
Abstract
The suborder Heteroptera (Hemiptera) includes zoophagous and zoophytophagous species which conform diverse natural enemies’ systems with potential to control several horticultural pests. In this study, we report the assemblage structure of heteropteran bugs species inhabiting open-field and greenhouse organic tomato crops and one common adjacent non-crop solanaceous plant, Solanum sisymbriifolium, in North Buenos Aires province, Argentina, aimed to select promissory biocontrol species. Biweekly direct inspection of selected plants was carried out during a 3-year period (2017–2019) to collect hemipteran nymphs and adults. As a result, nine species and morphospecies belonging to Berytidae (zoophytophagous), Lygaeidae (phytophagous), and Miridae (phytophagous and zoophytophagous) were found, with ≥ 75% of species belonging to the latter family. The zoophytophagous mirid Tupiocoris cucurbitaceus (Spinola) was the most frequent and dominant species in all sites studied. Among the phytophagous species, Nysius simulans Stål was mostly present in greenhouse crops. The community found in greenhouse tomato crops was more diverse than that registered in open-field crops. The characterization of the heteropteran complex in organic tomato farms provides basic knowledge necessary to design pest control strategies in the region studied.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Community ecology provides the theoretical framework to study the mortality factors of insects of agricultural importance by analyzing the diversity and interspecific interactions occurring in an agroecosystem (Morin 2011). Exploring the structure of the ecological community, which involves crop and non-crop plants and the herbivorous and entomophagous insects associated, has been pointed out as useful to gather baseline knowledge for pest management, especially because it allows the identification of potential biocontrol agents, particularly for conservation biological control (DeBach 1964; Salas Gervassio et al. 2016a; López-Núñez et al. 2017). In this respect, the search for native or spontaneous natural enemies is considered a better strategy instead of using imported species as biological control agents, due to the environmental risks related to the introduction of exotic species in a new biogeographic area (van Lenteren 2012; Cédola et al. 2021).
Insect pests are a major constraint to horticultural production and many of them are common species of vegetable and fruits crops worldwide (Knapp et al. 2020). The greenhouse whitefly Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae) and the South American tomato moth Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) are two main species of a complex of native insect pests of tomato crop, Solanum lycopersicum L. (Solanales: Solanaceae), in the Neotropical region (Desneux et al. 2010; López et al. 2019). The whitefly T. vaporariorum, a highly polyphagous species reported as pest of hundreds of crops, is a major pest in temperate regions causing direct crop damage due to nymphal and adult feeding on leaves and indirect damage by vectoring plant viruses. Regarding T. absoluta, this gelechiid has gained worldwide economic importance after its invasion into European, African, and Asian continents (Desneux et al. 2011; Biondi et al. 2018; Ferracini et al. 2019). Larvae mine leaves, stems, and fruits, yielding important tomato production losses (Desneux et al. 2021). Tuta absoluta feeds on tomato but it can develop using other solanaceous plant species, cultivated and wild, and among them, the sticky nightshade Solanum sisymbriifolium Lam. (Solanales: Solanaceae) has been reported as an alternative host plant for this pest (Salas Gervassio et al. 2016a). Control of these pests is difficult due to their population growth rates, their overlapping generations, and the rapid development of genetic resistance to insecticides (Knapp et al. 2020). Biological control is widely used for whitefly species management, being predatory heteropteran bugs, coccinellids and lacewings, and nymphal and adult parasitoids important natural enemies employed against this pest by means of augmentative and conservation strategies (López et al. 2012; van Lenteren et al. 2020). In relation to T. absoluta natural enemies, there have been reported over 100 entomophagous species worldwide, and among them, mirid bugs and egg and larval parasitoids show great potential as biocontrol agents (Desneux et al. 2010; Biondi et al. 2018; Salas Gervassio et al. 2019; Ferracini et al. 2019).
The suborder Heteroptera (Hemiptera) includes ca. 45,000 species recorded throughout the world, with a wide range of feeding habits (Henry 2017; Schuh and Weirauch 2020). Particularly, the species of agricultural importance vary from strictly phytophagous, phytozoophagous, zoophytophagous to strictly zoophagous (Schaefer and Panizzi 2000; Wheeler 2001). Phytophagous species that cause different degrees of damage in crops belong to several families, i.e., Pentatomidae, Coreidae, Lygaeidae, and some Miridae (Schuh and Weirauch 2020), meanwhile others, belonging to the Anthocoridae, Geocoridae, some genera of Miridae and Nabidae families, are important biocontrol agents of insect pests (van Lenteren et al. 2017). Indeed, mirid predators are used under conservation techniques or by releasing mass reared individuals in biofactories in Europe, Asia, New Zealand, and North and South America to control several horticultural pests, as whiteflies and T. absoluta (van Lenteren 2012; van Lenteren et al. 2017; 2020).
In the Mediterranean basin, several species of Heteroptera have been reported inhabiting tomato crops and feeding on eggs and small larvae of Lepidoptera, nymphs and adults of aphids, scales, and whiteflies (van Lenteren et al. 2017). In Brazil, studies on biocontrol of pests in greenhouses showed the potential of various heteropteran predators (Anthocoridae and Miridae) against them, especially to control T. absoluta (Bueno et al. 2012; 2020; van Lenteren et al. 2016; 2017; 2018). In Argentina, most of research was focused on parasitoid species of these pests (López et al. 2020; Luna et al. 2015; Salas Gervassio et al. 2019; Viscarret et al. 2000), and recently, a predatory species Tupiocoris cucurbitaceus (Spinola) (Hemiptera: Miridae) has gained attention as a biocontrol agent (Cagnotti et al. 2016; López et al. 2019). Progress has been made to study some biological traits of this heteropteran species and currently is commercially used to control whiteflies and T. absoluta in horticultural crops in Argentina and Uruguay (Polack et al. 2017; MGAP 2020).
Considering the economic importance of T. vaporariorum and T. absoluta for tomato production, the search for entomophagous insects associated with this crop, such as heteropteran species, deserves attention. Moreover, studying the co-occurrence of predators and prey in crops at plant level can help to understand the encountering opportunities among them, which is crucial to determine the potential of these natural enemies as biocontrol agents in field conditions. Our study hypothesis was that the complex of heteropteran species naturally present in tomato farms is more diverse than that currently known. Thus, we aimed to assess the assemblage of heteropteran bugs inhabiting tomato farms in a main horticultural region of Argentina with emphasis on those species that could have potential as biological control agents of T. vaporariorum and T. absoluta. We focused our study in organic farms since they rely on cropping practices which ban chemical inputs and promote crop and non-crop plant diversity, among others, increasing species richness and natural biocontrol services (Zalazar and Salvo 2007; Rouaux et al. 2020). The objectives were (1) to identify the heteropteran species present in tomato crops and in S. sisymbriifolium—a frequent non-crop solanaceous plant present in those farms—; (2) to analyze the community structure of heteropteran species in the organic tomato farms across the cropping cycle; and (3) to assess the co-occurrence between the main predators and the two main pest species, T. vaporariorum and T. absoluta, at plant level. The information of local diversity of natural enemies is relevant to design further augmentative and conservation biological control strategies in the region studied.
Materials and methods
The study was located at La Plata Horticultural Belt, NE Buenos Aires province, Argentina. In this important horticultural region, tomato crops are planted applying a two-cycles cropping scheme (spring and summer seasons) and cultivated under open-field and protected conditions (greenhouses). Although prevention and control of pests and diseases based on spraying with neurotoxic pesticides are conventional phytosanitary practices, organic farming, which relies on conservation biological control through habitat manipulation and botanical and Bt-based insecticides use, is increasing in the region.
Six organic farms were chosen to assess the assemblage structure of heteropteran species including both open-field and greenhouse tomato crops (each crop with ≈ 200 plants). Four of them were located at La Plata and the other two sites were one at Berazategui and one at Magdalena counties (Fig. 1). Biweekly samplings were carried out along both production cycles (from August to December and from January to May) and during three consecutive years (2017 to 2019) as follows: Guillermo Hudson and Lisandro Olmos 1 2017–2018; Gorina 2018–2019; Lisandro Olmos 3 2018; Lisandro Olmos 2 2017–2018–2019; and General Mansilla 2019. The number of samples taken in each season and field varied because of the dynamic farming activities, where the producers rotate crops according to economic and market conditions. All nymphs and adults of heteropteran bugs were captured by beating leaves, fruits, and stems of randomly selected plants over a white plastic tray (40 cm × 35 cm × 10 cm, length × width × depth), and the insects were collected from the tray with an aspirator. The number of revised plants per sample date ranged between 10 and 15, estimated according to heteropteran bug density and considering a standard error of 20% (Gotelli and Ellison 2004). Moreover, the presence of T. absoluta and T. vaporariorum were recorded in each sampled plant. Plants of the solanaceous S. sisymbriifolium were also revised in a similar manner when they were present in the surroundings of open-field tomato crops, since T. absoluta (Salas Gervassio et al. 2016a) and the mirids T. cucurbitaceus and Campyloneuropsis cincticornis (Stål) (Salas Gervassio and Rocca, unpublished data) have been recorded in that species. Because S. sisymbriifolium was not always found, only the heteropteran species collected are listed, but no statistical analyses were carried out.

Sampling sites to search for heteropteran species inhabiting horticultural crops located in NE Buenos Aires province, Argentina. GH, greenhouse; OP, open-field
The collected insects were transported to the laboratory and conditioned for further studies. All specimens were kept alive, individually placed in plastic Petri dishes (10 cm × 2 cm, diameter × height), and provided with tomato leaves and eggs of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) (Brometán S.R.L. Argentina SA) as food. Dishes were labelled with information regarding sampling site and date. Subsequently, they were maintained in a walk-in chamber under controlled environmental conditions (14:10 h L:D photoperiod, 25 ± 2 °C temperature, and 70% relative humidity).
Insect species identification
First separation in morphospecies was made in vivo based on visual morphological characteristics using a stereoscopic microscope (Nikon SMZ1270). Additionally, photographic images were taken with a video camera (Micrometrics 519CU CMOS, 5.0 megapixel), attached to the microscope. Dead specimens were stored in 70% ethanol for identification. Individuals were identified using literature and available keys by specialists (EM and PMD) and information about the food habits of the species was sought using literature and online catalogues (Henry et al. 2015; Wheeler and Krimmel 2015; Konstantinov et al. 2018; López et al. 2019; Nogueira et al. 2019; Dellapé and Henry 2021).
Characterization of the heteropteran species assemblage on tomato crop
The structure of the heteropteran species community present in open-field and greenhouse organic tomato crops in the NE of Buenos Aires province was assessed by means of Hill numbers (or the effective number of species). The purpose of this analysis was to standardize the variable number of samples, while extrapolation enabled to predict the real diversity by taking into account the expected number of species undetected by the sampling effort considered. This methodology measures the degree to which proportional abundances are distributed among the species and their units are species (Hill 1973). We first calculated for all sites and cropping techniques the sample coverage that indicates the proportion of the statistical population represented by the species sampled (Chao and Jost 2012). Additionally, we estimated, based on abundance data, the three most widely used members of the family of Hill numbers: q = 0 (species richness, 0D), which counts species equally without regard to their relative abundances, q = 1 (Shannon diversity, 1D) that can be interpreted as the effective number of common species in the assemblage, and q = 2 (Simpson diversity, 2D) that accounts for the effective number of dominant species in the assemblage (Chao et al. 2014).
We used the package iNEXT (iNterpolation/EXTrapolation), using R software, to compute and plot coverage-based rarefaction and extrapolation (R/E) sampling curve.
Co-occurrence analysis
To estimate the co-occurrence between the main heteropteran predators and T. vaporariorum and T. absoluta pest species in the same tomato plant (i.e., at plant level), at each farm, we first prepared a datasheet with information that included all sampled plants in which at least one predator and/or pest species (T. absoluta and T. vaporariorum) was present. Also, we considered the T. absoluta–T. vaporariorum combination to explore which is the probability that the predator could find both prey species together in the same plant. All analyses were performed using the R software (version 3.5.1) with the package Co-occur, which uses combinatory to determine the probability that the observed frequency of co-occurrence of a species pair is significantly equal to that expected (random association α = 0.05), greater than expected: P (gt) ≤ 0.05 (association positive), or significantly lower than expected: P (lt) ≤ 0.05 (negative association) (Veech 2013).
Results
Insect species identification
A total of 40 samplings were carried out during the period studied: three in Gorina and four in General Mansilla (La Plata and Magdalena), five in Lisandro Olmos 1, 16 in Lisandro Olmos 2 and four in Lisandro Olmos 3 (all La Plata), and eight in Guillermo Hudson (Berazategui). We collected 2081 heteropteran bugs which were sorted out in nine species and morphospecies, belonging to three families (Table 1). Eight of the nine species/morphospecies were found in tomato crops, and only two species, C. cincticornis and T. cucurbitaceus (Fig. 2), were registered in S. sisymbriifolium plants (Table 1). Tupiocoris cucurbitaceus was present in all tomato farms and Jalysus sobrinus Stål was registered in five out of six farms.

Dorsal view of adults of a Tupiocoris cucurbitaceus (Spinola) and b Campiloneuropsis cincticornis (Stål), two main predatory mirids registered in organic tomato farms at La Plata Horticultural Belt (Buenos Aires province, Argentina)
Based on information of feeding habits reported for hemiptera bugs, most of species/morphospecies registered have been described as zoophytophagous, except for Nysius simulans Stål and Collaria scenica Stål which are phytophagous. The food habit of the morphospecies Dyciphini 1 could not be corroborated since species of this tribe have various feeding habits.
In relation to the common pest species present in the tomato crops sampled, besides T. absoluta, whiteflies T. vaporariorum were recorded in all farms.
Characterization of the heteropteran species assemblage on tomato crop
The highest relative abundance was observed for the mirid predatory bug T. cucurbitaceus (90.43%) in the tomato farms sampled, followed by N. simulans (6.15%), J. sobrinus (2.35%), and C. scenica (0.38%); meanwhile, the rest of mirids were rare (0.67%).
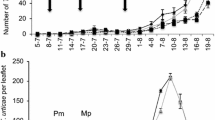
Tupiocoris cucurbitaceus was ubiquitous in all farms and recorded almost in all sampled dates, in which it generally presented the highest relative abundance. The phytophagous bug N. simulans was collected mainly from October to December during 2017, and it presented the highest relative abundance in most of these sampling dates. Jalysus sobrinus showed variable abundances in most of the sites; meanwhile, individuals of Dicyphus sp. 1 and Dicyphus sp. 2 and Nesidiocoris sp. (considered together as “other mirids”) were rarely found during the study being present only at General Mansilla site (Fig. 3).

Phenological variation of the relative abundances (%) of the heteropteran bug species found in six organic tomato farms at La Plata Horticultural Belt (Buenos Aires province, Argentina)
Regarding the characterization of the community structure of heteropteran bugs, Table 2 summarizes the values of the effective number of species calculated for both different sites and cropping techniques. The curve of sample completeness vs. sample size for each of the six sites showed that for any sample size ≥ 25 individuals, ≥ 97% sample coverage was obtained (Fig. 4a). Considering greenhouse and open-field cropping techniques, for sample size ≥ than 100 individuals, the curves showed almost 100% sample coverage (Fig. 4b).

Plots of sample coverage for samples as a function of sample size estimated for heteropteran communities associated to organic tomato farms in NE of Buenos Aires province for 3 years (2017 to 2019). a Independent six farms and b farms sorted out as greenhouse and open-field. Shaded regions show the 95% confidence intervals obtained by a bootstrap method. Reference samples are denoted by solid dots
In relation to Hill numbers, the heteropteran species richness (0D) for tomato crops varied from 1 to 4 in all sites, being Olmos 3 the community with the lower number of species (Fig. 5a). Furthermore, General Mansilla and Guillermo Hudson sites yielded the highest number of common species (1D) and very abundant species (2D) than the other sites (Fig. 5b and c). Considering the cropping techniques, richness values observed in greenhouse crops does not differ significantly from those observed in the open-field crops at abundances ≥ 100 individuals, i.e., the CI curves overlap 95% (Fig. 5d). When comparing the other Hill numbers, protected crops showed higher number of common species (1D) and greatest “very abundant” species (2D) than non-protected crops (Fig. 5e and f).

Sample-size-based rarefaction (solid lines) and extrapolation (dashed lines, up to double the reference sample size) of heteropteran communities associated to organic tomato farms in NE of Buenos Aires province for 3 years (2017 to 2019) based on the Hill numbers: a–c independent six farms and d–f farms sorted out as greenhouse and open-field. a and d 0D: q = 0, b and e 1D: q = 1, and c and f 2D: q = 2. Shaded regions show the 95% confidence intervals obtained by a bootstrap method. Reference samples are denoted by solid dots
Co-occurrence analysis
This analysis was performed for the predatory species T. cucurbitaceous because it was the most frequent and abundant heteropteran species in all farms. As a result, the presence of the predatory bug was independent (at random) of the presence of either of the two pest species on the same plant (Table 3), except in Olmos 1, where the T. cucurbitaceus–T. vaporariorum and T. absoluta–T. vaporariorum combinations were segregated at plant level with a low probability of co-occurrence, lower than 35%, while T. cucurbitaceus–T. absoluta combination was positively associated, with a higher probability of co-occurrence. In Olmos 2, prey combinations were also segregated at plant level (Table 3).
Discussion
The use of predatory biocontrol agents has been largely underestimated in augmentative releasing programs compared to parasitoids and entomopathogenic microorganisms because of their potential negative effects on other species in the agroecosystem, due to polyphagia or omnivory, preying on non-target species, and interspecific competition and intraguild predation (Labbé 2005; van Lenteren 2012). In spite of that, predatory species can play an important role in reducing pest populations, and those with a narrower diet range or oligophagous have been subject of various conservation biological control tactics and are currently gaining importance for augmentative biocontrol programs (Snyder 2019). In the last decade, several successful biocontrol programs based on predatory arthropods (mites and heteropteran bugs) have been reported in European countries to control thrips and whiteflies in sweet pepper and tomato crops (Calvo et al. 2012; Urbaneja et al. 2012). In Latin American countries, this biotechnology is considered promissory and some native predatory biocontrol agents are being evaluated (van Lenteren et al. 2020).
In this study, we found a heteropteran species assemblage inhabiting organic tomato farms in NE Buenos Aires province composed of nine species and morphospecies, seven belonging to the family Miridae, and one to Lygaeidae and Berytidae each, with various feeding habits. The finding of a greater number of mirid species in the community agrees with the taxonomic diversity of this family, represented by ca. 11,300 species worldwide (Schuh and Weirauch 2020), and > 520 described for Argentina (Ferreira et al. 2015). Species of this family have been pointed out as early colonizers into new habitats, showing that their inherent high dispersal capacity and the zoophytophagous feeding habit could allow them to exploit a variety of food resources and to rapidly increase their populations (Becker 1992). Notably, one of the phytophagous mirid species found, C. scenica, has been reported of economic importance as a pest of fodder crops (Barreto et al. 2012). For this reason, its presence in organic tomato crops in NE Buenos Aires province is worth studying. Tupiocoris cucurbitaceus, C. cincticornis, Dicyphus sp. 1 and Dicyphus sp. 2, and Nesidiocoris spp. are well-known zoophytophagous species/genera. These taxa have been reported in South America and Europe as predators of eggs and nymphs of whiteflies, and eggs and larvae of T. absoluta (Urbaneja et al. 2009; Bueno et al. 2012; Calvo et al. 2012; van Lenteren 2012; Ingegno et al. 2013; Baños-Díaz et al. 2017; López et al. 2019; Montiel Cáceres 2021). Among them, T. cucurbitaceus was a very frequent and abundant species in the heteropteran bugs assemblage, both in open-field and greenhouse tomato crops, and also found in S. sisymbriifolium plants adjacent to crops. Other mirid species, C. cincticornis, was found exclusively on the latter solanaceous species, which also hosts T. absoluta (Salas Gervassio et al. 2016a). A better understanding of the biology of C. cincticornis is necessary to determine its potential as a biocontrol agent in horticultural crops through conservation or augmentative tactics.
Both T. cucurbitaceus and C. cincticornis have been reported previously in northern Buenos Aires province by Carpintero et al. (2014). This author mentioned other eight mirid species in the region, and among them, another species of the genus Campyloneuropsis, C. infumatus Carvalho, which was not found in the present study. This species, together with Engytatus varians (Distant), and Macrolophus basicornis (Stål) are zoophytophagous bugs that have been evaluated in Brazil and were considered suitable T. absoluta control agents in tomato crops in that country (Bueno et al. 2012; 2018; Silva et al. 2016).
The species belonging to Dicyphus and Nesidiocoris have presumably great potential as natural enemies in tomato farms in NE Buenos Aires province, since other species of these genera are being commercialized as biocontrol agents (e.g., several bioproducts based on the predatory bug Nesidiocoris tenuis Reuter) (Urbaneja et al. 2009; Salas Gervassio et al. 2016b). Because of the low numbers recorded of these mirids, complementary prospections are recommended to be able to identify the species.
The greater abundance of mirid species in tomato crops, especially of T. cucurbitaceus, from the beginning of the cropping cycle (August to December) is probably related to the availability of prey (numerical response). Tupiocoris cucurbitaceus is the only species from the assemblage that has been reported as a natural enemy of T. vaporariorum and T. absoluta in Argentina (Cagnotti et al. 2020; López et al. 2019; Montiel Cáceres 2021). A decade ago, the company Brometán S.R.L. Argentina S.A. started its commercial production (Polack et al. 2017); however, it is noteworthy to mention that in this study, the co-occurrence of the predatory bug with these pests in field conditions was found mainly as independent. This result is relevant when considering augmentative releases of this biological control agent, since its effectiveness will depend, to some extent, on the encounter rates between the predator and the prey.
In relation to the other two families of heteropteran bugs recorded, the lygaeid N. simulans is a phytophagous species which was reported damaging several crops in Argentina (Dellapé 2014), and its pest status has been cited for soybean crops since it damages early vegetative crop stages (Aragón and Flores 2006). Populations of Nysius can increase in presence of its preferred host plant and eventually move to colonize new plant species, often of agricultural importance (Henry et al. 2015).
Conversely, despite most berytids appear to be predominantly phytophagous, many species exhibit predatory tendencies, especially on insects entrapped on the sticky stems of glandular plants (Henry 1997). There are no reports about predatory activity as a biocontrol agent of the berytid bug J. sobrinus; consequently, more studies are required to determine the feeding habits of this species in tomato crops.
At least other four species/morphospecies that conform the heteropteran bugs assemblage, i.e., C. cincticornis, Dicyphus sp. 1 and Dicyphus sp. 2, and Nesidiocoris sp., reported as zoophytophagous, could be also potential biocontrol candidates to manage horticultural pests — particularly T. absoluta — in Argentina. Preliminary studies showed that C. cincticornis nymphs and adults preyed on T. absoluta (Montiel Cáceres 2021). Regarding the other three taxa, as we mentioned above, species of the same genus are being used successfully in augmentative biocontrol programs (Calvo et al. 2012; Giorgini et al. 2019). Besides, it is well-known that habitat manipulation in an agroecosystem promotes biological diversity of natural enemies through differing patterns of crop colonization and niche partition (Snyder 2019).
Organic horticultural farms in NE Buenos Aires can support a diverse and abundant spontaneous heteropteran bug community, being some of them predatory species well-adapted to open-field and greenhouse tomato crops, and to spontaneous solanaceous species, such as S. sisymbriifolium. This study showed also that protected crops probably provide better conditions for the establishment of their populations, as is the case for T. cucurbitaceus, which becomes dominant in greenhouses. This constitutes an evidence that producers that reduce or eliminate applications of chemical pesticides at their farms could profit through conserving these natural enemies’ populations in crop and non-crop vegetation to maximize pest control (van Driesche et al. 2008). An exception can be observed at Gorina site where only two species were found although most of individuals belonged to T. cucurbitaceus, a result reflected by the wider confidence interval observed (Fig. 5a).
In Argentina, there is important progress in the evaluation of other natural enemies of T. vaporariorum and T. absoluta. López and Botto (1995) and López et al. (2005) have studied various biological features of the parasitoid Encarsia formosa (Hymenoptera: Aphelinidae) that attacks T. vaporariorum. In relation to T. absoluta, the larval parasitoids Dineulophus phthorimaeae de Santis (Hymenoptera: Eulophidae) and Pseudapanteles dignus Muesebeck (Hymenoptera: Braconidae), which are spontaneously present in conventional and organic tomato farms, are also under assessment as biocontrol agents of this pest (Savino et al. 2012; Nieves et al. 2015; Salas Gervassio et al. 2016a; 2018; 2019). Thus, the combined action of these parasitoids, added to the mortality inflicted by predatory species that remain to be studied, could help to maintain these pests under the threshold action. Nonetheless, it is important to consider that predatory species can engage in competitive or intraguild predation interactions with other entomophagous species, which could affect biocontrol (Symondson et al. 2002). Hence, we propose to deepen biological and ecological studies of the natural enemies’ complex of T. vaporariorum and T. absoluta, with the aim of synergizing their action in augmentative and conservation biological control programs and integrated pest management in Argentinian tomato crops.
References
Aldrich JR (1988) Chemical ecology of the Heteroptera. Ann Rev Entomol 33:211–238
Aragón J, Flores F (2006) Control integrado de plagas en soja en el sudeste de Córdoba. https://inta.gob.ar/documentos/control-integrado-de-plagas-en-soja-en-el-sudeste-de-cordoba. Accessed December 2021.
Baños-Díaz HL, Ruiz-Gil T, Del Toro-Benítez M, Mirada Cabrera I, Martínez-Rivero MA (2017) Desarrollo, reproducción y tablas de vida de Nesidiocoris tenuis Reuter empleando como presa estadios inmaduros de mosca blanca. Rev Protección Veg 32(2):1–10
Barreto N, Arce B, Rodríguez G, Arreaza LC, Rojas E (2012) Ajuste y validación de un prototipo de modelo para el manejo de la chinche de los pastos Collaria scenica para fincas lecheras del Altiplano Cundiboyacense. Informe Final, Corpoica, Centro de Investigación Tibaitatá, Mosquera, Colombia
Becker P (1992) Colonization of islands by carnivorous and herbivorous Heteroptera and Coleoptera: effects of island area, plant species richness, and ‘extinction’ rates. J Biogeogr 19(2):163–171
Biondi A, Guedes RNC, Wan FH, Desneux N (2018) Ecology, worldwide spread, and management of the invasive South American tomato pinworm, Tuta absoluta: past, present, and future. Ann Rev Entomol 63:239–258
Bueno VHP, van Lenteren JC, Lins JC Jr, Calixto AM, Montes FC, Silva DB, Santiaga LD, Pérez LM (2012) New records of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) predation by Brazilian Hemipteran predatory bugs. J Appl Entomol 137:29–34
Bueno VHP, Lins JC Jr, Silva DB, van Lenteren JC (2018) Is predation of Tuta absoluta by three Neotropical mirid predators affected by tomato lines with different densities in glandular trichomes? Arth-Plant Int. https://doi.org/10.1007/s11829-018-9658-1
Bueno VHP, Parra JRP, Bettiol W, van Lenteren JC (2020) Biological control in Brazil. In: Van Lenteren JC, Bueno VHP, Luna MaG, Colmenarez YC (eds), Biological control in Latin America and the Caribbean: its rich history and bright future, CABI, pp 78–107.
Cagnotti CL, Andorno AV, Hernández CM, Paladino LC, Botto EN, López SN (2016) Inherited sterility in Tuta absoluta (Lepidoptera: Gelechiidae): pest population suppression and potential for combined use with a generalist predator. Fla Entomol 99:87–94
Cagnotti CL, Arias AE, Ermantraut EN, Andorno AV, Viscarret MM, López SN (2020) Life history study of the mirid Tupiocoris cucurbitaceus feeding on Tuta absoluta eggs: implications for biological control and its combination with inherited sterility. Biocontrol 66:207–216
Calvo FJ, Lorente MJ, Stansly PA, Belda JE (2012) Preplant release of Nesidiocoris tenuis and supplementary tactics for control of Tuta absoluta and Bemisia tabaci in greenhouse tomato. Entomol Exp Appl 143:111–119
Carpintero DL, De Biase S, Konopko SA (2014) Hemiptera-Heteroptera de la Reserva Ecológica Costanera Sur (Ciudad Autónoma de Buenos Aires). Argentina Rev Mus Argent Cienc Nat 16(1):67–80
Cédola C, Luna MG, Achinelly MF, Sánchez NE (2021) Contributions to improve current environmental risk assessment procedures of GABCAs in Argentina. Biocontrol 66(1):153–166
Chao A, Jost L (2012) Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology 93:2533–2547
Chao A, Gotelli NJ, Hsieh TC, Sander EL, Ma KH, Colwell RK, Ellison AM (2014) Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84(1):45–67
Cornelissen T, Stiling P (2006) Does low nutritional quality act as a plant defense? An experimental test of the slow-growth, high-mortality hypothesis. Ecol Entomol 31:32–40
DeBach P (1964) Biological control of insect pests and weeds. Cambridge University Press, Cambridge, UK, p 844
Dellapé PM (2014) Lygaeoidea. In: Roig-Juñent S, Claps LE, Morrone JJ (eds), Biodiversidad de Artrópodos Argentinos. Volumen 3, Editorial INSUE - UTN, Argentina, pp 421–438.
Dellapé PM, Henry TJ (2021) Lygaeoidea species file. Version 5.0/5.0. Available from: http://Lygaeoidea.SpeciesFile.org/. Accessed December 2021.
Desneux N, Wajnberg E, Kag W, Burgio G, Arpaia S et al (2010) Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J Pest Sci 83:197–215
Desneux N, Luna MG, Guillemaud T, Urbaneja A (2011) The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. J Pest Sci 84:403–408
Desneux N, Han P, Mansour R, Arnó J, Brévaul T, Campos MT et al (2021) Integrated pest management of Tuta absoluta: practical implementations across different regions around the world. J Pest Sci. https://doi.org/10.1007/s10340-021-01442-8
Ferracini C, Bueno VHP, Dindo ML, Ingegno BL, Luna MG, Salas Gervassio NG, Sánchez NE, Siscaro G, van Lenteren JC, Zappala L, Tavella L (2019) Natural enemies of Tuta absoluta in the Mediterranean basin, Europe and South America. Biocontrol Sci Technol 29:578–609
Ferreira PSF, Henry TJ, Coelho LA (2015) Plant bugs (Miridae). In: Panizzi AR, Grazia J (eds) True bugs (Heteroptera) of the Neotropics. Springer Science, Dordrecht, pp 237–286
Giorgini M, Guerrieri E, Cascone P, Gontijo L (2019) Current strategies and future outlook for managing the Neotropical tomato pest Tuta absoluta (Meyrick) in the Mediterranean Basin. Neotrop Entomol 48:1–17
Gotelli NJ, Ellison AM (2004) A primer of ecological statistics, vol 1. Sinauer & Associates, Sunderland, MA
Henry TJ (1997) Monograph of the stilt bugs, or Berytidae, of the Western Hemisphere, memoirs of the Entomological Society of Washington, 19:149 pp.
Henry TJ, Dellapé PM, De Paula AS (2015) The big-eyed bugs, chinch bugs, and seed bugs (Lygaeoidea). In: Panizzi AR, Grazia J (eds) True bugs (Heteroptera) of the Neotropics. Springer Science, Dordrecht, pp 459–514
Henry TJ (2017) Biodiversity of Heteroptera. In: Foottit RG, Adler PH (eds), Insect biodiversity: science and society, vol. I, 2nd ed, John Wiley & Sons Ltd., pp 279–335.
Hill MO (1973) Diversity and evenness: a unifying notation and its consequences. Ecology 54:427–432
Ingegno BL, Ferracini C, Gallinotti D, Alma A, Tavella L (2013) Evaluation of the effectiveness of Dicyphus errans (Wolff) as predator of Tuta absoluta (Meyrick). Biol Control 67:246–252
Knapp M, Palevsky E, Rapisarda C (2020) Insect and mite pests. In: Albajes R, Nicot PC (eds) Gullino L. Integrated pest and disease management in greenhouse crops, Springer Nature Switzerland, pp 101–146
Konstantinov FV, Namyatova AA, Cassis G (2018) A synopsis of the Bryocorine tribes (Heteroptera: Miridae: Bryocorinae): key, diagnoses, hosts and distributional patterns. Invertebr Syst 32(4):866–891
Labbé R (2005) Intraguild interactions of the greenhouse whitefly natural enemies, predator Dicyphus hesperus, pathogen Beauveria bassiana and parasitoid Encarsia formosa. M.Sc. Thesis, Université Laval. Faculté des Sciences de l´ Agriculture et del´ Alimentation. Québec, Canadá
Logarzo GA, Williams L, Carpintero DL (2005) Plant bugs (Heteroptera: Miridae) associated with roadside habitats in Argentina and Paraguay: host plant, temporal, and geographic range effects. Ann Entomol Soc Am 98(5):694–702
López SN, Botto EN (1995) Parámetros biológicos del parasitoide Encarsia formosa (Gahan) [Hymenoptera: Aphelinidae] en condiciones de laboratorio. Ecol Austral 5:105–110
López SN, Viscarret MM, Andorno AV, Botto E (2005) Estudio de la interacción entre Encarsia formosa Gahan y Eretmocerus corni Haldeman [Hymanoptera: Aphelinidae], parasitoides de la mosca blanca de los invernáculos Trialeurodes vaporariorum (Westwood) [Hemiptera: Aleyrodidae]. Rev Investig Agropecu (RIA) 34(3):73–82
López SN, Arce Rojas F, Villalba Velásquez V, Cagnotti C (2012) Biology of Tupiocoris cucurbitaceus (Hemiptera: Miridae), a predator of the greenhouse whitefly Trialeurodes vaporariorum (Hemiptera: Aleyrodidae) in tomato crops in Argentina. Biocontrol Sci Technol 22(10):1107–1117
López S, Muñoz AO, Andorno AV, Cuello EM, Cagnotti CL (2019) Predatory capacity of Tupiocoris cucurbitaceous (Hemiptera Miridae) on several pests of tomato. Bull Insectology 72(2):201–205
López SN, D´Alessandro CP, Viscarret MM (2020) Control biológico de moscas blancas. In: POLACK A ET AL (Eds.), Control biológico de plagas en horticultura. Experiencias argentinas de las últimas tres décadas, INTA Ediciones Instituto de Microbiología y Zoología Agrícola–IMYZA–CICVyA–CNIA, Argentina, pp 373–409.
López-Núñez FA, Heleno RH, Ribeiro S, Marchante H, Marchante E (2017) Four-trophic level food webs reveal the cascading impacts of an invasive plant targeted for biocontrol. Ecology 98(3):782–793
Luna MG, Pereyra PC, Coviella CE, Nieves E, Savino V, Salas Gervassio NG, Luft E, Virla E, Sánchez NE (2015) Potential of biological control agents against Tuta absoluta (Lepidoptera: Gelechiidae): current knowledge in Argentina. Fla Entomologist 98:489–494
Mahdavi TS, Madadi H, Biondi A (2020) Predation and reproduction of the generalist predator Nabis pseudoferus preying on Tuta absoluta. Entomol Exp Appl 68:732–741
MGAP. Ministerio de Ganadería, Agricultura y Pesca (2020) https://www.gub.uy/ministerio-ganaderia-agricultura-pesca/comunicacion/noticias/promueven-control-plagas-agricolas-uso-controladores-biologicos-uruguay. Accessed December 2021.
Montiel Cáceres R (2021) Estudio de hemípteros depredadores en cultivos de tomate del noreste de Buenos Aires, Argentina: ¿posibles enemigos naturales de Tuta absoluta (Meyrick)? Tesis de maestría en Protección vegetal, EPG-UBA. http://ri.agro.uba.ar/greenstone3/library/collection/tesis/document/2021montielrocio
Moraes RCB, Haddad ML (2003) Software para análise faunística. In: Simpósio de Controle Biológico 8°, São Paulo, Resumos, Piracicaba: ESALQ/USP, p 185.
Morin PJ (2011) Communities. In: Morin PJ (ed), Community ecology, 2nd ed, Oxford, UK, pp 1–23.
Nieves E, Pereyra PC, Luna MG, Medone P, Sanchez NE (2015) Laboratory population parameters and field impact of the larval endoparasitoid Pseudapanteles dignus (Hymenoptera: Braconidae) on its host Tuta absoluta (Lepidoptera: Gelechiidae) in tomato crops in Argentina. J Econ Entomol 108:1553–1559
Nogueira BCF, Ferreira PSF, Coelho LA, Martins DS, Barcellos BD (2019) Plant bugs predators (Hemiptera: Heteroptera: Miridae) with references to arthropods and fungi in Brazil. An Acad Bras Cienc 91(3):e20181194
Polack LA, López SN, Silvestre C, Viscarret M, Andorno A, Del Pino M, Peruzzi G, Gomez J, Iezzi A (2017) Control biológico en tomate con el mírido Tupiocoris cucurbitaceus. Instituto Nacional de Tecnología Agropecuaria (INTA), [online] URL: https://inta.gob.ar/documentos/control-biologico-en-tomate-con-el-mirido-tupiocoris-cucurbitaceus. Accessed December 2021.
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.Rproject.org/. Accessed 3 Oct 2022
Rouaux J, Cabrera N, Martínez AS, Posse MC, Luna MG (2020) Diversity and phenology of epigeal Coleoptera assemblages in lettuce and tomato crops in Northern Buenos Aires province, Argentina. An Acad Bras Cienc 92 (suppl 1). https://doi.org/10.1590/0001-3765202020181391
Sakagami SF, Laroca L (1971) Relative abundance, phenology and flower visits of apid bees in eastern Paraná, southern Brazil (Hymenoptera, Apidae). Kontyû 39(3):217–230
Salas Gervassio NG, Luna MG, Lee S, Salvo A, Sánchez NE (2016) Trophic web associated with the South American tomato moth Tuta absoluta: implications for its conservation biological control in Argentina. Agric Forest Entomol 18:137–144
Salas Gervassio NG, Pérez-Hedo M, Luna MG, Urbaneja A (2016) Intraguild predation and competitive displacement between Nesidiocoris tenuis and Dicyphus maroccanus, two biological control agents in tomato pests. Insect Sci 24:809–817
Salas Gervassio NG, Luna MG, D’Auro F, Sánchez NE (2018) Performance of Pseudapanteles dignus (Hymenoptera: Braconidae), a natural enemy of Tuta absoluta (Lepidoptera: Gelechiidae) in eggplant. J Econ Entomol 111(2):528–532
Salas Gervassio NG, Aquino D, Vallina C, Biondi A, Luna MG (2019) A re-examination of Tuta absoluta parasitoids in South America for optimized biological control. J Pest Sci 92:1343–1357
Savino V, Coviella CE, Luna MG (2012) Reproductive biology of Dineulophus phhtorimaeae De Santis (Hymenoptera: Eulophidae), a natural enemy of the tomato moth Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). J Insect Sci 12:153. https://doi.org/10.1673/031.012.15301
Schaefer CW, Panizzi AR (2000) Heteroptera of economic importance. CRC Press, Boca Raton, p 856. https://doi.org/10.1201/9781420041859
Schuh RT, Weirauch C (2020) True bugs of the world (Hemiptera: Heteroptera): classification and natural history, 2nd edn. Siri Scientific Press, Manchester, UK, p 800
Silva DB, Bueno VHP, Calvo FJ, Van Lenteren JC (2016) Do nymphs and adults of three Neotropical zoophytophagous mirids damage leaves and fruits of tomato? Bull Entomol Res 107:200–207
Snyder WE (2019) Give predators a complement: conserving natural enemy biodiversity to improve biocontrol. Biol Control 135:73–82
Symondson WOC, Sunderland KD, Greenstone MH (2002) Can generalist predators be effective biocontrol agents? Annu Rev Entomol 47:561–594
Urbaneja A, Montón H, Mollá O (2009) Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. J Appl Entomol 133:292–296
Urbaneja A, Gonzalez J, Arno J, Gabarra R (2012) Prospects for the biological control of Tuta absoluta in tomatoes of the Mediterranean basin. Pest Manag Sci 68:1215–1222
Valladares G, Salvo A (1999) Insect-plant food webs could provide new clues for pest management. Environ Entomol 28:539–544
Van Driesche R, Hoddle M, Center T (2008) Control of pests and weeds by natural enemies: an introduction to biological control. Blackwell Publishing
van Lenteren JC (2012) The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. Biocontrol 57:1–20
van Lenteren JC, Hemerik L, Lins J, Bueno VHP (2016) Functional responses of three neotropical mirid predators to eggs of Tuta absoluta on tomato. Insects 7(34):1–10
van Lenteren JC, Bueno VHP, Smit J, Soares MA, Calixto AM, Montes FC, De Jong P (2017) Predation of Tuta absoluta eggs during the nymphal stages of three Neotropical mirid predators on tomato. Bull Insectology 70(1):69–74
van Lenteren JC, Bueno VHP, Montes FC, Hemerik L, De Jong PW (2018) Adult lifetime predation of Tuta absoluta eggs by three Neotropical mirid predators on tomato. Bull Insectology 71(2):179–188
van Lenteren JC, Bueno VHP, Luna MG, Colmenarez Y (2020) Biological control in Latin America and the Caribbean: its rich history and bright future. CABI, p 550
Veech JA (2013) A probabilistic model for analysing species co-occurrence. Glob Ecol Biogeogr 22:252–260
Veech JA (2014) The pairwise approach to analysing species co-occurrence. J Biogeogr. https://doi.org/10.1111/jbi.12318
Viscarret MM, Botto EN, Polak A (2000) Whiteflies (Hemiptera: Aleyrodidae) of economic importance and their natural enemies (Hymenoptera: Aphelinidae, Signiphoridae) in Argentina. Revista Chilena De Entomología 26:5–11
Wheeler AG Jr. (2001) Biology of the plant bugs (Hemiptera: Miridae). Predators, Opportunists, Cornell University Press, Pest, p 507
Wheeler AGJr, Krimmel BA (2015) Mirid (Hemiptera: Heteroptera) specialists of sticky plants: adaptations, interactions, and ecological implications. Annu Rev Entomol 60:393–414
Zalazar L, Salvo A (2007) Entomofauna asociada a cultivos hortícolas orgánicos y convencionales en Córdoba. Argentina Neotrop Entomol 36(5):765–773
Acknowledgements
We thank Eliana Nieves for the help in field sampling and the owners of the farms where the samplings were carried out. Laura Morote, Luis Gianbelluca, and M. Rosario Iglesias kindly helped designed Figs. 1 and 2. This research was supported by grants from the Agencia Nacional de Promoción Científica y Tecnológica PICT 2019-0745 and by the Universidad Nacional de La Plata Programa de Incentivos N 829.
Author information
Authors and Affiliations
Contributions
RMC carried out the samplings, performed the analysis of data, and the interpretation and discussion of results. NGSG collaborated in the interpretation and discussion of results, prepared the draft of the manuscript, and participated in the final version of the manuscript. EM and PMD carried out the identification of all insects collected and participated in the edition of the final version of the manuscript. MGL participated in the planning of the assays, interpretation, and discussion of results, as well as in the writing of the final version of the manuscript. She also obtained the funds to carry out the investigation. MR participated in the planning of the assays, collaborated in the analysis of data, interpretation, and discussion of results, and in the writing of the final version of the manuscript, and also obtained the funds to carry out the investigation.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Edited by German Vargas.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Montiel Cáceres, R., Salas Gervassio, N.G., Minghetti, E. et al. Heteropteran Bugs Assemblage Associated to Organic Tomato Farms: Knowledge for Pest Management. Neotrop Entomol 52, 251–262 (2023). https://doi.org/10.1007/s13744-022-01007-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13744-022-01007-z