
Abstract
To develop a new pest control device for greenhouses, we verified the effect of the noncontact force generated by ultrasonic transducers on two small insect pests: the sweet potato whitefly Bemisia tabaci Gennadius and the cotton aphid Aphis gossypii Glover. First, we examined their dispersion behavior in response to the device, and the dispersion rate of whiteflies from leaves was approximately 60% for the 60 s treatment with a modulation frequency of 1–240 Hz, which was comparable to that under the air blowing treatment of 5.0 m/s. In contrast, the dispersion rate of aphids from leaves was at most 25% for the 60 s treatment. Next, the flight behavior of whiteflies under 30 s of vibration treatment was analyzed; 40–70% of individuals flew within 5 s, and an average of 86% of dispersed adults flew downward. Moreover, we determined the suppression of whitefly fecundity by the device, and the number of eggs laid over 2 days was significantly decreased by 4 h/days treatment. These results suggest that this device also suppresses the fecundity of whiteflies. This device would be applicable for insect pest management following further studies on the optimization of treatments and the efficient collection of dispersed insects.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
For several vegetable and flower crops, large-scale greenhouses of more than 1 ha of floor area have emerged as a main type of protected agricultural system in Japan (Yamada 2008). In these greenhouses, meteorological factors, including air temperature, relative humidity, solar radiation, and vapor pressure deficit, are generally automatically recorded and controlled for 24 h a day throughout the year (Meng et al. 2017); thus, the productivity of vegetables or flowers in these systems is high because it is possible to cultivate them year-round. On the other hand, the risk of crop disease and insect pest spread is extremely high in greenhouses once introduced. In the case of tomato greenhouse farming, the sweet potato whitefly Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) is one of the most serious pests because this species is a vector of tomato yellow leaf curl virus (TYLCV) and tomato chlorosis virus (ToCV), both of which cause severe damage to tomato plants (Horowitz et al. 2007; Orfanidou et al. 2016). Thrips, spider mites and aphids are also among the most serious insect pests in greenhouses, and chemical pesticides are the conventional and main way of controlling these pests, while several pest species have developed insecticide resistance (Naveen et al. 2017; Ghosh et al. 2020; Osakabe et al. 2009; Edwards et al. 2008). In addition, in greenhouse tomato production in Japan, the buff-tailed bumblebee Bombus terrestris (Hymenoptera: Apidae) was introduced for pollination but was later designated by Japan as an invasive alien species in 2006; now, the use of alternative native bees has been promoted (Morimyo 2017). Moreover, the phenomenon of bees suddenly disappearing, i.e., “colony collapse disorder (CCD)”, has occurred in many parts of the world since the late 2000s and become a major problem (Rosner 2013; Klatt et al. 2014); thus, it is possible that it will be difficult to use bees for pollination in the future.
To develop a sustainable method for greenhouse farming independent of chemical pesticides, bees and chemical plant growth regulators for pollination, several studies on physical pest control and artificial pollination using vibration have been conducted. B. tabaci adults communicate via substrate-borne vibrations (Kanmiya 2006, 2011), and substrate-borne vibrations artificially generated on tomato plants infested with B. tabaci adults by a vibration exciter decreased the population density of nymphs on leaves (Yanagisawa et al. 2021). Ben-Ari and Inbar (2014) examined the reaction of pea aphid Acyrthosiphon pisum (Hemiptera: Aphididae) adults to several cues (mammalian breath, vibration, wind, and visual cues), and aphids were able to sense these cues and dropped from leaves to avoid the danger of being ingested by mammalian herbivores. Previous studies have also reported that noncontact vibration generated by ultrasonic transducers can achieve artificial pollination of strawberries and tomatoes (Shimizu and Sato 2018; Shimizu et al. 2015; Kobayashi 2013). Here, we aimed to develop a new physical pest control device using vibration and verified the effect of the noncontact force generated by ultrasonic transducers, which is the same device described by Shimizu et al. (2015) for artificial pollination, on two small insect pests: B. tabaci (Q-biotype) and the cotton aphid Aphis gossypii Glover (Homoptera: Aphididae), which is one of the most harmful pests to several vegetables and crops.
Materials and methods
A device for controlling insect pests using noncontact force
The same ultrasonic device as the artificial pollination system developed by Shimizu et al. (2015) was used: sound waves can exert forces on objects due to acoustic radiation pressure, and the size of the focal point of the sound wave decreases as the frequency increases. This phenomenon is attributed to the nonlinear acoustics of acoustic radiation pressure, and although the acoustic power is conventionally called “pressure”, it works in a specific direction, unlike atmospheric pressure (Shimizu et al. 2015). Utilizing these properties, Hoshi et al. (2014) developed an ultrasonic focusing device, and then this device was applied to artificial pollination (Shimizu et al. 2015; Shimizu and Sato 2018). The specifications of the ultrasonic focusing device, which were written in Shimizu et al. (2015), are shown in Table S1. The acoustic radiation pressure generated from a single ultrasonic transducer is weak; therefore, 285 ultrasonic transducers (T4010A1, Nippon Ceramic Co., Ltd.; 40 kHz resonance frequency, 1 cm diameter) were arrayed in a 170 × 170 mm square to generate a force of at most 16 mN. The phase of each transducer was appropriately controlled and focused on one point in space. The focal diameter was proportional to the focal length; for example, the focal diameter was 20 mm for a focal length of 20 cm. The focus could be moved by specifying spatial coordinates in 0.5 mm increments; for example, the focal point could be randomly moved within a 100 × 100 mm 2D area for 10 ms per position. The ultrasonic transducer was driven by a 40 kHz square wave obtained by amplifying the digital output of a field-programmable gate array (FPGA) using a driver, and the intensity of the acoustic radiation pressure was controlled by applying pulse width modulation (PWM) to the square wave. In addition, the pressure could be modulated by square waves whose frequency and duty ratio range from 1 to 1023 Hz and 50%; therefore, the ultrasonic carrier frequency of 40 kHz was modulated with the natural frequency of the object on a focal point. The rise time of the ultrasonic transducer was 1 ms; thus, the focal point could be moved from a target position to the next position within 1 ms. In this research, the acting force was the maximum value of 16 mN and modulated into vibration stimulation by ON/OFF control of the ultrasound treatment.
Insect materials
B. tabaci (Q biotype) individuals were collected from green peppers (Capsicum annuum L.) grown in Miyazaki, Japan in 2004 and reared for more than six generations on cabbage (Brassica oleracea L. var. capitata). Prior to the experiment, they were reared for more than six generations on kidney beans (Phaseolus vulgaris L.) in a cylindrical plastic cage (height: 330 mm, diameter: 135 mm) with nylon mesh in a growth chamber at 23 ± 2 °C and 60 ± 10% relative humidity with a photoperiod of 14L10D.
A. gossypii individuals were collected from cucumbers (Cucumis sativus) grown in Tsu, Mie, Japan in May 2004, and prior to the experiment, they were reared for more than six generations on eggplants (Solanum melongena) in an acrylic cage (400 × 360 × 250 mm) with nylon mesh in a growth chamber at 20 ± 1 °C and 60 ± 10% relative humidity with a photoperiod of 14L10D.
Plant materials
For the B. tabaci experiment, kidney bean seedlings (cv ‘Naga-uzura’) in a plastic pot (top diameter: 45 mm, height: 60 mm) were raised in a growth chamber with the temperature and photoperiod set to 24.5 °C and 14L10D, respectively, for one week. For the A. gossypii experiment, eggplant seedlings (cv ‘Senryo’) in a plastic pot (top diameter: 45 mm, height: 60 mm) were raised in a glasshouse for approximately 1 month. For both experiments, the same plant species used for rearing insects were used.
Dispersion experiment
For the B. tabaci experiment, 4–6 kidney bean seedlings and 200–300 adults (1–5 days after emergence; the sex ratio was approximately 1:1) were introduced into an acrylic cage (300 × 280 × 250 mm) with nylon mesh. After a couple of hours, a kidney bean leaf (leaf size: within 100 × 100 mm square) with 20–50 adults was cut off and attached to the inside wall of an acrylic cage (500 × 600 × 450 mm) with adhesive tape (Table S2). The ultrasonic device was placed 200 mm apart from the attached leaf (Fig. 1a). The focal distance of the ultrasonic device was set to 200 mm, and the ultrasonic carrier frequency of 40 kHz was modulated with frequencies of 1, 10, 30, 60, 120, 240, 480 and 1000 Hz. Here, the focal distance was set to 200 mm based on a previous study (Hoshi et al. 2014). The focal point was set to move randomly within a 100 × 100 mm area for 10 ms per position, and the period of movement of the focal point was set to 10 or 60 s. As a comparison group, air blowing treatments with three levels of wind velocity (10.5 m/s, 5.0 m/s and 2.6 m/s) generated by an air pump (KM-8EEA, E.M.P-Japan Ltd.) were applied to B. tabaci-infested leaves for 10 or 60 s. Wind velocity was measured with an air speedometer (CW-60, CUSTOM Co.). The experiment was conducted at room temperature and relative humidity of 24 ± 1 °C and 50 ± 10%, respectively, under a fluorescent lamp from 14:00 to 17:00. Pictures of a whole leaf before and after treatment were taken using a digital camera (STYLUS TG-4 Tough, Olympus Co.), and the number of whitefly adults was counted with ImageJ v.1.50i software (NIH).

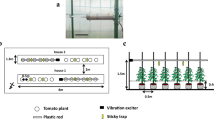
Images of the dispersion and suppression of fecundity of insect pest experiments. a An image of the B. tabaci or A. gossypii dispersion experiment; b an image of the B. tabaci fecundity suppression experiment
For the A. gossypii experiment, approximately 2–4 eggplant seedlings and 100–200 winged or wingless adults (1–3 d after emergence) were introduced into an acrylic cage (300 × 280 × 250 mm) with nylon mesh. After a couple of hours, an eggplant leaf (leaf size: within 100 × 100 mm square) with 20–40 adults was cut off and attached in the same way as in the experiment with B. tabaci (Fig. 1a; Table S2). The focal distance of the ultrasonic device was set to 200 mm, and the ultrasonic carrier frequency of 40 kHz was modulated with frequencies of 1, 10, 30, 60, 120, 240 and 1000 Hz. Here, the focal distance was set to 200 mm based on a previous study (Hoshi et al. 2014). The focal point was set to move randomly within a 100 × 100 mm area for 10 ms per position, and the period of movement of the focal point was set to 60 s. The experiment was conducted under the same light, temperature, and relative humidity conditions as the B. tabaci experiment from 14:00 to 17:00. The numbers of aphid adults on the leaves before and after the treatments were counted in the same way as in the B. tabaci experiment.
Flight behavior analysis
Similar to the B. tabaci dispersion experiment, a kidney bean leaf with B. tabaci adults was attached to the inner wall of an acrylic cage (500 × 600 × 450 mm) with adhesive tape, and the ultrasonic device was placed 200 mm apart from the leaf (Table S2). The focal distance of the ultrasonic focusing device was set to 200 mm, and the ultrasonic carrier frequency of 40 kHz was modulated with frequencies of 1, 10, 30, 60, 120, 240, 480 and 1000 Hz. Here, the focal distance was set to 200 mm based on a previous study (Hoshi et al. 2014). The focal point was set to move randomly within a 100 × 100 mm area for 10 ms per position, and the period of movement of the focal point was set to 30 s. The experiment was conducted under the same light, temperature, and relative humidity conditions as above from 14:00 to 17:00. A high-speed digital camera (EX-100PRO, Casio Computer Co., Ltd) was set next to the ultrasonic focusing device, and the flight behavior was recorded for 30 s at 240 fps (Fig. 1a). The period from the beginning of irradiation to take off from the leaf was measured, and the flight direction and the behavior before taking off were observed by using Photron Fastcam Analysis v.1.4.4 software (Photron Ltd.). The flight direction was set to three levels: upward, downward and horizontal (at an elevation angle above 30°, at a depression angle above 30° and from an elevation angle of 30° to a depression angle of 30°, respectively), and the behavior before taking off was categorized into two motions: walking or freezing.
Reproductive behavior suppression experiment
Ten pairs of prenuptial B. tabaci adults within 2 days after emergence and a kidney bean seedling with two cotyledons were introduced into an acrylic cage (220 × 150 × 250 mm) with nylon mesh, of which the whole surface of the bottom side was covered with mesh at 9:00 (Table S2). After a few hours, the cage was put over the ultrasonic focusing device, which was laid down to face upwards (Fig. 1b). The focal distance of the ultrasonic focusing device was set to the distance from the transducer array surface to the underside of the leaf surface, and the ultrasonic carrier frequency of 40 kHz was modulated with a frequency of 30 Hz. The focal point was set to move randomly among 200 × 200-mm discrete positions for 10 ms per position, and the period of movement of the focal point was set to 2 (11:30–13:30) or 4 h (11:30–15:30). On the next day at 11:30, the same treatment applied the previous day was started (Fig. S1), and just after the treatment, the numbers of adults on leaves and pairs in courtship behavior were visually counted, and all the eggs laid on leaves were counted under a stereoscopic microscope. Here, courtship behavior was defined as a sequence of stereotyped courtship behavior from “in parallel position” to “wing hanging in copula” described in previous studies (Kanmiya 1996, 2006). The experiment was conducted under the same 14L10D light, temperature, and relative humidity conditions as above.
Data analysis
For all the experiments, a generalized linear model (GLM) with a Poisson error distribution and log link function was used to discern significant differences among treatments. For the dispersion experiments with B. tabaci and A. gossypii, following GLM statistical analysis, a further Dunnett comparison test was performed to scrutinize differences between the control and other treatments. The dispersion rate of A. gossypii adults under each modulation frequency condition was also compared between winged and wingless adults by GLM with a Poisson error distribution and log link function. The dispersion rate after 60 s of treatment was also compared among B. tabaci adults and winged and wingless adults of A. gossypii by GLM with a Poisson error distribution and log link function. For the experiment of flight behavior of B. tabaci, the rate of flying or remaining adults was compared among modulation frequencies by the likelihood ratio chi-square test, and the histograms of duration time to take off the leaf were analyzed with a negative binomial model of discrete distribution, i.e., gamma Poisson distribution (Zuur et al. 2009). This distribution is a discrete probability distribution that models the number of successes in a sequence of independent and identically distributed Bernoulli trials before a specified and nonrandom number of failures occurs (Bliss and Fisher 1953); thus, it was appropriate for fitting this model in this case. For the experiment on the B. tabaci reproductive behavior suppression effect, the numbers of pairs in courtship behavior and laid eggs and the residual rates of adults on leaves were compared among varieties by Dunnett comparison test. The residual rate was defined as the number of adults on leaves after 2 days of treatment/the number of adults introduced into the test cage. All statistical analyses were performed with JMP v.13.2.1 software (SAS Institute, Cary, North Carolina, USA).
Results
Dispersion effect of the ultrasonic focusing device on B. tabaci
For the 10 s experiment, there was a significant difference in the dispersion rate of B. tabaci adults among the wind velocity, modulation frequency and control treatments (Fig. 2a; GLM, χ2 = 2751.429, df = 11, p < 0.0001). The dispersion rate was significantly higher than that under no treatment when the noncontact force was created using an ultrasonic focusing device or air blowing at a velocity of 5.0 m/s or greater (Fig. 2a; Dunnett’s test, p < 0.05 at 1000 Hz and p < 0.0001 at the other treatments) and not significantly different at a velocity of 2.6 m/s (Fig. 2a; Dunnett’s test; p = 1.00). For the 60 s experiment, there was also a significant difference in the dispersion rate of B. tabaci among the wind velocity, modulation frequency and control treatments (Fig. 2b; GLM, χ2 = 2200.878, df = 11, p < 0.0001). The dispersion rate of B. tabaci was significantly higher than that under no treatment when the noncontact force was created using an ultrasonic focusing device or air blowing at a velocity of 5.0 m/s or greater (Fig. 2b; Dunnett’s test, p < 0.0001) and not significantly different at a velocity of 2.6 m/s (Fig. 2b; Dunnett’s test, p = 1.00). The dispersion rate was highest under the 10.5 m/s air blowing conditions for both the 10 and 60 s treatments (78.8% and 88.6%, respectively).

Dispersion rate of B. tabaci adults from a kidney bean leaf after a 10 s and b 60 s of treatment with noncontact vibration or wind power. The vertical bars indicate SE (n = 16 ± 6). Dunnett’s comparison test was significant for comparisons with the corresponding control group values at *p < 0.05 and **p < 0.0001. NS indicates nonsignificant (p > 0.05)
Dispersion effect of the ultrasonic focusing device on A. gossypii
There were significant differences in the dispersion rate of winged or wingless A. gossypii adults among the modulation frequency and control treatments for 60 s (Fig. 3; GLM, χ2 = 743.89, df = 7, p < 0.0001 for winged adults and χ2 = 606.51, df = 7, p < 0.0001 for wingless adults). Comparing the effects of modulation frequencies on winged adults to the control condition, the dispersion rate from eggplant leaves was significantly higher under 1, 10, 30 and 240 Hz conditions (Fig. 3b; Dunnett’s test, p < 0.05 at 1, 10 and 240 Hz and p < 0.0001 at 30 Hz) compared to the control and not significantly different on 60, 120 and 1000 Hz conditions (Fig. 3b; Dunnett’s test, p = 0.09, p = 0.21 and p = 1.00, respectively). In the case of wingless adults, the dispersion rate was significantly higher under the 30 and 240 Hz conditions (Fig. 3a; Dunnett’s test, p < 0.05 at 240 Hz and p < 0.0001 at 30 Hz) and not significantly different under the 1, 10, 60, 120 and 1000 Hz conditions (Fig. 3a; Dunnett’s test, p = 0.30, p = 0.19, p = 0.77, p = 0.24 and p = 1.00, respectively).

Dispersion rate of a wingless and b winged A. gossypii adults after 60 s of treatment with noncontact vibration or wind power. The vertical bars indicate SE (n = 17 ± 6). Dunnett’s comparison test was significant for comparisons with the corresponding control group values at *p < 0.05 and **p < 0.0001. NS indicates nonsignificant (p > 0.05)
The dispersion rate of adults was compared between winged and wingless adults, and there was a significant difference (Fig. 3; GLM, χ2 = 565.24, df = 1, p < 0.0001). Compared with B. tabaci, however, the dispersion rate of winged or wingless adults of A. gossypii was significantly lower compared to B. tabaci after 60 s treatments under 1 Hz, 10 Hz, 30 Hz, 60 Hz, 120 Hz, 240 Hz and 1000 Hz (Figs. 2, 3; Dunnett’s test, p < 0.0001).
Flight behavior of B. tabaci after treatment with modulation frequencies
The effects of irradiation on time on the leaf and flight behavior of B. tabaci adults were analyzed. The flight rate after 30 s of ultrasonic focusing device treatment was more than 70% under all modulation frequencies, and the rate of taking off or remaining on the leaf was not significantly different among modulation frequencies (Fig. 4; likelihood chi-square test, df = 6, χ2 = 8.3361, p = 0.2145). Out of all the modulation frequencies, the width of the histogram was set to 1 s or 0.125 s under 1, 30, 60, 120, 240, 480 and 1000 Hz conditions for optimization of the fitting of the models. The histogram of B. tabaci adults that took 1–30 s to take off the leaf was fitted to a gamma Poisson distribution under every modulation frequency condition (Fig. S2), and approximately 60–90% of individuals flew within 5 s regardless of modulation frequency (Fig. 4).

Behavioral analysis of B. tabaci adults after 30 s of noncontact vibration treatment. Percentage of whitefly adults that took less than 1 s, 1–5 s, 5–10 s, 10–20 s or 20–30 s to fly away or remained on the leaf (left) and percentage of dispersed whitefly adults that took less than 1 s, 1–5 s, 5–10 s, 10–20 s or 20–30 s to fly away (right)
On average, among the seven levels of modulation frequencies, 86%, 13% and 1% of dispersed adults flew downward, horizontal, and upward, respectively, and the rate of B. tabaci adults flying downward, horizontal, or upward was significantly different (Fig. 5; GLM, df = 8, χ2 = 922.31, p < 0.0001). Moreover, the number of adults walking before taking off the leaf was counted, and the rates of walking individuals were between 10 and 33% at 1–1000 Hz.

Flight direction and percentage of B. tabaci adults that walked around the leaf before taking off
Suppression effect on B. tabaci fecundity by the ultrasonic focusing device
There were significant differences in the residual rates of adults on leaves (Table 1; GLM, df = 2, χ2 = 82.68, p < 0.0005), number of pairs in courtship behavior (Table 1; GLM, df = 2, χ2 = 19.68, p < 0.0001) and laid eggs (Table 1; GLM, df = 2, χ2 = 73.34, p < 0.0001) among the three treatments. The mean residual rate of adults on leaves just after 2 h/days or 4 h/days of noncontact vibration treatments was 49.4% or 41.4%, respectively, and significantly lower than that under no treatment (Table 1; Dunnett’s test, p < 0.01 and p < 0.001, respectively). The mean number of pairs in courtship behavior after just 2 h/days or 4 h/days of noncontact vibration was 0.5 or 0.4, respectively, and significantly less than that under no treatment (Table 1; Dunnett’s test, p < 0.001 and p < 0.0001, respectively). The mean number of laid eggs under the 4 h/days treatment was 14.5 and significantly less than that under no treatment (Table 1; Dunnett’s test, p < 0.05), but that under the 2 h/days treatment was not significantly different compared to that under no treatment (Table 1; Dunnett’s test, p = 0.24).
Discussion
These experiments showed that the noncontact vibration with 16 mN of acting force generated by an ultrasonic focusing device not only dispersed B. tabaci and A. gossypii but also suppressed B. tabaci fecundity after treatment for longer than 4 h/days. First, we examined the dispersion effect on the two insect pests, and the dispersion rate was significantly higher for B. tabaci than for A. gossypii. In the B. tabaci experiment, air blowing treatments (2.6 m/s, 5.0 m/s and 10.5 m/s) were also applied as a comparison group, and the number of dispersed B. tabaci significantly increased under 5.0 m/s and 10.5 m/s conditions compared to that under no treatment. The dispersion rate of B. tabaci at a wind velocity of 5.0 m/s for 60 s was 62.1% and was similar to that at a modulation frequency of 1–240 Hz (Fig. 2). The wind velocity of the noncontact vibration generated by an ultrasonic focusing device was also measured and lower than 0.2 m/s, the detection limit. It has been reported that take-off and flight of B. tabaci are influenced by wind velocity and increasing wind velocity from 0 to 0.3 m/s significantly reduced the number of B. tabaci flying away (Isaacs et al. 1999). Here, we tested several wind velocities from 2.6 to 10.5 m/s, which were more than 10 times as fast as those in the previous study; thus, it is suggested that adults on the leaves could not hold on and dispersed by such a strong wind. According to the Beaufort wind scale (Beaufort 1805), 5.0 m/s is classified as a “gentle breeze”, which makes leaves and small twigs move constantly, and 10.5 m/s is a “fresh breeze”, which makes small trees and leaves begin to sway. As these levels of wind power can break branches of crops in greenhouses, the dispersion of B. tabaci using air blowing is suggested to be inappropriate. The noncontact vibration generated by an ultrasonic focusing device is more appropriate for crop protection in greenhouses because it can make B. tabaci and A. gossypii adults disperse from leaves not by wind power but by noncontact vibration.
In contrast to B. tabaci, the dispersion effect of the modulation frequency on A. gossypii adults, regardless of whether they were winged or wingless, was at most less than half as strong as that on B. tabaci. Several species of aphids, such as the sweet pepper aphid Aphis nerii and large cat's ear aphid Uroleucon hypochoeridis, are known to have herbivore detection mechanisms that use vibration to defend against natural predators (Hartbauer 2010). It is suggested that A. gossypii individuals could also detect noncontact vibration stimuli, and some of them dropped from eggplant leaves.
Comparing these two insect pests, the ultrasonic focusing device was more effective on B. tabaci than on A. gossypii in terms of dispersion, and thus we investigated the flight behavior of B. tabaci under 30 s of the 30 Hz modulation frequency treatment. The rate of flying or remaining adults was not significantly different among modulation frequency conditions, and the latency time to take off the leaf fitted the negative binomial model on every modulation frequency condition (Figs. 4, S2). Generally, if every unit in the series was exposed equally to the chance of containing the organism, the distribution would follow the Poisson series, each unit having the population mean as its expected frequency, and the negative binomial distribution is a discrete probability distribution that models the number of successes in a sequence of independent and identically distributed Bernoulli trials before a specified nonrandom number of failures, which is denoted r, occurs (Bliss and Fisher 1953). For example, Martini et al. (2018) reported that the dispersion of duration time of the Asian citrus psyllid Diaphorina citri Kuwayama (Hemiptera: Liviidae) on an insect flight mill after taking off fitted to the negative binomial model. In this experiment, the latency time of B. tabaci adults to take off the leaf after noncontact vibration was comparable to the number of failures (i.e., remaining on the leaf) before r successes (i.e., taking off the leaf); thus, it is suggested that the histograms of B. tabaci adults flew 1–30 s were fitted to the negative binomial model regardless of modulation frequency. Approximately 60–90% of dispersed B. tabaci individuals flew within 5 s, 86% of them flew downward, and 10–33% of them walked around before flying away. Many species of insects show startle responses, such as fast jerky movements, freezing, and escaping under vibration stimuli (Bullock 1984; Friedel 1999), and such responses are considered to have evolved for evasion from predation (Takanashi et al. 2019). Yanagisawa et al. (2021) reported that B. tabaci can also show startle responses following substrate-borne vibrations. Therefore, it is suggested that B. tabaci adults showed startle responses such as freezing during vibration and escaping under noncontact vibration, i.e., some of them walked around the leaf and flew off of the leaf within a few seconds, but most of them took off immediately after the noncontact vibration stimulus working on their bodies. Ribak et al. (2016) investigated the stabilizing mechanism of whiteflies when they performed take-off jumps with closed wings and revealed that the take-off speed and angle from the horizontal ground were 0.61 ± 0.097 m/s and 64 ± 10.7°, respectively (mean ± SD). Here, we conducted this experiment by attaching leaves infested with B. tabaci adults to the vertical wall of a cage, and it was revealed that the individuals taking off could not engage in normal flight behavior due to continuous noncontact vibration.
Second, we examined the suppression effect on B. tabaci fecundity using an ultrasonic focusing device, and the number of eggs laid by female adults significantly decreased by 4 h/days treatment at a 30 Hz modulation frequency. Male and female whiteflies, including B. tabaci, communicate via substrate-borne vibrations (Kanmiya 2006). Yanagisawa et al. (2021) reported that substrate-borne vibrations artificially generated on tomato plants infested with B. tabaci adults by a vibration exciter decreased the population density of nymphs on leaves, which might be due to startle responses induced by the signals irrelevant to courtship sounds. While further confirmation is necessary, the noncontact vibration force generated by ultrasonic transducers on B. tabaci adults might have caused a disturbance in mating. In this study, however, we did not examine the effect of continuous vibration treatment on the hatching rate, growth of nymphs or emergence rate of B. tabaci or optimize the treatment conditions (e.g., treatment period, interval, modulation frequency) using this device. Further studies are required to apply this device for insect pest control in large-scale greenhouses, and the following prospective applications should be considered: (1) the single use of this device for suppression of fecundity of B. tabaci as well as artificial pollination of fruit vegetables such as tomatoes or strawberries (Kobayashi 2013; Shimizu and Sato 2018), and (2) the combination of this device and a vacuum with a light attractant (Stukenberg and Poehling 2019; Kim et al. 2012) or sticky traps (Sharaf 1982; Gerling and Horowitz 1984) to remove insect pests.
References
Beaufort F (1805) Beaufort wind scale. http://www.spc.noaa.gov/faq/tornado/beaufort.html. Accessed 29 Jul 2021
Ben-Ari M, Inbar M (2014) Aphids link different sensory modalities to accurately interpret ambiguous cues. Behav Ecol 25:627–632. https://doi.org/10.1093/beheco/aru033
Bliss CI, Fisher RA (1953) Fitting the negative binomial distribution to biological data. Biom 9:176–200
Bullock TH (1984) Comparative neuroethology of startle, rapid escape, and giant fiber-mediated responses. In: Eaton RC (ed) Neural mechanisms of startle behavior. Plenum Press, New York, pp 1–13
Edwards OR, Franzmann B, Thackray D, Micic S (2008) Insecticide resistance and implications for future aphid management in Australian grains and pastures: a review. Aust J Exp Agric 48:1523–1530. https://doi.org/10.1071/EA07426
Friedel T (1999) The vibrational startle response of the desert locust Schistocerca gregaria. J Exp Biol 202:2151–2159. https://doi.org/10.1242/jeb.202.16.2151
Gerling DAN, Horowitz AR (1984) Yellow traps for evaluating the population levels and dispersal patterns of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae). Ann Entomol Soc Am 77:753–759. https://doi.org/10.1093/aesa/77.6.753
Ghosh A, Jagdale SS, Dietzgen RG, Jain RK (2020) Genetics of Thrips palmi (Thysanoptera: Thripidae). J Pest Sci 93:27–39. https://doi.org/10.1007/s10340-019-01160-2
Hartbauer M (2010) Collective defense of Aphis nerii and Uroleucon hypochoeridis (Homoptera, Aphididae) against natural enemies. PLoS ONE 5:e10417
Horowitz R, Denholm I, Morin S (2007) Resistance to insecticides in the TYLCV vector, Bemisia tabaci. In: Czosnek H (ed) Tomato yellow leaf curl virus disease. Springer, Dordrecht, pp 305–325
Hoshi T, Ochiai Y, Rekimoto J (2014) Three-dimensional noncontact manipulation by opposite ultrasonic phased arrays. Jpn J Appl Phys 53:KE07. https://doi.org/10.7567/JJAP.53.07KE07
Isaacs R, Willis MA, Byrne DN (1999) Modulation of whitefly take-off and flight orientation by wind velocity and visual cues. Physiol Entomol 24:311–318. https://doi.org/10.1046/j.1365-3032.1999.00144.x
Kanmiya K (1996) Discovery of male acoustic signals in the greenhouse whitefly, Trialeurodes vaporariorum (WESTWOOD) (Homoptera: Aleyrodidae). Appl Entomol Zool 31:255–262. https://doi.org/10.1303/aez.31.255
Kanmiya K (2006) Mating behaviour and vibratory signals in whiteflies (Hemiptera: Aleyrodidae). In: Drosopoulos S, Claridge MF (eds) Insect sounds and communication: physiology, behaviour, ecology and evolution. Taylor and Francis, London, pp 365–379
Kanmiya K (2011) Mating behaviour and vibratory signals in Aleyrodidae (Hemiptera). In: Miyatake Y (ed) Insect communication by sounds and vibrations. Hokuryukan, Tokyo, pp 217–240 (in Japanese)
Kim MG, Yang JY, Chung NH, Lee HS (2012) Photo-response of tobacco whitefly, Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae), to light-emitting diodes. J Korean Soc Appl Biol Chem 55:567–569. https://doi.org/10.1007/s13765-012-2115-4
Klatt B, Holzschuh A, Westphal C, Clough Y, Smit I, Pawelzik E, Tscharntke T (2014) Bee pollination improves crop quality, shelf life and commercial value. Proc Biol Sci. https://doi.org/10.1098/rspb.2013.2440
Kobayashi N (2013) Development of pollinating device -specifying character frequency of flowers of fruit vegetable and measuring success or failure of pollination-. Rep Kansai Soc Agric Mach Jpn 113:28–29 (in Japanese)
Martini X, Rivera M, Hoyte A, Sétamou M, Stelinski L (2018) Effects of wind, temperature, and barometric pressure on Asian citrus psyllid (Hemiptera: Liviidae) flight behavior. J Econ Entomol 111:2570–2577
Meng Z, Duan A, Chen D, Dassanayake KB, Wang X, Liu Z, Liu H, Gao S (2017) Suitable indicators using stem diameter variation-derived indices to monitor the water status of greenhouse tomato plants. PLoS ONE 12:e0171423. https://doi.org/10.1371/journal.pone.0171423
Morimyo N (2017) A background and a summary of “the manual on utilization of native bumblebees, as an alternative species of Bombus terrestris”. Plant Prot 71:697–699 (in Japanese)
Naveen NC, Chaubey R, Kumar D, Rebijith KB, Rajagopal R, Subrahmanyam B, Subramanian S (2017) Insecticide resistance status in the whitefly, Bemisia tabaci genetic groups Asia-I, Asia-II-1 and Asia-II-7 on the Indian subcontinent. Sci Rep 7:1–15. https://doi.org/10.1038/srep40634
Orfanidou CG, Pappi PG, Efthimiou KE, Katis NI, Maliogka VI (2016) Transmission of Tomato chlorosis virus (ToCV) by Bemisia tabaci biotype Q and evaluation of four weed species as viral sources. Plant Dis 100:2043–2049. https://doi.org/10.1094/PDIS-01-16-0054-RE
Osakabe M, Uesugi R, Goka K (2009) Evolutionary aspects of acaricide-resistance development in spider mites. Psyche 2009:1–11. https://doi.org/10.1155/2009/947439
Ribak G, Dafni E, Gerling D (2016) Whiteflies stabilize their take-off with closed wings. J Exp Biol 219:1639–1648. https://doi.org/10.1242/jeb.127886
Rosner H (2013) Return of the natives: how wild bees will save our agricultural system. Sci Am 309:70–75
Sharaf NS (1982) Determination of the proper height, direction, position and distance of a yellow sticky trap for monitoring adult sweet potato whitefly populations (Bemisia tabaci Genn., Homoptera: Aleyrodidae). Dirasat 9:169–182
Shimizu H, Sato T (2018) Development of strawberry pollination system using ultrasonic radiation pressure. IFAC-PapersOnLine 51:57–60. https://doi.org/10.1016/j.ifacol.2018.08.060
Shimizu H, Hoshi T, Nakamura K, Park JE (2015) Development of a noncontact ultrasonic pollination device. Environ Control Biol 53:85–88. https://doi.org/10.2525/ecb.53.85
Stukenberg N, Poehling HM (2019) Blue–green opponency and trichromatic vision in the greenhouse whitefly (Trialeurodes vaporariorum) explored using light emitting diodes. Ann Appl Biol 175:146–163. https://doi.org/10.1111/aab.12524
Takanashi T, Uechi N, Tatsuta H (2019) Vibrations in hemipteran and coleopteran insects: behaviors and application in pest management. Appl Entomol Zool 54:21–29. https://doi.org/10.1007/s13355-018-00603-z
Yamada M (2008) Emergence of large-scale greenhouse farms as a main figure of protected cropping in Japan. Res Bull Aichi Agric Res Ctr 40:1–7 (In Japanese with English abstract)
Yanagisawa R, Suwa R, Takanashi T, Tatsuta H (2021) Substrate-borne vibrations reduced the density of tobacco whitefly Bemisia tabaci (Hemiptera: Aleyrodidae) infestations on tomato, Solanum lycopersicum: an experimental assessment. Appl Entomol Zool 56:157–163. https://doi.org/10.1007/s13355-020-00711-9
Zuur AF, Ieno EN, Walker NJ, Saveleiev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
We would like to thank Mr. T. Sato for technical support in recording high-speed movies and measuring the wind velocity of the air pump.
Funding
This research was supported by the research program on development of innovative technology grants from the Project of the Bio-oriented Technology Research Advancement Institution (BRAIN).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Urairi, C., Hoshi, T. & Ohta, I. Development of a new insect pest control device using noncontact force generated by ultrasonic transducers. Appl Entomol Zool 57, 183–192 (2022). https://doi.org/10.1007/s13355-022-00774-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13355-022-00774-w