
Abstract
Sex pheromones produced by females of Rhagoletis conversa Bréthes and Rhagoletis brncici Frías are deposited on the surface of fruits after the eggs are laid. These pheromones repel other females, preventing repeated oviposition on the same fruit. They also attract males, thus assisting mating. Observations were made on wild populations, and cross-species behavioral tests were performed on males. The pheromone released by females was significantly more attractive for males of the same species. The two species showed remarkable differences in mating behavior, in the duration of oviposition, and in the number of circuits made around the fruit after eggs were laid. A morphological analysis of flies collected from their respective host plants indicated great host fidelity and the reproductive isolation of the two species. Possibly, the specific male-arresting effect of this pheromone was important for the sympatric speciation and evolution of these sibling species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The genus Rhagoletis Loew (Diptera: Tephritidae) is composed of about 64 species extensively distributed in the Neotropical, Nearctic, and Palearctic regions, with few species being collected in the Oriental region (Norrbom et al. 1998). Many of these species have economic importance because their life cycles are associated with the fruits of cultivated plants. Individual species may be monophagous, oligophagous, or polyphagous. After mating, females show elaborate oviposition behavior in which they puncture the fruit and lay one egg per puncture (Bush 1966). Their oviposition behavior includes an exploratory phase in which the female contacts the surface of the fruit with the ovipositor to obtain information concerning its size, shape, and chemical properties. After the eggs are deposited, the ventral zone of ovipositor is dragged across the surface of the fruit impregnating it with an oviposition-deterring pheromone (ODP). This pheromone repels other females, thus impeding repeated oviposition in the same fruit (Prokopy & Bush 1972, Katsoyannos 1975, Prokopy et al 1976, Prokopy & Roitberg 1984, Díaz-Fleischer et al 2000, Silva et al 2012).
The ODP persists on the fruit for several days. It is soluble in water and methanol, and therefore can be easily washed off the substrate (Katsoyannos 1975, Mumtaz & Aliniazee 1983). The pheromones of Tephritidae are low-molecular-weight volatile compounds derived from alkanes (Wicker-Thomas 2007). In Rhagoletis cerasi (Linnaeus), palmitylglucopyranoside was identified as an active component of the ODP (Boller et al 1987). In Rhagoletis, multiple ovipositions within the same fruit are inhibited by the ODP, preventing intraspecific competition for food (Katsoyannos 1975). The action of marking the oviposition site has been reported for 23 frugivorous species within the genera Anastrepha Schiner, Ceratitis Bezzi, and Rhagoletis (Silva et al 2012). For Rhagoletis, these studies have been performed only in Nearctic and Palearctic species (Prokopy & Bush 1972).
It was reported that the ODP also attracts males that court and mate with females arriving at the fruit (Katsoyannos 1975). Species belonging to different groups produce different pheromones, but closely related species may be attracted by pheromones of other species within the group (Prokopy et al 1976). Nearctic and Palearctic species of Rhagoletis, including the Nearctic species Rhagoletis conversa Bréthes and Rhagoletis brncici Frías, have been observed initiating copulation on the fruits of their larval host plants (Katsoyannos 1975, Frías et al 1984). In this article, we report on aspects of the reproductive behavior of two sibling species, R. conversa and R. brncici, whose host plants are Solanum ligustrinum and Solanum nigrum, respectively. Daily activity of these flies occurs on the fruits of these host plants (Frías et al 1984), which grow in various localities in the Central Valley of Chile. Ecological and morphological data concerning the adults and immature stages indicate reproductive isolation of these species. Both species are monophagous and follow a model of sympatric speciation (Frías 2001). The effects of their ODPs have been evaluated in relation to their intraspecific roles and from the perspective of their application in the pest control. However, the influences of ODPs on phylogenetically closely related species have not been studied from an evolutionary point of view. Such behavioral studies might better explain the reproductive behavior and isolation of sibling species observed in wild sympatric populations. Here, we examine the interspecific effects of the ODP on males of these species, i.e., the effects of the pheromone of female R. conversa on males of R. brncici, and vice versa.
Thus, the objectives of this study were (1) to observe mating and the effects of the pheromones on male and female behavior in wild populations of both species; (2) to make laboratory observations of the durations of mating, oviposition, and ovipositor dragging, and of the number of circuits around the fruit made by females; (3) to estimate the average time spent by males of R. brncici and R. conversa that alight on fruits impregnated with the ODPs of R. brncici and R. conversa females; and (4) to compare this behavior with other species of the nova group, including Rhagoletis tomatis Foote and Rhagoletis nova (Schiner).
Material and Methods
Fruits of wild plants of S. ligustrinum infested by R. conversa and S. nigrum infested by R. brncici were collected in Las Vertientes, 35 km southeast of Santiago (33°35′17′′S, 70°27′45′′W). The fruits were brought to the laboratory and deposited on to a grid placed over a plastic tray. The larvae were able to pass through the grid and were collected in the tray, where they pupated. Males and females were obtained from the pupae. The experiments used 8-day-old sexually mature virgin female and male flies. Observations were made at room temperature (about 20°C) in population boxes (30 × 30 × 30 cm) with glass tops. Using a stopwatch, we recorded the durations of mating, oviposition, and ovipositor dragging by females, and we also noted the number of circuits around the fruit made by females during the dragging behavior. To estimate mating duration, one female/male pair was deposited inside each box. After mating, the females were transferred to a box containing 2-cm-diameter artificial agar fruits wrapped in a parafilm “M” (Laboratory Film Pechney Plastic Packacing Chicago, IL 60631). For the preparation of artificial fruits, a ratio of 1 g of agar-agar Winkler Ltda, Santiago, Chile and 100 mL of water was used. This mixture was placed in a glass beaker and heated waving to boiling. This mixture was then emptied into plastic molds and allowed to cool. Subsequently, agar spheres were wrapped with parafilm. The females laid eggs in these artificial fruits and then dragged their ovipositors across the surface, depositing the ODP. These fruits were used in the behavioral experiments with males. Artificial fruits were used to exclude the effect of plant volatiles on the insect behavior (Aluja et al 1989, Aluja & Prokopy 1992, Linn et al 2003).
To investigate whether R. brncici ODP attracts males of R. conversa and if ODP of R. conversa attracts males of R. brncici, non-choice tests were conducted. Rhagoletis brncici males were offered artificial fruits marked either by R. brncici ODP or by R. conversa ODP. In both treatments, controls lacking ODP were included. One male was introduced into each box, and the time spent on the surface of the fruit was recorded over a period of 15 min. Males that did not alight on the fruit in this time were replaced. In each experiment, a total of 100 males that settled on fruit were observed.
In another experiment, a crossover choice test was performed. Individual males of either R. brncici or R. conversa were introduced into a box containing two artificial fruits, one impregnated with ODP from conspecific females and the other with ODP from the other species. When the male arrived at one of these fruits, its residence time was measured over 15 min. Males that did not alight on the fruits were replaced. In total, 100 males of each species were observed. Student’s t test was applied to determine the statistical significance of differences between the mean residence times obtained in each experiment.
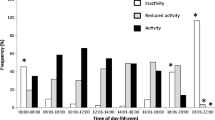
Field observations were made on female and male flies in wild populations of both species on their host plants. Observations were made on 73 females of R. conversa laying their eggs inside of fruits of S. ligustrinum and 63 females of R. brncici ovipositing on S. nigrum. A total of 105 males perched on fruits of S. ligustrinum and 109 males on fruits of S. nigrum were observed. X 2 test was applied to determine the statistical significance of these field observations. We observed 110 matings on the host plants: 65 on S. ligustrinum and 45 on S. nigrum. All of these flies were collected and transported to the laboratory for identification based on their morphology (Frías 2001, 2007) to analyze their host fidelity. If males were observed perching on a fruit, the fruit was collected to determine whether it had been infested. Similarly, when a female was observed to lay eggs inside a fruit, the fruit was collected to determine whether it had been previously infested. For each fly species, the percentage of males and females that perched on infested and uninfested fruits was estimated. The field observations were made in Las Vertientes, where the host plants of these species coexist, in November and December 2012 and 2013. For both species, the largest number of ovipositions and matings occur during this period (Frías 1989). Observations were made between 10:00 and 11:00 h, and between 18:00 and 19:00 h, which are the periods of maximum activity of these flies (Frías et al 1984).
Results
Field observations
Observations in natural populations of both species showed that the females dispersed and searched for fruit in which to lay their eggs. Females of R. conversa oviposited more frequently in uninfested fruits (83%) than in infested fruits (17%). Similarly, females of R. brncici laid eggs in a greater percentage of uninfested fruits (89%) (X 2 = 0.42, p > 0.50). Conversely, males perched for longer on the infested fruits of their respective host plants. Thus, in R. conversa 81% of males were observed perched on infested fruits of their host plant S. ligustrinum while 78% males of R. brncici were perched on infested fruits of S. nigrum (X 2 = 0.13, p > 0.70). These observations indicate that, for males of both species, infested fruits were more attractive than uninfested fruits and suggest that the ODP might be involved in their reproductive behavior. When intraspecific behavior of males and females from each species are compared in the field, differences are highly statistically significant; among R. conversa males and females (X 2 = 69.69, p < 0.0001) and between males and females of R. brncici (X 2 = 69.10, p < 0.0001). Many matings were observed in which females approached males. In these flies in wild populations, no courtship was observed before matings. Moreover, many attempted matings between males were observed.
Mating and oviposition behavior under experimental conditions
The duration of the ovipositor-dragging behavior was similar in the two species (nearly 33.5 s). Mating was significantly longer in R. brncici (15.6 ± 3.95 min) than in R. conversa (11.8 ± 4.20 min) (t = 6.84, p < 0.05). The oviposition time was significantly greater in R. brncici (6.8 ± 4.30 min) than in R. conversa (1.3 ± 0.62 min) (t = 12.68, p < 0.025). During the dragging, the females of R. conversa carry out a significantly greater number of circuits (about nine) than the females of R. brncici (about seven) (t = 9.05, p < 0.05).
Non-choice test
The non-choice test showed that the males of R. brncici perched on fruits impregnated with the ODP of R. brncici for a significantly longer time (2.72 ± 1.47 m) than did males of R. conversa (1.1 ± 1.05 min) (t = 6.40, p < 0.05). Likewise, males of R. conversa remained for significantly longer period (7.1 ± 4.81 min) on fruits treated with the ODP of females of R. conversa than did males of R. brncici (1.6 ± 1.32 min) (t = 10.94, p < 0.05).
Choice test
The choice test between fruits marked with the ODP of R. conversa and R. brncici females showed that males of both species spent more time perching on fruits impregnated with the ODP of their own species. Males of R. brncici perched for a significantly longer period (11.7 ± 3.13 min) on fruits impregnated with the ODP from female R. brncici than the males of R. conversa (1.3 ± 0.80 min) (t = 32.50, p < 0.025) and males of R. conversa perched significantly longer on fruits saturated with the ODP of females of their own species (2.9 ± 1.31 m) than the males of R. brncici (1.5 ± 1.64 min) (t = 6.62, p < 0.05).
Discussion
The deterrent effect of the ODP on females of R. conversa and R. brncici is similar to that described in other Palearctic and Nearctic species of the genus Rhagoletis, e.g., Rhagoletis pomonella Walsh and R. cerasi (Katsoyannos 1975, Prokopy et al 1976, Boller et al 1987). Females of all of these species preferentially lay their eggs in fruits that have not been previously infested. However, this behavior differs from that described in females of R. tomatis in which the pheromone released by females attracts other females (Frías 1995), allowing aggregation for the oviposition of females in a single fruit. As a consequence, many immature stages may coexist in one fruit (Frías 2014). Like R. conversa and R. brncici, female R. tomatis oviposit a single egg in each puncture hole. However, in the latter species, after oviposition, the ODP is deposited by marking a circle around the puncture. Subsequent females may visit the fruit and oviposit near the puncture made by a previous female. The fruits of wild Solanaceae in which the larvae of R. brncici and R. conversa develop are small in size (diameter about 1 cm), and usually, no more than two or three larvae develop within one fruit (Frías 2014). The distribution of eggs among a number of fruits prevents intraspecific larval competition. In the case of R. tomatis, females deposit eggs on the larger fruits of Lycopersicum esculentum (>5 cm in diameter) and up to 14 larvae may grow inside a single tomato fruit. A similar situation has been described in R. nova, which is associated with Solanum muricatum; females preferably lay their eggs in 6-cm-long fruits and many larvae have been observed in a single fruit (Frías 1986). These behavioral and ecological differences between these congeneric species are adaptive (Frías 2014), and they are consistent with the morphological differences between the adults and larvae of these species. Rhagoletis tomatis is allopatric with R. conversa and R. brncici and distributed in northern Chile and southern Peru (Foote 1981, Frías et al 2006, 2008, Frías 2007). Rhagoletis conversa, R. brncici, R. nova, and R. tomatis all belong to the nova group of species and are phylogenetically related (Foote 1981). However, significant differences were detected between the effects of the ODP on females of R. tomatis and its effects on R. brncici and R. conversa. These changes have evolved relatively recently (Frías 1988, Smith & Bush 2000). In both cases, the ODP facilitates a better use of resources by the larvae and contributes to the survival of the species.
The field observations on wild populations showed that infested fruits were more frequently visited by males than were uninfested fruits. In response to the ODP, females tended to be more widely distributed among their host plants and preferentially visited uninfested fruits to lay their eggs. When a female perched on a nearby fruit with a resident male, mating occurred. Morphological analysis of the couples collected in the field from their host plants showed that there is strong host fidelity in these flies. This suggests that, despite their sympatry, both species are reproductively isolated.
These species are of great economic importance because of the damage caused to fruits of cultivated plants (e.g., R. nova associated with S. muricatum). It has been demonstrated that the application of pheromone in cherry orchards reduced infestation by R. cerasi by up to 90% (Katsoyannos & Boller 1976, 1980, Boller 1981, Boller et al 1987).
The non-choice and choice tests showed that males perched for longer on infested fruit impregnated with ODP from females of their own species than from the other species. This suggests that the ODP might be involved in sexual selection and in the reproductive isolation of these sibling species. Together with other factors, such as plant architecture and odors, this could explain the host fidelity and sympatric origin which has been proposed (Frías 2001). Besides its theoretical importance, this oviposition-deterring and male-arresting pheromone could have practical application for population control of species of nova group.
References
Aluja M, Prokopy RJ (1992) Host search behavior by Rhagoletis pomonella flies: inter-tree movement patterns in response to wind-borne fruit volatiles under field conditions. Physiol Entomol 17:1–8
Aluja M, Prokopy RJ, Elkinton JS, Laurence F (1989) Nobel approach for tracking and quantifying the movement patterns of insects in three dimensions under seminatural conditions. Environ Entomol 18:1–7
Boller EF (1981) Oviposition-deterring pheromone of the European cherry fruit fly: status of research and potential applications. In: Mitchell ER (ed) Management of insect pests with semiochemicals. Plenum Publishing Corp, New York, pp 457–462
Boller EF, Shoni R, Bush GL (1987) Oviposition deterring pheromone in Rhagoletis cerasi: biological activity of a pure single compound verified in semi-field test. Entomol Exp Appl 45:17–22
Bush GL (1966) The taxonomy, cytology, and evolution of the genus Rhagoletis in North America (Diptera: Tephritidae). Bull Mus Comp Zool 134:431–562
Díaz-Fleischer F, Papaj DR, Prokopy RJ, Norrbom AL, Aluja M (2000) Evolution of fruit fly oviposition behavior. In: Aluja M, Norrbom AL (eds) Fruit flies (Tephritidae): phylogeny and evolution of behavior. CRC Press, Boca Raton, pp 811–841
Foote RH (1981) The genus Rhagoletis Loew South of the United States. United States Department of Agriculture. Tech Bull 1607:75
Frías LD (1986) Biología poblacional de Rhagoletis nova (Schiner) (Díptera :Tephritidae). Rev Chil Entomol 13:75–84
Frías LD (1988) Tiempos y modos de especiación. En: Ibcia Santibañez (ed). Manejo del tiempo en biología y sus ingeniosas maneras de estudiarlo. Facultad de Medicina, Universidad de Chile, Copyrigth, Biblioteca Nacional, pp 145-158
Frías LD (1989) Ecological and reproductive differentiation of two host races of Rhagoletis conversa (Bréthes) (Diptera Tephritidae). Acta Entomol Chilena 15:163–170
Frías LD (1995) Comportamiento de oviposición de Rhagoletis tomatis en tomate (Lycopersicum esculentum) (Díptera: Tephritidae). Acta Entomol Chilena 19:159–162
Frías LD (2001) Genetic and morphological differences of the immature stages of two races of Rhagoletis conversa (Bréthes) (Diptera Tephritidae), geographic distribution and possible sympatric origin of a new species. Rev Chil Hist Nat 74:73–90
Frías LD (2007) Especiación simpátrica y sus implicaciones genéticas y morfológicas en moscas de las fruta En: Hernández-Ortiz V (ed), Moscas de la fruta en Latinoamérica (Díptera: Tephritidae). Diversidad, biología y manejo. SyG Editores, Distrito Federal, México pp 1-26
Frías LD (2014) Modes of sympatric speciation in phytophagous insects of diptera Tephritidae and the importance of epigenetics and behavior imprinting in host fidelity. J Insect Sci Photon 115:179–194
Frías LD, Malavasi A, Morgante JS (1984) Field observations of distribution and activities of Rhagoletis conversa (Díptera Tephritidae) on two host in nature. Ann Entomol Soc Am 77:548–551
Frías LD, Hernández- Ortíz V, Vaccaro NC, Bartolucci A, Salles LA (2006) Comparative morphology of immature stages in some species of the genera Anastrepha, Toxotrypana, Rhagoletis, Bactrocera and Ceratitis (Díptera Tephritidae). In: Freidberg A (ed) Biotaxonomy of Tephritoidea. Isr J Entomol 35-36: pp: 423-457
Frías LD, Selivon D, Hernández-Ortiz V (2008) Taxonomy of immature stages: new morphological characters for Tephritidae larvae identification. In: Sugayama RL, Zucchi RA, Ovruski SM, Sivinski J (eds) Fruit flies of economic importance: from basic to applied knowledge. Proceedings 7th International Symposium on Fruit Flies of Economic Importance. Biofábrica Mosca Med, Salvador, Brazil, pp 29-44
Katsoyannos BI (1975) Oviposition-deterring, male-arresting, fruit-marking pheromone in Rhagoletis cerasi. Environ Entomol 4:801–807
Katsoyannos BI, Boller EF (1976) First field application of oviposition-deterring marking pheromone of European cherry fruit fly. Environ Entomol 5:151–152
Katsoyannos BI, Boller EF (1980) 2nd field application of oviposition-deterring pheromone of the European cherry fruit-fly, Rhagoletis cerasi (Diptera, Tephritidae). J Appl Entomol 89:278–281
Linn C, Feder JL, Nojima S, Danbroski HR, Berlocher SH, Roelofs W (2003) Fruit odor discrimination and sympatric speciation host race formation in Rhagoletis. Proc Natl Acad Sci 100:11490–11493. doi:10.1073/pnas.1635049100
Mumtaz MM, Aliniazee MT (1983) The oviposition-deterring pheromone in the western cherry fruit fly, Rhagoletis indifferens Curran (Dipt., Tephritidae). Biological properties. Z Angew Entomol 96:83–93
Norrbom AL, Carroll LE, Freidberg A (1998) Status of knowledge. In: Thompson FC (ed) Fruit fly expert identification system and systematic information database. North American Dipterists, Society Backhuys Publishers, Leiden, pp 9-47
Prokopy RJ, Bush GL (1972) Mating behavior in Rhagoletis pomonella (Diptera: Tephritidae). III. Male aggregation in response to an arrestant. Can Entomol 104: 275-283. Prokopy RJ, Reissig WH, Moericke V (1976) Marking pheromone deterring repeated oviposition in Rhagoletis flies. Entomol Exp Appl 20: 170-178
Prokopy RJ, Roitberg BD (1984) Resource partitioning. In: Bell WJ, Carde RT (eds) Chemical ecology of insects. Chapman and Hall, London, pp 301–330
Silva MA, Dias Bezerra-Silva GC, Mastrangelo TH (2012) The host marking pheromone application on the management of fruit flies. Braz Arch Biol Technol 55:835–842
Smith JJ, Bush GL (2000) Phylogeny of the subtribe Carpomyina (Trypetinae), emphasizing relationships of the genus Rhagoletis. In: Aluja M, Norrbom AL (eds) Fruit flies (Tephritidae): phylogeny and evolution of behavior. CRC Press, Boca Raton, pp 187–217
Wicker-Thomas C (2007) Pheromonal communication involved in courtship behavior in Diptera. J Insect Physiol 53:1089–1100
Acknowledgments
Thanks to Dr. Raúl Godoy-Herrera for the enriching commentaries on the manuscript, as well as the suggestions of an anonymous reviewer. This research was supported by Project Code: FIBE 07-12, Dirección de Investigación de la Universidad Metropolitana de Ciencias de la Educación (DIUMCE).
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by Stefano Colazza – Univ of Palermo
Rights and permissions
About this article

Cite this article
Frías-Lasserre, D. Effects of Female fruit-marking Pheromones on Oviposition, Mating, and Male Behavior in the Neotropical Species Rhagoletis conversa Bréthes and Rhagoletis brncici Frías (Diptera: Tephritidae). Neotrop Entomol 44, 560–564 (2015). https://doi.org/10.1007/s13744-015-0326-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13744-015-0326-1