
Abstract
Diatoms, silicious microalgae, have been used successfully as bioindicators of water quality assessment in aquatic ecosystems. Diatoms have a degree of tolerance to the water quality and some diatoms are a good indicator for several water quality variables. Diatom indices have been developed to assess river water quality, mostly in Europe. This study aims to apply diatom indices developed in Europe for the tropical lake of Galela adjacent to residential areas influenced by human activities. Galela Lake is one of the biggest lakes in Halmahera Utara, Indonesia with its main functions being domestic water supply, irrigation, fisheries, and tourism. Human activities have impacted the area around the lake. The 90-cm and 85-cm long sediment cores were collected using a piston corer from Site 1 and 2, respectively. Sediment samples were sliced every 5 cm, separated from sediment by adding HCl and H2O2. The diatom valves were identified under a microscope with 1,000 × magnification. The water quality status of each layer was inferred with diatom indices performed using OMNIDIA software version 6.0. Forty-nine and 63 diatoms species were identified from Site 1 and Site 2, respectively. The number of species and diversity of diatoms was higher in the lower layers than those in the upper layers. The preserved diatom assemblages reflect past physical and chemical water quality. Generic Diatom Index and Specific Pollution Sensitivity Index provided the best evidence for change in Galela Lake—they integrated 70–100% of the diatom taxa from the sediment core samples.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Ecological status is the quality of structure and function of an aquatic ecosystem, that can be expressed by biological quality elements such as macrophytes and phytobenthos, mostly diatoms (Kelly et al. 2009). Diatoms are microalgae that inhabit a wide variety of aquatic habitats, even growing in humic soils (Kociolek et al. 2020). Diatoms play an important role in paleolimnological studies as they are often well preserved in lake sediments due to their siliceous cell wall. Across the world, diatom communities are used in ecological status assessments of aquatic ecosystems due to their quick reactions to environmental changes and provide a wide range of information for diagnosing changes in aquatic ecosystems (Szczepocka and Wieczorek 2018; Abdelkarim 2020; Lobo et al. 2020; Trabert et al. 2020). Some diatom species have a well-known autecological indicator for pH, salinity (Van Dam et al. 1994), acidity, mineralization, oxygen requirements, saprobity, trophic state, organic nitrogen, and nitrates (Carayon et al. 2019). Therefore, diatoms have been used to evaluate environmental conditions in many countries as indicators of water pollution (Tokatlı et al. 2020).
Diatom index was developed based on autecological study and species pollution sensitivity. Nowadays, the diatom index is one of the most widely used water quality assessment techniques in Europe, such as Descy’s Index (DES, Descy 1979), Specific Pollution Sensitivity Index (IPS, Coste 1982), Commission for Economical Community Metric-European Index (CEE, Descy and Coste 1991), Generic Diatom Index (IDG, Coste and Ayphassorho 1991), Artois-Picardie Diatom Index (IDAP, Prygiel and Coste 1996), and Biological Diatom Index (IBD, Lenoir and Coste 1996) for general pollution. Diatom index for trophic status had been developed, such as Trophic Diatom Index (TDI, Kelly and Whitton 1995), Trophic-Saprobic index (T-SI, Lobo et al. 2004), and Eutrophication/Pollution Index-Diatom-based (EPID, Dell’uomo and Torrisi 2011). Furthermore, Metal Pollution Index (MPI, Fernandez et al. 2018), and the FGDI (French Guiana Diatomic Index; Carayon et al. 2020) had also been established. Diatom communities are in place of were also good bioindicators of soil condition and quality change related to human disturbances (Minaoui et al. 2021).
Diatom indices are one of the most widely used water quality assessment techniques and have been widely researched in many ecosystems and countries such as Gomez and Licursi 2001 (rivers and streams in Argentina), Kelly et al. 2008 (rivers in the UK), Trabert et al. 2017 (Danube, the second-longest river in Europe), Ali et al. 2018 (Tigris River, Iraq), Mangatze et al. 2019 (austral temperate river system, Bloukrans River system, South Africa), Wondmagegn et.al. 2019 (tropical rift valley lake, Lake Hawassa, in Ethiopia), Lobo et al. 2020 (Amazon floodplain, Brazil), Solak et al. 2020 (Sakarya River Basin, Turkey), Tokatly et al. 2020 (Ergene River Turkey), Yang et al. 2020 (dish lake from Nanjishan nature reserve, Lake Poyang, China), Gebler et al. 2021 (Polish lakes), Kennedy and Buckley 2021 (Irish lakes). Because of their sensitivity and rapid response to environmental change, diatom communities have been widely used in the world to provide robust assessments for ecosystem impairment and the biotic integrity of ecosystems (Mannion 2012).
Diatoms are considered successful bioindicators worldwide, different levels of water quality have been known to have a significant effect on diatom community composition and diatom index (Tokatly et al. 2020). Most of the diatom indices were developed and well-performed for river water quality in Europe. Where the diatom composition is similar and the water quality is comparable, it is possible to implement the European diatom indices in other regions, other countries, or even across continents (Carayon et al. 2019; Wondmagegn et al. 2019).
Those successful applications of diatoms as a bioindicator in the river water quality monitoring were followed by their use as environmental markers in the lake sediment. Paleoenvironmental analyses help to determine the past environmental conditions through the changes in preserved diatom assemblages that reflect the lake’s condition at the time of their deposition (Jensen et al. 2020). Critically, diatoms are known to be highly sensitive to water quality and the ecological preferences of a wide range of species are well documented from studies modeling species abundances across water quality gradients. Databases of the preferences of species can be used to infer changes to lake conditions over time based on the relative abundance of known species extracted from dated sediment sequences (Battarbee et al. 2011a; Bennion et al. 2014; North et al. 2018; Wondmagegn et al. 2019; Yang et al. 2020).
Since the 1980s, the development of mining and logging sharply increased in Halmahera Utara, causing conflict with local inhabitants and increased religious tension in Halmahera Utara Regency. Communal violence between Muslim and Christian communities broke out from December 1999 until 2001 (Duncan 2009). During the conflict, the people refuged to Galela and lived near the lake, and the communities are well-developed until now.
Recent research has been conducted in Galela Lake on the composition of phytoplankton to establish the effect of land usage on the water quality of local lakes and to examine how phytoplankton diversity could act as a tool for environmental assessment (Suhry et al. 2020a, 2020b). The water quality of Galela Lake in 2017 was in a good to moderate category. The phytoplankton community in Galela lake consists of 12 species of Chlorophytes, 9 species of Bacillariophytes, 4 species of Cyanophytes, 1 species of Euglenophyte, and Dinophyte, respectively. Aulacoseira granulata dominated the phytoplankton community in Galela lake. The Bacillariphyte dominated the phytoplankton community in the water column at 10–20 m and was replaced with Chlorophyte at the 0–5 m water column (Suhry et al. 2020b).
This study was conducted to analyze the temporal change of Galela Lake with the help of diatom indices from a 90-cm sediment core sample retrieved from the lake adjacent to residential areas influenced by human land-use activities and an 85-cm sediment core sample from the middle of Galela Lake.
Material and methods
Sampling area
There are six lakes in the Halmahera Utara Regency, North Maluku, namely Galela, Makete, Kapupu, Talaga Biru, Talaga Paca, and Talaga Lina lakes. These lakes play a crucial role in several ecosystem services, such as water supply, flood regulation, fisheries, and recreation. Galela Lake is the largest covering 390 hectares. It is surrounded by the villages of Seki, Togawa, Sukonora, Igobula, Bale, Soatabaru, Dokolamo, Duma, and Goatalamo. In recent decades, Galela Lake has transformed due to land-use changes as part of settlement development (Djangu et al. 2017). A classification of the land use of the catchment of the lake shows 73.34% to be farmland, 11.12%, forest, 4.87% resident area, and 10.67% lake. Almost one-quarter (22.1%) of the lake is covered with water hyacinth (Suhry et al. 2020a). An increase in the floating net cage area (FNCA) has not only affected the water quality but has also increased the organic content due to the decomposition of residual food and feces from the measured area of the lake.
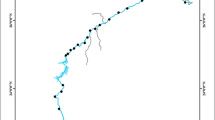
Galela Lake is situated at 1°49′6.05 S, 127°48′38.46 E. The 90 cm sediment core was collected in September 2019 from Site1, a location adjacent to the residential area in the Togawa village, and Site 2, a location in the middle of the lake (Fig. 1). The sediment sample was obtained using a piston corer with an internal diameter of 6.35 cm. Once extracted, the cores were kept in a vertical position to preserve the sediment layering. The sediment was extracted using a plastic spatula by cutting the pipe into 2 parts. Core sections were sliced every 5 cm and separated into subsamples that were placed in plastic zip-lock bags and refrigerated. Sub-sampling was carried out in the field due to the remote location (3,477 km round trip).

Research site at Galela Lake Halmahera Utara. Site 1 near residence area of Togawa Besi village, Site 2 in the midle of the lake
Diatom analysis
Diatom analysis was carried out according to Soeprobowati et al. (2016) which is a modified version of Battarbee (1986). One gram sediment sample was digested with 50 mL of 10% HCl for 2 h at 80 °C, settled for 6 h and the supernatant was discharged. Furthermore, the sample was digested with 50 mL of 10% H2O2 to remove the organic material. The samples were washed repeatedly and rinsed in distilled water between each stage until the pH was approximately neutral.
A 400 µL subsample of the clean slurry of diatoms frustules was dried onto a coverslip and permanent slides were mounted with Naphrax. A minimum of 300 diatoms valves were counted and identified under oil immersion at 1,000 × magnification. Diatom taxa were identified using standard literature (Kramer and Lange-Bertalot 1986, 1988, 1991, 2000, 2004, Gell et al. 1999, Sonneman et al. 2000, Taylor 2007a, 2016, Bahl 2017, Bahl et al. 2018), and by checking photomicrographs in AlgaBase.org (Guiry and Guiry 2021).
Data analysis
The Shannon–Wiener Index (H’) was calculated from the abundance of diatom species preserved in each sediment layer, as a measure of the α diversity. The calculation of indices of diversity and Evenness (E) was performed using PAST version 2.17c (Hammer 2017). To determine points of change between zones of similar diatom communities, the unweighted pair group method was applied with arithmetic mean (UPGMA) clustering analysis based on Bray–Curtis similarity measure using PAST. The vertical distribution of diatom species was presented using C2 software version 1.7.7 ((Juggins 2016).
The relative abundance of diatoms was used to calculate the diatom indices (Lecointe et al. 1993). Data were transformed to percent abundance and rare species (maximum abundance < 2%) were removed from the dataset before analysis, as they are not present in sufficient quantity to be statistically significant. The analysis of 18 diatom indices was performed using OMNIDIA software version 6.0 (Leconte et al. 1993) to determine relative water quality as shown in Table 1. The percentage (%) species for diatom indices and the diatom indices scores in corresponding ecological status were presented with heatmaps produced by Python version 3.7.13 with the platform of google colab. The library that had been used were numpy, pandas, matplotlib, and seaborn.
Results and discussion
The indices
In the sediment record from Site 1 of Galela Lake, the Shannon–Wiener index (H’) was generally higher for samples in the lower layers. The diversity indices tended to increase from 1.9 (90 cm), 2.07 (85 cm), and 2.52 (80 cm), respectively, and reached a peak at layer 75 cm. It then decreased slightly to 2.77 at layer 35 cm. Except for layers 30 and 20 cm, the upper layers (45–0 cm) has higher diversity compared to the lower layers (90–55 cm). In Site 2, the upper layers (30–0 cm) have higher diversity than the lower layers (85–35 cm, H’ less than 2, Fig. 2).

Shannon–Wiener diversity (H') index from Galela Lake
Layers 30, 75, and 80 cm of Site 1 have a high Evenness index. The Evenness indices at layers 90- 65 cm tend to reduce and slightly increase from layers 40 to 0 cm (Fig. 3). In layer 60–45 cm diatom were poorly preserved and only some small fragments were observed that could not be identified. Species diversity and Evenness indices differed significantly among layers tending to be higher relatively in the middle into bottom layer compared to the upper layer. The lack of diatom at 60—45 cm layers, might be related to the frustules undergoing dissolution in the water column at the sediment–water interface or in the pore waters of the sediments itself before conciliation (North et al. 2018). Alternatively, the diatom assemblage maybe is diluted from the increased sedimentation during high carbonate precipitation which may also have damaged the diatom valves. In contrast, Site 2 has lower Evenness indices in the upper layers (33–0 cm), high Evenness indices were found at layers 40 (0.79) and 50 cm (0.87, Fig. 3).

Evenness index from Galela Lake
Fossil groups and key species
A total of 49 diatom taxa belonging to 24 genera were recorded from the sampled core of Site 1 and 63 species were found from Site 2. At Site 1, the highest number of species were found at layer 70 cm (42 species). Layers 90–80 cm had 11–17 species and layers 30–0 cm has 5–34 species, with exceptions at layer 20 cm (12 species) and 30 cm (5 species), respectively. At Site 2, layers 30–0 cm have a high number of species (26–46), and layers 85–35 cm have several species in the range of 3–24. Layer 40 cm has 4 species and layer 50 cm has 3 species diatoms.
Based on the cluster analysis with Bray–Curtis similarity, it was possible to divide the samples from Site 1 into two groups with significantly different diatom compositions which are Zone 1 (90–65 cm) and Zone 2 (40–0 cm) as described in Fig. 4. The blank zone is the layers 60–45 cm. Site 2 was also divided into 2 clusters, Zone 1 (85–35 cm) and Zone 2 (30–0 cm, Fig. 5).

Vertical distribution of diatom in the Galela lake based on species diatom compositions > 2% abundance from Site 1

Vertical distribution of diatom in the Galela lake based on species diatom compositions > 2% abundance from Site 2
Fossil groups and key species
The cutoff value for separation of these clusters was set at < 2% similarity to reveal the main patterns in the diatom communities without providing excessive detail. Most taxa showed a particular preference for one group, except Encyonema minutum, Navicula leptostriata and Nitzchia gracilis which were present in most sediment layers.
Zone 1 of Site 1 (90–65 cm) consisted of subzone 1a (layers 90–80 cm) and subzone 1b (layers 75–65 cm). Subzone 1a was dominated by Achnanthidium minutissimum with maximum abundance in layers 90 cm, which was slightly reduced in the layers 85–80 cm (Fig. 4). Ulnaria acus was the dominant species at the 85 cm layer and declined into the top layers. In zone 1b, the diatom assemblage comprised a reduced abundance of Achnanthidium minutissimum and a greater representation of tolerant diatoms such as Encyonema minutum, Encyonema sileasiacum, Navicula spp, and Nitzschia spp. which entered the record in response to nutrient sources impacting the lake. This is likely due to the diversification in the land usage that has occurred over recent decades and other direct factors such as domestic discharges and agricultural runoff. This finding is consistent with that expected in nutrient-impacted lakes such as those in Europe (Kelly et al. 2014) and the UK (Bennion et al. 2014).
Subzone1b of Site 1 (layers 75–65 cm) was dominated by Encyonema minutum, Encyonema sileasiacum, Navicula leptostriata, Nitzchia gracilis, Nitzschia perminuta, and Ulnaria acus (Fig. 4). Layer 30 cm, was in contrast to others with only 5 species of diatoms recorded and was dominated by Platessa oblongella and Platessa lanceolata. Zone 2 (40–0 cm) was dominated by Encyonema minutum, Encyonema sileasiacum, Navicula leptostriata, Nitzschia gracilis, and Ulnaria acus that were reduced at the Zone 2. Pinularia viridis was dominant at layer 20 cm. It was uncommon but with a tendency, Achnanthidium minutissimum to increase in layers 15 cm and above. Achnanthidium minutissimum was used to indicate oligotrophy in lochs (Bennion et al. 2014), the most alkaline pre-acidification conditions in 121 low alkalinity lakes in the UK (Battarbee et al. 2011b). The Achnanthidium minutissimum complex is observed across various water status categories, but it is particularly abundant in high-status habitats (Stubbington et al. 2019). The presence of sensitive taxa such as Achnanthidium minutissimum reveals that the original condition of Galela Lake was of low nutrient levels.
A high dominance of Aulacoseira ambigua and Aulacoseira granulata since layer 50 cm from Site 2 recorded the eutrophication process, mostly in layers 50–35 cm (sub-zone 1b, Fig. 5). Encyonema minutum, Encyonema sileasiacum, Fragillaria tenera, Gomphonema parvulum, Navicula angusta, Navicula leptostriata, Navicula notha, Nitzchia gracilis, Nitzschia perminutta, and Ulnaria acus were dominant at Zone 2.
An increase of pollution-tolerant taxa such as Navicula and Nitzschia was associated with the formation of sedimentation (Peszek et al. 2021). A high assemblage of Nitzschia perminutta in zone 2 indicated a neutrophilic, β-mesosaprobous, mesotraphentic strictly aquatic form, (Van Dam et al. 1994) is further evidence for poor water quality (Table 2). Usually, Nitzschia perminutta originates from the rivers (Solak et al. 2020).
Diatom indices
Eighteen diatom indices were calculated with OMNIDIA software version 6.0, noting that species with abundance < 2% were not used for analysis. The diatom index scores of the 15 layers from Site 1 and Site 2 of Galela Lake sediment cores are shown in Tables 3 and 4. The greater the number of species from Galela Lake included in an index calculation the more efficient the resulting metric was at explaining the ecological status of the lake (Table 4).
The indices were found to have 5.6 to 100 percent (Site 1, Table 3) and 9.5 – 100 percent (Site 2, Table 4) representation found from Galela Lake cores that were available at OMNIDIA database. On average, for Site 1, the indices that included the highest species diversity, with more than 70% identified species, were IDG, IPS, and ACID (Table 3), whereas for Site 2 were IBD, IPS, IDG, TDI, and ACID (Table 4). IDG represented 100% of the genera recorded from the 2 cores. IBD, EPID, Pfister new Trophic indices (PTI), and TDI had more than 50% identified species from Site 1 (Table 3), and Sládeček’s Index (Sla), EPID, Swiss Diatom Index, Hurl (DI-CH), PTI, and Steinber and Schiefele trophic metric (SHE) had more than 50% identified diatoms species from Site 2 (Table 4).
Inferred change in condition
Based on the IBD, IPS, EPID, Rott’s Saprobic Metric (Rott SI), and Pfister’s new Saprobic indices (PSI), Zone 1b Site 1 was categorized as being in a high ecological status (oligotrophic), indicated by value > 15 (Table 5). Zone 1a tended to an oligotrophic state based on the IPS, was oligo-mesotrophic based on IBD, IDG, EPID, and PTI, and mesotrophic based on TDI indices. Based on the IBD and IPS Zone 2 was in the moderate to high ecological status (oligotrophic – mesotrophic), whereas based on IDG and TDI indices it was in poor – moderate condition (oligotrophic to meso-eutrophic) (Table 2). At Site 2, based on the Rott SI, PSI, and WAT, zone 1a (85–65 cm) was categorized as being in a high ecological status (oligotrophic), indicated by value > 15 (Tables 2 and 6). However, based on IBD, zone 1 reflects poor to moderate conditions. Based on the IBD and IPS, zone 2 (30–0 cm) was moderate to a high condition, whereas based on IDG in the moderate and good level (Tables 2 and 6).
All species found from Galela Lake were represented in IDG (100%) because the IDG index is limited to the genus level for simplification of the technique (Descy and Coste 1991). IDG was shown to perform well in monitoring water quality in tropical African rift lakes (Wondmagegn et.al. 2019), as well as in Turkey (Solak et al. 2020) and China (Yang et al. 2020). Based on IPS, IDG, and IBD, Galela Lake was experiencing oligo-mesotrophic to meso-eutrophic conditions. At zone 2, most species reflect circumneutral to slightly alkaline conditions yet the presence of Aulacoseira granulata at layers 15–0 cm reflected the ecological change from oligotrophic at zone 1 to meso-eutrophic at zone 2 and can therefore be beneficial as a paleoenvironmental indicator of catchment area changes.
Inferred from the diatoms indices, zone 1a Site 1 tended to be alkaline (pH 7.3–7.9) and oligotrophic. Zone 1b was circumneutral (pH 6.7–7.3) to alkaline, oligotrophic (orthophosphate < 0.11 mg/L) to oligo-mesotrophic, and oligonitrophic (Nitrate < 6.10 mg/L). Zone 2 tended to circumneutral to alkaline, mesotrophic to meso-eutrophic (orthophosphate 0.21–0.3 mg/L) conditions, organic nitrogen < 0.73–0.88 mg/L, and mesotrophic (10.63–13.91 mg/L). At Site 2 zone 2 was tent to alkaline (pH 7.56–8.53). This was indicated by the dominance of Aulacoseira ambigua and Aulacoseira granulata.
The ecological status of aquatic ecosystem based on the diatom indices is well developed and has performed well in Europe, mostly for determining the ecological status of rivers (Elorana and Soininen 2002, Taylor et al. 2007b, Kelly et al. 2008, Wang et al. 2014, Trabert et al. 2017, Pham and Nguyen, 2018, Xue et al. 2019, Cantonati et al. 2020, Solak êt al. 2020, Tokatly et al. 2020, Jakovljević et al. 2021). The diatom indices were well performed for assessing the ecological status of lakes (Battarbee 2000, Sayer and Robert 2001, Hall 2010, Battarbee et al. 2011a, 2011b, Bennion et al. 2014, Kelly et al. 2014, North et al. 2018, Wondmagegn et al. 2019, Gell 2019, Yang et al. 2020).
The data obtained from this research demonstrated that European indices can be applied to tropical lakes, specifically in this instance to Galela Lake. Of the 18 indices applied, seven diatom indices were noted as a potential metric for application to Galela Lake. On average for all layers from Site 1, the IDG, IPS, ACID, TDI, IBD, EPID, and PTI indices worked with 100%, 82.7%, 70.41%, 63.42%, 59.81%, 51.66%, 50.71%, respectively, concerning the identified taxa in the basin (Table 3). These indices represent fossil records, mostly cosmopolitan species for altitude. The indices that are not strong indicators for Galela Lake (by supported 50% or less of diatom) are Descy, IDS/E, IDAP, Lobo, DI-CH, Rott SI, PTI, PSI, TDIL, CEE, WAT, PDI, and EPI-L, which are not applicable for tropical lakes, consist of tropical specific or endemic species. The average of species form Site 2 for all layers were for the indices of IDG (100%), IPS (89.93%),ACID (84.49%), TDI (83.49%), IBD (78.47%), EPID (57.66%), SHE (57.32%) PTI (56.43%), DI-CH (52.6%), and Sla (51.77%), respectively (Table 4).
IPS and IDG indices were the best-performed indices of the trophic state of Galela Lake. This result supported the finding that diatom indices could be applied outside their original geographic region, provided climatic conditions in lowland Europe has a similar climate to upland Indonesia – cool. Possible warm lowland Indonesian waters would have different species. The data obtained from this research highlighted that many indices developed in Europe can be applied to tropical settings such as Galela Lake. This is because a high proportion of the diatom species recorded as a fossil in the Galela Lake sediments are represented in the databases from which the indices were developed. This high level of commonality between temperate European flora, and tropical Indonesian flora, may be on account of the cooler climates.
On the other hand, care must be taken as diatom indices developed in one geographic area were applied less successfully in other areas and this may cause uncertain results (Taylor et al. 2007b), hardly define ecological profiles for some taxa, several taxa have the same ecological preference or different ecological profiles for the same species (Pinto et al. 2021). This arose on account of the high number of endemic species in the test tropical lake so few were common to the species in the OMNIDIA database. This may likely be resolved by the establishment of a tropical lake diatom database that would better capture the biogeography and limnology of Indonesian lakes.
Usually, diatom indices are used as indicators of varying waterway conditions across spatial regions. The novelty of this research was applying diatom indices to reveal the temporal change from cores that was retrieved from a site impacted by human activities compare to the site from the middle of the lake.
So, diatom indices are a potential tool to elucidate the possible impact of human activity differences on the lake ecosystem. Diatom indices that were developed for temperate regions, when applied in a tropical lake, delivered satisfactory results. The usefulness index reveals whether there is a high level of commonality of species in the index, and the species in the sediment core, i.e., there are good analogs in the data sets. Species’ preferences for nutrients and temperature may differ in lowland tropical lakes. The development of a data set of diatoms from warm, tropical waters would be valuable to fill this gap.
Conclusion
This study assessed the suitability of diatom indices for water quality produced in temperate Europe to the tropical setting of Galela Lake. The diatom indices were tested against the diatom assemblages extracted from 90 and 85 cm sediment cores from the lake in a location likely to be affected by nearly land development. Several indices performed well owing to the high level of commonality between the species in the database and those identified from the fossil material. This is likely due to the relatively cool climate at Galela Lake. Likely that these European indices would not perform as well in lowland Indonesian waters due to the warmer conditions. The development of local index systems for warm tropical waters would better test the suitability of temperate-based diatom indices, and pave the way for the better application of these approaches in tropical situations.
Data availability statements
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Abdelkarim MS (2020) Biomonitoring and bioassessment of running water quality in developing countries: a case study from Egypt. Egypt J Aquat Res 46(4):313–324. https://doi.org/10.1016/j.ejar.2020.11.003
Ali SF, Abdul-Jabar RA, Hassan FM (2018) Diversity measurement indices of diatom communities in the tigris river within wasit province. Iraq. Baghdad Sci J. 15(2):117–122. https://doi.org/10.21123/bsj.2018.15.2.0117
Bahls L, Boynton B, Johnston B (2018) Atlas of diatoms (Bacillariophyta) from diverse habitats in remote regions of western Canada. PhytoKeys 105:1–186. https://doi.org/10.3897/phytokeys.105.23806
Bahls LL (2017). Diatoms from Western North America 1. Some new and notable Biraphid species Montana diatom collection; Lubrecht and cramer. Helena, MT, USA. pp. 1–52
Battarbee RW (2000) Palaeolimnological approaches to climate change, with special regard to the biological record. Quat Sci Rev 19(1–5):107–124. https://doi.org/10.1016/S0277-3791(99)00057-8
Battarbee RW, Morley D, Bennion H, Simpson GL, Hughes M, Bauere V (2011a) A palaeolimnological meta-database for assessing the ecological status of lakes. J Paleolimnol 45:405–414. https://doi.org/10.1007/s10933-010-9417-5
Battarbee RW, Simpson GL, Bennion H, Curtis C (2011b) A reference typology of low alkalinity lakes in the UK based on pre-acidification diatom assemblages from lake sediment cores. J Paleolimnol 45:489–505. https://doi.org/10.1007/s10933-010-9426-4
Battarbee RW (1986) Diatom analysis in Berglund, B.E. (ed), Handbook of Holocene Paleoecology and Paleohydrology. Wiley, Chichester. pp. 527–570. Doi: https://doi.org/10.1002/jqs.3390010111
Bennion H, Kelly MG, Juggins S, Yallop ML, Burgess A, Jamieson J, Krokowski J (2014) Assessment of ecological status in UK lakes using benthic diatoms. Freshw Sci 33:639–654. https://doi.org/10.1086/675447
Cantonati M, Kelly MG, Demartini D, Angeli N, Doringer G, Papatheodoulou A, Armanini DG (2020) Overwhelming role of hydrology-related variables and river types in driving diatom species distribution and community assemblage in streams in Cyprus. Ecol Indic 117:106690. https://doi.org/10.1016/j.ecolind.2020.106690
Carayon D, Tison-Rosebery J, Delmas F (2019) Defining a new autoecological trait matrix for French stream benthic diatoms. Ecol Indic 103:650–658. https://doi.org/10.1016/j.ecolind.2019.03.055
Carayon D, Eulin-Garrigue A, Vigouroux R, Delmas F (2020) A new multimetric index for the evaluation of water ecological quality of French Guiana streams based on benthic diatoms. Ecol Indic 113:106248. https://doi.org/10.1016/j.ecolind.2020.106248
Coste M, Ayphassorho H (1991) Etude de la qualité des eaux du Bassin Artois-Piccardie á l’ aide des communautés des diatomées benthiques (application des indices diatomiques). In: Rapport CEMAGREF, Bordeaux; Agence de l’Eau Artois-Picardie: Douai France, pp. 1–227
Coste M (1982) Etude des méthods biologiques quantatatives d’appréciation de la qualité des eaux. Rapport Division Qualité des Eaux Lyon. Agence Financiére de Bassin Rhône. Mediterranée-Corse: Lyon French, pp. 1–218
Dell’uomo A (1996) Assessment of water quality of an Apennine river as a pilot study for diatom-based monitoring of Italian watercourses. In: The Use of Algae for Monitoring Rivers II. Whitton BA, Rott E, (Eds.), Institut fϋr Botanik Universität Innsbruck, Innsbruck Austria, pp. 1–65
Dell’uomo A, Torrisi M (2011) The eutrophication/pollution index-diatom based (EPI-D) and three new related indices for monitoring rivers: the case study of the river Potenza (the Marches, Italy). Plant Biosystems 145(2):331–341. https://doi.org/10.1080/11263504.2011.569347
Descy JP (1979) A new approach to water quality estimation using diatoms. Nova Hedwig 64:305–323
Descy JP, Coste M (1991) A test of methods for assessing water quality based on diatoms. Verh Intern Limnol 24(4):2112–2216. https://doi.org/10.1080/03680770.1989.11899905
Djangu A, Rondonuwu DM, Sela, RLE (2017) Analisis Perubahan Penggunaan Lahan di Kawasan Tepi Danau Galela Kabupaten Halmahera Utara. Spatial 4 3: 96–108. https://ejournal.unsrat.ac.id/index.php/spasial/article/view/17331/16873 (in Bahasa Indonesia)
Duncan C (2009) Monuments and martyrdom: memorializing the dead in post-conflict North Maluku. Bijdragen Tot De Taal, Land- En Volkenkunde, 165 4: 429–458. Retrieved July 4, 2021, from http://www.jstor.org/stable/43817773
Eloranta P, Soininen J (2002) Ecological status of some Finnish rivers evaluated using benthic diatom communities. J Appl Phycol 14(1):1–7. https://doi.org/10.1023/A:1015275723489
Fernández MR, Martín G, Corzo J, de la Linde A, García E, López M, Sousa M (2018) Design and testing of a new diatom-based index for heavy metal pollution. Arch Environ Contam Toxicol 74(1):170–192. https://doi.org/10.1007/s00244-017-0409-6
Gebler D, Kolada A, Pasztaleniec A, Szoszkiewicz K (2021) Modelling of ecological status of Polish lakes using deep learning techniques. Environ Sci Pollut Res 28:5383–5397. https://doi.org/10.1007/s11356-020-10731-1
Gell PA (2019) Watching the tide roll away – contested interpretations of the nature of the Lower Lakes of the Murray Darling Basin. Pac Conserv Biol 26:130–141. https://doi.org/10.1071/PC18085
Gell P, Sonneman J, Reid K, Illman MA, Sincock AJ (1999) An illustrated key to common diatom genera from southern Australia. Co-operative research centre for freshwater ecology: Hurgoona, Australia, Thurgoona, NSW. pp.1–68
Gomez N, Licursi M (2001) The Pampean Diatom Index (IDP) for assessment of rivers and streams in Argentina. Aquat Ecol 35:173–181. https://doi.org/10.1023/A:1011415209445
Guiry MD, Guiry GM (2021) AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https:www.algaebase.org (Accessed on 4 July 2021)
Hall R, Smol J (2010) Diatoms as indicators of lake eutrophication. In: Smol J, Stoermer E (eds) The diatoms: applications for the environmental and earth sciences. Cambridge University Press, Cambridge, pp 122–151. https://doi.org/10.1017/CBO9780511763175.008
Hammer O (2017) PAST: Paleontological Statistics Software Package for Education and Data Analysis. Version 2.17c. Available online: http://priede.bf.lu.lv/ftp/pub/TIS/datu_analiize/PAST/2.17c/download.html (Accessed on 6 July 2021).
Hürlimann J, Niederhauser P (2002) Méthode d’analyse et d’appréciation des cours d’eau en Suisse, Diatomées, niveau R région. Office Fédéral de l’Environnement, des Forêts et du Paysage OFEFP: Berne, Switzerland, pp. 1–111
Jakovljević OS, Popović SS, Živić IM, Stojanović KZ, Vidaković DP, Naunovic ZZ, Krizmanić JZ (2021) Epilithic diatoms in environmental bioindication and trout farm’s effects on ecological quality assessment of rivers. Ecol Indic 128:107847. https://doi.org/10.1016/j.ecolind.2021.107847
Jensen TC, Zawiska I, Oksman M, Slowinski M, Woszczyk M, Luoto TP, Tylmann W, Nevalainen L, Obremska M, Schartau AK, Walseng B (2020) Historical human impact on productivity and biodiversity in a subalpine oligotrophic lake in Scandinavia. J Paleolimnol 63:1–20. https://doi.org/10.1007/s10933-019-00100-5
Juggins S (2016) C2 Software for ecological and palaeoecological data analysis and visualization, Version 1.7.7; University of New Castle: New Castle, UK. Available online: https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm (Accessed on 8 July 2021)
Kelly MG, Whitton BA (1995) The trophic diatom index: A new index for monitoring eutrophication in rivers. J Appl Phycol 7:433–444. https://doi.org/10.1007/BF00003802
Kelly M, Juggins S, Guthrie R, Pritchard S, Jamieson J, Rippey B, Hirst H, Yallop M (2008) Assessment of ecological status in U.K. rivers using diatoms. Freshw Biol 53:403–422. https://doi.org/10.1111/j.1365-2427.2007.01903.x
Kelly M, Urbanic G, Ács É, Bennion H, Bertrin V, Bur-gess A, Denys L, Gottschalk S, Kahlert M, Karjalainen SM, Kennedy B, Kosi G, Marchetto A, Morin S, Picinska-Fałtynowicz J, Poikane S, Rosebery J, Schoenfelder I, Schoenfelder J, Varbiro G (2014) Comparing aspirations: intercalibration of ecological status concepts across European lakes for littoral diatoms. Hydrobiologia 734(1):125–141. https://doi.org/10.1007/s10750-014-1874-9
Kelly M, King L, Ní Chatháin B (2009) The conceptual basis of ecological status assessments using diatoms. Biol. Environ. Proc. R. Ir. Acad.109, 3: 175–189. DOI: https://doi.org/10.3318/BIOE.2009.109.3.179. https://www.jstor.org/stable/20694892?seq=1
Kennedy B, Buckley YM (2021) Use of seasonal epilithic diatom assemblages to evaluate ecological status in Irish lakes. Ecol Indic 129:107853. https://doi.org/10.1016/j.ecolind.2021.107853
Kociolek JP, You Q, Liu Q, Liu Y, Wang Q (2020) Continental diatom biodiversity discovery and description in China: 1848 through 2019. PhytoKeys 160:45–97. https://doi.org/10.3897/phytokeys.160.54193
Krammer K, Lange-Bertalot H (1986) Bacillariophyceae 1. Teil: Naviculaceae. In Süsswasserflora von Mitteleuropa, Band 2/1; Ettl, H., Gärtner, G., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, pp. 1–876. (in German)
Krammer K, Lange-Bertalot H (1988) Bacillariophyceae 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Süsswasserflora von Mitteleuropa, Band 2/2; Ettl, H., Gärtner, G., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, pp. 1–596. (in German)
Krammer K, Lange-Bertalot H (1991) Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In Süsswasserflora von Mitteleropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, pp. 1–576. (in German)
Krammer K, Lange-Bertalot H (2000) Bacillariophyceae 5. Teil: Bacillariophyceae. In Süsswasserflora von MitteleuropaII, Springer Spektrum Verlag, Jena, Germany. Volume 2, pp. 1–311. (in German)
Krammer K, Lange-Bertalot H (2004) Bacillariophyceae 4. Teil: Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae), Gomphonema Gesamtliteraturverzeichnis Teil 1-4 [second revised edition] [With “Ergänzungen und Revisionen” by H. Lange-Bertalot]. In Suesswasserflora von Mitteleuropa; Ettl, H., Ed.; Spektrum Akademischer Verlag: Heidelberg, Germany, Volume 2, pp. 1–468. (in German)
Leclerq L, Maquet B (1987) Deux nouveaux índices chimique et diatomique de qualité d’eau courante: Comparaison avec diérents indices existants. Cah Biol Mar 28:303–310 ((in French with English abstract))
Lecointe C, Coste M, Prygiel J (1993) “Omnidia”: software for taxonomy, calculation of diatom indices and inventories management. Hydrobiologia 269(1):509–513. https://doi.org/10.1007/BF00028048
Lenoir A, Coste M (1996) Development of a practical diatom index of overall water quality applicable to the French National Water Board network. In the Use of Algae for Monitoring Rivers II. Whitton BA, Rott E, (Eds) Institut fϋr Botanik Universität Innsbruck, Schmitten, Austria, pp. 29–45
Lobo EA, Callegaro VLM, Hermany G, Bes D, Wetzel CA, Oliveira MA (2004) Use of epilithic diatoms as bioindicator from lotic systems in southern Brazil, with special emphasis on eutrophication. Acta Limnol Bras 16(1):25–40
Lobo MTMPS, Scalize PS, Kraus CN, da Silva WJ, Garnier J, da Motta MD, de Souza NI (2020) Biological index based on epiphytic diatom assemblages is more restrictive than the physicochemical index in water assessment on an Amazon floodplain. Brazil Environ Sci Pollut Res 27:10642–10657. https://doi.org/10.1007/s11356-020-07658-y
Mangadze T, Taylor JC, Froneman WP, Dalu T (2019) Water quality assessment in a small austral temperate river system (Bloukrans River system, South Africa): Application of multivariate analysis and diatom indices. S Afr J Bot 125:353–359. https://doi.org/10.1016/j.sajb.2019.08.008
Mannion AM (2012) The diatoms: applications for the environmental and earth sciences. J Biol Educ 46(3):200–201. https://doi.org/10.1080/00219266.2011.645850
Minaoui F, Hakkoum Z, Douma M, Mouhri K, Loudiki M (2021) Diatom communities as bioindicators of human disturbances on suburban soil quality in arid marrakesh area (Morocco). Water Air Soil Pollut 232:146. https://doi.org/10.1007/s11270-021-05094-3
North SM, Stockhecke M, Tomonaga Y, Mackay AW (2018) Analysis of a fragmentary diatom record from Lake Van (Turkey) reveals substantial lake-level variability during previous interglacials MIS7 and MIS5e. J Paleolimnol 59(1):119–133. https://doi.org/10.1007/s10933-017-9973-z
Peszek L, Zgrundo A, Noga T, Kochman-Kędziora N, Poradowska A, Rybak M, Puchalski C, Lee J (2021) The influence of drought on diatom assemblages in a temperate climate zone: a case study from the Carpathian Mountains, Poland. Ecol Indic 125:107579. https://doi.org/10.1016/j.ecolind.2021.107579
Pham TL, Nguyen TD (2018) Use of benthic diatom indices for assessing ecological status of the Saigon River. Vietnam Can Tho Univ J Sci 54(2):106–111. https://doi.org/10.22144/ctu.jen.2018.014
Pinto R, Vilarinho R, Carvalho AP, Moreira JA, Guimarães L, Oliva-Teles L (2021) Raman spectroscopy applied to diatoms (microalgae, Bacillariophyta): prospective use in the environmental diagnosis of freshwater ecosystems. Wat Res 198:117102. https://doi.org/10.1016/j.watres.2021.117102
Prygiel J, Coste M (1996) Recent trends in monitoring French rivers using algae, especially diatoms. In: Whitton BA, Rott E (eds) Use of Algae for Monitoring Rivers II. Institut für Botanik Universität Innsbruck, Schmitten, Austria, pp 1–87
Rott E, Hofmann G, Pall K, Pfister P, Pipp E (1997) Indikationslisten für Aufwuchsalgen in Österreichischen Fliessgewässern, Teil 1: Saprobielle Indikation Wasserwirtschaftskataster; Bundeministerium für Land- und Forstwirtschaft: Wienna, Astralia, pp. 1–73. (in German)
Rott E, Pfister P, Van Dam H, Pipp E, Pall K, Binder N, Ortler K (1999) Indikationslisten fϋr Aufwuchsalgen Teil 2: Trophieindikation und autökologische Anmerkungen Bundesministerium für Land- und Forstwirtschaf, Wienna, Astralia, pp1–248. (in German)
Sayer CD, Roberts N (2001) Establishing realistic restoration targets for nutrient-enriched shallow lakes: linking diatom ecology and palaeoecology at the Attenborough Ponds, U.K. Hydrobiologia 448:117–142. https://doi.org/10.1023/A:1017597221052
Schiefele S, Schreiner C (1999) Use of diatoms for monitoring nutrient enrichment acidification and impact salts in Germany and Austria. In: Whitton BA, Rott E, Friedrich G (eds) Use of Algae for Monitoring Rivers. Institut fϋr Botanik Universität Innsbruck, Schmitten, Austria, pp 1–193
Sládeček V (1986) Diatoms as indicators of organic pollution. Acta Hydrochim Hydrobiol 14:555. https://doi.org/10.1002/aheh.19860140519
Soeprobobowati TR, Tandjung SD, Sutikno S, Hadisusanto S, Suedy SWA, Gell P (2016) The minimum number of valves for diatom identification in Rawapening Lake Central Jawa. Biotropia 23(2):96–104. https://doi.org/10.11598/btb.2016.23.2.486
Solak CN, Peszek Ł, Yilmaz E, Ergül HA, Kayal M, Ekmekçi F, Varbiro G, Yüce AM, Canli O, Binici MS, Acs É (2020) Use of diatoms in monitoring the Sakarya River Basin. Turkey Water 12:703. https://doi.org/10.3390/w12030703
Sonneman JA, Sincock A, Fluin J, Reid M, Newall P, Tibby J, Gell P, Tyler P (2000) An illustrated guide to common stream diatom species from temperate Australia, Co-operative research centre for freshwater ecology: Hurgoona, Australia, pp. 1–166
Steinberg C, Schiefele S (1988) Biological indication of trophy and pollution of running waters. Zeitschrift Für Wasser Und Abwasser Forschung 21(6):277
Stubbington R, Paille A, England J, Barth`es A, Bouchez A, Rimet F, Sa ́nchez- Montoya MM, Westwood CG, Datry T (2019) A comparison of biotic groups as dry-phase indicators of ecological quality in intermittent rivers and ephemeral streams. Ecol Indic 97:165–174. https://doi.org/10.1016/j.ecolind.2018.09.061
Suhry HC, Soeprobowati TR, Jumari J (2020a) Land use change around Galela lake (Perubahan Tata Guna Lahan Sekitar Danau Galela. J Ilmu Lingkungan 18(1):140–145. https://doi.org/10.14710/jil.18.1.140-145
Suhry HC, Soeprobowati TR, Jumari J (2020b) Water quality and pollution indix in Galela lake (Kualitas air dan Indeks Pencemaran Danau Galela). J Ilmu Lingkungan 18(2):236–241. https://doi.org/10.14710/jil.18.2.236-241
Szczepocka E, Wieczorek JZ (2018) Diatom biomonitoring–scientific foundations, commonly discussed issues and frequently made errors. Oceanol Hydrobiol Stud. https://doi.org/10.1515/ohs-2018-0030
Taylor JC, Harding WR, Archibald CGM (2007a) An illustrated guide to some common diatom species from South Africa. Water Research Commission, Pretoria, South Africa, pp 1–225
Taylor JC, Van Vuuren MS, Pieterse AJ (2007b) The application and testing of diatom-based indices in the Vaal and Wilge Rivers, South Africa. Water SA 33:51–59. https://doi.org/10.4314/wsa.v33i1.47871
Taylor JC, Cocquyt C, Mayama S (2016) Navicula nielsfogedii sp. nov: A new diatom (Bacillariophyta) from tropical and sub–tropical Africa. Fottea 16:201–208. https://doi.org/10.5507/fot.2016.015
Tokatli C, Solak CN, Yilmaz E (2020) Water quality assessment by means of bioindication: A case study of Ergene river using biological diatom index. Aquat. Sci. Eng. 36: 43–51. Doi: https://doi.org/10.26650/ASE2020646725
Trabert Z, Kiss KT, Varbiro G, Dobosy P, Grigorszky I, Acs E (2017) Comparison of the utility of a frequently used diatom index (IPS) and the diatom ecological guilds in the ecological status assessment of large rivers. Fundam Appl Limnol 189(2):87–103. https://doi.org/10.1127/fal/2016/0933
Trabert Z, Duleba M, Biro T, Dobosy P, Foldi A, Hidas A, Kiss KT, Ovari M, Takacs A, Varbiro G, Zaray G, Acs E (2020) Effect of land use on the benthic diatom community of the Danube river in the region of Budapest. Water 12(2):479. https://doi.org/10.3390/w12020479
Van Dam H, Mertens A, Sinkeldam J (1994) A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Neth J Aquat Ecol 28(1):117–133. https://doi.org/10.1007/BF02334251
Wang H, Zheng B, Liu L, Li L (2014) Use of diatoms in river health assessment. Annu Res Rev Biol 4(24):4054–4074. https://doi.org/10.9734/ARRB/2014/11963
Watanabe T, Asai K, Houki A (1986) Numerical estimation to organic pollution of flowing water by using the epilithic diatom assemblage - diatom assemblage index (DAIpo). Sci Total Environ 55:209–218. https://doi.org/10.1016/0048-9697(86)90180-4
Wondmagegn T, Mengistou S, Barker PA (2019) Testing of the applicability of European diatom indices in the tropical rift valley lake, Lake Hawassa, in Ethiopia. Afri J Aquat Sci 44(3):209. https://doi.org/10.2989/16085914.2019.1645640
Xue H, Zheng B, Meng F, Wang Y, Zhang L, Cheng P (2019) Assessment of aquatic ecosystem health of the Wutong river based on Benthic diatoms. Water 11:727. https://doi.org/10.3390/w11040727
Yang J, Ji Y, Yan R, Liu X, Zhang J, Wu N, Wang K (2020) Applicability of benthic diatom indices combined with water quality valuation for dish lake from Nanjishan nature reserve Lake Poyang. Water 12:2732. https://doi.org/10.3390/w12102732
Acknowledgements
Thanks to Widodo, Hendro Suhry, and Suyono Lateke who helped during field work, and Kenanga Sari who help in the laboratory work.
Funding
The research leading to these results received funding from the Universitas Diponegoro Research Grant of RPIBT (Riset Publikasi Internasional Bereputasi Tinggi = Research for the High Reputable International Publications) under Grant Agreement No. 329–118/UN7.6.1/PP/2021.
Author information
Authors and Affiliations
Contributions
Tri Retnaningsih Soeprobowati: Conceptualization, Methodology, Investigation, Analysis, Writing–original draft, Writing–review & editing, and Funding Acquisition. Jumari: Conceptualization, Methodology, Investigation, Analysis, Writing–review & editing, and Funding Acquisition. Tyas Rini Saraswati: Conceptualization, Investigation, Writing–review & editing, and Funding Acquisition. Kenanga Sari: Methodology, Analysis, and Writing–review & editing. Peter Gell: Conceptualization, fieldwork Methodology, and Writing–review & editing.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The authors declare that they have no Ethical Approval.
Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Additional information
Editorial responsibility: Fatih ŞEN.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Soeprobowati, T.R., Saraswati, T.R., Jumari, J. et al. Diatom index of Galela Lake, Halmahera, Indonesia in relation to human activities. Int. J. Environ. Sci. Technol. 20, 7707–7722 (2023). https://doi.org/10.1007/s13762-022-04463-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13762-022-04463-7