
Abstract
Cynanchum thesioides (Freyn) K. Schum. with higher drought tolerance has received much research attention in arid and semiarid areas of China. However, no full evaluation of drought resistance had been made. In the current study, we compared relative water content, water saturation deficit, root vitality, root–cap ratio, photosynthetic pigments and ten physiological indices in C. thesioides and C. thesioides (Freyn) K. Schum. var. australe (Maxim.) Tsiang et P.T. Li. seedlings under a gradient of drought stress. The ten indices included superoxide dismutase, peroxidase, catalase, ascorbate peroxidase and malondialdehyde, hydrogen peroxide, superoxide, proline, soluble protein and sugar content of leaves and roots. Based on the analysis of average membership functional values, the drought tolerance of C. thesioides var. australe was better than C. thesioides. In the future, C. thesioides var. australe should be given priority when selecting varieties for use as medicines in arid and semiarid areas of China.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Drought is a major stress factor in plants, under drought stress plant morphology, physiology and gene expression changes (Comas et al. 2013; Zhang et al. 2015). For example, when confronted with a deficit of soil water, root activity drops and growth of the aerial parts of the plant are inhibited. However, this also promotes elongation of the root, increases root–shoot ratio and improves water use efficiency (Shan et al. 2008). Leaf color is a clear expression of the pigments present in leaves. Chlorophyll is the predominant pigment, and the quantity of chlorophyll determines plant growth capacity and is an indicator of stress level. Water stress during drought can change the quantity of chlorophyll a and chlorophyll b in plants leading to changes in photosynthesis capacity (Miao et al. 2015; Shukla et al. 2015).
Osmotic regulation occurs in plants under water stress, which results in cell synthesis and the absorption and accumulation of particular compounds; within-plant levels of these compounds are positively correlated with drought tolerance as they maintain cell turgor under low water potential, reducing wilting and maintaining the capacity for growth. At the same time, accumulation of solutes can protect cell proteins, enzymes, organelles and cell membranes from damage due to dehydration. These solutes also have an anti-ageing function as they scavenge free radicals under conditions of severe drought stress (Sapeta et al. 2015; Nolan et al. 2017).
The balance between production and scavenging of reactive oxygen species (ROS) is disrupted in plant cells during drought stress, and this results in a variety of harmful cytological effects (Choudhury et al. 2017; Verma et al. 2019; Wei et al. 2019). Malondialdehyde (MDA) is an indicator of lipid peroxidation and plasma membrane damage in plant cells (Mohammadi et al. 2011; Maqbool et al. 2017). The non-enzymatic and enzymatic antioxidants that scavenge ROS contribute to the long-term survival of plants as they retain ROS at a moderate level thus preventing cell damage (Huang et al. 2013; Zhu 2016). Recent studies have suggested that some antioxidative enzymes protect plants from damage during mild and moderate drought stress, but are less effective under severe drought stress (Jia et al. 2015).
Cynanchum thesioides (Freyn) K. Schum. are erect, xerophytic subshrubs (Chen and Qian 1959), present in arid and semiarid regions. C. thesioides is economically and ecologically important: It produces edible fruits; is used in the production of medicines, animal feed and industrial raw materials; and improves soil quality and water conservation in pastures (Zhang et al. 2019). In recent years, more and more people have become familiar with C. thesioides as a medicinal plant that can cure tonifying qi, lactogenesis and warts (Liang 2012) and have begun cultivating them.
The aim of the present study was to quantify osmotically active compounds, chlorophyll, ROS and antioxidant enzymes in the leaves and roots of C. thesioides under drought stress. We aimed to elucidate the physiological response mechanisms that C. thesioides uses to tolerate water deficit and moreover to determine whether C. thesioides or C. thesioides (Freyn) K. Schum. var. australe (Maxim.) Tsiang et P. T. Li were more drought resistant.
2 Materials and methods
Plant materials and experimental design
– Seeds of both C. thesioides and C. thesioides var. australe were collected in Chifeng city in the Inner Mongolia Autonomous Region of China. The seeds were surface-sterilized in 2% (v/v) NaOCl for 15 min, rinsed three times in distilled water and pre-germinated at 25 °C in darkness for 3 days on wet filter paper. Germinated seeds were sown 2-3 cm deep in plastic pots (260 cm height × 180 cm diameter, 0.9 kg weight) each containing 2.4 kg of compost that was comprised of a mixture of sand, soil and fermented sheep dung (7:5:2; v/v/v); eight seeds were sown in each pot. Seedlings were grown in a greenhouse (mean temperatures of 26/19 °C [day/night]) at the Inner Mongolia Agricultural University, Huhhot, Inner Mongolia Province, Northwest China (111°69′E,40°80′N).
At the ten-leaf stage, morphologically uniform seedlings were selected for experimentation. Seedlings were randomly divided into four groups (n = 20 per treatment group for both C. thesioides and C. thesioides var. australe) and assigned to different drought stress treatments as described by Hsiao (1973). The four drought stresses were related to soil moisture content and represented different percentages of the maximum water holding capacity (24.4%) of the soil, specifically: 65-70% (control), 45-50% (mild drought), 25-30% (moderate drought) and 5-10% (severe drought), respectively. To achieve the different drought treatments, we first saturated each pot with water and then let them dry naturally until they reached the required moisture levels, as determined by weighing, drying and re-drying samples of the soil to determine water content. Every day at 5 p.m., the soil moisture content was determined in the same way, and any water lost was replaced to ensure the moisture content was maintained at the required level for each treatment. After 5 days the leaves and roots were harvested and the related physiological indices were determined, each experimental data were repeated four times.
Estimation of relative water content and water saturation deficit
– Relative water content and water saturation deficit were measured four replicates in each treatment, following Barrs and Weatherley (1962). Fresh leaves were weighed (Wf) and soaked in water overnight until the weight of leaves was constant. The saturated leaves were weighed (Wt) and then dried for 24 h at 80 °C to determine the dry weight (Wd). The RWC and WSD were calculated using the formula:
Quantifying photosynthetic pigments
– Photosynthetic pigments were extracted by soaking 0.2 g of fresh leaves in 25 mL of 95% ethanol. The absorbance of each extract was measured at 665 nm, 649 nm and 470 nm using a spectrophotometer (Li 2000). Chlorophyll a (Chl a), chlorophyll b (Chl b), total chlorophyll (T-Chl) and carotenoids (Cx.c) were calculated using the equations of Li (2000).
Estimation of root vitality and root–cap ratio
– The 2,3,5-triphenyltetrazolium chloride (TTC) method was used to determine the root vitality. Root vitality was calculated using the equations of Li (2000), and RCR was calculated as the ratio of root dry weight to shoot dry weight.
Determining the content of the osmolytes: soluble protein and soluble sugar
– To measure the soluble sugar content, 0.3 g fresh leaves from each replicate was homogenized in 10 mL distilled water. The samples were stirred and maintained in a water bath at 100 °C for 30 min. Extract was merged and made a total volume to 25 mL. Total soluble protein was extracted from 0.25 g leaf with 5 mL of distilled water and then 5 mL Coomassie brilliant blue G-250 after centrifugation 3000 r min−1 for 10 min. Protein and sugar content was determined by the procedure of Li (2000).
Extraction and quantification of H2O2, malondialdehyde (MDA) and O2
– From each replicate, 0.25 g of fresh leaves was immersed in liquid nitrogen to which 20% polyvinylpolypyrrolidone (PVPP) had been added. The mixture was homogenized with 5 mL 0.1% trichloroacetic acid (TCA) and centrifuged at 10000 g for 10 min at 4 °C. The supernatant was used to determine hydrogen peroxide and MDA concentrations. H2O2 was measured using an ultraviolet–visible spectrophotometer in accordance with Patterson et al. (1984). MDA concentration was determined following the method of Li (2000) and used as a measure of the level of membrane damage. The production ratio of superoxide (O.−2) was measured according to the methods of Elstner and Heupel (1976).
Enzymatic activity
– ÒEnzymes were extracted by immersing 0.5 g of leaves from each replicate in liquid nitrogen, to which 1.5 mL of extraction buffer had been added; extraction buffer was comprised of 100 mM potassium phosphate buffer (pH 7.0), 1 mM ethylenediaminetetraacetic acid (EDTA), 2 mM dithiothreitol (DTT), 0.8 mM phenylmethylsulfonyl fluoride (PMSF), 1% PVPP and 1 mM ascorbic acid (ASA). The mixture was centrifuged at 14000 rpm per 30 min at 4 °C, and the supernatant was collected and stored at -80 °C prior to analysis. Each supernatant was evaluated for activity of all the enzymes of interest as described below.
The activity of superoxide dismutase (SOD) was determined using the method of Giannopolitis and Ries (1977) with some modifications. The absorbance of each solution was determined at 560 nm using an ultraviolet–visible spectrophotometer. One unit of enzyme activity was defined as the amount of enzyme that would inhibit 50% of blue tetrazolium (NBT) photoreduction.
The activity of catalase (CAT) was determined in relation to its ability to oxidize H2O2 using the method of Chance and Maehly (1955). Changes in absorbance at 240 nm were read every 1 min for 3 min using an ultraviolet–visible spectrophotometer. One unit of CAT activity was defined as a change in absorbance of 0.01 units per minute.
The activity of peroxidase (POD) was evaluated in relation to its ability to oxidize guaiacol. Changes in absorbance at 470 nm were read every 1 min for 3 min using an ultraviolet–visible spectrophotometer. One unit of POD activity was defined as a change in absorbance of 0.01 units per minute.
Ascorbate peroxidase (APX) activity was measured in relation to its ability to oxidize ascorbate using the methods of Nakano and Asada (1981). Changes in absorbance at 290 nm were read every 1 min for 5 min using an ultraviolet–visible spectrophotometer. One unit of APX activity was defined as a change in absorbance of 0.01 units per minute.
Statistical methods
– Data were presented as mean values ± SE. Significant differences among different water stress treatments were determined by ANOVA. All data were analyzed using the statistical software package SAS version 9.0. The least significant difference (LSD) was used to compare means at P ≤ 0.05. Drought resistance evaluation was adopted by membership function (Chen et al. 2012; Zhang et al. 2017) and principal components analysis (Shen 2018).
3 Results
Relative water content and water saturation deficit of leaves
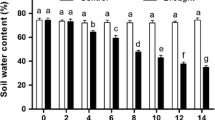
– As drought stress increased, the relative water content of C. thesioides leaves first increased and then decreased; it was largest under mild drought conditions, followed by the control and then moderate drought and smallest under severe drought. Relative water content of CT and CA plants under severe drought was reduced by 46.45% and 69.25%, respectively, compared with the control, and this was statistically significant (P ≤ 0.05). Under mild drought, relative water content was significantly higher than in all other treatments (P ≤ 0.05). However, changes in water saturation deficit followed the opposite trend to the relative water content of leaves. Under mild drought, water saturation deficit was significantly lower than in all other treatments (P ≤ 0.05) and reached the maximum value under severe drought (Table 1).
Quantity of photosynthetic pigments
– As drought stress increased, the quantity of photosynthetic pigments increased (Table 2). This increase is not significantly different between the control and mild drought stress conditions. Compared with the control, the increases in chlorophyll a, chlorophyll b, total chlorophyll and carotenoids in CT and CA plants under severe drought stress were 44.07%, 35.44%, 67.88%, 18.23%, 50.54% and 37.72%, respectively. The ratio of total chlorophyll/carotenoid in CT plants increased with increasing drought stress; it was 7.67, 7.70, 9.08 and 10.13, respectively, under the different drought conditions. However, this ratio in CA plants increased initially, but then decreased; it was 8.52, 8.61, 8.72 and 8.15, respectively, under the different drought conditions.
Root vitality and root–cap ratio
– As drought stress increased, the root vitality of CT plants gradually decreased; compared with the control, root vitality under mild, moderate and severe drought decreased by 45.40%, 44.90% and 46.37%, respectively, and this was statistically significant (P ≤ 0.05). As drought stress increased, the root vitality and root–cap ratio of CA plants increased; compared with the control, they increased 1 and 0.4 times under severe water stress, respectively. The root–cap ratio of CT plants was higher than that of CA plants (Table 3).
Quantity of osmolytes
– The soluble sugar in leaves of CT plants decreased under drought stress, but it increased in CA plants (Fig. 1a). Soluble sugar content of roots under severe drought was two times higher than that in leaves under the same conditions. The soluble protein content of leaves of CT and CA plants under severe drought was higher than in roots (Fig. 1b). The proline content did not change significantly in leaves of CT plants under drought stress (Fig. 1c). Compared with the control, the proline content of leaves from CA plants and roots from CT and CA plants increased significantly (Fig. 1c).

Soluble sugar (a), protein (b) and proline (c) content of the leaves and roots of two varieties of C. thesioides grown under drought stress. CTL: leaves of C. thesioides; CAL: leaves of C. thesioides var. australe; CTR: roots of C. thesioides; and CAR: roots of C. thesioides var. australe. Different lowercase letters indicate significant difference for the same organs and plant variety at P ≤ 0.05 level among treatments
Quantity of H2O2, MDA and O2
– As drought stress increased, the production ratio of superoxide (O2−) in leaves and roots of CT and CA plants increased; compared with the control, it increased by 35.69% (CT leaves), 36.87% (CT roots), 23.88% (CA leaves) and 54.99% (CA roots) under severe drought stress. In leaves, it was at least two times higher than in roots (Fig. 2a). As drought stress increased, H2O2 content in roots and leaves of CT and CA plants also increased. H2O2 content of leaves was higher than in roots under different drought stress conditions. H2O2 content of leaves from CT plants was higher than that in leaves from CA plants in the same treatment, while that in roots of CT plants was lower than in roots of CA plants in the same treatment (Fig. 2b). MDA content in leaves and roots of CT and CA plants increased as drought stress increased; compared with the control, it increased by 31.91% (CT leaves), 54.30% (CT roots), 37.17% (CA leaves) and 31.72% (CA roots) under severe drought stress. MDA content of CT plants was higher than in CA plants (Fig. 2c).

O.−2generation rate (a), H2O2 (b) and MDA (c) content in leaves and roots of two varieties of C. thesioides grown under drought stress. CTL: leaves of C. thesioides; CAL: leaves of C. thesioides var. australe; CTR: roots of C. thesioides; CAR: roots of C. thesioides var. australe; O.−2: superoxide radical; H2O2: hydrogen peroxide; and MDA: malondialdehyde. Different lowercase letters indicate significant difference for the same organs and plant variety at P ≤ 0.05 level among treatments
Activity of antioxidant enzymes
– The activities of SOD, CAT, POD and APX were all affected by drought stress. CAT activity in leaves and roots of C. thesioides increased slightly compared with the control, peaking under moderate drought stress conditions; while it declined under severe drought (Fig. 3a).

CAT (a), SOD (b), POD (c), APX (d) activity in leaves and roots of two varieties of C. thesioides grown under drought stress. CTL: leaves of C. thesioides; CAL: leaves of C. thesioides var. australe; CTR: roots of C. thesioides; CAR: roots of C. thesioides var. australe; CAT: catalase; SOD: superoxide dismutase; POD: peroxidase; and APX ascorbate peroxidase. Different lowercase letters indicate significant difference for the same organs and plant variety at P ≤ 0.05 level among treatments
As drought stress increased, SOD activity in roots and leaves of CT plants first increased and then decreased; however, in roots and leaves of CA plants, it increased gradually. SOD activity of leaves from CT plants was higher than in leaves from CA plants, but its activity in roots of CT plants was higher than in roots of CA plants under low drought stress conditions and lower in severe drought stress conditions (Fig. 3b).
POD activity in CT and CA plants was at least 2.65 and 1.77 times higher in roots than in leaves, respectively. In roots of CT and CA plants, POD activity reached its maximum under severe drought (2.67 U mg−1 min−1) and moderate drought (3.61 U mg−1 min−1) conditions, respectively. In leaves of CT and CA plants, it reached its maximum under control and severe water stress conditions, respectively (Fig. 3c).
APX activity in leaves of CT and CA plants was at least 3.52- and 4.09-fold higher than in roots, respectively, and CA plants had greater activity than CT plants. The highest specific activity of APX was observed under severe drought stress conditions in leaves and roots of CT plants and in the roots of CA plants (Fig. 3d).
Drought resistance evaluation
– In this experiment, the cumulative rates of variance for the first five components measured accounted for more than 80% of the total variance: 39.79%, 26.04%, 15.08%, 9.20% and 6.07% (Table 4). The absolute value was greater than 0.8 in the five principal component matrix following fuzzy membership function analysis. The average of membership function of CA plants was 0.4713, which was greater than CT plants (Table 4).
4 Discussion
In recent years, water shortages have become increasingly more serious throughout the world; it is thus very important to determine how to achieve normal plant growth without excessive water use. The relative water content and water saturation deficit of leaves reflect the water holding capacity of plants and are the link between physiological characteristics and levels of drought tolerance (Marshall et al. 2000; Farooq et al. 2009). Maintaining an appropriate water state in plant tissues helps them avoid dehydration and protects against inactivation and denaturation of carboxylase and other enzymes (Sanchez-Rodriguez et al. 2010). The opposite trend is observed under drought stress, as demonstrated in Sorghum bicolor L. Moench and blackberry (Rubus L.) (Ogbaga et al. 2014; Zhang et al. 2017). A strong capacity for water retention in plants allows them to limit the extent of decreased relative water content and increase the water saturation deficit. Thus, it can be seen that water capacity of C. thesioides was better than that of C. thesioides var. australe. Chlorophyll a, chlorophyll b, total chlorophyll and carotenoids all increased to various degrees under drought stress; this may be as a result of compensation and over-compensation effects and was similar to the results of Zhang et al. (2017).
Previous studies have shown that an increase in osmotic regulators can improve water use efficiency, eliminate oxidative stress and improve drought resistance in plants (Elansary and Yessoufou 2015). So, proline accumulation in C. thesioides suggests that it plays an important role in osmotic protection. In our research, proline levels increased more in leaves than in roots under severe drought, but when there was a water deficit, there was no significant accumulation in leaves. Peak levels were observed in roots of C. thesioides and in leaves of C. thesioides var. australe under severe drought. Recent evidence suggests that proline synthesis may occur in the shoots and is probably transported to the roots in order to sustain normal root growth under low water potential (Sharma et al. 2011).
Accumulation of ROS is induced by various stresses but is scavenged by other components, such as antioxidant enzymes, non-enzymatic antioxidants and secondary metabolites (Gill et al. 2011; Wei et al. 2019), as has been demonstrated recently in Diospyros L. (Wei et al. 2015) and blackberry (Rubus sp.) (Yang et al. 2015). Drought resistance of Rubus sp. ‘Ningzhi 1’ increased due to a decrease in lipid peroxidation and larger quantities of antioxidant enzymes (Yang et al. 2015). An increase in antioxidant enzyme activity is a common adaptive response of plants to drought stress (Gill and Tuteja 2010). The main function of SOD is to transform O.−2 into O2 and H2O2, which are toxic lipid peroxidation, but can be eliminated by POD and CAT (Sabine et al. 2004). We observed increased POD and CAT activity as a result of drought stress, which may protect plants from oxidation. POD also plays an important role in eliminating MDA, thus protecting the permeability of cell membranes (Hojati et al. 2011). In our study, POD activity was increased under mild, moderate and severe drought stress, which may limit any increases in MDA incurred as a result of drought stress. Our results for increased POD activity were similar to studies on Carthamus tinctorius L. (Hojati et al. 2011). In our study, SOD activity increased in response to mild and moderate drought stress, which confirmed previous reports by Lum et al. (2014) and Pandey and Shukla (2015). APX is a component of the AsA–GSH cycle, which can efficiently protect plants from ROS. In our study, APX levels significantly increased under drought stress and compared with the control; in leaves of C. thesioides and C. thesioides var. australe, it was at 3.52 and 4.09 times higher levels than in roots, respectively. This suggested that APX in the AsA–GSH cycle could contribute to antioxidant activity during drought stress and that leaves were more sensitive than roots. We also observed decreases in the activity of most enzymes under severe drought, because the synthesis of ROS exceeded the capacity of the enzyme protection system, resulting in extensive membrane lipid peroxidation. Moreover, the relative water content decreased and MDA levels increased, peaking under severe drought conditions. We observed that antioxidant enzymes could reduce ROS, under certain drought conditions, unless the production rate of antioxidant enzymes was less than that of ROS. The plants were capable of appropriate responses to adapt to drought environments.
The membership function gave a comprehensive assessment based on the theory of fuzzy mathematics. In this study, drought-affected physiological responses differed between C. thesioides and C. thesioides var. australe; we described drought tolerance by evaluating the membership function of C. thesioides based on physiological indices. The greater membership function represented the higher drought resistance. The drought resistance of C. thesioides var. australe was better than C. thesioides.
In conclusion, C. thesioides demonstrated comprehensive drought tolerance features including accumulation of osmotic regulators and activation of enzymatic antioxidant systems. These actions prevented tissue damage under drought stress. This is the first paper to report changes in physiological indices in two varieties of C. thesioides under drought stress.
Abbreviations
- CT:
-
Cynanchum thesioides (Freyn) K. Schum.,
- CA:
-
Cynanchum thesioides (Freyn) K. Schum. var. australe (Maxim.) Tsiang- et P.T. Li
- CK:
-
Control
- LS:
-
Mild drought stress
- MS:
-
Moderate drought stress
- SS:
-
Severe drought stress
- O2− :
-
Superoxide
References
Barrs HD, Weatherley PE (1962) A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust J Biol Sci 15:413–428. https://doi.org/10.1071/BI9620413
Bonos SA, Rush D, Hignight K, Meyer WA (2004) Selection for deep root production in tall fescue and perennial ryegrass. Crop Sci 44:1770–1775. https://doi.org/10.2135/cropsci2004.1770
Chance B, Maehly AC (1955) Assay of catalase and peroxidase. Method Enzymol 136:764–775. https://doi.org/10.1016/S0076-6879(55)02300-8
Chen CS, Qian HY (1959) Flora reipublicae popularis sinicae. Beijing, China
Chen X, Min D, Yasir T, Hu Y (2012) Evaluation of 14 morphological yield-related and physiological traits as indicators of drought tolerance in Chinese winter bread wheat revealed by analysis of the membership function value of drought tolerance (MFVD). Field Crop Res 137:195–201. https://doi.org/10.1016/j.fcr.2012.09.008
Choudhury FK, Rivero RM, Blumwald E, Mittler R (2017) Reactive oxygen species, abiotic stress and stress combination. Plant J 90:856–867. https://doi.org/10.1111/tpj.13299
Comas LH, Becker SR, Cruz VM, Byrne PF, Dierig DA (2013) Root traits contributing to plant productivity under drought. Front Plant Sci 4:442–457. https://doi.org/10.3389/fpls.2013.00442
Elansary HO, Yessoufou K (2015) Growth regulators and mowing heights enhance the morphological and physiological performance of Seaspray turfgrass during drought conditions. Acta Physiol Plant 37:232–242. https://doi.org/10.1007/s11738-015-1986-5
Elstner EF, Heupel A (1976) Inhibition of nitrite formation from hydroxylammoniumchloride: a simple assay for superoxide dismutase. Anal Biochem 2:616–620. https://doi.org/10.1016/0003-2697(76)90488-7
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009) Plant drought stress: effects, mechanisms and management. Agron Sustain Dev 29:185–212. https://doi.org/10.1051/agro:2008021
Giannopolitis CN, Ries SK (1977) Superoxide dismutase. I. Occurrence in higher plants. Plant Physiol 59:309–314. https://doi.org/10.1104/pp.59.2.309
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930. https://doi.org/10.1016/j.plaphy.2010.08.016
Gill SS, Khan NA, Anjum NA, Tuteja N (2011) Amelioration of cadmium stress in crop plants by nutrients management: morphological, physiological and biochemical aspects. Plant Stress 5:1–23
Glombitza S, Dubuis P, Thulke O, Welzl G, Bovet L, Götz M, Affenzeller M, Geist B, Hehn A, Asnaghi C, Ernst D, Seidlitz H, Gundlach H, Mayer K, Martinoia E, Werck-reichhart D, Mauch F, Schäffner A (2004) Crosstalk and differential response to abiotic and biotic stressors reflected at the transcriptional level of effector genes from secondary metabolism. Plant Mol Biol 54:817–835. https://doi.org/10.1007/s11103-004-0274-3
Gou ZP, Yang YJ, Zhao RN (2001) Revision of latin names of the medicinal plants of genus Cynanchum in Gansu Province. Northwest Pharma J 16:56–57. https://doi.org/10.3969/j.issn.1004-2407.2001.02.005
Hojati M, Modarres-Sanavy SAM, Karimi M, Ghanati F (2011) Responses of growth and antioxidant systems in Carthamus tinctorius L. under water deficit stress. Acta Physiol Plant 33:105–112. https://doi.org/10.1007/s11738-010-0521-y
Hsiao TC (1973) Plant response to water stress. Annu Rev Plant Physiol 24:519–570. https://doi.org/10.1146/annurev.pp.24.060173.002511
Huang XS, Wang W, Zhang Q, Liu JH (2013) A basic helix-loop-helix transcription factor, PtrbHLH, of Poncirus trifoliata confers cold tolerance and modulates peroxidase-mediated scavenging of hydrogen peroxide. Plant Physiol 162:1178–1194. https://doi.org/10.1104/pp.112.210740
Jia X, Sun CS, Li GY, Li GB, Chen GL (2015) Effects of progressive drought stress on the physiology, antioxidative enzymes and secondary metabolites of Radix Astragali. Acta Physiol Plant 37:262–275. https://doi.org/10.1007/s11738-015-2015-4
Karcher ED, Richardson MD, Hignight K, Rush D (2008) Drought tolerance of tall fescue populations selected for high root/shoot ratios and summer survival. Crop Sci 48:771–777. https://doi.org/10.2135/cropsci2007.05.0272
Li HS (2000) Principles and techniques of plant physiological biochemical experiment. Beijing, China
Liang LM (2012) Study on the medical ethonobotany of the naiman banner mongolians in Inner Mongolia. Dissertation, Inner Mongolia Normal University
Liu CC, Liu YG, Guo K, Fan DY, Li GP, Zheng YR, Yu LF, Yang R (2011) Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China. Environ Exp Bot 71:174–183. https://doi.org/10.1016/j.envexpbot.2010.11.012
Liu DL, Zhang WH, Miao YJ, Yan TF, Lin YH, Xu YM (2017) Drought stress responses of the seedlings of three wild forages in Tibet. Acta Ecol Sin 37:7277–7285. https://doi.org/10.5846/stxb201609021789
Lum MS, Hanafi MM, Rafii YM, Akmar ASN (2014) Effect of drought stress on growth, proline and antioxidant enzyme activities of Upland rice. J Anim Plant Sci 24:1487–1493
Maqbool MA, Aslam M, Ali H (2017) Breeding for improved drought tolerance in Chickpea (Cicer arietinum L.). Plant Breed 136:300–318. https://doi.org/10.1111/pbr.12477
Marshall J, Rutledge R, Blumwald E, Dumboroff E (2000) Reduction in turgid water volume in jack pine, white spruce and black spruce in response to drought and paclobutrazol. Tree Physiol 20:701–707. https://doi.org/10.1093/treephys/20.10.701
Miao YY, Zhu ZB, Guo QS, Ma HL, Zhu LF (2015) Alternate wetting and drying irrigation-mediated changes in the growth, photosynthesis and yield of the medicinal plant Tulipa edulis. Ind Crop Prod 66:81–88. https://doi.org/10.1016/j.indcrop.2014.12.002
Mohammadi A, Habibi D, Rohami M, Mafakheri S (2011) Effect of drought stress on antioxidant enzymes activity of some chickpea cultivars. Am-Eur J Agric Environ Sci 11:782–785
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22:867–880. https://doi.org/10.1093/oxfordjournals.pcp.a076232
Nolan RH, Tarin T, Santini NS, McAdam SAM, Ruman R, Eamus D (2017) Differences in osmotic adjustment, foliar abscisic acid dynamics, and stomatal regulation between an isohydric and anisohydric woody angiosperm during drought. Plant Cell Environ 40:3122–3134. https://doi.org/10.1111/pce.13077
Ogbaga CC, Stepien P, Johnson GN (2014) Sorghum (Sorghum Bicolor) varieties adopt strongly contrasting strategies in response to drought. Physiol Plant 152:389–401. https://doi.org/10.1111/ppl.12196
Pandey V, Shukla A (2015) Acclimation and tolerance strategies of rice under drought stress. Rice Sci 22:147–161. https://doi.org/10.1016/j.rsci.2015.04.001
Patterson BD, Macrae EA, Ferguson IB (1984) Estimation of hydrogen peroxide in plant extracts using titanium (IV). Anal Biochem 139:487–492. https://doi.org/10.1016/0003-2697(84)90039-3
Qiu YX, Yang ZQ, Liu ZX, Chen QY, Tian W (2016) The Effects of soil moisture stress on the growth of root and above-ground parts of greenhouse tomato crops. Acta Ecol Sin 36:748–757. https://doi.org/10.5846/stxb201403310606
Sanchez-Rodriguez E, Rubio-Wilhelmi M, Cervilla LM, Blasco B, Rios JJ, Rosales MA, Romero L, Ruiz JM (2010) Genotypic differences in some physiological parameters symptomatic for oxidative stress under moderate drought in tomato plants. Plant Sci 178:30–40. https://doi.org/10.1016/j.plantsci.2009.10.001
Sapeta H, Lourenço T, Lorenz S, Grumaz C, Kirstahler P, Barros PM, Costa JM, Sohn K, Oliveira MM (2015) Transcriptomics and physiological analyses reveal co-ordinated alteration of metabolic pathways in Jatropha curcas drought tolerance. J Exp Bot 67:845–860. https://doi.org/10.1093/jxb/erv499
Shan LS, Zhang XM, Wang YK (2008) Influence of moisture on the growth and biomass allocation in Haloxylon ammodendron and Tamaracks ramosissima seeding in the shelterbelt along the Tarim Desert Highway, Xinjiang, China. Chin Sci Bull 53:93–101. https://doi.org/10.1007/s11434-008-6010-7
Shan LS, Yang CH, Li Y, Duan YN, Geng DM, Li ZY, Zhang R, Duan GF (2015) Effects of drought stress on root physiological traits and root biomass allocation of Reaumuria soongorica. Acta Ecol Sin 35:155–159. https://doi.org/10.1016/j.chnaes.2015.06.010
Sharma S, Villamor JG, Verslues PE (2011) Essential role of tissue specific proline synthesis and catabolism in growth and redox balance at low water potential. Plant Physiol 157:292–304. https://doi.org/10.1104/pp.111.183210
Shen HT (2018) Principal component analysis. In: Liu L, Özsu MT (eds) Encyclopedia of database systems. Springer, New York, pp 42–71. https://doi.org/10.1007/978-1-4614-8265-9_540
Shukla PS, Gupta K, Agarwal P, Jha B, Agarwal PK (2015) Overexpression of a novel SbMYB15 from Salicornia brachiata confers salinity and dehydration tolerance by reduced oxidative damage and improved photosynthesis in transgenic tobacco. Planta 242:1291–1308. https://doi.org/10.1007/s00425-015-2366-5
Verma G, Srivastava D, Tiwari P, Chakrabarty D (2019) ROS modulation in crop plants under drought stress. In: Hasanuzzaman M, Fotopoulos V, Nahar K, Fujita M (eds) Reactive oxygen, nitrogen and sulfur species in plants. Chapter 13. https://doi.org/10.1002/9781119468677.ch13
Wei P, Yang Y, Wang F, Chen HJ (2015) Effects of drought stress on the antioxidant systems in three species of Diospyros L. Hortic Environ Biote 56:597–605. https://doi.org/10.1007/S13580-015-0074-5
Wei T, Wang Y, Xie Z, Guo D, Chen C, Fan Q, Deng X, Liu J (2019) Enhanced ROS scavenging and sugar accumulation contribute to drought tolerance of naturally occurring autotetraploids in Poncirus trifoliata. Plant Biotechnol J 17:1394–1407. https://doi.org/10.1111/pbi.13064
Yang HY, Zhang CH, Wu WL, Li WL, Wei YL, Dong SS (2015) Physiological responses of blackberry cultivar ‘Ningzhi 1’ to drought stress. Russ J Plant Physiol 62:472–479. https://doi.org/10.1134/S1021443715040184
Yu HP, Huang JP, Guan XD, Wang GY, Guo RX (2016) Accelerated dryland expansion under climate change. Nat Clim Change 6:166–171. https://doi.org/10.1038/nclimate2837
Zhang M, Jin ZQ, Zhao J, Zhang GP, Wu FB (2015) Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul 75:567–574. https://doi.org/10.1007/s10725-014-0022-x
Zhang CH, Yang HY, Wu WL, Li WL (2017) Effect of drought stress on physiological changes and leaf surface morphology in the blackberry. Braz J Bot 40:625–634. https://doi.org/10.1007/s40415-017-0377-0
Zhang XY, Yang ZR, Li Z, Zhang FL, Hao LZ (2019) De novo transcriptome assembly and co-expression network analysis of Cynanchum thesioides: identification of genes involved in resistance to drought stress. Gene 710:375–386. https://doi.org/10.1016/j.gene.2019.05.055
Zhu JK (2016) Abiotic stress signaling and responses in plants. Cell 167:313–324. https://doi.org/10.1016/j.cell.2016.08.029
Acknowledgements
We thank a National Natural Science Foundation of China (31101541;31160393) Science Fund of Inner Mongolia of China (2015MS0359) Ph.D. award; Start-up Fund of Inner Mongolia Agricultural University (BT09-17); Scientific Research Project of Public Welfare Industry (Agriculture) (201203004); Inner Mongolia President Fund; and Inner Mongolia Agricultural University Science and Technology Innovation (Nurturing) Team (NDPYTD2013-3) for its financial support.
Author information
Authors and Affiliations
Contributions
H, LZ and Y, ZR supervised the experiment. Z, XY and LZ performed the experiment work and prepared the figures and tables. Z, XY wrote the manuscript. Z, FL edited the final version of the manuscript. Z, XY and Y, ZR contributed equally to this work. All authors have approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
Authors declare that there is no conflict of interest regarding the publication of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zhang, X., Yang, Z., Li, Z. et al. Effects of drought stress on physiology and antioxidative activity in two varieties of Cynanchum thesioides. Braz. J. Bot 43, 1–10 (2020). https://doi.org/10.1007/s40415-019-00573-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40415-019-00573-8