
Abstract
The tumor microenvironment (TME) has been recognized as an integral component of malignancies in breast and prostate tissues, contributing in confounding ways to tumor progression, metastasis, therapy resistance, and disease recurrence. Major components of the TME are immune cells, fibroblasts, pericytes, endothelial cells, mesenchymal stroma/stem cells (MSCs), and extracellular matrix (ECM) components. Herein, we discuss the molecular and cellular heterogeneity within the TME and how this presents unique challenges and opportunities for treating breast and prostate cancers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Breast and prostate cancers are leading causes of cancer-related death in women and men, respectively, worldwide. In fact, prostate cancer is the second most common cancer among men after non-melanoma skin cancer in the USA (https://www.cdc.gov/cancer/). While mortality in these reproductive malignancies has been significantly reduced over the past several decades by applying early diagnostic biomarker discovery and targeted therapeutic approaches, therapy resistance, metastasis, and/or tumor relapse remain critical hurdles preventing successful treatment of all breast and prostate cancer patients.
Hanahan and Weinberg defined the hallmarks of cancer as a multistep process that includes biological functions such as sustaining proliferative signaling, evading growth suppression, resisting cell death, enabling replicative immortality, inducing angiogenesis, activating invasion/metastasis, acquiring therapy resistance, and escaping immune system control [39]. Solid tumors of the breast and prostate usually contain heterogeneous populations of neoplastic cells and collections of tissue-specific resident and recruited stromal cell types, which together form the tumor microenvironment (TME) [8, 38, 46, 66, 84, 86]. The normal breast ductal epithelium consists of a luminal epithelial cell layer surrounded by myoepithelial cells attached to the basement membrane. Several studies demonstrated that components of the TME play critical roles in mammary duct morphogenesis as well [46, 76, 89]. The cellular architecture of the prostate is quite similar to that of the breast in that it also consists of ducts with epithelial luminal and basal layers. As in the breast, the structure of these prostate ductal epithelial layers as they become neoplastic is strongly influenced by the TME [8, 24, 34, 79, 84]. In cancer of both the breast and prostate, components of TME vary greatly between and within each patient contributing to great disease diversity and difficulty in prescribing the optimal treatment plans for each patient.
In this regard, it is well established that bidirectional communication exists between cells and their microenvironment during both normal tissue homeostasis and tumor growth/metastasis. Importantly, the interactions between tumor cells and the associated stromal cells represent an opportune relationship that influences not only disease initiation and progression but also patient prognosis. Historically, cancer prognosis was thought to primarily depend upon the acquisition of aberrant mutations in tumor cells causing them to be governed by genetic and epigenetic alterations of neoplastic cells during the development/progression of a tumor. More recently, however, the TME has emerged as a significant and an equally important determinant of tumor behavior and pathogenesis. It is speculated that the TME also continues to change and evolve as a given tumor evolves in response to systemic changes (e.g., therapy-induced stresses). Further research into the specifics outlined here, in the setting of both breast and prostate cancers, is certain to reveal the dynamic and highly adaptable TME, helping scientists and clinicians to better diagnose and treat these malignancies to improve patient outcomes.
Tumor Cell Interactions with the Extracellular Matrix
Composition
The extracellular matrix (ECM) is a complex mixture of structural proteins, glycoproteins, and proteoglycans that provide essential physical scaffolds to maintain tissue structure [101]. The ECM regulates tissue development and homeostasis whereas, dysregulation of ECM functions promote neoplastic progression. Common matrix proteins embedded in breast and prostate cancers include fibrillar collagens, fibronectin, specific laminins, and proteoglycans [43]. Furthermore, the ECM represents an abundant source of soluble factors such as growth factors, angiogenic factors, cytokines and chemokines, proangiogenic platelets, and bone marrow-derived cells (BMDCs) [52]. Thus, the ECM provides a dynamic and complex network for cancer development, progression, and metastasis mediated by extensive reorganization of ECM and increasing matrix stiffness [1, 74, 97, 101].
Several studies [7, 9, 42, 70, 74, 99] suggest that many ECM proteins play a major functional role in breast and prostate cancer progression and metastasis. Further, a number of the induced ECM proteins take part as essential components of metastatic niches and promote stem/progenitor signaling pathways and survival of cells within the metastatic niche. ECM remodeling enzymes also lead to changes in the matrix structure and matrix biomechanical properties [43]. Collagens are one of the most abundant fibrous proteins in the extracellular matrix. During cancer invasion and metastasis, collagen I fibers are deposited and make tight cross-linked structures with other structural matrix proteins such as elastins, laminins, or fibronectin. Crosslinking of collagen I with ECM modifying enzymes create more rigid phenotype of the whole tumor.
In breast tumors, the presence of dense clusters of collagen fibrils indicates increased matrix stiffness, which correlates with poor patient survival. It was found that TWIST1 expression drives essential mechano-mediated changes in the TME that promote epithelial-mesenchymal transition (EMT) in response to increasing matrix stiffness [97]. High matrix stiffness induces nuclear translocation of TWIST1 and releases TWIST1 from its cytoplasmic binding partner G3BP2 [97]. The study suggests that TWIST1-G3BP2, a mechanotransduction pathway, actually responds to biomechanical signals from the tumor microenvironment to drive EMT, invasion, and metastasis. Another study has reported [42] that stiffness of the ECM potently modulates the repertoire of prolactin signals in human ERα-positive breast cancer cells. In the context of increased tumor invasion, stiff matrices can activate cross-talk signals between Janus Kinase (JAK) 2/STAT5 and pro-tumorigenic extracellular regulated kinase (ERK1/2). Several studies have also shown that extracellular matrix stiffness regulates prostate cancer cell behavior including tumor cell migration and aggressiveness [6, 87, 106].
Physical Properties
The ECM extracellular matrix has been shown to confer not only biochemical but also biophysical cues including matrix rigidity to the TME that significantly alters cellular behavior such as cell proliferation, stem cell properties, and metastatic growth [90]. A recent study has characterized the independent roles of mechanical, structural, and adhesion characteristics of 3D hydrogels and found that human breast cancer invasion and aggressive behavior correlate with ECM stiffening and immune cell infiltration. The study undertook a biophysical and biochemical assessment of stromal-epithelial interactions in noninvasive, invasive, and normal adjacent human breast tissue and in breast cancers of enhanced aggressive subtype. The data revealed that excessive collagen deposition and progressive linearization/thickening of collagen fibers correlated with human breast cell transformation. When stiffness of the stroma and cellular mechano-signaling were the highest, the linearization of collagen and tissue birefringence were also. Further, the greatest number of infiltrating macrophages and the highest level of TGF-β signaling were also observed within the cells at the invasive front [9]. Additionally, it was found that stiffened substrates aberrantly engage cellular mechanosignaling networks in cells, which promotes the assembly of invadosomes and lamellae for cell invasion and migration [45]. Finally, the impact of mechanotransduction and YAP-dependent matrix remodeling is required for the generation and maintenance of cancer-associated fibroblasts (CAFs) [16].
Remodeling
Several studies have been done to investigate the ECM remodeling within the TME on cancer progression. A study showed that pleiotrophin (PTN, Ptn), an 18-kDa secretory cytokine, could induce remodeling of the ECM through stimulation of stromal cells and is significantly associated with features of cancer progression [20]. It was observed that in a co-culture model of equal numbers of NIH 3T3 stromal fibroblasts and MCF-7-Ptn cells, PTN secreted from MCF-7-Ptn cells could induce a more malignant MCF-7-Ptn breast cancer cell phenotype via extensive remodeling of the MCF-7-Ptn/NIH 3T3 cell microenvironment. Further, PTN secretion could upregulate the expression of markers of aggressive breast cancers, including PKCδ and matrix metalloproteinase-9 (MMP-9) in both MCF-7-Ptn and NIH 3T3 cells. A growing body of evidence also indicates that interactions between neoplastic cells and tumor-associated macrophages (TAMs) in the TME are crucial in promoting tumor cell invasion and progression through remodeling [61]. In this study, it was shown that interactions between mouse breast tumor cells and TAMs remodel ECM, leading to the upregulation of proto-oncogene Fra-1, a member of the FOS family of transcription factors. In turn, Fra-1 initiates activation of the IL-6/JAK/Stat3 signaling pathway, inducing a malignant switch in breast tumor cells and increasing the release of proangiogenic factors MMP-9, vascular endothelial growth factor (VEGF), and transforming growth factor-beta (TGF-β). Another study showed that MMP-9 has different effects on breast cancer progression depending on whether insulin-like growth factor binding proteins (IGFBPs) are expressed [50].
Mesenchymal stem cells (MSCs) may also play a critical role in ECM remodeling. It was found that co-culture of MSCs with breast cancer cells causes upregulation of lysyl oxidase (LOX), a collagen crosslinker that promotes breast cancer progression. During invasion/metastasis, cancer cells must navigate and remodel dense ECM for migration through the 3D TME. Two major types of migration (i.e., amoeboid and mesenchymal) are utilized by individual cancer cells. Amoeboid migration is characterized by rounded cells that circumnavigate ECM without the use of adhesion proteins or matrix degradation. On the other hand, through mesenchymal migration, cells elongate, establish integrin-mediated adhesion to the ECM, and degrade ECM with MMPs [67].
Signaling
Matrix stiffness induces integrin clustering to enhance growth factor-dependent ERK activation. Integrin clustering can enhance PI3K signaling to regulate invasion of a premalignant mammary epithelium in vitro and tumor progression. Cytochrome P450 (CYP) 4Z1, a novel CYP4 family member, is overexpressed in human mammary carcinoma and associated with high-grade tumors and poor prognosis. It was found that CYP4Z1 overexpression promotes tumor angiogenesis and growth in breast cancer. Increased CYP4Z1 expression promotes tumor angiogenesis and growth in breast cancer, partly via PI3K/Akt and ERK1/2 activation [104]. ECM stiffness enhances cancer invasion and aggressive tumor cell behavior through TGF-β signaling and immune cell infiltration [1, 19]. ECM signaling with associated immune cells also influences the cancer stem cell niche. It was demonstrated that TAMs could create a CSC-niche via juxtacrine signaling with cancer stem cells (CSCs) [59].
Adhesion
Cancer cell invasion and dissemination are regulated by the matrix adhesion molecules. It was found TME signals can induce dissemination of normal and malignant epithelial cells without requiring the fibrillar structure of collagen I or containing collagen I-specific adhesion sequences [9]. Further, metastatic tumors preferentially disseminate in specific ECM microenvironments in which breaks in the basement membrane can induce invasion and dissemination through the direct contact between cancer cells and collagen I [70]. Further, alterations in the mechanical environment of the surrounding ECM induce the tumor cells to regulate secretion, which in turn, may influence cell adhesion/dissemination. A recent study reports that extracellular matrix rigidity-dependent sphingosine-1-phosphate (SIP) secretion regulates metastatic cancer cell invasion and adhesion in breast cancer [49].
Carcinomas commonly invade as a cohesive multicellular unit, usually known as a process of collective invasion [33, 37]. Collective invasion of breast cancer requires a conserved basal and epithelial program and heterotypic interactions between epithelial subpopulations [21]. A study was conducted using three-dimensional (3D) organoid assays to identify the most invasive cancer cells in primary breast tumors. It was found that collective invasion was led by specialized cancer cells, which prominently expressed basal epithelial genes, such as cytokeratin-14 (K14) and p63. Further, it was found that K14+ cells led collective invasion in the major human breast cancer subtypes. Importantly, luminal cancer cells were observed to convert phenotypically to invasive after induction of basal epithelial genes. On the other hand, during collective invasion, ductal carcinoma cells physically connect with expressed cadherins (ecadherin and P-cadherin), tight (JAM-A, claudin-3, and -4), and immunoglobulin-based junctions (ALCAM and L1-CAM). But most of these cell–cell junction proteins are downregulated in invading carcinoma cells, with the loss of E-cadherin. Most invasive carcinomas invade collectively as thin multicellular sheets or strands (in single-file patterns) which are parallel to collagen bundles. Although candidate adhesion mechanisms for multicellular cohesion within single file E-cadherin structures include N-cadherin and other cadherins, their mechanical and signaling contribution still need to be investigated [85].
Tumor Cell Interactions with the Mesenchyme
Fibroblasts
Fibroblasts are derived from primitive mesenchyme with elongated, spindle-like morphology and responsible for structural integrity of most connective tissues. Fibroblasts are predominantly multifunctional cell types involved in the synthesis and degradation of ECM and basement membrane components, epithelial homeostasis, proliferation, differentiation, and modulation of immune responses [11, 15, 65, 71, 77, 92]. Several studies suggest that normal fibroblasts can promote tumor growth, progression, and metastasis in breast and prostate cancers [71, 83, 92]. It was reported that tumor-derived osteopontin reprograms normal mammary fibroblasts through paracrine signaling to promote inflammation and breast tumor growth. Further, fibroblasts recruited, tumor-infiltrating regulatory T cells stimulate tumor metastasis through RANKL-RANK signaling. Therefore, fibroblasts are disrupted to promote tumorigenesis and those tumor supportive fibroblasts are defined as CAFs. CAFs are activated subpopulations of stromal fibroblasts, many of which express the myofibroblast marker alpha-smooth muscle actin (α-SMA). As such, it has been reported that isolation of normal and CAFs from fresh tissues (mouse and human) by fluorescence activated cell sorting (FACS) revealed that PDGFRα is abundantly expressed as surface markers by both normal fibroblasts and CAFs [83]. Therefore, in the TME, CAFs have been an ongoing challenge due to the lack of CAF-specific markers and the vast heterogeneity of these cells. Accumulating evidence support that CAFs facilitate tumor initiation, growth and progression [15, 17, 77], and tumor invasion and metastasis by mediating tumor-enhancing inflammation [31]. The data suggests that CAFs strongly promote the development of an aggressive cancer cell phenotype [28]. It was found by differential expression profile analysis that three types of microRNAs (miRs -21, -378e, and -143) increased in exosomes from breast CAFs as compared from normal fibroblasts. In this study, cancer cells (BT549, MDA-MB-231, and T47D lines) exposed to CAF exosomes or transfected with those miRs showed a significant increase in tumorspheres, stem cell and epithelial-mesenchymal transition (EMT) markers, and anchorage-independent cell growth. Current research has focused to understand how CAFs originate and maintain their phenotype in the TME. Studies show that MSCs and matrix remodeling influence the origin and help to maintain the CAF phenotype in the TME of breast cancer [16, 31, 67]. Fibroblasts were isolated from different stages of breast cancer and were analyzed for molecular and biological functions. These analyses revealed that activation of the YAP transcription factor is a signature feature of CAFs. Remodeling of the ECM and promotion of cancer cell invasion requires the actomyosin cytoskeleton. YAP regulates the expression of several cytoskeletal regulators; matrix stiffening further enhances YAP activation, thus establishing a feed-forward self-reinforcing loop that helps to maintain the CAF phenotype [16].
The TME of breast and prostate cancer is regulated by subpopulations of cells including fibroblasts, CAFs, stromal fibroblasts, CSCs, MSCs, and immune cells through networks of cytokines and growth factors. Accumulating evidence suggests that both stromal fibroblasts and CSCs mediate tumor growth and metastasis [10, 30, 35, 63, 93]. Since these components have a direct influence in the TME of these reproductive cancers, they represent attractive targets for therapeutic development.
Mesenchymal Stem Cells
An important component of breast TME are mesenchymal stem cells or multipotent mesenchymal stromal cells (MSCs), usually found in the bone marrow and in adipose tissue. They represent the adult stem cells that are able to differentiate into osteogenic, adipogenic, myogenic, and chondrogenic lineages [75]. Further, MSCs express stromal cell markers reported as CD73, CD105, and CD90 in the absence of hematopoietic and endothelial cell markers (CD34, CD31, CD45, and CD14) [27]. Normally, MSCs are recruited to injured areas or hypoxic tumor microenvironments and this homing of MSCs to tumors is reported as the earliest phenomena of MSC-cancer interactions. Contributions of MSCs within the tumor microenvironment allow tumor cells to exhibit altered biological functions. Accumulating evidence has supported that MSCs play critical roles in tumor development and progression, by increasing stemness of tumor cells, stimulating tumor cell migration, promoting angiogenesis, supporting immune responses, and inducing drug resistance. Additionally, MSCs increase the metastatic potential of tumor cells by advancing their motility and invasiveness and thereby play a role in the nature of a metastatic niche at the secondary site [14, 28, 36, 61, 67, 100]. Therefore, a comprehensive knowledge on the biology and mechanism of interaction between MSCs and TME of breast and prostate cancer is essential for the development of therapeutics that capitalize on vulnerabilities in these tumor cell-MSC interplay.
A recent study shows that MSCs in the TME has an impact on breast cancer progression by creating a pro-inflammatory milieu within the tumor [14]. This study identified six putative oncomiRs, strongly correlating with poor overall patient survival. Out of the six candidates, miR-1246 was upregulated in aggressive breast cancer subtypes and expressed at highest levels in MSCs. Functionally, miR-1246 induced the release of pro-inflammatory mediators IL-6, CCL2, and CCL5 in MSCs, and increased NF-κB activity. Further, incubation with conditioned medium (CM) of MSC overexpressing miR-1246 increased Stat3 phosphorylation in breast epithelial (MCF10A) and cancer cells (SK-BR-3, MCF7, T47D). In human breast cancer cells, metastasis is promoted by mesenchymal stem/multipotent stromal cells (MSC) with increased collagen deposition. It has been reported that the collagen receptor discoidin domain receptor 2 (DDR2) is concordantly upregulated in metastatic breast cancer and MSCs and also maintains a fibroblastic phenotype by activation of DDR2 signaling in breast cancer cells [36].
MSCs also support CSCs through various kinds of interactions and cell fusion to form hybrid tumor cells [69]. Since plasticity is a fundamental feature of MSCs, MSCs can also inhibit tumors by activating various MSC-based signaling pathways. Understanding the mechanism of MSC interacted with tumors, TME, and inflammatory TME will reveal new targets for cancer therapy.
Signaling
Interactions between tumor cells and mesenchymal cells involves a number of MSC-secreted signaling molecules that stimulate various signaling pathways leading to cell growth and regulation of apoptosis in the tumor cells. Cross talk between tumor-MSC in the TME is complex in the case of both normal development and tumor pathogenesis. This change in the cell phenotype is induced by contextual signals that epithelial cells receive from their microenvironment. Signals from tumor cells can stimulate MSCs as well as MSCs secreted factors involved in tumor initiation and early tumor growth. A study shows that paracrine and autocrine signals induce and maintain mesenchymal and stem cell states in the breast [81]. Spontaneously arising mesenchymal subpopulations (MSP) of cells were isolated from immortalized human mammary epithelial cells (HMECs) and were analyzed to understand how signals coordinate in the breast TME. It was revealed that a set of extracellular signals operated in a paracrine manner to induce entrance of HMLE cells into the mesenchyme/ SC state and subsequently function as autocrine factors. A recent study, however, suggests that paracrine NF-κB activation promotes the initiation of Notch in basal-type breast cancer cells. Several studies identified MSCs as signal-sending cells of Notch signaling whereas breast cancer cells were able to receive the signals. Blocking the Notch signal during co-culture could reduce the expression of MSC marker CD90 and thus suggest a functional role of this pathway in interaction of MSC-tumor cells [107].
Increased collagen deposition by breast cancer (BC)-associated MSCs promotes metastasis, but the exact mechanisms still need to be investigated. A recent study has reported that the collagen receptor discoidin domain receptor 2 (DDR2) is essential for stromal-breast cancer communication. In MSCs isolated from human BC metastasis, DDR2 induces activation of DDR2 signaling and maintains a fibroblastic phenotype with collagen deposition. Loss of DDR2 in MSCs inhibits DDR2 phosphorylation in BC cells, alignment, migration, and metastasis [36].
MSCs support CSC development and enhance the proportions of CSC in tumors and promote EMT by activation of various signaling pathways. Further, EMT confers on cancer cells mesenchymal traits and an ability to enter the CSC state. Recent studies show that breast tumor progression and EMT are activated by ERK/GSK-3β/Snail [53] and both NF-κB and β-catenin signaling [62] mediated by MCP-1 and VEGF/NPR, respectively. In this context, it may speculate that ERK/GSK-3β/Snail and NF-κB and β-catenin signaling may induce MSCs to secret tumor-initiating growth factors that stimulate sphere formation or growth of CSC-like cell. Although the interactions between MSCs, CSCs, and their surrounding microenvironments are poorly understood, it has been shown by quantitative proteomic profiling that tumor-associated monocytes and macrophages (TAMs) create a CSC-niche via juxtacrine signaling with CSCs [58]. The EMT program upregulates the expression of CD90/Thy1 and EphA4 receptors, which allow the physical interactions of CSCs with TAMs by directly binding with their respective counter-receptors on these cells. Further, the EphA4 receptor on the carcinoma cells activates Src and NF-κB in which the signaling of NF-κB induces the secretion of a variety of cytokines. Cytokines from CSCs and MSCs may help to maintain the stem-cell state.
Exosomes
Exosomes are small vesicles formed in vesicular bodies with a diameter of 30–100 nm with a distinctive “cup” or “dish” morphology that play an important role in the TME of breast cancer. Exosomes in the TME are considered as potential transporters and biomarkers since they can contain microRNAs, mRNAs, DNA fragments, and proteins, and transfer from a donor cell to recipient cells [95]. Exosome-derived tumor cells are called tumor-derived (TD) exosomes, and accumulating evidence suggest that TD exosomes can construct a suitable TME to support tumor proliferation, angiogenesis, invasion, and metastasis [13, 23, 48, 60] through exosome-mediated signaling. Breast cancer-derived exosomes can directly suppress T cell proliferation and inhibit NK cell cytotoxicity, and thereby suppress the anticancer immune response in premetastatic organs [98]. Studies show that the small GTPases RAB27A and RAB27B regulate exocytosis of multivesicular endosomes, which lead to exosome secretion, in human HeLa cells. Further, Rab27a maintains both exosome-dependent and exosome-independent mechanisms, modifies TME, and promotes tumor progression [13]. A recent study examined the influence of breast cancer-derived exosomes on MSCs by treating adipose tissue-derived MSCs (ADSCs) with TD exosomes. The exosome-treated ADSCs exhibited the phenotypes of tumor-associated myofibroblasts with enhanced expression of α-SMA, tumor-promoting factors SDF-1, VEGF, CCL5, and TGFβ. This study suggests that tumor-derived exosomes can promote tumor progression and metastasis by switching MSCs within the tumor stroma into tumor-associated myofibroblasts. Another study showed that fibroblast-secreted exosomes mobilize autocrine Wnt-PCP signaling to drive breast cancer cell invasive behavior [60]. Exosomes can deliver invasion-potentiating microRNA (miRNA) and thereby decrease sensitivity of breast cancer cells to chemotherapeutic drugs [64, 102]. A study has shown that the enrichment of a specific cargo secreted from breast cancer-derived exosomes can promote cell survival and thereby potentially provide a marker of chemotherapy resistance [51]. Exosomes can be mutually exchanged by tumor cells and adjacent cell populations in the tumor microenvironment [44]. Exosome levels in the serum of breast cancer patients are higher when compared to normal subjects. Further, breast cancer cells secrete exosomes with specific capacity for cell-independent miRNA biogenesis, which is absent in normal cell-derived exosomes [68]. Several studies show that some miRNAs such as miR-222/223 from MSC-released exosomes promote dormancy/quiescence and drug resistance in a subset of breast cancer [12, 72, 80]. It is reported that internalization of MSC-derived exosomes by breast and ovarian cancer cells provide tumor cells with new properties and functions through the expression of MMP2 and ecto-5′-nucleotidase (CD73, a MSC surface marker) that are potentially reorganized in the TME [69]. On the other hand, exosomes derived from tumor cells are able to transport tumor factors and appear to be involved in tumor invasion, angiogenesis, chemoresistance, immune evasion, and circumventing cell death [44]. By proteomic profiling analysis, it has been identified that Annexin II is one of the most highly expressed proteins in exosomes that causes macrophage-mediated activation of the p38MAPK, NF-κB, and STAT3 pathways and increased secretion of IL6 and TNFα [128]. TD-exosomes can provide autocrine, paracrine, endocrine, and other signals that promote cancer growth and maintain its invasion and metastasis [105]. In this context, since Annexin II plays an important role in angiogenesis and breast cancer metastasis, it can be exploited as a potential biomarker as well as a therapeutic target for diagnosis and treatment of metastatic breast cancer.
Metastatic Niches
Local and Systemic Dissemination
Dissemination of tumor cells is an essential step in metastasis. The TME is an important supporter of tumorigenesis in all forms of breast and prostate cancers and has been associated with the risk of metastasis and disease relapse. Resident CAFs and MSCs modulate early dissemination, tumorigenesis, and metastasis through alteration of ECM characteristics [29]. Further, several studies support the notion that tumor cell dissemination is predominantly affected by endothelial adhesion and related to functional changes of adhesion receptors. Therefore, the contribution of tumor-secreted factors to tumor-adhesion represents a therapeutic opportunity for preventing cancer metastasis. A recent study shows a network of paracrine signals between carcinoma, myeloid, and endothelial cells that drives both metastasis and chemoresistance in breast cancer [2]. In this study, cancer cells that overexpress CXCL1 and 2 are attracted by CD11b + Gr1+ myeloid cells into the tumor and enhance cancer cells survival by producing chemokines (S100A8/9). TNF is produced by endothelial or other stromal cells via NF-kB which amplifies the CXCL1/2 expression, thus strengthening the CXCL1/2- S100A8/9 loop, which causes chemoresistance in the TME. Another study demonstrated a novel mechanism to control tumor-cell dissemination. NEDD9 is a recognized marker of invasive and metastatic cancers that is required for matrix metalloproteinase 14 (MMP14). MMP14 secretion recycles through late endosomes and allows the release of tissue inhibitors of matrix metalloproteinase 2 (TIMP2) for tumor invasion. This finding illustrates a novel mechanism to control tumor-cell dissemination through NEDD9/Arf6-dependent regulation of MMP14/TIMP2 trafficking [57]. Several studies support that other factors such as BRD4, IL-30, serpin –E2, VEGF-A and Tenascin-C regulate breast cancer dissemination and metastatic colonization [3, 4, 71, 88] in the TME. Using real-time imaging, it was observed that for tumor cell dissemination, vascular permeability is transient, restricted only to TME of metastasis [40]. VEGF-A signaling from Tie2Hi TMEM macrophages causes local loss of vascular junctions, transient vascular permeability, and tumor cell intravasation, which explains a role of TME within the primary mammary tumor.
Composition
Metastasis is the systemic dissemination of tumor cells that profoundly depends on its microenvironment or “niche” interactions with resident components. It is a multistep process whether primary tumors detach and invade into the blood vessels or lymphatic system and finally develop as secondary tumors [78]. Emerging research is illuminating the underlying cellular and molecular events controlling the metastatic cascade from onset to colonization. Now, it is defined that a reciprocal interplay between tumor cells, stromal cells, and the ECM mediates metastasis in epithelial cancers [47]. The essential components of metastatic niches include ECM proteins, secreted enzymes, growth factors, cytokines, and other molecules that carry information to cancer cells. The sources of this molecular milieu are multiple cell types including cancer cells themselves, which are local or recruited to the site of metastasis [26]. Further, the stromal compartment contains various bone marrow-derived cells (BMDCs) such as macrophages, mast cells, neutrophils, and lymphocytes recruited by the primary tumor cells to increase tumor cell migration, angiogenesis, and invasion. Observation of immune infiltrates at sites of focal disruptions of the myoepithelial cell layer in breast tissues suggests a potential role of these cells in tumor malignancy and metastatic spread [47, 52]. Collective dissemination is a frequent mechanism for metastatic spread and whether the most invasive, keratin 14+ (K14+) expression in breast cancer cells regulate cell–cell adhesion, cell–matrix adhesion, and immune evasion [21, 22]. Understanding the molecular basis of collective dissemination will provide novel prognostics and therapies to improve patient outcomes.
Research has demonstrated that metastatic niches are shaped by the interactions of stroma and tumor and is mediated by monocarboxylate metabolism [73]. This study suggests that stromal carcinoma-associated fibroblasts (CAFs), generated from bone marrow-derived MSCs, may (i) recycle tumor-derived lactate for their own energetic requirements and thus sparing glucose for neighboring glycolytic tumor cells, and also (ii) discharge extra valuable metabolites of lactate oxidation, such as pyruvate for supporting tumor growth. Breast cancer metastasizes to the lung, bone, liver, and brain where an emerging paradigm suggests that primary tumor cells may secrete factors capable of inducing a productive microenvironment, termed the “pre-metastatic niche.” Further, pre-metastatic niches favor the seeding and proliferation of metastatic cells at unique sites. A study has shown that breast cancer-secreted miR-122 reprograms glucose metabolism in the pre-metastatic niche to promote metastasis. High miR-122 levels in the circulation have been associated with metastasis in breast cancer patients. This study demonstrated that by modifying glucose utilization by recipient pre-metastatic niche cells, cancer-derived extracellular miR-122 can reprogram energy metabolism and facilitate disease progression [32]. Another study has revealed the role of TAMs within the stroma of the pre-metastatic niche. It was found that human breast cancer specimens exhibited significant association between mRNA expression levels of MIP-1β and MYO3A and both, whether MIP-1β and MYO3A showed positive correlation with an established molecular determinant of cancer cell invasion, MMP9. Higher expression of these genes correlated with poor survival of breast cancer patients [5].
Recently, O’Connell and colleagues found that S100A4-positive fibroblasts provide the suitable metastatic niche to support metastatic colonization. Fibroblasts can promote angiogenesis and protect against apoptosis, respectively, through production of VEGF-A and tenascin-C. This study of the metastatic microenvironment of breast cancer in respect of clinical implication revealed that postmenopausal women treated with adjuvant bisphosphonates showed an 18% improvement in DFS (hazard ratio [HR] = 0.82; 95% CI 0.74–0.92, 2P = <0.001) by meta-analysis data. Reductions in relapse rates not only in bone but also at extra-skeletal and locol-regional sites indicate that bone-targeted treatments may modify the course of the disease through both direct and indirect inhibitory effects on niche components and signallings between tumor and bone cells [25].
Plasticity
Both tumor microenvironmental signals and cell-intrinsic modifications contribute to epigenetic changes that can induce cancer cells to adapt under challenging conditions. Successful invasion of the stroma, allow them to survive in lymphatic or blood vessels, and colonization of distant/different organs, as well as become resistant to cytotoxic drugs. Common adaptive responses include enhanced plasticity in cell motility, resistance to apoptosis, and survival in a hostile environment characterized by hypoxia. Moreover, several components of the TME including tumor mesenchyme such as fibroblasts, MSCs, CAFs, and macrophages are directly or indirectly linked with oxidative stress (OS), and serve as drivers of adaptive strategies [91].
To understand plasticity and the metastatic niche, the interplay between cancer cells and the microenvironment of intact xenograft tumors at distinct stages of metastasis is determined using a dual species-specific microarray platform. The study focused on three organ sites (brain, bone, and lung) to which breast cancer commonly metastasizes. It was found stage-specific changes happen in tumor cell-derived proteases and their inhibitors in expression during metastasis. However, stromal-derived genes are primarily regulated in a tissue-specific manner. It was found that cathepsin S expression enhances breast-to-brain metastasis and is regulated in both a cell type-specific and stage-dependent manner [82]. Another study showed that microenvironmental factors and recurrence rates are not strictly related to cell intrinsic properties. Outgrowth of the same population of incipient tumors is accelerated in mice with triple-negative breast cancer (TNBC) relative to those with luminal breast cancer (LBC). A tumor-supportive microenvironment enriched for EGF and IGF-1 is created by systemic signals provided by overt TNBCs at distant indolent tumor sites. Further, EGF and IGF-1 enhances expression of transcription factors associated with pluripotency, proliferation, and EMT [18]. Macrophage phenotypic subtypes completely regulate epithelial-to-mesenchymal plasticity in breast cancer cells [103]. Co-culturing of epithelial and mesenchymal cell lines with macrophages revealed that M2 macrophages might impart outgrowth and M1 macrophages may contribute to dormancy behaviors in metastatic breast cancer cells. A study has reported that embryonic stem cells precondition microenvironments through signal transducer activator of transcription 3 (Stat3) and suppress tumorigenic properties in breast cancer [41]. This study may act as a platform to better characterize tumor cell plasticity within the metastatic niche, providing new therapeutic strategies to inhibit breast and/or prostate cancer progression.
Concluding Remarks and Prospects
The complexity of the tumor microenvironment and its evolution throughout the course of breast and prostate cancer progression presents significant challenges to how basic scientists and translational clinicians collaborate to best identify effective strategies for curing patients. As such, it is essential that future work carefully study and characterize the complex cellular states that exist within the primary and metastatic tumor microenvironments of these malignancies, and more specifically, how these cellular states change in response to surgical or pharmaceutical intervention. Understanding these changing cellular states will likely unveil new tumor and microenvironment cell vulnerabilities that can further be exploited to adjust treatment strategies—even possibly in real time.
Importantly, large data efforts have begun to make progress along this avenue, but more work is needed in this area. The Human Protein Atlas project is an online repository providing large quantities of subcellular immunofluorescence and clinical immunohistochemistry (IHC) imaging data from normal and malignant cells and patient tissues [94,95,96]. This resource provides a wealth of information for oncology researchers. Figures 1 and 2 represent IHC images from breast and prostate malignancies that were extracted from the Human Protein Atlas and show markers of both tumor and stromal cell compartments in these malignancies. While helpful to query the relevance of specific markers in various cancer types—no doubt instructing follow-up experiments, these data do not provide co-labeling of these various markers across multiple patient samples or in heterogeneous co-cultures. Furthermore, this resource does not enable access to patient samples pre-/post-resection or therapeutic intervention.

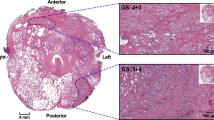
Immunihistochemistry of cell state markers indicating prostate cancer heterogeneity. Patient samples were retrieved from the Human Protein Atlas

Immunihistochemistry of cell state markers indicating breast cancer heterogeneity. Patient samples were retrieved from the Human Protein Atlas
Examples of new methodologies that are opening distinct avenues for the study tumor cell state evolution within the complex microenvironment in response to treatment regimens are the cyclic immunofluorescence (CycIF) [54, 55] and tissue-based CycIF (t-CycIF) [56] techniques. These open-source platforms enable high-resolution, multi-plexed, subcellular immunofluorescence analysis of cells in culture (CycIF) and in frozen or fixed tissues (t-CycIF). Thus, researchers can now profile over 30 unique antigens simultaneously and obtain single-cell information about the spatial heterogeneity of tumor and stroma cells within their native or more defined microenvironments. Achieving such resolution will further inform research efforts aimed at identifying vulnerabilities for better treatment strategies, ultimately leading to better breast and prostate cancer patient outcomes.
References
Acerbi I, Cassereau L, Dean I, Shi Q, Au A, Park C, et al. Human breast cancer invasion and aggression correlates with ECM stiffening and immune cell infiltration. Integr Biol (Camb). 2015;7:1120–34.
Acharyya S, Oskarsson T, Vanharanta S, Malladi S, Kim J, Morris PG, et al. A CXCL1 paracrine network links cancer chemoresistance and metastasis. Cell. 2012;150:165–78.
Airoldi I, Cocco C, Sorrentino C, Angelucci D, Di Meo S, Manzoli L, et al. Interleukin-30 promotes breast cancer growth and progression. Cancer Res. 2016;76:6218–29.
Andrieu G, Tran AH, Strissel KJ, Denis GV. BRD4 regulates breast cancer dissemination through Jagged1/Notch1 signaling. Cancer Res. 2016;76:6555–67.
Baghel KS, Tewari BN, Shrivastava R, Malik SA, Lone MU, Jain NK, et al. Macrophages promote matrix protrusive and invasive function of breast cancer cells via MIP-1beta dependent upregulation of MYO3A gene in breast cancer cells. Oncoimmunology. 2016;5:e1196299.
Baker EL, Bonnecaze RT, Zaman MH. Extracellular matrix stiffness and architecture govern intracellular rheology in cancer. Biophys J. 2009;97:1013–21.
Barcus CE, O’Leary KA, Brockman JL, Rugowski DE, Liu Y, Garcia N, et al. Elevated collagen-I augments tumor progressive signals, intravasation and metastasis of prolactin-induced estrogen receptor alpha positive mammary tumor cells. Breast Cancer Res. 2017;19:9.
Barron DA, Rowley DR. The reactive stroma microenvironment and prostate cancer progression. Endocr Relat Cancer. 2012;19:R187–204.
Beck JN, Singh A, Rothenberg AR, Elisseeff JH, Ewald AJ. The independent roles of mechanical, structural and adhesion characteristics of 3D hydrogels on the regulation of cancer invasion and dissemination. Biomaterials. 2013;34:9486–95.
Benyahia Z, Dussault N, Cayol M, Sigaud R, Berenguer-Daize C, Delfino C, et al. Stromal fibroblasts present in breast carcinomas promote tumor growth and angiogenesis through adrenomedullin secretion. Oncotarget. 2017;8:15744–62.
Bertos NR, Park M. Breast cancer - one term, many entities? J Clin Invest. 2011;121:3789–96.
Bliss SA, Sinha G, Sandiford OA, Williams LM, Engelberth DJ, Guiro K, et al. Mesenchymal stem cell–derived exosomes stimulate cycling quiescence and early breast cancer dormancy in bone marrow. Cancer Res. 2016;76:5832–44.
Bobrie A, Krumeich S, Reyal F, Recchi C, Moita LF, Seabra MC, et al. Rab27a supports exosome-dependent and -independent mechanisms that modify the tumor microenvironment and can promote tumor progression. Cancer Re. 2012;72(19):4920–30.
Bott A, Erdem N, Lerrer S, Hotz-Wagenblatt A, Breunig C, Abnaof K, et al. miRNA-1246 induces pro-inflammatory responses in mesenchymal stem/stromal cells by regulating PKA and PP2A. Oncotarget. 2017;8(27):43897–43914.
Calvo F, Sahai E. Cell communication networks in cancer invasion. Curr Opin Cell Biol. 2011;23:621–9.
Calvo F, Ege N, Grande-Garcia A, Hooper S, Jenkins RP, Chaudhry SI, et al. Mechanotransduction and YAP-dependent matrix remodelling is required for the generation and maintenance of cancer-associated fibroblasts. Nat Cell Biol. 2013;15:637–46.
Castaño Z, Fillmore CM, Kim CF, McAllister SS. The bed and the bugs: interactions between the tumor microenvironment and cancer stem cells. Semin Cancer Biol. 2012;22:462–70.
Castano Z, Marsh T, Tadipatri R, Kuznetsov HS, Al-Shahrour F, Paktinat M, et al. Stromal EGF and igf-I together modulate plasticity of disseminated triple-negative breast tumors. Cancer Discov. 2013;3:922–35.
Chaffer CL, Marjanovic ND, Lee T, Bell G, Kleer CG, Reinhardt F, et al. Poised chromatin at the ZEB1 promoter enables breast cancer cell plasticity and enhances Tumorigenicity. Cell. 2013;154:61–74.
Chang Y, Zuka M, Perez-Pinera P, Astudillo A, Mortimer J, Berenson JR, et al. Secretion of pleiotrophin stimulates breast cancer progression through remodeling of the tumor microenvironment. Proc Natl Acad Sci. 2007;104:10888–93.
Cheung KJ, Gabrielson E, Werb Z, Ewald AJ. Collective invasion in breast cancer requires a conserved basal epithelial program. Cell. 2013;155:1639–51.
Cheung KJ, Padmanaban V, Silvestri V, Schipper K, Cohen JD, Fairchild AN, et al. Polyclonal breast cancer metastases arise from collective dissemination of keratin 14-expressing tumor cell clusters. Proc Natl Acad Sci U S A. 2016;113:E854–63.
Cho JA, Park H, Lim EH, Lee KW. Exosomes from breast cancer cells can convert adipose tissue-derived mesenchymal stem cells into myofibroblast-like cells. Int J Oncol. 2012;40:130–8.
Chung LW, Baseman A, Assikis V, Zhau HE. Molecular insights into prostate cancer progression: the missing link of tumor microenvironment. J Urol. 2005;173:10–20.
Coleman RE, Gregory W, Marshall H, Wilson C, Holen I. The metastatic microenvironment of breast cancer: clinical implications. Breast. 2013;22(Suppl 2):S50–6.
Descot A, Oskarsson T. The molecular composition of the metastatic niche. Exp Cell Res. 2013;319:1679–86.
Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–7.
Donnarumma E, Fiore D, Nappa M, Roscigno G, Adamo A, Iaboni M, et al. Cancer-associated fibroblasts release exosomal microRNAs that dictate an aggressive phenotype in breast cancer. Oncotarget. 2017;8:19592–608.
Dumont N, Liu B, Defilippis RA, Chang H, Rabban JT, Karnezis AN, et al. Breast fibroblasts modulate early dissemination, tumorigenesis, and metastasis through alteration of extracellular matrix characteristics. Neoplasia. 2013;15:249–62.
Elkabets M, Gifford AM, Scheel C, Nilsson B, Reinhardt F, Bray MA, et al. Human tumors instigate granulin-expressing hematopoietic cells that promote malignancy by activating stromal fibroblasts in mice. J Clin Invest. 2011;121:784–99.
Erez N, Truitt M, Olson P, Arron ST, Hanahan D. Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-kappaB-dependent manner. Cancer Cell. 2010;17:135–47.
Fong MY, Zhou W, Liu L, Alontaga AY, Chandra M, Ashby J, et al. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat Cell Biol. 2015;17:183–94.
Friedl P, Alexander S. Cancer invasion and the microenvironment: plasticity and reciprocity. Cell. 2011;147:992–1009.
Ganguly SS, Li X, Miranti CK. The host microenvironment influences prostate cancer invasion, systemic spread, bone colonization, and osteoblastic metastasis. Front Oncol. 2014;4:364.
Ghesquiere B, Wong BW, Kuchnio A, Carmeliet P. Metabolism of stromal and immune cells in health and disease. Nature. 2014;511:167–76.
Gonzalez ME, Martin EE, Anwar T, Arellano-Garcia C, Medhora N, Lama A, et al. Mesenchymal stem cell-induced DDR2 mediates stromal-breast cancer interactions and metastasis growth. Cell Rep. 2017;18:1215–28.
Gritsenko PG, Ilina O, Friedl P. Interstitial guidance of cancer invasion. J Pathol. 2012;226:185–99.
Hanahan D, Coussens LM. Accessories to the crime: functions of cells recruited to the tumor microenvironment. Cancer Cell. 2012;21:309–22.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74.
Harney AS, Arwert EN, Entenberg D, Wang Y, Guo P, Qian BZ, et al. Real-time imaging reveals local, transient vascular permeability, and tumor cell Intravasation stimulated by TIE2hi macrophage-derived VEGFA. Cancer Discov. 2015;5:932–43.
He N, Feng G, Li Y, Xu Y, Xie X, Wang H, et al. Embryonic stem cell preconditioned microenvironment suppresses tumorigenic properties in breast cancer. Stem Cell Res Ther. 2016;7:95.
Iijima J, Konno K, Itano N. Inflammatory alterations of the extracellular matrix in the tumor microenvironment. Cancers. 2011;3:3189–205.
Insua-Rodríguez J, Oskarsson T. The extracellular matrix in breast cancer. Adv Drug Deliv Rev. 2016;97:41–55.
Kahlert C, Kalluri R. Exosomes in tumor microenvironment influence cancer progression and metastasis. Journal of molecular medicine (Berlin, Germany). 2013;91:431–7.
Kai F, Laklai H, Weaver VM. Force matters: biomechanical regulation of cell invasion and migration in disease. Trends Cell Biol. 2016;26:486–97.
Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016;16:582–98.
Khamis ZI, Sahab ZJ, Sang QX. Active roles of tumor stroma in breast cancer metastasis. Int J Breast Cancer. 2012;2012:574025.
King HW, Michael MZ, Gleadle JM. Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer. 2012;12:421.
Ko P, Kim D, You E, Jung J, Oh S, Kim J, et al. Extracellular matrix rigidity-dependent sphingosine-1-phosphate secretion regulates metastatic cancer cell invasion and adhesion. Sci Rep. 2016;6:21564.
Kong W, Philipps AF, Dvorak B, Anderson GG, Lake M, Koldovsky O. Presence of insulin-like growth factor I but absence of the binding proteins in the bile of rats. Am J Phys. 1995;268:R266–71.
Kreger BT, Johansen ER, Cerione RA, Antonyak MA. The enrichment of survivin in exosomes from breast cancer cells treated with paclitaxel promotes cell survival and chemoresistance. Cancers (Basel). 2016;8(12). https://doi.org/10.3390/cancers812011.
Kuznetsov HS, Marsh T, Markens BA, Castano Z, Greene-Colozzi A, Hay SA, et al. Identification of luminal breast cancers that establish a tumor-supportive macroenvironment defined by proangiogenic platelets and bone marrow-derived cells. Cancer Discov. 2012;2:1150–65.
Li S, Lu J, Chen Y, Xiong N, Li L, Zhang J, et al. MCP-1-induced ERK/GSK-3beta/Snail signaling facilitates the epithelial-mesenchymal transition and promotes the migration of MCF-7 human breast carcinoma cells. Cell Mol Immunol. 2016;
Lin JR, Fallahi-Sichani M, Sorger PK. Highly multiplexed imaging of single cells using a high-throughput cyclic immunofluorescence method. Nat Commun. 2015;6:8390.
Lin JR, Fallahi-Sichani M, Chen JY, Sorger PK. Cyclic immunofluorescence (CycIF), a highly multiplexed method for single-cell imaging. Curr Protoc Chem Biol. 2016;8:251–64.
Lin J-R, Izar B, Mei S, Wang S, Shah P, Sorger P. A simple open-source method for highly multiplexed imaging of single cells in tissues and tumours. bioRxiv. 2017. https://doi.org/10.1101/151738.
Loskutov YV, Kozyulina PY, Kozyreva VK, Ice RJ, Jones BC, Roston TJ, et al. NEDD9/Arf6-dependent endocytic trafficking of matrix metalloproteinase 14: a novel mechanism for blocking mesenchymal cell invasion and metastasis of breast cancer. Oncogene. 2015;34:3662–75.
Lu H, Clauser KR, Tam WL, Frose J, Ye X, Eaton EN, et al. A breast cancer stem cell niche supported by juxtacrine signalling from monocytes and macrophages. Nat Cell Biol. 2014a;16:1105–17.
Lu H, Clauser KR, Tam WL, Fröse J, Ye X, Eaton EN, et al. A breast cancer stem cell niche supported by Juxtacrine signaling from monocytes and macrophages. Nat Cell Biol. 2014b;16:1105–17.
Luga V, Zhang L, Viloria-Petit AM, Ogunjimi AA, Inanlou MR, Chiu E, et al. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell. 2012;151:1542–56.
Luo YP, Zhou H, Krueger J, Kaplan C, Liao D, Markowitz D, et al. The role of proto-oncogene Fra-1 in remodeling the tumor microenvironment in support of breast tumor cell invasion and progression. Oncogene. 2009;29:662–73.
Luo M, Hou L, Li J, Shao S, Huang S, Meng D, et al. VEGF/NRP-1axis promotes progression of breast cancer via enhancement of epithelial-mesenchymal transition and activation of NF-kappaB and beta-catenin. Cancer Lett. 2016;373:1–11.
Mao Y, Keller ET, Garfield DH, Shen K, Wang J. Stroma cells in tumor microenvironment and breast cancer. Cancer Metastasis Rev. 2013;32:303–15.
Mao L, Li J, Chen WX, Cai YQ, Yu DD, Zhong SL, et al. Exosomes decrease sensitivity of breast cancer cells to adriamycin by delivering microRNAs. Tumour Biol. 2016;37:5247–56.
Marsh T, Pietras K, McAllister SS. Fibroblasts as architects of cancer pathogenesis. Biochim Biophys Acta (BBA) - Mol Basis Dis. 2013;1832:1070–8.
McAllister SS, Weinberg RA. The tumour-induced systemic environment as a critical regulator of cancer progression and metastasis. Nat Cell Biol. 2014;16:717–27.
McAndrews KM, McGrail DJ, Ravikumar N, Dawson MR. Mesenchymal stem cells induce directional migration of invasive breast cancer cells through TGF-beta. Sci Rep. 2015;5:16941.
Melo SA, Sugimoto H, O'Connell JT, Kato N, Villanueva A, Vidal A, et al. Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell. 2014;26:707–21.
Melzer C, von der Ohe J, Lehnert H, Ungefroren H, Hass R. Cancer stem cell niche models and contribution by mesenchymal stroma/stem cells. Mol Cancer. 2017;16:28.
Nguyen-Ngoc KV, Cheung KJ, Brenot A, Shamir ER, Gray RS, Hines WC, et al. ECM microenvironment regulates collective migration and local dissemination in normal and malignant mammary epithelium. Proc Natl Acad Sci U S A. 2012;109:E2595–604.
O'Connell JT, Sugimoto H, Cooke VG, MacDonald BA, Mehta AI, LeBleu VS, et al. VEGF-A and Tenascin-C produced by S100A4+ stromal cells are important for metastatic colonization. Proc Natl Acad Sci U S A. 2011;108:16002–7.
Ono M, Kosaka N, Tominaga N, Yoshioka Y, Takeshita F, Takahashi R-U, et al. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci Signal. 2014;7:ra63.
Patel BB, Ackerstaff E, Serganova IS, Kerrigan JE, Blasberg RG, Koutcher JA, et al. Tumor stroma interaction is mediated by monocarboxylate metabolism. Exp Cell Res. 2017;352:20–33.
Pickup MW, Mouw JK, Weaver VM. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 2014;15:1243–53.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–7.
Place AE, Jin Huh S, Polyak K. The microenvironment in breast cancer progression: biology and implications for treatment. Breast Cancer Research : BCR. 2011;13:227.
Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013;19:1423–37.
Rahman M, Mohammed S. Breast cancer metastasis and the lymphatic system. Oncol Lett. 2015;10:1233–9.
Santoni M, Piva F, Scarpelli M, Cheng L, Lopez-Beltran A, Massari F, et al. The origin of prostate metastases: emerging insights. Cancer Metastasis Rev. 2015;34:765–73.
Santos JC, Ribeiro ML, Sarian LO, Ortega MM, Derchain SF. Exosomes-mediate microRNAs transfer in breast cancer chemoresistance regulation. Am J Cancer Res. 2016;6:2129–39.
Scheel C, Eaton EN, Li SH, Chaffer CL, Reinhardt F, Kah KJ, et al. Paracrine and autocrine signals induce and maintain mesenchymal and stem cell states in the breast. Cell. 2011;145:926–40.
Sevenich L, Bowman RL, Mason SD, Quail DF, Rapaport F, Elie BT, et al. Analysis of tumour- and stroma-supplied proteolytic networks reveals a brain-metastasis-promoting role for cathepsin S. Nat Cell Biol. 2014;16:876–88.
Sharon Y, Alon L, Glanz S, Servais C, Erez N. Isolation of normal and cancer-associated fibroblasts from fresh tissues by fluorescence activated cell sorting (FACS). J Vis Exp. 2013;14(71);e4425.
Shiao SL, Chu GC, Chung LW. Regulation of prostate cancer progression by the tumor microenvironment. Cancer Lett. 2016;380:340–8.
Shree T, Olson OC, Elie BT, Kester JC, Garfall AL, Simpson K, et al. Macrophages and cathepsin proteases blunt chemotherapeutic response in breast cancer. Genes Dev. 2011;25:2465–79.
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66:7–30.
Sieh S, Taubenberger AV, Rizzi SC, Sadowski M, Lehman ML, Rockstroh A, et al. Phenotypic characterization of prostate cancer LNCaP cells cultured within a bioengineered microenvironment. PLoS One. 2012;7:e40217.
Smirnova T, Bonapace L, MacDonald G, Kondo S, Wyckoff J, Ebersbach H, et al. Serpin E2 promotes breast cancer metastasis by remodeling the tumor matrix and polarizing tumor associated macrophages. Oncotarget. 2016;7:82289–304.
Soysal SD, Tzankov A, Muenst SE. Role of the tumor microenvironment in breast cancer. Pathobiology. 2015;82:142–52.
Spill F, Reynolds DS, Kamm RD, Zaman MH. Impact of the physical microenvironment on tumor progression and metastasis. Curr Opin Biotechnol. 2016;40:41–8.
Taddei ML, Giannoni E, Comito G, Chiarugi P. Microenvironment and tumor cell plasticity: an easy way out. Cancer Lett. 2013;341:80–96.
Tan W, Zhang W, Strasner A, Grivennikov S, Cheng JQ, Hoffman RM, et al. Fibroblast-recruited, tumor-infiltrating CD4(+) T cells stimulate mammary cancer metastasis through RANKL-RANK signaling. Nature. 2011;470:548–53.
Tripathi M, Billet S, Bhowmick NA. Understanding the role of stromal fibroblasts in cancer progression. Cell Adhes Migr. 2012;6:231–5.
Uhlen M, Bjorling E, Agaton C, Szigyarto CA, Amini B, Andersen E, et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol Cell Proteomics. 2005;4:1920–32.
Uhlen M, Oksvold P, Fagerberg L, Lundberg E, Jonasson K, Forsberg M, et al. Towards a knowledge-based human protein atlas. Nat Biotechnol. 2010;28:1248–50.
Uhlen M, Fagerberg L, Hallstrom BM, Lindskog C, Oksvold P, Mardinoglu A, et al. Proteomics. Tissue-based map of the human proteome. Science. 2015;347:1260419.
Wei SC, Fattet L, Tsai JH, Guo Y, Pai VH, Majeski HE, et al. Matrix stiffness drives epithelial-mesenchymal transition and tumour metastasis through a TWIST1-G3BP2 mechanotransduction pathway. Nat Cell Biol. 2015;17:678–88.
Wen SW, Sceneay J, Lima LG, Wong CS, Becker M, Krumeich S, et al. The biodistribution and immune suppressive effects of breast cancer-derived exosomes. Cancer Res. 2016;76:6816–27.
Wolf K, Friedl P. Extracellular matrix determinants of proteolytic and non-proteolytic cell migration. Trends Cell Biol. 2011;21:736–44.
Wolfe AR, Trenton NJ, Debeb BG, Larson R, Ruffell B, Chu K, et al. Mesenchymal stem cells and macrophages interact through IL-6 to promote inflammatory breast cancer in pre-clinical models. Oncotarget. 2016;7:82482–92.
Xiong G-F, Xu R. Function of cancer cell-derived extracellular matrix in tumor progression. Journal of Cancer Metastasis and Treatment. 2016;2:357–64.
Yang M, Chen J, Su F, Yu B, Su F, Lin L, et al. Microvesicles secreted by macrophages shuttle invasion-potentiating microRNAs into breast cancer cells. Mol Cancer. 2011;10:117.
Yang M, Ma B, Shao H, Clark AM, Wells A. Macrophage phenotypic subtypes diametrically regulate epithelial-mesenchymal plasticity in breast cancer cells. BMC Cancer. 2016;16:419.
Yu W, Chai H, Li Y, Zhao H, Xie X, Zheng H, et al. Increased expression of CYP4Z1 promotes tumor angiogenesis and growth in human breast cancer. Toxicol Appl Pharmacol. 2012;264:73–83.
Yu S, Cao H, Shen B, Feng J. Tumor-derived exosomes in cancer progression and treatment failure. Oncotarget. 2015;6:37151–68.
Zaman MH, Trapani LM, Sieminski AL, Mackellar D, Gong H, Kamm RD, et al. Migration of tumor cells in 3D matrices is governed by matrix stiffness along with cell-matrix adhesion and proteolysis. Proc Natl Acad Sci U S A. 2006;103:10889–94.
Zhang W, Grivennikov SI. Top notch cancer stem cells by paracrine NF-kappaB signaling in breast cancer. Breast Cancer Res. 2013;15:316.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
F. Runa, S. Hamalian, K. Meade, P. Shisgal, P.C. Gray, and J.A. Kelber declare that they have no conflict of interest.
Human and Animal Rights and Informed Consent
This article contains no studies with human or animal subjects performed by any of the authors.
Additional information
This article is part of the Topical Collection on Molecular Biology of Prostate Cancer
Rights and permissions
About this article

Cite this article
Runa, F., Hamalian, S., Meade, K. et al. Tumor Microenvironment Heterogeneity: Challenges and Opportunities. Curr Mol Bio Rep 3, 218–229 (2017). https://doi.org/10.1007/s40610-017-0073-7
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40610-017-0073-7