
Abstract
The vereda (Brazilian savanna palm swamp) has a distinguishable flora and high floristic beta diversity. The factors that influence dissimilarities among vereda floras have not been clearly elucidated. Here we investigated how key physicochemical soil factors modulate the species composition of this vegetation. Soil and plants were sampled from three veredas, species composition and diversity were compared among them. To investigate the effects of soil physicochemical properties on species richness and cover of each vereda, we fitted a linear mixed-effect model. The plant assemblages in the veredas diverged in terms of species composition but converged in two dominant species graminoids. We found significant differences in soil properties among the veredas, except in texture. Soil properties had significant effects on species richness and cover as a whole, and on richness and relative cover for the individual growth forms. Potential cation-exchange capacity alone negatively affected species richness, especially of graminoids and subshrubs. The cation-exchange capacity of soils and its complex interactions with phosphorus and organic matter were found to be the main drivers of species occurrence and vegetation cover in the veredas. The distinct flora of veredas is controlled by the interaction of species and its growth forms with soil filters.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Wetlands are considered ecosystems of global importance because they harbor a variety of endemic species and promote essential ecosystem services (Keddy 2010). However, they are one of the most sensitive ecosystems to degradation and are increasingly threatened (Junk et al. 2014). In this sense, understanding the environmental factors that control wetlands has become relevant. Soil plays a fundamental role for vegetation, serving as a source of nutrients for plants, and the hydrological condition in the environment strongly influences its characteristics (Weil and Brady 2016). Due to different hydrological conditions, wetlands exhibit high variability of soil physicochemical properties at different spatial scales (Shi et al. 2010; Nkheloane et al. 2012). Previous studies have shown that soil properties, such as organic matter, phosphorus (Liu et al. 2018), and pH can affect positively the composition and species richness in wetlands (Xiaolong et al. 2014). Nevertheless, the complexity of plant–soil interactions in determining differences in species composition, between different Brazilian savanna wetlands sites, are still not clear yet (Bijos et al. 2017).
The Cerrado (Brazilian savanna) biome extends over a wide latitudinal amplitude of 20° and features a high degree of climatic, edaphic, and altitudinal variation. Differences in topography, soil drainage, soil physical and chemical properties, and fire history frequency, affect the heterogeneity of vegetation types in the Cerrado (Ribeiro and Walter 2008). Among the Cerrado vegetation types the vereda (palm swamps) occurs in shallow valleys or flat areas, following undefined drainage routes, as mosaic patches of wet-savanna vegetation. This characteristic vegetation consists of continuous herbaceous layer with stands of the buriti palm (Mauritia flexuosa L.f.) and sparse shrubs and trees emerging in the wettest zones.
The dry-vegetation types neighboring the veredas and/or the variations in topography create a moisture gradient in the soil, which provides a mosaic of microhabitats that form a vegetation complex with species having different ecological requirements (Oliveira et al. 2009). The vereda harbors a high proportion of distinct herbaceous species (Silva et al. 2018). In addition to the typical flora and internal heterogeneity, the floristic richness and diversity vary widely among different vereda sites (Bijos et al. 2017). It is supposed that the floristic differences between veredas increase with increasing distances between them, probably in response to the larger differences in climate and soil conditions (Bijos et al. 2017). Few studies have evaluated the effects of these factors on the differences in floristic richness and diversity among vereda sites (Guimarães et al. 2002).
The hydromorphic soils in veredas are acidic, poor in nutrients, and with high levels of aluminum saturation (Ramos et al. 2006). The soil has high carbon, clay, and organic matter contents, which vary according to the moisture gradient, being higher in the waterlogged patches where nutrient levels rise and the pH is acidic (Ramos et al. 2006; Sousa et al. 2015). Changes in soil drainage can cause changes in its physicochemical and textural properties (Sigua et al. 2006), promoting encroachment by woody species and colonization by invasive species (Barbosa da Silva et al. 2016), and can also reduce the heterogeneity of microhabitats in the environment (Bleich et al. 2014). Although many studies show that fire frequency influences soil properties and vegetation, most of the studies on the effects of fire focus on dry savannas in the Cerrado (e.g., Silva and Batalha 2008; Pilon et al. 2021) and few have examined wet savannas (Araújo et al. 2013).
In view of the close relationship between soil and savanna vegetation (Furley and Ratter 1988) and that small-scale differences in soil properties may contribute to the distribution of species in the Cerrado (Dantas and Batalha 2011), we investigated the soil and vegetation attributes in veredas to explain how key physicochemical soil factors modulate the species composition and diversity of this vegetation at regional scale. We addressed the hypothesis that changes in floristic differences are driven by soil properties. Within the same Arenosol (sandy soils) matrix, we expected to find differences in species composition and soil properties among the vereda sites, since the edaphic factor has been strongly associated with plant species distribution in the Cerrado (Bueno et al. 2018).
2 Materials and methods
Study area –
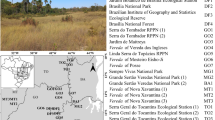
We collected floristic and soil data at the following three vereda sites, located in the northeastern Cerrado in Brazil: a vereda in Posse municipality, Goiás state (PO), an area of native cerrado vegetation belonging to the Goiano Federal Institute of Education, Science and Technology; a vereda in Chapada Gaúcha municipality, Minas Gerais state (GS1); and a vereda in Côcos municipality, Bahia state (GS2) (Fig. 1). The last two veredas are in the Grande Sertão Veredas National Park, one of the largest protected areas in the Cerrado, which covers more than 231,000 ha. The vereda PO is located next to the road, near an urban area. The regional climate is tropical savanna (Aw, according to the Köppen climate classification system), with a rainy summer from October to April and a dry winter from May to September. The annual mean maximum temperature at the three sites ranged from 30.2 °C to 31.5 °C, and the annual precipitation ranged from 1024 to 1314 mm (Supplementary Table S1). Quartzipsamment soils predominate in the region (Santos et al. 2018). This soil is poorly developed, deep, well drained, sandy (more than 90% sand), acidic, low fertility, with little differentiation between horizons, and with high aluminum saturation (Furley and Ratter 1988). Burned area maps for the veredas were produced with data from the Thematic Mapper (TM) sensor on board the Landsat-4-5 satellite, the Enhanced Thematic Mapper plus (ETM +) on board Landsat-7, and Landsat 8 (Operational Land Imager and the Thermal Infrared Sensor), for the months of April (end of the wet season) to November (early in the wet season), between 1999 and 2019. The Landsat images were downloaded from the ESPA—USGS (United States Geological Survey—http://espa.cr.usgs.gov/). The vereda PO had the highest frequency of burns, with nine fire scars in the last 21 years. In contrast, we recorded only one fire scar on GS1 and three on GS2 (Supplementary Table S2). Fire scars on the veredas occurred mainly in August and September, associated with burning in the adjacent savanna (Supplementary Fig. S1).

Locations of the sampling sites: vereda GS1—Grande Sertão Veredas National Park, municipality of Chapada Gaúcha in the state of Minas Gerais (MG); vereda GS2—Grande Sertão Veredas National Park, municipality of Côcos in the state of Bahia (BA); and vereda PO—municipality of Posse in the state of Goiás (GO)
Vegetation sampling –
Vegetation in the three areas was sampled in the rainy season between February and May in 2018 (GS1), 2019 (PO), and 2020 (GS2), when more species are likely to be in the reproductive period (Munhoz and Felfili 2006), facilitating identification. We determined the floristic composition and cover using the line intercept method (Canfield 1941, adapted by Munhoz et al. 2008). Three parallel transections were installed 80–100 m apart, extending throughout the entire humidity gradient of the vereda, from the waterlogged area at the lowest point of the vereda, where the buriti stands occur, to the border between the vereda and the cerrado sensu stricto. On each transect, five 20-m sections were sampled. The sample units were spaced 10 or 20 m apart along the transect, totaling 15 sample units in two of the veredas and 13 sample units in the smaller vereda (PO). In the sampling units, a millimeter measuring tape, placed 50 cm above the ground, was extended and the horizontal projection of each species that touched or came within 1 cm of the line, both below and above, was recorded. The sum of the horizontal projection of each species in the sample units corresponded to the value of its absolute cover (Munhoz et al. 2008). Cover percentage (relative cover) of each species was estimated by dividing the total length intercepted by the species (absolute cover) by the sum of the length of all species in the 15 or 13 sample units, multiplied by 100. Following the same principle, the percentage of relative frequency was determined, based on the number of sample units where each species occurred, instead of the cover (Munhoz et al. 2008). We estimated the importance value of the species by adding the values of relative cover and relative frequency.
The species collected were classified according to the main groups of plant growth forms (graminoids, herbs, subshrubs, shrubs, and trees), following Dansereau (1951). The relative cover of each growth form per transect was then obtained by summing the relative cover of all species of that growth form on every 13 or 15 20-m transect in the three veredas. The relative richness of each growth form by transect was calculated by dividing the number of species of each growth form present on the transect by the total number of species of all growth forms on the same transect, multiplied by 100. Species were identified by comparison with herbarium vouchers, specialized literature, and consultations with taxonomists. We updated all scientific names according to the List of Species of Flora do Brasil (Flora do Brasil 2020).
Soil sampling –
Soil samples were collected from a depth of 0–20 cm at a perpendicular distance of 1 m from the center (10 m) of each sample unit, totaling 43 soil samples. Soil texture was measured by the Bouyoucos densimeter method. The percentage of organic matter was measured using the Walkley–Black method (% organic C × 1.724). The soil pH was determined in 0.01 mol L–1 CaCl2 (pH CaCl2). The cations Ca2+ and Mg2+ were extracted with 1 mol L–1 KCl solution. Phosphorus and K+ were extracted with Mehlich-1 solution (0.0125 mol L–1 H2SO4 + 0.05 mol L–1 HCl). Potential acidity (H + Al) was evaluated by extracting with 0.5 mol L–1 calcium acetate solution at pH 7.0 and titrating with 0.025 mol L–1 NaOH. Potential cation exchange capacity (CEC) was estimated according to the equation \((\mathrm{T}= {\mathrm{Ca}}^{2+}+{\mathrm{Mg}}^{2+}+{\mathrm{K}}^{+}+{\mathrm{H}}^{+}+{\mathrm{Al}}^{3+})\) and soil cation saturation by \(\{\mathrm{BS}=[({\mathrm{Ca}}^{2+}+{\mathrm{Mg}}^{2+}+{\mathrm{K}}^{+}/\mathrm{CEC})]\times 100\}\). Aluminum saturation index was determined by \([\mathrm{m\%}=(\mathrm{Al}/\mathrm{CEC})\times 100]\). Detailed descriptions of methodological procedures of the laboratory soil analyses were given by Teixeira et al. (2017).
Data analysis –
The species diversity was compared among the veredas by diversity profiles based on Rényi’s entropy, displaying exponential series at 5% significance using the BiodiversityR package version 2.6-1 (Kindt and Coe 2005) in the R software, version 3.6.2 (R Core Team 2019). This analysis generated a visual representation that compared the values of several diversity metrics (e.g., number of species and Shannon and Simpson diversity indexes) simultaneously on the graph. The parameter α = 0 is highly sensitive to rare species, and as the scale increases, more weight is given to dominant species, following the Shannon diversity index (α = 1), Simpson diversity index (α = 2), and the Berger–Parker dominance index (α = ∞).
We used non-metric multidimensional scaling (metaMDS function; maximum number of iterations = 2000) to visualize the sites in the species composition space. We ran the NMDS twice: once with the species presence/absence matrix and once with the species cover matrix, using the Jaccard dissimilarity index and the Bray–Curtis index, respectively (Legendre and Legendre 2012). We also built ellipses with 95% confidence intervals around the centroids (stat_ellipse function) for each vereda site. To assess the similarity in species cover and presence/absence among the veredas, we performed a permutational multivariate analysis of variance (PERMANOVA) using the function adonis in the vegan package, with 999 permutations. We then performed a dispersion homogeneity test (PERMDISP) applying the betadisper function in vegan, which assesses the degree to which the significant PERMANOVA results are determined by differences in multivariate centroid location, relative dispersion, or both. We ran the analyses in the vegan package version 2.4-0 (Oksanen et al. 2015) of the R software, version 3.6.2 (R Core Team 2019).
To elucidate the differences in the mean values of relative richness and relative cover of growth forms among the three veredas, we ran a nested analysis of variance (nested ANOVA) (Zar 2010). Considering our nested experimental design, we created appropriate models to treat the effect of sites on the sample units, so we nested sample units into the vereda site. Transformation by log (x + 1.1) was applied to the relative percentage of the data to stabilize variances and to keep data normality. Data normality were tested previously to each analysis using a Shapiro–Wilk test. Model assumptions and homoscedasticity were verified when examining the model residuals. We used a mixed-effect model to fit a nested ANOVA, using vereda site as a fixed factor and the sample units as random effects. The effect of the sample units on growth form was tested by comparing this model to a model without the sample units. The model was fitted using the lme function of the nlme package version 3.1–149 (Pinheiro et al. 2019) in the R software, version 3.6.2 (R Core Team 2019), and the ANOVA was performed using the anova.lme function of the same package. To compare mean values and implicit statistical significance (P ≤ 0.05), we applied a Tukey test.
In order to compare the edaphic physicochemical properties among the three veredas, we used the Kruskal–Wallis test because the assumptions of normality of the residuals or the homogeneity of variances were not satisfied. The Nemenyi test, a non-parametric Tukey-type multiple comparison, was used to identify homogeneous subsets of means after applying the Kruskal–Wallis test (Zar 2010), using the R package DescTools version 0.99.34. A Mann–Whitney U test was applied to determine where differences existed among the soil physicochemical properties in the three veredas (Zar 2010).
To investigate the effects of soil on species richness and cover of the sample units of each vereda, we fitted a linear mixed-effect model (LME) to accommodate the spatial autocorrelation among the sample units and transects and to reduce unexplained residuals in our analysis. Relative cover, species richness (i.e. total species number per transect), and growth forms (relative cover and number of species by each growth form per transect, except for trees of their small numbers) were used as separate response variables in the models. The LME analysis was conducted using the lme function of the nlme package (Pinheiro et al. 2019) version 3.1–149 in R software, version 3.6.2 (R Core Team 2019). To avoid incorrect identification of important soil predictors due to high collinearity, we built a correlation matrix (Spearman correlation), discarding variables with coefficient |r|≥ 0.7 before fitting the model (Legendre and Legendre 2012). The fixed-effect predictor variables used in the analyses were: CEC, Phosphorus, pH, OM, and Clay (Supplementary Table S3), and all possible interactions of these variables. Veredas sites were used as a random factor in the models (Zuur et al. 2009), to take account for the different fire frequencies in the areas and the distance between the areas. We checked the residuals of the models for normal distribution, and when necessary to achieve normality of the distributed residuals, richness data were square-root transformed, and relative cover data were arcsine transformed (Zuur et al. 2009). The significance of the variables in the models were assessed by an Analysis of Deviance (ANODEV) with an F test. We used Akaike's Information Criterion (AIC) to obtain the minimum appropriate models based on the smallest AIC values (with both forward and backward search directions) using the stepAIC function from the MASS package (Venables and Ripley 2002).
3 Results
Vegetation cover, composition and diversity –
We recorded 142 species, 81 genera, and 36 families on the three vereda sites (Supplementary Table S4). Only 16 species were shared among all three vereda sites (Fig. 2). The most species-rich families were Poaceae (25 species), Cyperaceae (20), Eriocaulaceae (16), Xyridaceae (13), and Melastomataceae (12). Among the species that represent half of the total importance value (200%) in the three veredas, approximately 60% occurred in only one area, with four species exclusive to GS1, two to GS2, and two to PO (Fig. 3). The only species with the highest importance value in all three veredas were Trachypogon spicatus (L.f.) Kuntze and Paspalum lineare Trin. (Fig. 3). While Trachypogon spicatus (L.f.) Kuntze reached the highest importance value in veredas GS1 and PO, Otachyrium versicolor (Döll) Henrard had the highest importance value in GS2 (Fig. 3). Analysis of the diversity profiles showed that GS1 (e.g., α = 0, α = 1 and α = 2) and PO (e.g., α = ∞) had high species richness and diversity, while GS2 held lowest diversity (Fig. 4).

Shared and exclusive species sampled in the veredas in Grande Sertão Veredas National Park (GS1 and GS2) and in Posse (PO), central Brazil

Species that represent 50% of the total importance value (200%) in the veredas in Grande Sertão Veredas National Park (GS1 and GS2) and Posse (PO)

Diversity profiles for species in the veredas in Grande Sertão Veredas National Park (GS1 and GS2) and in Posse (PO). Parameter α = 0, diversity value equivalent to species richness; α = 1, diversity value equivalent to the Shannon index; α = 2, diversity value equivalent to Simpson’s inverse index (1/D); α = ∞ is the Berger-Parker index
The nMDS ordination analysis showed similar conformations in the multidimensional space for the presence/absence matrix and the cover matrix, with a clear separation of the vereda sites but with higher separation for presence/absence (Fig. 5). This was supported by the lack of overlap among the 95% confidence intervals for the three vereda centroids (Fig. 5). Besides the species cover, which differed significantly among the veredas (PERMANOVA: R2 = 0.478, P < 0.001), there was no significant difference in the dispersion of the groups (PERMDISP: F = 1.158, P < 0.34). The lack of significance generated by PERMDISP showed that the abundance of species varied similarly, although the veredas had distinct floristic compositions, as demonstrated in the PERMANOVA analysis. In contrast, the vereda sites differed significantly in species presence/absence composition (PERMANOVA: R2 = 0.315, P < 0.001; PERMDISP: F = 12.792, P < 0.001), with GS1 being the most heterogeneous (average distance to the median of the samples: 0.5519) and GS2 the least heterogeneous (average distance to the median: 0.4041). These results indicated that the three veredas had a high degree of floristic difference.

Cover (a) (stress = 0.14) and presence/absence (b) (stress = 0.17) NMDS ordinations for the species sampled on the 15 line transects in Grande Sertão Veredas National Park (GS1 and GS2) and 13 line transects in Posse (PO). Ellipses indicate 95% confidence intervals around the centroids of the vereda sites
We found a significant difference between herbs and subshrubs among the veredas, with the highest mean values of relative cover and richness for herbs in PO, followed by GS2. Vereda GS1 showed the highest mean values of relative cover and richness for subshrubs (Table 1). No significant differences were found in graminoid, shrub, and tree species richness and relative cover among the veredas (Table 1).
Soil properties and its influence on vegetation –
In general, the soils in the veredas were acidic, sandy, poor in nutrients, and with a high organic matter content. However, we found significant differences in soil properties among the veredas, except in texture (clay, silt, and sand contents) (Table 2). GS1 and PO showed higher soil cation saturation and phosphorus contents than GS2 (Table 2). On the other hand, GS2 showed the highest cation-exchange capacity and aluminium saturation (Table 2). Vereda PO had high organic matter and carbon contents, while GS1 had lowest contents of both (Table 2).
Soil properties had significant effects on species richness and cover as a whole and on richness and relative cover for the individual growth forms in the veredas (Table 3; Table S5). Potential cation-exchange capacity alone negatively affected species richness, especially of graminoids and subshrubs (Table 3). However, cation-exchange capacity in interaction with phosphorus positively affected the species richness and overall relative cover, especially for shrub richness, although the relative cover of graminoids was negatively affected. The interaction of organic matter, pH, and clay was positive for species richness and relative cover, especially for the relative cover of graminoids. For herbaceous species, the effect was the opposite. Organic matter alone positively affected the richness of herbaceous plants while reducing the richness of shrubs and subshrubs. The results indicated the complexity of the effects of soil properties on species richness and relative cover and on growth forms, by their varied effects on different interactions in each response variable measured (Table 3).
4 Discussion
We found that the plant assemblages in the three veredas diverged in terms of species composition but converged in two dominant species graminoids. We also found that the distinct flora of veredas is controlled by soil filters and the ability of species and growth forms to interact with them. The cation-exchange capacity of soils and its complex interactions with phosphorus, organic matter, and other soil properties were found to be the main drivers of species occurrence and vegetation cover in the veredas. The complex interactions described here show that we need to broaden our understanding of soil-vegetation relationships of savannas such as the veredas, to improve and direct conservation efforts.
Vegetation cover, composition and diversity –
Although geographically close veredas usually show floristic affinity (Silva et al. 2018), the floras of the veredas studied here, within the same Arenosol (sandy soils) matrix, differed in species composition and shared only two dominant species. This floristic distinction among veredas may be related to their isolation as islands of humid savannas within the dry savanna matrix. Therefore, each vereda site may represent a different floristic unit, as suggested by Cianciaruso and Batalha (2008) for wet grassland in the Cerrado. As well as in the veredas that we studied, the occurrence of distinct species, high richness, and high diversity is common in other vereda sites in central Brazil, as well as the high richness of the families Poaceae, Cyperaceae, Eriocaulaceae and Xyridaceae, (Bijos et al. 2017; Silva et al. 2018). Many wetland species have physiological and/or anatomical adaptations to lead with waterlogging, and consequently situations of hypoxia or even anoxia, such as the presence of aerenchyma cells (Keddy 2010).
More than half of the species with the highest importance values for each area were exclusive, showing that each vereda had its specific flora even though the plants belonged to the same vegetation type. Trachypogon spicatus (L.f.) Kuntze showed the highest importance values in all three areas. This species is dominant in humid grasslands in the Cerrado of northeastern Brazil (Mendes et al. 2014) and in Venezuelan savannas (Baruch et al. 2004) but is rare in moist grasslands, in cerrado sensu stricto, and (as far as known) in veredas in the Cerrado of central Brazil (Bijos et al. 2017; Souza et al. 2021). The second species with one of the highest importance values in the three areas, Paspalum lineare Trin., was recorded in all nine veredas in central Brazil studied by Bijos et al. (2017) and appeared to be associated with humid environments (Munhoz et al. 2008). The species Otachyrium versicolor, which was exclusive to GS2 and had the highest importance value in this area, also seems to be associated with wetter areas in veredas (Oliveira et al. 2009). Therefore, there is a convergent pattern of dominant species adapted to humid environments, but their occurrence at a specific site seems to be dependent on the isolation of the site. Site isolation may act as a filter for species from the same vegetational type at a given site. This pattern is most likely related to the degree of species diversity in the landscape, where higher species diversity would increase the number of unique communities locally (Ter Steege et al. 2013).
Graminoid species were predominant in terms of richness and cover. Members of the family Poaceae are abundant in the Cerrado biome, predominantly in moist grasslands and in dry and moist savannas (Munhoz et al. 2008; Bijos et al. 2017; Souza et al. 2021). This growth form tends to form dense clusters or has rhizomes or stolons that increase the plants ability to establish themselves throughout the vegetation (Munhoz and Felfili 2006; Pilon et al. 2021). Besides, the fire suppression in the Cerrado favors woody species over herbaceous species (Durigan 2020), as well as found in the African savanna wet grassland (Luvuno et al. 2016). However, our veredas (GS1 and GS2) with low fire frequency, showed low and high herbaceous diversity, respectively, and very low or no occurrence of tree in this vegetation type. Most likely, the degree and temporal extent of soil moisture in the veredas may act as an environmental filter that prevents woody species from invading, regardless of fire suppression (Xavier et al. 2019), besides contributing to a greater diversity of herbaceous-subshrub species in moist grasslands (Leite et al. 2018). Given the limitation of our study, we suggest that veredas be included in research on fire-exclusion policies in the Cerrado, because the effect of fire on these environments has not yet been clearly elucidated.
Soil properties and its influence on vegetation –
Overall, veredas show low cation saturation values and high levels of acidity and organic matter content. Similar conditions have been found in other veredas (e.g., Ramos et al. 2006). However, the physicochemical properties of the soil usually differ among veredas possibly explained by the variation in the seasonality of the water table, differences in sediment deposits among the sites, and/or human interference (Ramos et al. 2006). Also, different hydrological conditions in veredas influence the variation of soil properties (Ramos et al. 2006; Sousa et al. 2015). Taking into account that vereda GS2 showed a lower standard deviation in the mean values of soil properties, it is possible that the lower floristic heterogeneity in GS2 is related to the lower soil heterogeneity in this vereda, and that the higher floristic heterogeneity in GS1 and PO is related to a larger variety of microhabitats generated by differences in soil moisture. Habitat heterogeneity in wetlands are one of the main determinants of species richness, contributing to the beta diversity of these areas (Shi et al. 2010). The variation in soil moisture acts as a filter, selecting species more adapted to the environment; and the number of species tends to be higher in better-drained areas and lower in permanently waterlogging areas (Xavier et al. 2019; Souza et al. 2021). Waterlogged soils have higher aluminum saturation (Amorim and Batalha 2006), as found in GS2, presumably it is a homogeneously wet area and consequently with lower species richness than the other veredas.
The increase in the potential cation-exchange capacity was a major drive in reducing overall species richness. Nevertheless, this attribute in interaction with phosphorus was associated with an increase in species richness and the entire relative species cover. There is an intimate relationship between soil and plant that can affect nutrient uptake (Furley and Ratter 1988). The potential cation-exchange capacity favors the maintenance of soil fertility while retaining nutrients to release them to the plants gradually. However, the higher the percentage of aluminum saturation and the lower the percentage of cation saturation, the more cation-exchange capacity is occupied by exchangeable aluminum, and consequently, the poorer the soil becomes (Weil and Brady 2016). Aluminum, being trivalent, is more easily retained by the colloids, limiting the absorption capacity of nutrients such as calcium, magnesium, and potassium, which for this reason remain free in the soil solution and are lost by leaching (Weil and Brady 2016). Aluminum toxicity and phosphorus deficiency are among the main factors limiting agricultural production in the Cerrado, and native species are highly tolerant to these conditions (Haridasan 2008). Their adaptation to poor Cerrado soils does not mean that Cerrado plants do not respond positively to soils with higher fertility (Haridasan 2008). The interaction of phosphorus with aluminum continues to be investigated and many studies have shown that the presence of phosphorus reduces aluminum toxicity, for various reasons, such as increasing aluminum resistance as well as ensuring better root development and nutrient uptake (Chen et al. 2011). Our results agree with these findings since phosphorus interaction with cation-exchange capacity (occupied mainly by aluminum) showed a positive effect on species richness and relative cover.
The interaction among organic matter, pH, and clay also positively affected species richness and relative cover. We believe that together these soil attributes promote higher soil fertility in these environments. Organic matter is an important indicator of soil quality (Sousa et al. 2015) and has a strong influence on the cation-exchange capacity, assisting in improving the chemical quality of weathered soils (Ramos et al. 2018); while humic substances present in organic matter decrease the soil aluminum saturation through their strong affinity for aluminum (Haynes and Mokolobate 2001). The increase in pH indicates a reduction of H+ ions, another element considered toxic, as it erodes the structure of minerals, releasing aluminum that is easily retained by colloids (Weil and Brady 2016). Clay, in turn, is considered one of the main colloids responsible for cation-exchange capacity, due to its electrically charged surface (Weil and Brady 2016), which in environments with lower amounts of available aluminum enables greater retention of nutrients essential for plant development (Ellis and Mellor 1995).
Soil properties affect species richness and relative cover, contributing to the distinction of the herbaceous-shrub layer among the Cerrado vegetation types (Souza et al. 2021). Among growth forms, cation-exchange capacity negatively affected the subshrubs and graminoids species in the veredas. The cation-exchange capacity and phosphorus interaction positively affected shrubs, but graminoids were negatively affected. In part, this disagrees with the finding of Amorim and Batalha (2006) that higher phosphorus availability favors Poaceae and Cyperaceae in humid environments. In a wetland in Minnesota, USA, only graminoid species were limited by phosphorus (Chapin et al. 2004). However, it is not easy to relate these studies to each other, since the possible interactions with other soil properties in these environments was not considered. Very few studies have investigated these interactions, which makes it challenging to reach better-founded conclusions and avoid misinterpretations. We wonder if the higher fire frequency in vereda PO might be influencing the higher amount of phosphorus, and consequently, negatively affecting the graminoid species, since the global meta-analysis showed that phosphorus tends to increase in fire-prone environments (Butler et al. 2018). In addition, in an experimental burn conducted in a vereda the phosphorus content was also elevated (Araújo et al. 2013). In this sense, further studies should be carried out to investigate the effect of fire in these environments.
In contrast, organic matter alone positively affected the herbaceous plants, while the subshrubs and shrubs were negatively affected. We presume that this is more related to the ability of herbaceous plants to produce dryer biomass and consequently more organic matter than subshrubs and shrubs, since changes in organic matter can be caused by the vegetation itself (Silva et al. 2008). Furthermore, the higher frequency of fire in vereda PO may be increasing the organic matter content of its soil, as found in a cerrado site (Silva and Batalha 2008) and in a humid subtropical grassland in the USA (Brye 2006). It is known that frequent fires alter the chemical structures of organic matter, leading to increased resistance to chemical and biological degradation, and increased stability to carbon degradation (González-Pérez et al. 2004). Only herbaceous plants were not positively or negatively affected by cation-exchange capacity, nor were their interactions with other soil properties. The higher organic matter content related to herbaceous plants may have been an advantage since organic matter can reduce soil aluminum saturation (Haynes and Mokolobate 2001).
Finally, most of the effect of soil properties on species richness and overall relative cover and growth forms depended on the interactions of these properties. This highlights the complexity of interactions among the different soil properties, particularly of synergistic interactions, illustrating the need for further studies on their interactions and greater caution in interpreting their effects. The soil–plant relationship is still complex to unravel, as it is highly influenced by moisture, temperature, and even the composition of the vegetation itself (Ehrenfeld et al. 2005), and it is often difficult to determine whether a particular soil characteristic is a cause or effect of differences in vegetation (Haridasan 2008).
In this study, we presented results about the soil-vegetation interaction of three vereda sites located in the Chapadão do São Francisco ecoregion, which is among the most endangered areas of the Cerrado due to land-use change and a lack of protected areas (Sano et al. 2019). Veredas are highly important as filters and water reservoirs in the Cerrado (Lima 2011) and its graminoid species are essential to protect the soil from erosion and runoff (Guimarães et al. 2002). Thus, many studies are still needed to better understand this environment (Fagundes and Ferreira 2016). We hope to contribute information regarding the influence of soil properties on the structure and floristic diversity of veredas. We believe that better understanding of the ways that environmental factors influence the variation in structure and floristic diversity of veredas can help to maintain these environments, since a better understanding of soil-vegetation relationships enables us to predict changes in vegetation dynamics and assists in decisions about areas that require conservation and restoration.
Data availability
The datasets used and/or analyzed during the study are available from the corresponding author on reasonable request.
References
Amorim PK, Batalha MA (2006) Soil characteristics of a hyperseasonal cerrado compared to a seasonal cerrado and a floodplain grassland: implications for plant community structure. Braz J Biol 66:661–670. https://doi.org/10.1590/S1519-69842006000400010
Araújo GM, Amaral AF, Bruna EM, Vasconcelos HL (2013) Fire drives the reproductive responses of herbaceous plants in a neotropical swamp. J Plant Ecol 214:1479–1484. https://doi.org/10.1007/s11258-013-0268-9
Barbosa da Silva FH, Arieira J, Parolin P, Nunes da Cunha C, Junk WJ (2016) Shrub encroachment influences herbaceous communities in flooded grasslands of a neotropical savanna wetland. App Veg Sci 19:391–400. https://doi.org/10.1111/avsc.12230
Baruch Z, Nassar JM, Bubis J (2004) Quantitative trait, genetic, environmental, and geographical distances among populations of the C4 grass Trachypogon plumosus in neotropical savannas. Divers Distrib 10:283–292. https://doi.org/10.1111/j.1366-9516.2004.00102.x
Bijos NR, Eugênio CUO, Mello TDRB, de Souza GF, Munhoz CBR (2017) Plant species composition, richness, and diversity in the palm swamps (veredas) of central Brazil. Flora 236:94–99. https://doi.org/10.1016/j.flora.2017.10.002
Bleich ME, Mortati AF, André T, Piedade MTF (2014) Riparian deforestation affects the structural dynamics of headwater streams in Southern Brazilian Amazonia. Trop Conserv Sci 7:657–676. https://doi.org/10.1177/194008291400700406
Brye KR (2006) Soil physiochemical changes following 12 years of annual burning in a humid–subtropical tallgrass prairie: a hypothesis. Acta Oecol 303:407–413. https://doi.org/10.1016/j.actao.2006.06.001
Bueno ML, Dexter KG, Pennington RT et al (2018) The environmental triangle of the Cerrado domain: ecological factors driving shifts in tree species composition between forests and Savannas. J Ecol 106:2109–2120. https://doi.org/10.1111/1365-2745.12969
Butler OM, Elser JJ, Lewis T, Mackey B, Chen C (2018) The phosphorus-rich signature of fire in the soil–plant system: a global meta-analysis. Ecol Lett 3:335–344. https://doi.org/10.1111/ele.12896
Canfield RH (1941) Application of the line intercept method in sampling range vegetation. J For 39:388–394
Chapin CT, Bridgham SD, Pastor J (2004) pH and nutrient effects on above-ground net primary production in a Minnesota. USA Bog Fen Wetl 24:186–201. https://doi.org/10.1672/0277-52122004024[0186:PANEOA]2.0.CO;2
Chen RF, Zhang FL, Zhang QM, Sun QB, Dong XY, Shen RF (2011) Aluminium-phosphorus interactions in plants growing on acid soils: does phosphorus always alleviate aluminium toxicity? J Sci Food Agric 92:995–1000. https://doi.org/10.1002/jsfa.4566
Cianciaruso MV, Batalha MA (2008) A year in a Cerrado wet grassland: a non-seasonal island in a seasonal savanna environment. Braz J Biol 68:495–501. https://doi.org/10.1590/S1519-69842008000300005
Dansereau P (1951) Description and recording of vegetation upon a structural basis. Ecology 32:172–229. https://doi.org/10.2307/1930415
Dantas V, Batalha MA (2011) Vegetation structure: fine scale relationships with soil in a Cerrado site. Flora 206:341–346. https://doi.org/10.1016/j.flora.2010.11.003
Durigan G (2020) Zero-fire: not possible nor desirable in the Cerrado of Brazil. Flora 268:151612. https://doi.org/10.1016/j.flora.2020.151612
Ehrenfeld JG, Ravit B, Elgersma K (2005) Feedback in the plant-soil system. Annu Rev Environ Resour 30:75–115. https://doi.org/10.1146/annurev.energy.30.050504.144212
Ellis S, Mellor A (1995) Soils and environment. Routledge, New York
Fagundes NCA, Ferreira EJ (2016) Veredas (Mauritia flexuosa palm swamps) in the southeast Brazilian Savanna: floristic and structural peculiarities and conservation status. Neotrop Biol Conserv 11:178–183. https://doi.org/10.4013/nbc.2016.113.07
Flora do Brasil (2020) Jardim Botânico do Rio de Janeiro. http://floradobrasil.jbrj.gov.br/. Accessed: 12 ago. 2021
Furley PA, Ratter JA (1988) Soil resources and plant communities of the central Brazilian Cerrado and their development. J Biogeogr 15(1):97–108. https://doi.org/10.2307/2845050
González-Pérez JA, González-Villa FJ, Almendros G, Knicker H (2004) The effect of fire on soil organic matter—a review. Environ Int 30:855–870. https://doi.org/10.1016/j.envint.2004.02.003
Guimarães AJM, Araújo GMD, Corrêa GF (2002) Estrutura fitossociológica em área natural e antropizada de uma vereda em Uberlândia. MG Acta Bot Bras 16:317–329. https://doi.org/10.1590/S0102-33062002000300007
Haridasan M (2008) Nutritional adaptations of native plants of the Cerrado biome in acid soils. Braz J Plant Physiol 20:183–195. https://doi.org/10.1590/S1677-04202008000300003
Haynes RJ, Mokolobate MS (2001) Amelioration of Al toxicity and P deficiency in acid soils by additions of organic residues: a critical review of the phenomenon and the mechanisms involved. Nutr Cycl Agroecosyst 59:47–63. https://doi.org/10.1023/A:1009823600950
Jardim CH, Moura FP (2018) Variações dos totais de chuvas e temperatura do ar na bacia do rio Pandeiros, norte do estado de Minas Gerais-Brasil: articulação com fatores de diferentes níveis escalares em área de transição climática de cerrado para semiárido. Revista Brasileira de Climatologia. https://revistas.ufpr.br/revistaabclima/article/view/61013/36761. Accessed 20 July 2020
Junk WJ, Piedade MTF, Lourival R et al (2014) Brazilian wetlands: their definition, delineation, and classification for research, sustainable management, and protection. Aquatic Conserv 24:5–22. https://doi.org/10.1002/aqc.2386
Keddy PA (2010) Wetland ecology: principles and conservation. Cambridge University Press, Cambridge, UK
Kindt R, Coe R (2005) Tree diversity analysis: a manual and software for common statistical methods for ecological and biodiversity studies. World Agroforestry Centre, Nairobi, Kenya
Legendre P, Legendre L (2012) Numerical ecology. Elsevier, Amsterdam
Leite MB, Xavier RO, Oliveira PTS, Silva FKG, Matos DMS (2018) Groundwater depth as a constraint on the woody cover in a neotropical Savanna. Plant Soil 426:1–15. https://doi.org/10.1007/s11104-018-3599-4
Lima JEFW (2011) Situação e perspectivas sobre as águas do cerrado. Ciência e Cultura 63:27–29
Liu S, Hou X, Yang M, Cheng F, Coxixo A, Wu X, Zhang Y (2018) Factors driving the relationships between vegetation and soil properties in the Yellow River Delta, China. CATENA 165:279–285. https://doi.org/10.1016/j.catena.2018.02.004
Luvuno LB, Kotze DC, Kirkman KP (2016) Long-term landscape changes in vegetation structure: fire management in the wetlands of KwaMbonambi South Africa. Afr J Aquat Sci 41:279–288. https://doi.org/10.2989/16085914.2016.1177482
Mendes MRA, Silva MC, Castro AAJF, Takahashi FSC, Munhoz CBR (2014) Temporal change in species and functional plant traits in the moist grassland on the Sete Cidades National Park, Piauí Brazil. Braz J Biol 74:111–123. https://doi.org/10.1590/1519-6984.18812
Munhoz CBR, Felfili JM (2006) Fitossociologia do estrato herbáceo-subarbustivo de uma área de campo sujo no Distrito Federal. Brasil Acta Bot Bras 20:671–685. https://doi.org/10.1590/S0102-33062006000300017
Munhoz CBR, Felfli JM, Rodrigues C (2008) Species-environment relationship in the herb-subshrub layer of a moist Savanna site, Federal District, Brazil. Braz J Biol 68:25–35. https://doi.org/10.1590/S1519-69842008000100005
Nkheloane T, Olaleye AO, Mating R (2012) Spatial heterogeneity of soil physico-chemical properties in contrasting wetland soils in two agro-ecological zones of Lesotho. Soil Res 50:579–587. https://doi.org/10.1071/SR12145
Oksanen J, Blanchet F, Friendly M, et al. (2015) Vegan community ecology package: ordination methods. Diversity analysis and other functions for community and vegetation ecologists. R package, 2–3
Oliveira GC, Araújo GM, Barbosa AAA (2009) Florística e zonação de espécies vegetais em veredas no Triângulo Mineiro Brasil. Rodriguesia 60:1077–1085. https://doi.org/10.1590/2175-7860200960417
Pilon NA, Cava MG, Hoffmann WA et al (2021) The diversity of post-fire regeneration strategies in the cerrado ground layer. J Ecol 109:154–166. https://doi.org/10.1111/1365-2745.13456
Pinheiro J, Bates D, DebRoy S, Sarkar D (2019) 'Nlme: linear and nonlinear mixed effects models. R Package version 3.1–140
R Core Team (2019) R version 3.6.0: a language and environment for statistical computing. R Found. Stat. computing, Vienna, Austria. http://www.R-project.org/. Accessed 01 January 2021
Ramos MVV, Curi N, Motta PEFD et al (2006) Veredas do Triângulo Mineiro: Solos, água e uso. Ciênc Agrotec 30:283–293. https://doi.org/10.1590/S1413-70542006000200014
Ramos FT, Dores EFC, Weber OLS, Beber DC, Campelo JH, Maia JCS (2018) Soil organic matter doubles the cation exchange capacity of tropical soil under no-till farming in Brazil. J Sci Food Agric 98:3595–3602. https://doi.org/10.1002/jsfa.8881
Ribeiro JF, Walter BMT (2008) As principais fitofisionomias do bioma Cerrado. In Sano SM, Almeida SP, Ribeiro JF (ed) Cerrado: ecologia e flora. Embrapa, Brasília, 151–212
Sano EE, Rodrigues AA, Martins ES et al (2019) Cerrado ecoregions: a spatial framework to assess and prioritize Brazilian savanna environmental diversity for conservation. J Environ Manag 232:818–828. https://doi.org/10.1016/j.jenvman.2018.11.108
Santos HG, Jacomine PKT, Anjos LHC, et al. (2018) Brazilian soil classification system, 5th. Embrapa, Brasília, DF
Shi J, Ma K, Wang J, Zhao J, He K (2010) Vascular plant species richness on wetland remnants is determined by both area and habitat heterogeneity. Biodivers Conserv 19:1279–1295. https://doi.org/10.1007/s10531-009-9757-5
Sigua GC, Kang WJ, Coleman SW (2006) Soil profile distribution of phosphorus and other nutrients following wetland conversion to beef cattle pasture. J Environ Qual 35:2374–2382. https://doi.org/10.2134/jeq2006.0092
Silva DM, Batalha MA (2008) Soil–vegetation relationships in Cerrados under different fire frequencies. Plant Soil 311:87. https://doi.org/10.1007/s11104-008-9660-y
Silva LCR, Sternberg L, Haridasan M et al (2008) Expansion of gallery forests into central Brazilian savannas. Glob Change Biol 149:2108–2118. https://doi.org/10.1111/j.1365-2486.2008.01637.x
Silva DPD, Amaral AG, Bijos NR, Munhoz CBR (2018) Is the herb-shrub composition of veredas (Brazilian palm swamps) distinguishable? Acta Bot Bras 32:47–54. https://doi.org/10.1590/0102-33062017abb0209
Sousa RFD, Brasil EPF, Figueiredo CCD, Leandro WM (2015) Soil organic matter fractions in preserved and disturbed wetlands of the Cerrado biome. Rev Bras Ciênc Solo 39:222–231. https://doi.org/10.1590/01000683rbcs20150048
Souza GF, Almeida RF, Bijos NR, Fagg CW, Munhoz CBR (2021) Herbaceous-shrub species composition, diversity and soil attributes in moist grassland, shrub grassland and savanna in Central Brazil. Braz J Bot 44:227–238. https://doi.org/10.1007/s40415-020-00672-x
Teixeira PC, Donagemma GK, Fontana A, Teixeira WG (2017) Manual de métodos de análise de solo. Embrapa, Rio de Janeiro, p 573
Ter Steege H, Pitman NC, Sabatier D et al (2013) Hyperdominance in the Amazonian tree flora. Science 342:1243092. https://doi.org/10.1126/science.1243092
Venables WN, Ripley BD (2002) Modern applied statistics with S-PLUS. Springer, New York
Weil RR, Brady NC (2016) The nature and properties of soils. Pearson, Edinburgh Gate, Harlow, Reino Unido, UK
Xavier R, Leite MB, Dexter K, da Silva Matos DM (2019) Differential effects of soil waterlogging on herbaceous and woody plant communities in a neotropical savanna. Oecologia 190:471–483. https://doi.org/10.1007/s00442-019-04423-y
Xiaolong W, Jingyi H, Ligang X, Rongrong W, Yuwei C (2014) Soil characteristics in relation to vegetation communities in the wetlands of Poyang Lake, China. Wetlands 34:829–839. https://doi.org/10.1007/s13157-014-0546-x
Zar JH (2010) Biostatistical analysis. Pearson Prentince-Hall, Upper Saddle river, NJ New Jersey
Zuur A, Ieno EN, Walker N, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
We are grateful to ICMBio, especially for hosting the team in the Grande Sertão Veredas Park lodgings. For identifications of botanical material, we thank J.B. Bringel (Asteraceae), J.E.Q. Faria Junior (several families), V.L. Rivera (Asteraceae), C.E.B. Proenca (several families), and A.L.R. Oliveira (Eriocaulaceae).
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brazil (CAPES), Finance Code 001. Collection of field data was supported by the Fundação de Apoio à Pesquisa do Distrito Federal (FAPDF 193.001.549/2017).
Author information
Authors and Affiliations
Contributions
EVN and CBRM developed the ideas and designed the experimental methods. EVN, CBRM, NRB and VLT collected the data. EVN, CBRM, NRB, PHBT and GPH analyzed the data. EVN and CBRM wrote the first draft of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Nogueira, E.V., Bijos, N.R., Trindade, V.L. et al. Differences in soil properties influence floristic changes in the Veredas of the Brazilian Cerrado. Braz. J. Bot 45, 763–774 (2022). https://doi.org/10.1007/s40415-022-00795-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40415-022-00795-3