.x.
Wilson, J. A., & Sereno, P. C. (1998). Early evolution and higher-level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology, 18(suppl. 2), 1–68. doi:10.2307/3889325.
Yagüe, P., Upchurch, P., Sanz, J.L., & Gasulla, J.M. (2001). New Sauropod material from the Early Cretaceous of Spain. In: 49 Annual Symposium of Vertebrate Paleontology and Comparative Anatomy, Yorkshire Museum.
Acknowledgements
This work was supported by “Fundação para a Ciência e Tecnologia” (Portugal) under grant SFRH/BD/68450/2010 PhD scholarship, Haaga Family Postdoctoral Fellow, and by the Ministerio de Economía y Competitividad (FPDI-2013-18986). This research received support from the Synthesys Project (http://synthesys3.myspecies.info/) which is financed by the European Community Research Infrastructure Action under the FP7 (FR-TAF-5072) and (DE-TAF-6138). We want to thank the following people for allowing us access to specimens in their care: B. Sánchez Chillón, P. Pérez Dios and S. Fraile (MNCN, Spain); M. Ramalho and R. Silva (MG, LNEG, Portugal); R. Allain (MNHN, France); S. Chapman (NHMUK, UK); D. Schwarz (HNM, Germany); E. Howlett (OUMNH, UK); the Herrero family (MPG, Spain); and L. Alcalá (FCPT-Dinópolis, Spain). We are also grateful to F. Escaso and I. Narváez (GBE-UNED, Spain) and J.L. Sanz (UAM) for the discussion about the upper Barremian fauna of Morella. We also appreciate the critical comments of S.F. Poropat and J.L. Carballido and the editors F. Torcida Fernández-Baldor and J.I. Canudo for suggested improvements to the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mocho, P., Pérez-García, A., Gasulla, J.M. et al. High sauropod diversity in the upper Barremian Arcillas de Morella Formation (Maestrat Basin, Spain) revealed by a systematic review of historical material. J Iber Geol 43, 111–128 (2017). https://doi.org/10.1007/s41513-017-0012-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-017-0012-8