
Abstract
High-temperature stress influences the growth and development of grapevines, and plant responses to this stress vary between laboratory and natural conditions. In the present study, the responses to high-temperature stress in four grapevine cultivars (‘Summer Black’, ‘Zuijinxiang’, ‘Hupei1#’, and ‘Shenfeng’) were studied by comparing chlorophyll a fluorescence and the levels of heat-shock protein 21 (HSP21) after exposure to control (35 °C in controlled laboratory conditions or at 37 °C in a greenhouse with naturally fluctuating temperatures) and high-temperature stress treatments (45 °C in laboratory conditions or 42 °C in the field conditions). Leaf water loss in ‘Summer Black’ was less than that in the three other cultivars after treatment at 45 °C. Some parameters (ΨEo, Wk, RCQA, and Mo) of photosystem II (PSII) activity were altered in ‘Shenfeng’, ‘Zuijinxiang’, and ‘Hupei1#’, but were unaltered in ‘Summer Black’ after treatment at 45 °C. Other parameters (maximum photochemical efficiency (Fv/Fm), performance index on absorption basis (PIABS), ΦPo, ΦEo, and HSP21 levels) were altered in ‘Summer Black’, but to a lesser extent than in the three other cultivars under high-temperature stress. Unlike ‘Summer Black’, extreme injury owing to leaf water loss to ‘Shenfeng’, ‘Zuijinxiang’, and ‘Hupei1#’ could be explained by disruption of PSII activity. Furthermore, there were the observed differences in PSII activity between in laboratory and field conditions, which could be considered as the mechanisms for high-temperature acclimation and other environment factors.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Temperature is a key parameter that controls plant growth and development, and rising temperatures are expected in the future (Karl et al. 2015). When grapevines are exposed to heat shock at 45 °C for 3 h, severe injury may occur. Grapevine leaves exposed to such high temperature exhibit chlorophyll photooxidation mediated by reactive oxygen species and severe alterations in the photosynthetic apparatus (Zha et al. 2016). In order to breed grapevine crops for better heat tolerance, we need to better understand the physiological damage that occurs during heat shock.
The photosynthetic apparatus is sensitive to high temperatures and is an ideal system for evaluating the degree of damage from high-temperature stress (Rachmilevitch et al. 2006). These evaluations are based on changes in chlorophyll a fluorescence and the OJIP transient, defined by the steps of the O, J, I, and P phases corresponding to the redox states of photosystem (PS) II and PSI and the efficiencies of electron transfer to the final electron acceptors of PSI (Strasser et al. 2000, 2004). The steps of O, J, I, and P reflect energy absorption, trapping, and electron transport (Schansker et al. 2005). The O step reflects the minimum fluorescence when the primary quinone electron acceptor (Q −A ) is fully oxidized. The P step corresponds to the state in which Q −A is fully reduced. The rise from phase O to phase J reflects a reduction in Q −A and is associated with the primary photochemical reactions of PSII. Therefore, the intermediate step, I, and the final step, P, reflect the existence of reducing plastoquinone (PQ) centers as well as different redox states of the reaction center (RC) complex in PSII (Strasser and Srivastava 1995; Strasser et al. 2000, 2004). The OJIP transient is an ideal signal and a reliable method exists for detecting and quantifying high-temperature-induced changes in the activity of PSII in plant leaves (Tóth et al. 2005; Wen et al. 2005; Strauss et al. 2006).
As a molecular chaperone, it is mainly involved in the folding, assembly and transfer of protein.
Small heat-shock proteins (sHSPs, ranging from 15 to 42 kDa), act as molecular chaperones, are mainly involved in the folding, assembling and transferring proteins (Basha et al. 2012). In plants, sHSPs are abundant and are highly expressed under high-temperature stress to protect cells against damage (Sun et al. 2002; Sun and MacRae 2005). Heat-shock protein 21 (HSP21) protects PSII against environmental stresses including high-temperature stress (Wang and Luthe 2003; Neta-Sharir et al. 2005; Kim et al. 2012).
Most studies focusing on the changes in PSII under high-temperature stress have been conducted in grapevines cultivated under laboratory conditions. Studies on the response of different grapevine cultivars to high-temperature stress in both laboratory and field conditions are needed. Accordingly, our study evaluated the activity of PSII and levels of HSP21 in four table grapevine cultivars under high-temperature stress induced in controlled laboratory conditions and in a field with naturally fluctuating temperatures in order to compare their performance and to identify heat-tolerant grapevine cultivars.
2 Materials and methods
2.1 Plant materials and treatments under field conditions
Six-year-old plants of the four table grapevine cultivars (Vitis vinifera × V. labrusca L.) ‘Summer Black’, ‘Zuijinxiang’, ‘Hupei1#’, and ‘Shenfeng’, were grown in an 8 m × 20 m greenhouse in Shanghai, China (30°89 N, 121°39 E). They received standard horticultural practices including disease and pest control. Air temperatures were measured using a HOBO U23-002 (Onset Computer Corporation, Cape Cod, MA, USA). In 2015, the average daily temperature before July 24 was below 30 °C and the highest temperature was below 40 °C (Fig. 1, Table S1). After July 24, the average daily temperature was above 30 °C until July 31st, and the highest temperature during the day was above 40 °C. On July 23, 2015, the highest daily temperature in the greenhouse was recorded at 37 °C, and on July 31, 2015, the recorded highest daily temperature was 42 °C; these were the days chosen as the control and high temperature, respectively, to measure chlorophyll a fluorescence and levels of HSP21. Fully expanded mature leaves were collected at 13:00 h for the measurements, which was the time at which the temperature reached the highest point each day. Three biological replicates with five technical replicates were used for each measurement.

Temperatures from July 21 to July 31 in 2015. July 21 is denoted by 721; all other dates are indicated in the same format. Max: highest daily temperature; Avg: average daily temperature
2.2 Plant materials and treatments under laboratory conditions
Stem cuttings were taken from grapevine cultivars of the same growth stage grown in a greenhouse in Shanghai, China (31°96 N, 121°48 E) and were rooted in a plastic container (radius: height = 15 cm: 40 cm) containing a mixture of peat moss and perlite (1:1, v/v). The stem cuttings were acclimated in a laboratory environment using a 1000-L artificial culture light box (Qianjiang Instrument and Equipment Co. Ltd., Hanzhou, China) for about 2 weeks at 25 °C, 65–70% relative humidity, and a light intensity of 200 μmol m–2s–1. Thereafter, one group was maintained at 35 °C and the other group was exposed to a 45 °C treatment for 3 h. Fully expanded mature leaves were used for the measurements of chlorophyll a fluorescence and HSP21 levels at 0 and 3 h after the treatments. These experiments were performed on three biological replicates with five technical replicates.
2.3 Measurements of chlorophyll a fluorescence
Chlorophyll a fluorescence was measured according to the method of Strasser and Srivastava (1995) with a Plant Efficiency Analyzer (Hansatech Instruments Ltd., King’s Lynn, Norfolk, UK), which provides information on the photochemical activity of PSII and the status of the PQ pool (Strauss et al. 2006). Before measurement, the leaves were dark-acclimated for 20 min. The indicators and their descriptions are presented in Table 1.
2.4 Protein immunoblot assay
Total protein was extracted with the trichloroacetic acid/acetone method (Shultz et al. 2005). The anti-HSP21 polyclonal antibody (Abcam, Bristol, UK) was provided by Wang et al. (2010). The protein immunoblot was analyzed according to the methods of Zha et al. (2016).
2.5 Statistical analysis
Data were processed with Microsoft Excel 2007. Differences were evaluated using two-sample t-tests assuming equal variances at p < 0.05 or p < 0.01.
3 Results
In the field, daily temperatures reached 40 °C from July 25 to July 31 for 1–7 h each day, and the highest temperature was recorded on July 31 at 44.3 °C (Table S1). The temperature treatments in the laboratory conditions (35 and 45 °C) were set for further verification of the results of the field treatments. In the laboratory treatment, the ‘Zuijinxiang’, ‘Hupei1#’, and ‘Shenfeng’ plants were dehydrated more significantly at 45 °C for 3 h than those at 35 °C. However, only young leaves and stems of ‘Summer Black’ were dehydrated under the 45 °C treatment (Fig. 2).

Phenotypes of the four grapevine cultivars during the different laboratory heat treatments at 6 h. ‘Summer Black’ at the 35 °C treatment (a); ‘Summer Black’ at the 45 °C treatment (b); ‘Zuijinxiang’ at the 35 °C treatment (c); ‘Zuijinxiang’ at the 45 °C treatment (d); ‘Hupei1#’ at the 35 °C treatment (e); ‘Hupei1#’ at the 45 °C treatment (f); ‘Shenfeng’ at the 35 °C treatment (g); ‘Shenfeng’ at the 45 °C treatment (h)
The maximum photochemical efficiency (Fv/Fm) and performance index on absorption basis (PIABS) were significantly decreased under the high temperature treatments (45 °C/42 °C) compared with the control temperatures (35 °C/37 °C) for ‘Zuijinxiang’, ‘Hupei1#’, and ‘Shenfeng’ (Fig. 3). The values of Fv/Fm and PIABS for ‘Summer Black’ decreased after treatment at 45 °C, but not at 42 °C compared with the controls (Fig. 3).

Fv/Fm and PIABS of PSII in the leaves of grapevines. The field conditions on July 23 and July 31, 2015 are denoted by “37” and “42 °C”, respectively. SB: ‘Summer Black’; ZJX: ‘Zuijinxiang’; HP1#: ‘Hupei1#’; SF: ‘Shenfeng’. Significant differences among the different temperature treatments at the same time point are indicated by *(p < 0.05) or **(p < 0.01). Vertical bars represent standard errors of the means from three replications
PIABS is a combination of the indices of three parameters: (1) ΨEo: the probability that a trapped exciton moves an electron into the electron transport chain beyond Q −A , (2) ΦPo: the maximum quantum yield of primary photochemistry, and (3) ΦEo: the efficiency with which a trapped exciton can move an electron into the electron transport chain. PIABS decreased following the 45 °C treatment in ‘Zuijinxiang’, ‘Hupei1#’, and ‘Shenfeng’. The ΨEo of ‘Summer Black’ did not differ significantly between the laboratory treatment at 45 °C and the field treatment at 42 °C (Fig. 4). Electron transport decreased as the temperature increased. Additionally, the values of ΨEo, ΦPo, and ΦEo decreased after treatment at 42 °C for the ‘Zuijinxiang’ cultivar throughout the experiment (Fig. 4).

ΦPo and electron acceptor parameters (ΨEo and ΦEo) of PSII in the leaves of grapevines. The field conditions on July 23 and July 31, 2015 are denoted by “37” and “42 °C”, respectively. SB: ‘Summer Black’; ZJX: ‘Zuijinxiang’; HP1#: ‘Hupei1#’; SF: ‘Shenfeng’. Significant differences among the different temperature treatments at the same time point are indicated by *(p < 0.05) or **(p < 0.01). Vertical bars represent standard errors of the means from three replications
No changes were found in the amplitude of phase K (Wk) in ‘Summer Black’ and ‘Zuijinxiang’ (Fig. 5); however, the Wk was significantly increased after treatment at 45 or 42 °C in ‘Hupei 1#’ and ‘Shenfeng’. These data indicated that the oxygen-evolving complexes of ‘Summer Black’ and ‘Zuijinxiang’ were not damaged under the high-temperature treatments. However, these high-temperature treatments altered the density of the Q −A reducing reaction centers of PSII (RCQA) in the three other grapevine cultivars, but did not influence the RCQA of ‘Summer Black’.

Donor side indicator (Wk) and reaction center indicator (RCQA) of PSII in the leaves of grapevines. The field conditions on July 23 and July 31, 2015 are denoted by “37” and “42 °C”, respectively. SB: ‘Summer Black’; ZJX: ‘Zuijinxiang’; HP1#: ‘Hupei1#’; SF: ‘Shenfeng’. Significant differences among the different temperature treatments at the same time point are indicated by *(p < 0.05) or **(p < 0.01). Vertical bars represent standard errors of the means from three replications
‘Summer Black’ did not show significant alteration of the approximate initial slope of the fluorescence transient (Mo) after treatment at 45 or 42 °C, but changes in Mo were observed in ‘Shenfeng’, ‘Zuijinxiang’, and ‘Hupei1#’ (Fig. 6). The redox state of PSI (δRo) for the four grapevine cultivars significantly increased after the laboratory treatment at 45 °C; however, there were no changes in δRo after the field treatment at 42 °C compared with δRo after treatment at 37 °C (Fig. 6).

Acceptor side parameters Mo and δRo in the leaves of grapevines. The field conditions on July 23 and July 31, 2015 are denoted by “37” and “42 °C”, respectively. SB: ‘Summer Black’; ZJX: ‘Zuijinxiang’; HP1#: ‘Hupei1#’; SF: ‘Shenfeng’. Significant differences among the different temperature treatments at the same time point are indicated by *(p < 0.05) or **(p < 0.01). Vertical bars represent standard errors of the means from three replications
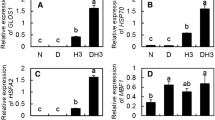
As shown in Fig. 7, HSP21 levels, representing the immune signal, increased in ‘Summer Black’, but decreased in ‘Shenfeng’, ‘Zuijinxiang’, and ‘Hupei1#’ after treatment for 3 h at 45 °C. In the field treatment, this signal increased in ‘Summer Black’ and ‘Hupei 1#’, but decreased in ‘Shenfeng’ and ‘Zuijinxiang’ after treatment at 42 °C as compared with that after treatment at 37 °C.

HSP21 expression levels in the leaves of grapevine. The field conditions on July 23 and July 31, 2015 are denoted by “37” and “42 °C”, respectively
4 Discussion
In the present study, all four cultivars suffered various degrees of leaf blade dehydration under the 45 °C treatment for 3 h. ‘Summer Black’ showed slight dehydration, whereas ‘Shenfeng’, ‘Zuijinxiang’, and ‘Hupei1#’ showed marked dehydration. Thus, ‘Summer Black’ may be more tolerant to high-temperature stress than the other cultivars.
The OJIP transients were used to evaluate several biophysical reactions that allow us to quantify the function of PSII. Changes in OJIP transients caused by high temperature stress have been used to study heat tolerance in apples and beans (Chen and Cheng 2009; Stefanov et al. 2011). Fv/Fm, the maximum quantum yield of primary PSII photochemistry, has been used to determine the health of plants in many studies (Stirbet and Govindjee 2011; Chen et al. 2013; Murchie and Lawson 2013). Our results of the Fv/Fm (ranging from 0.78 to 0.84 for the four grapevine cultivars) showed that the PSII activity were at the normal levels,. High-temperature stress decreased the Fv/Fm in previous studies (Bjorkman and Demmig 1987; Murchie and Lawson 2013), and in our study, the Fv/Fm decreased markedly after treatment at 45 °C in all four grapevine cultivars, with ‘Summer Black’ showing the smallest changes. Moreover, only ‘Summer Black’ had no obvious changes in the Fv/Fm after treatment at 42 °C, whereas the Fv/Fm of the other cultivars decreased significantly, indicating that ‘Summer Black’ was more tolerant to high temperatures and ‘Zuijinxiang’, ‘Hupei 1#’, and ‘Shenfeng’ were heat-sensitive.
PIABS, which is associated with ΨEo, ΦPo, and ΦEo, provides useful information regarding plant health (Perboni et al. 2012; Singh and Sarkar 2014). In an earlier study, the ratios of ΨEo, ΦPo, ΦEo, Fv/Fm, and PIABS of the co-dominant species Quercus ilex and Phillyrea latifolia were decreased under the combined effect of heat and drought stresses (Ogaya et al. 2011). In our study, the reductions in some of these parameters were more dramatic in the high temperature-sensitive cultivars (‘Zuijinxiang’, ‘Hupei 1#’, and ‘Shenfeng’) than in the heat-tolerant grapevine cultivar (‘Summer Black’) under high-temperature stress.
In our study, Wk (donor side parameter of PSII), RCQA (reaction center parameter of PSII), and Mo and δRo (acceptor side parameters of PSII) of the grapevine cultivars were altered under the high-temperature treatments, consistent with the findings of a previous study (Wang et al. 2010). Notably, the degree of those changes was smallest for ‘Summer Black’, further supporting that ‘Summer Black’ is more heat tolerant with regard to the stability of PSII activity.
HSP21 stabilizes proteins and prevents protein aggregation, thereby functioning in the protection of PSII (Neta-Sharir et al. 2005). In the present study, HSP21 levels increased in ‘Summer Black’ and decreased in the three other cultivars at 45 °C, indicating that the leaves of ‘Shenfeng’, ‘Zuijinxiang’, and ‘Hupei1#’ were heavily damaged due to high temperature stress, and that they were unable to effectively synthesize proteins to maintain normal physiological processes.
The patterns of variation in the chlorophyll a fluorescence parameters and HSP21 levels between the laboratory and field conditions were similar, which suggested that the major driver of the response of PSII in grapevines is temperature. The differences among the four grapevine cultivars might be due to differences in their genetic backgrounds. Additionally, the extreme injury to ‘Shenfeng’, ‘Zuijinxiang’, and ‘Hupei1#’ cultivars could be explained by disruption of the activity of PSII during high-temperature stress.
Different behaviors of PSII and HSP21 levels in the four grapevine cultivars were seen in the laboratory and field conditions. Accordingly, the lower ΦPo value occurring at 45 °C was considered as a positive adaptation for the downregulation of photosynthetic excitation pressure (Raven 2011). However, these changes did not occur after the field treatment at 42 °C, suggesting the involvement of other factors. In the field, temperatures gradually increased, which may have enhanced the ability of the plants to adapt to the higher temperatures (42 °C), thereby, improving the resistance to heat stress. This high-temperature acclimation theory was consistent with that of previous reports (Havaux 1993; Horowitz 2002; Larkindale and Vierling 2008; Hasanuzzaman et al. 2013). Additionally, the differences in performance between the different cultivars may be attributed to their inherent vigor. Other environmental factors may have attributed to the differences in some parameters, including HSP21 levels, between the laboratory and field conditions. Although it is difficult to simulate natural high-temperature stress, our findings provide experimental and theoretical guidance for the production of heat-tolerant cultivars.
Abbreviations
- PIABS:
-
Performance index on absorption basis
- PQ:
-
Plastoquinone
- PS:
-
Photosystem
- RC:
-
Reaction centers
- QA:
-
Quinione electron acceptor
- HSP:
-
Heat shock protein
References
Basha E, O’Neill H, Vierling E (2012) Small heat shock proteins and α-crystallins: dynamic proteins with flexible functions. Trends Biochem Sci 37:106–117
Bjorkman O, Demmig B (1987) Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170:489–504
Chen LS, Cheng L (2009) Photosystem II is more tolerant to high temperature in apple (Malus domestica Borkh.) leaves than in fruit peel. Photosynthesis 47:112–120
Chen Y, Zhou Y, Yin TF, Liu CX, Luo FL (2013) The invasive wetland plant Alternanthera philoxeroides shows a higher tolerance to water logging than its native congener Alternanthera sessilis. PLoS ONE 8:e81456
Hasanuzzaman M, Nahar K, Alam MM, Roychowdhury R, Fujita M (2013) Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Intern J Mol Sci 14:9643–9684
Havaux M (1993) Rapid photosynthetic adaptation to heat stress triggered in potatoleaves by moderately elevated temperatures. Plant Cell Environ 16:461–467
Horowitz M (2002) From molecular and cellular to integrative heat defense during exposure to chronic heat. Comp Biochem Physiol A 131:475–483
Karl TR, Arguez A, Huang B, Lawrimore JH, McMahon JR, Menne MJ, Peterson TC, Vose RS, Zhang HM (2015) Possible artifacts of data biases in the recent global surface warming hiatus. Science 348:1469–1472
Kim KH, Alam I, Kim YG, Sharmin SA, Lee KW, Lee SH, Lee BH (2012) Overexpression of a chloroplast-localized small heat shock protein OsHSP26 confers enhanced tolerance against oxidative and heat stresses in tall fescue. Biotechnol Lett 34:371–377
Larkindale J, Vierling E (2008) Core genome responses involved in acclimation to high temperature. Plant Physiol 146:748–761
Murchie EH, Lawson T (2013) Chlorophyll fluorescence analysis: a guide to good practice and understanding some new applications. J Exp Bot 64:3983–3998
Neta-Sharir I, Isaacson T, Lurie S, Weiss D (2005) Dual role for tomato heat shock protein 21: protecting photosystem II from oxidative stress and promoting color changes during fruit maturation. Plant Cell 17:1829–1838
Ogaya R, Penuelas J, Asensio D, Llusià J (2011) Chlorophyll fluorescence responses to temperature and water availability in two co-dominant Mediterranean shrub and tree species in a long-term field experiment simulating climate change. Environ Exp Bot 73:89–93
Perboni AT, Cassol D, Silva F, Sliva D, Bacarin M (2012) Chlorophyll a fluorescence study revealing effects of flooding in canola hybrids. Biologia 67:338–346
Rachmilevitch S, Huang B, Lambers H (2006) Assimilation and allocation of carbon and nitrogen of thermal and nonthermal Agrostis species in response to high soil temperature. New Phytol 170:479–490
Raven JA (2011) The cost of photoinhibition. Physiol Plant 142:87–104
Schansker G, Tóth SZ, Strasser RJ (2005) Methylviologen and dibromothymoquinone treatments of pea leaves reveal the role of photosystem I in the Chl a fluorescence rise OJIP. Biochim Biophys Acta (BBA)-Bioenerg 1706:250–261
Shultz RW, Settlage SB, Hanley-Bowdoin L, Thompson WF (2005) A trichloroacetic acid-acetone method greatly reduces infrared autofluorescence of protein extracts from plant tissue. Plant Mol Biol Rep 23:405–409
Singh DP, Sarkar RK (2014) Distinction and characterization of salinity tolerant and sensitive rice cultivars as probed by the chlorophyll fluorescence characteristics and growth parameters. Funct Plant Biol 41:727–736
Stefanov D, Petkova V, Denev ID (2011) Screening for heat tolerance in common bean (Phaseolus vulgaris L.) lines and cultivars using JIP-test. Sci Hortic 128:1–6
Stirbet A, Govindjee B (2011) On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: basics and applications of the OJIP fluorescence transient. J Photochem Photobiol 104:236–357
Strasser RJ, Srivastava A (1995) Polyphasic chlorophyll a fluorescence transients in plants and cyanobacteria. Photochem Photobiol 61:37–42
Strasser RJ, Srivastava A, Tsimilli-Michael M (2000) The fluorescence transient as a tool to characterize and screen photosynthetic samples. In: Yunus M, Pathre U, Mohanty P (eds) Probing photosynthesis: mechanisms, regulation and adaptation. Taylor and Francis, London, pp 445–483
Strasser RJ, Tsimilli-Micheal M, Srivastava A (2004) Analysis of the chlorophyll a fluorescence transient. In: Papageorgiou GC, Govindjee (eds) Chlorophyll a fluorescence. Advanced in photosynthesis and respiration, vol 19. Springer, Dordrecht, pp 371–376
Strauss AJ, Krüger GHJ, Strasser RJ (2006) Ranking of dark chilling tolerance in soybean genotypes probed by the chlorophyll a fluorescence transient OJIP. Environ Exp Bot 56:147–157
Sun Y, MacRae TH (2005) Small heat shock proteins: molecular structure and chaperone function. Cell Mol Life Sci 62:2460–2476
Sun W, Van Montagu M, Verbruggen N (2002) Small heat shock proteins and stress tolerance in plants. Biochem Bioph Acta 1577:1–9
Tóth SZ, Schansker G, Kissimon J, Kovács L, Strasser RJ (2005) Biophysical studies of photosystem II-related recovery processes after a heat pulse in barley seedlings (Hordeum vulgare L.). J Plant Physiol 162:181–194
Wang D, Luthe DS (2003) Heat sensitivity in a bentgrass variant. Failure to accumulate a chloroplast heat shock protein isoform implicated in heat tolerance. Plant Physiol 133:319–327
Wang LJ, Fan L, Loescher W, Duan W, Liu GJ, Cheng JS, Li SH (2010) Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Boil 10:1
Wen XG, Qiu NW, Lu QT, Lu C (2005) Enhanced thermotolerance of photosystem II in salt-adapted plants of the halophyte Artemisia anethifolia. Planta 220:486–497
Zha Q, Xi XJ, Jiang AL, Tian YH (2016) Changes in the protective mechanism of photosystem II and molecular regulation in response to high temperature stress in grapevines. Plant Physiol Biochem 101:43–53
Acknowledgements
This research was partially funded by the Youth Talent Development Plan of Shanghai Municipal Agricultural System, China [Grant Number 20170103]; and the Modern Agricultural Industry Technology System (Grape) [Grant Number CARS-29-10].
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Table S1
Daily air temperature (°C) from July 23 to July 31, 2015 (DOC 61 kb)
Rights and permissions
About this article

Cite this article
Zha, Q., Xi, X., Jiang, A. et al. Comparison of the activities of photosystem II of four table grapevine cultivars during high-temperature stress. Hortic. Environ. Biotechnol. 59, 363–371 (2018). https://doi.org/10.1007/s13580-018-0041-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13580-018-0041-z