
Abstract
Exopolysaccharides (EPS) are polymeric substances composed of sugars and are produced by microbes to form colonies and inhibit the growth of other organisms. In the present study, the bacterial strain Bacillus sp. YP03 was isolated from soil sample using a high-sucrose selective medium, the produced EPS recovered using ethanol precipitation, and lyophilized. In sugar analysis, it was found to be rich in fructose content, amounting to 86.1%, and further displayed appreciable antioxidant, emulsification, and flocculation activities. EPS concentration of 140 µg/ml, pH of 7.5, and the presence of trivalent metal cations augmented its flocculating ability. When applied as a coagulant aid in the treatment of municipal wastewater, up to 47 and 89% reductions of chemical oxygen demand and total suspended solids, respectively, were witnessed. Owing to these encouraging results, Bacillus sp. EPS could be applied as a potentially beneficial flocculant in wastewater treatment, for achieving accentuated TSS and organic load removal during primary sedimentation.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Microbes are capable of producing diverse biopolymers by utilizing simple to complex substrates. These biopolymers are classified into four major types, namely polyesters, polyamides, polyanhydrides, and polysaccharides. Polymeric substances composed of sugar moieties are called polysaccharides. There are many disadvantages in the production of polysaccharides from plant and algal sources. Change in environmental conditions like seasonal changes, aging cycle, and time of harvesting might influence the polysaccharide composition. Moreover, since plants contain cell walls, the isolation of polysaccharide involves harsh methods such as alkali treatment, acid precipitation, and bleaching, which may alter its structure and stability. Such difficulties can be overcome by the bacterial production of polysaccharides. Since the polysaccharides are produced extracellularly, their isolation becomes easier in bacteria. The enzyme that helps in the production of exopolysaccharides (EPS) is levansucrase. It cleaves the sucrose into glucose and fructose moieties and joins the fructose units to the growing fructan chain (Poli et al. 2009).
There are three main types of microbial polysaccharides: intracellular, structural, and extracellular. The intracellular polysaccharides serve as carbon reserve and help the cell during starvation. Structural polysaccharides are involved in the formation of cell structures. Extracellular polysaccharides are those produced and released by the microbes in their extracellular regions and are hence called EPS. Several bacteria and fungi are known to produce EPS (Qurashi and Sabri 2012; Domingos-Lopes et al. 2017; Indira et al. 2016; Jeong et al. 2017; Li et al. 2017). They prevent the growth of other competing microbes and also help in the formation of biofilms. EPS are commonly produced by deep sea bacteria to withstand high salinity and pressure. They may also act as a cryopreservant. Some may act as capsule for the microbe or completely detach from the bacterial surface and exist in the environment as slimes. Since EPS are non-toxic and biocompatible, they have innumerable applications in various fields such as medicine, cosmetics, pharmaceuticals, and dairy industries. They also function as insecticides and herbicides, anti-thrombotic and anticancer agents, and as bio-flocculants (and Patel and Prajapat 2013; Al-Wasify et al. 2015).
Pullulan, xanthan gum, hyaluronic acid, dextran, gellan, alginate, cellulose, succinoglycan, glucan, and levan are important EPS produced by microbes. Of these, levan is composed of fructose units and has low viscosity, high water solubility, adhesive strength, film-forming capacity, anti-tumor, and anti-inflammatory activities. Alcaligenes viscosus, Zymomonas mobilis, and Bacillus subtilis are bacteria that have been shown to produce levan. It finds application in food (as a prebiotic), feed, pharmaceutical, and cosmetic industries (Rehm 2010; Freitas et al. 2011).
In the present study, EPS-producing bacterial strains were isolated from soil sample. The compound was partially purified, and its carbohydrate, protein, and fructose contents were analyzed. The antioxidant, emulsifying, and flocculating activities of the EPS were tested. Further, its application potential as a natural flocculant in wastewater treatment was determined. The entire work was carried out in the Department of Biotechnology, Kumaraguru College of Technology, India, from June 2015 to April 2016.
Materials and methods
Chemicals
Activated carbon and bovine serum albumin (BSA) were purchased from Sigma-Aldrich, USA. Microbiological media components and all other chemicals were of reagent grade and procured from Hi-Media, India.
Isolation and identification of bacterial strains
Garden soil samples were collected from three different locations within the premises of Kumaraguru College of Technology, Coimbatore, India, in the month of June 2015. To screen for EPS production, the sample was serially diluted and 0.1 ml aliquots were spread on the surface of agar plates containing high-sucrose selective medium. It contained (per l) sucrose—250 g, peptone—2 g, yeast extract—2 g, (NH4)2SO4—2 g, K2HPO4—2 g, MgSO4—0.3 g, agar—15 g. After 24-h incubation, discrete bacterial colonies were subcultured and maintained as pure cultures in agar slants. They were subjected to Gram staining and MALDI–TOF Mass Spectroscopy for culture identification. The identity of the best EPS-producing isolate was further established by 16S rRNA gene sequencing. 16S rRNA gene primers 27F (5′-AGAGTTTGATC(A/C)TGGCTCAG-3′) and 1492R (5′-ACGG(C/T)TACCTTGTTACGACTT-3′) were used to amplify ~1.4 Kb gene from the isolated genomic DNA (Giovanoni 1991). The amplified PCR product was gel-purified, quantified, and sequenced using an ABI Prism 3100 Genetic Analyzer (Applied Biosystem). Consensus sequence was generated from forward and reverse sequences using ‘Aligner’ software. NCBI ‘BLAST’ was used to perform sequence similarity search in the GenBank database (Altschul et al. 1997). Phylogenetic tree was constructed using the neighbor joining method (Kimura 1980).
Production and partial purification of the bacterial EPS
For EPS production, the preserved stock cultures of the bacterial isolates were inoculated in the high-sucrose selective medium and cultured overnight. Subsequently, 2% (v/v) of this inoculum (53 × 106 CFU/ml) was transferred to the production medium and incubated at 37 °C and 110 rpm for 48 h. The cultures were centrifuged at 10,000 g for 10 min. To the supernatant, equal volume of ethanol was added, stored at −20 °C overnight, and again centrifuged for 30 min (Sam et al. 2011). The resulting pellet was dissolved in hot distilled water and dialyzed against distilled water using a 10-kDa membrane for 3 days at 4 °C. The sample was then lyophilized.
Analyses of total carbohydrate, protein, and fructose contents
Total carbohydrate content was analyzed by the phenol sulfuric acid method (Dubois et al. 1956). For this purpose, glucose standards (20–100 µg/ml) were prepared. One milliliter of phenol and 5 ml of 96% H2SO4 were added to each test tube and shaken well. After 10 min, they were placed in a water bath at 25 °C for 20 min. The intensity of the color developed was read at 490 nm against a reagent blank, and standard graph was plotted. The sample was suitably diluted, similarly processed and its carbohydrate content estimated from the standard graph.
Sucrose was used as the sole carbon source for the growth of bacteria, and the produced EPS was expected to be of poly-fructose moiety. Fructose content of the hydrolyzed EPS sample was estimated by resorcinol method (Roe 1934). Fructose standards were prepared in the concentration range of 20–100 µg/ml. One milliliter of resorcinol reagent and 7 ml of dilute HCl were added, and the tubes incubated at 80 °C for 10 min. They were cooled and optical density (OD) measured at 520 nm against a reagent blank. The sample was also similarly prepared and its fructose concentration estimated from the standard graph.
The bacteria not only produce EPS but also some extracellular proteins. The total protein content was estimated as per the protocol of Bradford (1976). To a series of test tubes containing BSA standards (20–100 µg/ml) and suitably diluted sample, 5 ml of Coomassie Brilliant Blue G 250 was added. The contents were mixed well and left for 5 min. The OD reading was measured at 595 nm against a reagent blank and protein content of sample estimated from the standard graph.
Antioxidant activity: DPPH, ABTS, and FRAP assays
The antioxidant activity of the exopolysaccharide was estimated by various assays. DPPH (2,2-diphenyl-1-picryl-hydrazyl-hydrate) free radical method is an antioxidant assay based on electron transfer that produces a violet solution in ethanol. This free radical is reduced in the presence of an antioxidant molecule, giving rise to colorless ethanol solution. The use of the DPPH assay provides an easy and rapid way to evaluate antioxidants by spectrophotometry (Huang et al. 2005). To the polysaccharide samples of varying concentrations (200–1000 µg/ml), 0.5 ml of 1 mM DPPH in 99.5% ethanol was added. The contents were vortexed and incubated for 30 min at room temperature (RT), and the absorbance read at 517 nm. In this assay, 1 mM DPPH was used as control.
In the ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) assay, molecules undergo nucleophilic attack by persulfate ions and form ABTS free radicals (Re et al. 1999). The EPS acts as a scavenging agent. For this assay, equal volumes of 14 mM ABTS and 5 mM APS were mixed and left for 16 h in dark. The mixture was diluted until the absorbance reached 0.7 ± 0.02. To 0.9 ml of this diluted mixture, 0.1 ml of sample was added. The contents were incubated for 6 min, and the OD read at 734 nm.
The FRAP (ferric-ion-reducing antioxidant power) assay is based on the principle that EPS reduces Fe3+ to Fe2+, which then complexes with ferricyanide (Benzie and Strain 1996). To the EPS sample in the concentration range of 200–1000 µg/ml, 0.9 ml of 96% ethanol, 5 ml of distilled H2O, 1.5 ml of 5 M HCl, 1.5 ml of 1% potassium ferricyanide, 0.5 ml of SDS, and 0.2% FeCl3 were added. The contents were incubated for 20 min at 50 °C, and the absorbance measured at 750 nm.
Emulsification activity
Emulsification activity is essential to maintain the stability of two immiscible substances. The assay was performed as per the protocol of Iyer et al. (2006). Various vegetable oils were added to EPS-containing aqueous solutions (1.0 mg/ml) in the ratio of 3:2. The mixtures were well agitated and left undisturbed. The emulsion heights were measured at an interval of 24 h for a period of 5 days. Emulsification indices were calculated using the formula H1/H2 * 100, where H1 is height of the emulsion layer and H2 is total height of the solution.
Flocculating ability of the EPS
Flocculation performance of the EPS is the most important if it is to be applied in wastewater treatment. The flocculants interact with the suspended solids present in wastewater, thereby increasing their mass, and cause rapid settling in a sedimentation tank. Flocculating ability of the EPS was estimated by the method of Li et al. (2008). Accordingly, 10 ml of activated carbon (surface area 600 m2/g, pore volume 0.95 ml/g dry basis, and maximum moisture content 12%) was taken in a test tube at a concentration of 1.0 g/l. One percentage CaCl2 and EPS were then added. A tube without the EPS served as control. The contents were vortexed for 30 s and allowed to stand for 30 min at RT. The turbidity of the upper layer was measured at 550 nm. The flocculation activity was calculated as (C–E)/C * 100, where C refers to OD of control and E refers to OD of the EPS sample. All experiments were carried out in triplicate, and the values given represent the mean.
Effect of various parameters on the flocculating ability
In order to test the influence of different parameters on the flocculating ability of the EPS, its concentration was varied from 20 to 240 µg/ml. pH plays an important role in flocculation. The presence of H+ and OH− ions may affect the binding of polysaccharide to the suspended particles. Hence, the pH was varied from 5.0 to 9.0. The valency of the cations is also a crucial factor in flocculation as they aid in binding of the EPS to the suspended particles. Monovalent cations NaCl and KCl; divalent cations FeCl2, MgCl2 and CaCl2; and trivalent cations FeCl3 and AlCl3 were used in the study.
Application in wastewater treatment
Flocculating ability of the EPS was tested using actual wastewater samples collected on 3 consecutive days from the effluent treatment plant of the local municipality (after the initial screening process). Chemical oxygen demand (COD) (closed reflux volumetric method), total solids (TS), and total suspended solids (TSS) were determined as per the protocols specified in Standard Methods for the Examination of Water and Wastewater (APHA 1995). Turbidity (Elico CL-52D), conductivity (Systronics 304), and pH (Elico LI-120) were the other effluent parameters measured. These characteristics are provided in Table 1.
The standard jar test experiment was used to test the coagulation–flocculation performance of the bacterial EPS. In this procedure, effluent samples (100 ml, pH 7.0) were treated either with FeCl3 alone or a combination of FeCl3 and EPS. The trivalent cations were added at a fixed dose of 30 mg/L, while the dose of the biopolymer was varied from 5.0 to 50 mg/L. Initially, FeCl3 was added to the effluent sample with constant stirring, followed by the addition of the EPS, with slow stirring of the suspension for another 15 min. It was then allowed to stand at RT for 30 min, and the supernatant was used for measurements of COD and TSS. Since coagulation efficiency is pH dependent, the pH of the effluent sample was then varied from 6.0 to 8.0, which is the range typically encountered in municipal wastewaters. In this study, the dose of the EPS was fixed at the optimum value.
Results and discussion
Isolation and identification of bacterial strains
Serial dilution and plating of the soil sample in sucrose-enriched selective medium led to the isolation of five different bacterial strains out of which one strain YP03 was capable of significant EPS production. It was a Gram positive and rod-shaped organism. Based on MALDI–TOF MS analysis, the identity of the bacterial isolate was established to be Bacillus mycoides, yielding a score value of 2.075, which could be considered a reliable identification (result not shown). 16S rRNA gene sequencing further confirmed the identity of the isolate as Bacillus sp., showing up to 99% homology to B. cereus and B. mycoides sequences in the NCBI GenBank database. The gene sequence was submitted in the database and was allotted the Accession No. KX404995.1 The phylogenetic tree depicting sequence relationship between Bacillus sp. YP03 and others in the database is depicted in Fig. 1. Previous reports on the production of EPS from Bacillus spp. are available for B. lentus (Abou-Taleb et al. 2015), B. subtilis (Shih and Yu 2005), and B. licheniformis (Porras-Domínguez et al. 2014). Production of fructo-oligosaccharide-based EPS by B. mycoides or B. cereus has not yet been reported.

Phylogenetic tree for Bacillus sp. YP03
Production and partial purification of EPS from Bacillus sp. YP03
Ethanol treatment of the culture supernatant resulted in precipitation of the EPS from Bacillus sp. YP03, which was then dissolved in hot distilled water and dialyzed. After dialysis, the EPS was lyophilized and finally obtained in the form of a gel-like substance. The weight of the sample was determined to be 1.52 g (produced per l of the culture). The amount was found to be comparable to that reported for Halomonas sp. by Poli et al. (2009). Further studies on the effects of parameters such as substrate concentration, pH, and temperature could optimize the production process and enhance the EPS yield obtained from YP03.
Analyses of total carbohydrate, protein, and fructose contents
Total carbohydrate content of the EPS sample was found to be 89.6%. The remaining percentage may be attributed to uronic acid and protein contents. This high carbohydrate content indicates the presence of polysaccharides. Moreover, the unhydrolyzed sample did not give any significant result in the carbohydrate assay as it does not form hydroxymethylfurfural. The monomer contents of the EPS were analyzed after hydrolyzing the sample, and its fructose content was found to be as high as 86.1%. This indicates that fructose moieties form a significant amount of the EPS. Total protein content in the partially purified EPS was found to be 5.6%, which could be removed upon further purification in chromatographic columns.
Antioxidant activity: DPPH, ABTS, and FRAP assays
Free radical scavenging property of the EPS isolated from Bacillus sp. YP03 was assessed using the DPPH method. DPPH is one of the compounds having a proton free radical with a characteristic absorption, which decreases significantly upon exposure to proton radical scavengers. The amount of DPPH free radical decreased in the presence of EPS. With an increase in concentration (200–1000 µg/ml) of the EPS, the percentage of inhibition increased from 22.7 to 38.2% (Fig. 2a). Thus, the produced EPS possess significant antioxidant potential. In the presence of antioxidants, the free radicals are converted into stable products and the chain reaction is terminated. This conversion leads to change in color from purple to yellow. Studies conducted by Li et al. (2014) had shown that the antioxidant potential of EPS from Lactobacillus helveticus MB2-1 varied from 33.5 to 42% when 4 mg/ml of sample was used.

a DPPH free radical scavenging activity of the EPS, b ABTS free radical scavenging activity of the EPS, c ferric ion reducing power of the EPS
Free radical scavenging activity of EPS was also examined by the ABTS method. In this case too, the EPS showed an increase in inhibition in a dose-dependent manner. Inhibition of free radical activity escalated from 28.9 to 44.1% as the concentration of EPS went up from 200 to 1000 µg/ml (Fig. 2b). The obtained result is consistent with the data of Li et al. (2012) who had reported the antioxidant potential of EPS isolated from fermented wheat. According to them, the ABTS free radical scavenging activity was around 40% when 1.25 mg/ml of EPS was used. They had shown 100% inhibition at a high concentration of 10 mg/ml. The antioxidant activity of a polysaccharide may depend on a combination of structural factors including its molecular weight, monosaccharide content, and configuration of the glycosidic bond (Zheng et al. 2014).
The EPS isolated from Bacillus sp. YP03 was tested for its reducing ability using FRAP assay. Enhanced OD reading from 0.010 ± 0.002 to 0.056 ± 0.004 at 750 nm was recorded as the concentration of EPS went up from 200 to 1000 µg/ml (Fig. 4). This increase is due to the formation of metal ion complex. The EPS reduces Fe3+ to Fe2+, which then binds with ferricyanide to form a complex. This result further testifies the antioxidant potential of YP03 EPS. The reducing power of Peanibacillus mucilaginosus TKU032 EPS at a dosage of 400 μg/ml was 0.65 and did not increase with concentration (Liang et al. 2016).
Emulsification activity
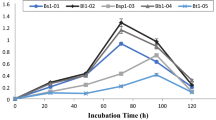
The emulsification indices over a 5-day period were represented as E 24, E 48, E 72, E 96, and E 120. In Table 2, it could be inferred that the EPS displays appreciable emulsifying ability for all the oils tested, and it was the greatest toward sesame and sunflower oils. Iyer et al. (2006) had studied the emulsification index of EPS from marine bacteria, and it was noted to be highest for groundnut oil. This property would enable the use of the compound as a biosurfactant for various applications including remediation of oil spills.
Flocculating ability of the EPS
The bacterial EPS showed appreciable flocculating ability toward the activated carbon particles and resultant turbidity removal. The effects of various factors on its performance were assessed. The EPS flocculated the suspended particles differently when its concentration was varied. The flocculation was found to increase with concentration in the range of 20–140 µg/ml, with 75% being the maximum flocculation achieved (Fig. 3a). Thereafter, it remained unchanged upon further increase in concentration. Li et al. (2008) had reported the flocculating ability of Pseudoalteromonas sp. SM9913 EPS. At a high concentration of 10 mg/ml, it had shown a maximum of 49.3% flocculation. Hence, the EPS isolated from Bacillus sp. YP03 has shown comparatively higher flocculating ability.

a Effect of EPS concentration on the flocculation of suspended particles, b effect of pH on the flocculation of suspended particles
A variation in pH influenced the flocculating characteristic of the EPS. It showed maximum performance in the pH range of 6.5–8.0. At pH below 6.0 or above 8.0, the EPS did not have good flocculating effect and its performance decreased (Fig. 3b). Suspensions of activated carbon and EPS lacking the presence of calcium ions showed poor flocculating performance. Prasertsan et al. (2006) have explained the mechanism behind the effect of pH on flocculation. When the pH increases above 8, the OH− ions influence the charge of Ca2+ ions. When the pH decreases below 6, the H+ ions are adsorbed by activated carbon and EPS. This weakens the stability of EPS-activated carbon complex. The coagulation of bentonite by chitosan was found to decrease with increase in pH of the suspension (Chatterjee et al. 2009).
Cations play an important role in flocculation as they function as mediators between the suspended particles and the EPS. In this study, the EPS showed heightened flocculation activity in the presence of trivalent and divalent cations. The flocculation performance was witnessed in the following order: Al3+ > Fe3+ > Ca2+ > Fe2+ > Mg2+ > K+ > Na+ (Fig. 4). Wu and Ye (2007) had reported the effect of cation valency on flocculation. They had shown superior flocculation of the suspended particles when trivalent cations were used. The strength of bond between the suspended particles and the flocculant influences the floc density, shear resistance of floc, and floc size. The strength of the bond is in turn influenced by the valency of cation. Since the strength of bonding is higher in trivalent cations, they show greater flocculating performance than divalent and monovalent cations.

Effect of cations on the flocculation of suspended particles
Application in wastewater treatment
The performance of primary sedimentation tank in an effluent treatment plant could be enhanced by coagulation–flocculation pretreatment. It is an efficient way of removing charged particles from wastewater. The formation of stable colloidal flocs is necessary for aggregation, and this occurs when the flocs form extended networks through self-assembly. Metal cations and synthetic polyelectrolytes are routinely used for this purpose. Replacing or supplementing them with biopolymer-based natural coagulants would improve the biodegradability of the resultant sludge and also protect us from the adverse effects of residual Al3+. In this study, the bacterial EPS was tested for its ability to bring down the pollution load of municipal wastewater by acting as an effective flocculating agent. Flasks containing only the standard coagulant FeCl3 served as control, and the test flasks additionally contained EPS as a coagulant aid. TSS and COD were the key effluent parameters monitored.
Figure 5 represents the effect of flocculant dosage on COD (a) and TSS (b) removal. From the graphs, it could be inferred that maximum TSS removal ranged from 79 to 89% for the effluent samples, while the maximum COD removal was around 41–47%. The optimum flocculant dose was seen to be 20 mg/l as higher concentrations did not yield any appreciable increase in the flocculating activity. In the absence of the coagulant aid, 24 and 44% were the maximum COD and TSS removal witnessed in the control flasks. This shows that the EPS acts as an effective coagulant aid and augments the performance of FeCl3. Suopajärvi et al. (2013) have reported turbidity reduction of 40–80% and COD removal of 40–60% using moderately low doses of nanocellulose flocculants in conjunction with ferric sulfate.

Effect of flocculant dosage on COD (a) and TSS (b) removal
Figure 6 represents the effect of effluent pH on COD (a) and TSS (b) removal. Maximum TSS removal ranged from 84 to 89%, and it was recorded at pH 7.5. Further increase in alkalinity dipped the removal efficiency down to 71–76% for the effluent samples. pH of the effluent did not have a clear pattern over turbidity removal by the biopolymer xanthan gum, and only specific pH values were suitable for the process. In the case of Locust bean gum, the turbidity removal was highest at lower pH levels and decreased as the effluent became alkaline (Mukherjee et al. 2014). COD removal ranged from 41 to 47% for the different effluent samples; this maximum efficiency also occurring at pH 7.5 and falling thereafter. In the absence of the coagulant aid, 21 and 48% were the maximum COD and TSS removals achieved in the control flasks, respectively (data not shown in graph). Hence, the removal efficiencies have almost doubled in the presence of the EPS. In studies with other biopolymers, tannin as a coagulant aid had been more effective than the synthetic polyelectrolyte AN913, and it had significantly reduced the required doses of alum (Ozacar and Sengil 2003).

Effect of effluent pH on COD (a) and TSS (b) removal
Conclusion
This study has culminated in the isolation of a promising exopolysaccharide-producing bacterial strain Bacillus sp. YP03. The compound of high-fructose content is of immense interest owing to the pronounced antioxidant, emulsifying, and flocculating activities displayed by it. The results from DPPH, ABTS, and FRAP assays have testified its free radical scavenging ability and reducing power. Moreover, its substantial flocculating ability toward activated carbon led to its exploitation in municipal wastewater treatment as a coagulant aid. 20 mg/l EPS and pH 7.5 were the optimum conditions that resulted in a maximum of 47 and 89% COD and TSS removal, respectively. In order to exploit the full potential of the isolated strain, statistical optimization of medium components for enhancing the EPS production could be an avenue for future work. Gene cloning and over-expression in heterologous hosts might also be explored for drastically improving the yield, thus making the compound highly amenable to practical application as a coagulant aid.
References
Abou-Taleb KA, Abdel-Monem MO, Yassin MH, Draz AA (2015) Production, purification and characterization of levan polymer from Bacillus lentus V8 strain. Br Microbiol Res J 5(1):22
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Al-Wasify RS, Al-Sayed A, Saleh SM, Aboelwafa AM (2015) Bacterial exopolysaccharides as new natural coagulants for surface water treatment. Int J Pharm Tech Res 8(9):198–207
APHA, Awwa, WPCF (1995) Standard methods for the examination of water and wastewater, 19th edn. American Public Health Association, American Water Works Association and Water Environment Federation, Washington
Benzie FF, Strain JJ (1996) The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal Biochem 239(1):70–76
Bradford MM (1976) Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Chatterjee T, Chatterjee S, Woo SH (2009) Enhanced coagulation of bentonite particles in water by a modified chitosan biopolymer. Chem Eng J 148(2):414–419
Domingos-Lopes MF, Nagy A, Stanton C, Ross PR, Gelencsér E, Silva CC (2017) Immunomodulatory activity of exopolysaccharide producing Leuconostoc citreum strain isolated from Pico cheese. J Funct Foods 33:235–243
DuBois KA, Gilles JK, Hamilton PA, Rebers PT, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28(3):350–356
Freitas F, Alves VD, Reis MA (2011) Advances in bacterial exopolysaccharides: from production to biotechnological applications. Trends Biotechnol 29(8):388–398
Giovanoni SJ (1991) The polymerase chain reaction. In: Stackebrandt E, Goodfellow M (eds) Nucleic acid techniques in bacterial systematics. Wiley, New York, pp 177–203
Huang DJ, Ou BX, Prior RL (2005) The chemistry behind antioxidant capacity assays. J Agric Food Chem 53:1841–1856
Indira M, Venkateswarulu TC, Chakravarthy K, Reddy A, Babu D, Kodali VP (2016) Morphological and biochemical characterization of exopolysaccharide producing bacteria isolated from dairy effluent. J Pharm Sci Res 8(2):88–91
Iyer A, Mody K, Jha B (2006) Emulsifying properties of a marine bacterial exopolysaccharide. Enzyme Microb Technol 38(1):220–222
Jeong D, Kim DH, Kang IB, Kim H, Song KY, Kim HS, Seo KH (2017) Characterization and antibacterial activity of a novel exopolysaccharide produced by Lactobacillus kefiranofaciens DN1 isolated from kefir. Food Control 78:436–442
Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16(2):111–120
Li WW, Zhou WZ, Zhang YZ, Wang J, Zhu XB (2008) Flocculation behavior and mechanism of an exopolysaccharide from the deep-sea psychrophilic bacterium Pseudoalteromonas sp. SM9913. Bioresour Technol 99(15):6893–6899
Li Y, Hu H, Zu X, Shi M, Zhang Z, Yang Y (2012) Improvement of physiological active substance of wheat distillers’ grains with solubles fermented by Preussia aemulans under optimum fermentation conditions. Int J Biol 4(4):91–100
Li W, Ji J, Chen X, Jiang M, Rui X, Dong M (2014) Structural elucidation and antioxidant activities of exopolysaccharides from Lactobacillus helveticus MB2-1. Carbohydr Polym 102:351–359
Li Y, Zhang G, Du C, Mou H, Cui J, Guan H, Hwang H, Wang P (2017) Characterization of high yield exopolysaccharide produced by sp. 921F exhibiting moisture preserving properties. Int J Biol Macromol 101:562–568. doi:10.1016/j.ijbiomac.2017.03.089
Liang TW, Tseng SC, Wang SL (2016) Production and characterization of antioxidant properties of exopolysaccharide(s) from Peanibacillus mucilaginosus TKU032. Mar Drugs 14(2):40
Mukherjee S, Mukhopadhyay S, Pariatamby A, Hashim MA, Sahu JN, Gupta BS (2014) A comparative study of biopolymers and alum in the separation and recovery of pulp fibres from paper mill effluent by flocculation. J Environ Sci 26(9):1851–1860
Özacar M, Şengil İA (2003) Evaluation of tannin biopolymer as a coagulant aid for coagulation of colloidal particles. Colloids Surf A 229(1):85–96
Patel A, Prajapat JB (2013) Food and health applications of exopolysaccharides produced by lactic acid bacteria. J Adv Dairy Res 1(2):107
Poli A, Kazak H, Gürleyendağ B, Tommonaro G, Pieretti G, Öner ET, Nicolaus B (2009) High level synthesis of levan by a novel Halomonas species growing on defined media. Carbohydr Polym 78(4):651–657
Porras-Domínguez JR, Ávila-Fernández Á, Rodríguez-Alegría ME, Miranda-Molina A, Escalante A, González-Cervantes R, Olvera C, Munguía AL (2014) Levan-type FOS production using a Bacillus licheniformis endolevanase. Process Biochem 49(5):783–790
Prasertsan P, Dermlim W, Doelle H, Kennedy JF (2006) Screening, characterization and flocculating property of carbohydrate polymer from newly isolated Enterobacter cloacae WD7. Carbohydr Polym 66(3):289–297
Qurashi AW, Sabri AN (2012) Bacterial exopolysaccharide and biofilm formation stimulate chickpea growth and soil aggregation under salt stress. Braz J Microbiol 43(3):1183–1191
Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice-Evans C (1999) Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol Med 26(9–10):1231–1237
Rehm BH (2010) Bacterial polymers: biosynthesis, modifications and applications. Nat Rev Microbiol 8(8):578–592
Roe JH (1934) A quantitative method for the determination of fructose in blood and urine. Biol Chem 107:15–22
Sam S, Kucukasik F, Yenigun O, Nicolaus B, Oner ET, Yukselen MA (2011) Flocculating performances of exopolysaccharide produced by a halophilic bacterial strain cultivated on agro-industrial waste. Bioresour Technol 102(2):1788–1794
Shih L, Yu YT (2005) Simultaneous and selective production of levan and poly (γ-glutamic acid) by Bacillus subtilis. Biotechnol Lett 27(2):103–106
Suopajärvi T, Liimatainen H, Hormi O, Niinimäki J (2013) Coagulation–flocculation treatment of municipal wastewater based on anionized nanocelluloses. Chem Eng J 231:59–67
Wu JY, Ye HF (2007) Characterization and flocculating properties of an extracellular biopolymer produced from a Bacillus subtilis DYU1 isolate. Process Biochem 42(7):1114–1123
Zheng JQ, Wang JZ, Shi CW, Mao DB, He PX, Xu CP (2014) Characterization and antioxidant activity for exopolysaccharide from submerged culture of Boletus aereus. Process Biochem 49:1047–1053
Acknowledgements
The authors are thankful to the management of Kumaraguru College of Technology, Coimbatore, India, for providing the laboratory facilities to carry out this research work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial responsibility: J Aravind.
Rights and permissions
About this article

Cite this article
Kanmani, P., Yuvapriya, S. Exopolysaccharide from Bacillus sp. YP03: its properties and application as a flocculating agent in wastewater treatment. Int. J. Environ. Sci. Technol. 15, 2551–2560 (2018). https://doi.org/10.1007/s13762-017-1416-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13762-017-1416-x