
Abstract
The increasing demand for sustainable agriculture has propelled researchers to envision novel technological innovations. In this situation, the implementation of nanoparticles eventually gained huge importance and proved to be beneficial in the field of sustainable agriculture. To date, there have been numerous reports that have focused on utilizing the nanotechnological approach for positively influencing plant growth, development, and yield-associated parameters. Over the years there have been several successful applications of nanoparticles to mitigate the toxic effects imposed by abiotic and biotic stress factors on plants. Among the three major approaches of nanoparticle application to plants, seed priming has played an important role in plant growth promotion owing to its ability to activate signaling pathways concerned with phytohormone and ROS synthesis. So far, the reports available on the role of nano-priming in boosting plant growth and tolerance against environmental stress have been mostly associated with Zn, Fe, Cu, and other essential micronutrients. But in recent years application of metallic and non-metallic nanoparticles like silver, gold, platinum, titanium, and carbon nanotubes is also worth mentioning. The outcome of nano-priming is also subject to the appropriate concentration, charge, and size of NPs used. Besides, nano-priming can elicit a plethora of physiological, biochemical, and molecular responses in the germinating seeds that require proper understanding. Hence this review aims to document the progress made in the nano-priming research and pave the way for other advancements that can be made in the future.
Graphical abstract

Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Agriculture is one of the indispensable domains of life. According to FAO (https://www.fao.org/fileadmin/templates/wsfs/docs/Issues_papers/HLEF2050_Global_Agriculture.pdf), about 60% of the world’s population depends on agriculture for survival. The World Bank has identified agricultural development as a compelling tool to purge poverty & hunger, boost shared prosperity, and ensure global food security. With the existing population growth rate, the food supply should be increased by 50–70% to ensure food for all [58]. Agriculture is challenged by unfavorable climate, anthropogenic activities, shrinking arable lands, soaring temperatures, and frequent spells of drought and floods. Flanked by this situation, the obsession to create and adopt new technologies for agricultural intensification and extension should be intended. However, sustainability should be of prime consideration to protect against environmental fragmentation. Thus, sustainable agriculture holds the key to global food security [54].
Since its advent, nanotechnology has been an innovative and groundbreaking tool to tackle transpiring global issues and agriculture is no exception. With a characteristic dimension of 100 nm or less, nanomaterials serve as an excellent toolset to render efficient and targeted delivery of nutrients and ions to living cells, which makes them a propitious candidate for implementation in agricultural sector. Nanomaterials have been widely used in agriculture and allied fields for precision farming, as nano-sensors, nano-pesticides, nano-fertilizers, nano-herbicides, and agents for seed encapsulation and priming. Slow and controlled release, targeted delivery, and micro resources have bestowed nanomaterials as potential alternatives for chemical fertilizers which usually lead to leaching, bioaccumulation, residual effects, and long-term detrimental effects on soil health and the microenvironment [73].
Seed invigoration has been of profound importance in the past decade and seed priming has emerged as a salient and widely used technique for seed invigoration. Seed germination can be divided into 3 distinct phases: imbibition phase (rapid water uptake phase), activation or preparatory phase (low rate of water uptake, synthesis of new proteins and other essential organelles & molecules) & germination phase (protrusion of radical and plumule). Seed priming is a controlled hydration of seeds that allows germination to extend till the activation phase but restricts the initiation of the third phase [57]. Thus priming ‘prepares’ the seed for future sowing and stress atrocities. Hydropriming (with water), Osmo-priming (with osmolytes like PEG, sugar alcohols), Hormo-priming (with gibberellic acid, polyamines, salicylic acid), Chemical priming (with salts like sodium selenite, silicon), Vitamin priming (with thiamine, ascorbate), Nutri-priming (with various micro and macronutrients), Redox priming (with hydrogen peroxide), Bio priming are some of the well-documented priming techniques till date [48, 72].
With the advent of nanotechnology, nanomaterials have been extensively used as seed priming agents, and the technique, termed ‘nano-priming’ has been serving as a useful and innovative tool for sustainable agriculture by enhancing seedling establishment, seed quality, and crop yields as well as increasing tolerance to environmental stresses. Nano-priming has been implemented in a wide range of plants and various metallic and non-metallic elements have been used as priming agents to improve the growth of the plants or mitigate abiotic and biotic stresses since the past decade. Numerous research and review articles have been published in last 5 years on nano-priming that has improved our current understanding of this technique and also highlighted several unresolved research gaps (Fig. 1). The molecular basis of nanopriming involves inducing a cascade of mechanisms within seeds that lead to enhanced germination and growth. These mechanisms include forming nanopores for improved water uptake, modulating reactive oxygen species (ROS) and antioxidant systems, generating hydroxyl radicals for cell wall loosening, and accelerating starch metabolism by enhancing starch catalysing enzymes [4]. Additionally, nano-priming has been shown to stimulate the production of growth-promoting metabolites, trigger hormone secretion, and reduce ROS levels, ultimately enhancing disease resistance in plants [72]. Seed priming with AgNP enhances aquaporin genes (PIP) and cotton seeds primed with (PNCs) exposed to salt stress showed induction of genes involved in ROS signaling and ion homeostasis [72] Despite the fact, that extensive research needs to be performed to accentuate the positive and negative impacts of nano-priming on sustainable agriculture, few countries have commercialized this approach by developing nanomaterials to amplify crop productivity. For instance, Nanovec TS80 curated by Laboratórios Bio-Médicin, Brazil, is a nanocapsule composed of Mo, Mn, and Zn which has been considered to be efficient in promoting seed germination, root development, nutrient uptake, and biological nitrogen fixation in leguminous plants [71]. Therefore, this comprehensive review provides a holistic view of the role of commonly used nanoparticles in growth enhancement and stress amelioration. It also highlights the mode of action of the nano-priming mediated responses. Although there are previous reviews on nano-priming [20, 38, 55], this review stands out as it discusses the most recent advances and summarizes the findings and research gaps to serve as a cue for future research direction.

Bar graph representing the number of articles published related to nano-priming in the last 5 years (Data source: PubMed)
Growth enhancement through nano-priming
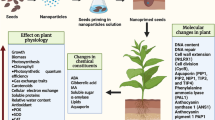
Nanoparticles (NPs) have indeed become an integral part of sustainable agriculture owing to their superior physical and chemical properties compared to its bulk counterparts. Over the years a wide range of nano-formulations have been curated where different carriers have been efficiently employed for gradual and specific release of macro as well as micronutrients. Depending upon their size, shape, surface charge, and degree of hydrophobicity nanoparticles can be taken up either by passive diffusion, active transport via transmembrane carrier proteins like aquaporins, or by endocytosis into the cell membrane. The entry of NPs can even be facilitated by creating small pores on the cell membrane or by forming a complex with organic compounds of root exudates [20, 55]. Post entry through the root cells, NPs are translocated to various plant tissues either via apoplastic (from root via xylem to shoot) or symplastic pathways (transport of water and substances between two cells via plasmodesmata). NPs can positively influence crucial physiological and metabolic activities in plants that make nano-priming a significant tool to facilitate breaking seed dormancy, enhancing seed germination, and seedling vigor (Table 1). NPs possibly function by creating nanopores that assist the uptake of water and nutrients into the seeds. Besides it triggers the formation of ROS (OH−, O2− radicals) that makes the seed coat permeable and facilitates germination (Fig. 2) [55]. They even initiate a downstream signaling cascade which includes upregulation of α- amylase or other hydrolytic enzyme activity to break down the reserve food present in the endosperm essential for optimum germination. Besides, it might also help in the production of secondary metabolites like phenols to eventually promote seedling growth and development. Nano-priming upregulates the expression of aquaporin genes; establishes cell-to-cell communication and allows the diffusion of H2O2 through the cell membrane which contributes to better and synchronized seedling emergence [36, 48]. The diffusion of ROS across the seeds can be interlinked with the phytohormonal GA/ABA balance (enhanced GA accumulation and lower ABA formation) which is considered to be indispensable for degradation of starch and initiate germination of the embryo. Though phytohormones including ABA and auxin have no positive impact on the germination of nano-primed seeds but they have been found to aid the attachment of nanoparticles to the seed coat [46]. Nano-priming has been recognized for its ability to enhance root development which eventually facilitates efficient nutrient uptake and mobilization within the plants. Moreover, they assist the activity of enzymes associated with increased nutrient accessibility and boost crop productivity [8].

Mode of action of nano-priming induced germination, growth, and stress resistance in plants. Nano-priming provides a ‘headstart’ to the seeds that promote germination and plant growth. It also ‘prepares’ the plant for future incidences of abiotic and biotic stress. NPs usually diffuse across the seed coat or create nanopores which facilitate their uptake into the seed. As soon as nanoparticles enter the seed, they can enhance the aquaporin gene expression which positively influences the uptake of water. NPs also initiate ROS formation at an optimum level which is regulated by elevated enzymatic antioxidative activities and they tend to act as a signaling molecule to activate GA and downregulate ABA synthesis. GA directly upregulates α-amylase activity and allows starch hydrolysis into soluble sugar which is indispensable for seed germination. This eventually leads to improvement in physiological parameters associated with growth. NP priming also activates the defense system against abiotic and biotic stress in plants by accumulation of secondary metabolites and strengthening the antioxidant system
The merits of nano-priming have further intrigued researchers to investigate targeted and sustained delivery of essential nutrients through nano-priming for growth promotion in plants. These nano-micronutrients have been found to improve the germination percentage, seedling emergence rates, seedling vigor, photosynthetic activity, carbon and nitrogen assimilation, antioxidative machinery, and even support reproductive growth. Among the several nano-micronutrients, Zn and Fe have been used extensively in various forms (Zinc oxide, Zerovalent Iron, Fe3O4, FeS2, etc.), concentrations, in different plant species (like rice, wheat, capsicum, maize, etc.).. Zn and Fe, both are responsible for maintenance of several important physiological functions in plants like photosynthesis, nitrogen assimilation, respiration and DNA metabolism [14], Priming of seeds by these nutrients impart a plethora of advantages to the plants. Nano-Nutri-priming is therefore a beneficial and sustainable approach to boost plant growth compared to their bulk counterparts [31, 37, 40, 60, 62, 64]. Often the beneficial effects of nano-nutrients depend on the rate of biotransformation of NPs into organically bound nutrients within plant cells and can be governed by the shapes, dimensions, and surface charge of NPs [64]. In addition, nano-macronutrients have also been utilized as efficient seed-priming agents to enhance biochemical parameters for overall growth and productivity [70]. However, it is imperative to assess the optimum concentration of NP to be used as a priming agent because NPs in large concentrations can result in toxicity [31]. Besides, an excess of one nutrient is often known to interact and inhibit the uptake of other essential nutrients. These should be taken into consideration before implementing nano-priming to ensure optimum results.
NPs synthesis also plays a pivotal role in ensuring proper nutrient delivery with minimal environmental residue. The coating material, thus, plays a crucial role in determining the efficacy of NPs. PEG-coated NPs are superior to other NPs in promoting germination and supplementing nutrients due to the hydrophilic nature of PEG, which makes them incapable of interacting with cell membranes and plant proteins, thus eliminating the chances of toxicity. PEG coating also helps water uptake by seeds and prevents the amalgamation of NPs that can otherwise restrict their usual functions [64]. Nano-priming has also provided exemplary results in field experiments. Wheat seeds treated with Fe2O3 NPs not only promoted seedling growth, vigor, and photosynthetic efficiency but also accelerated nutrient absorption by plants which eventually led to the biofortification of grains with Fe [77]. Similar results have been documented with rice [30]. But still, most of the reports on Nano-Nutri-priming are limited to the laboratories and lack proper field trials to validate the findings. While seedling performance is improved with nano-priming, it is also pertinent to assess whether the effect is persistent till maturity.
Besides the nutrients known to have significant roles in plants, some elements that are not utilized by plants directly, have also been tested as priming agents viz. Ag, Au, Ti, and Pt. These metals are biologically nonessential for plants as they are not required for their growth or maintenance but owing to their antioxidative potential and free radical scavenging activity they have been implemented to combat several abiotic and biotic stress incidences. For example, AgNPs reportedly augment photosynthetic activities by altering chloroplast ultra-structures [1], AuNPs increase the germination rate, overall growth of plant morphologically, chlorophyll content and yield by 23.9% [3], and TiO2 NPs increased seedling vigor, water content of leaves and antioxidant enzyme functioning in maize [39].
In recent decades carbon nanomaterials (CNMs), including single-walled carbon nanotubes (CNTs) (SWCNTs), multi-walled carbon nanotubes (MWCNTs), graphene (GR), and fullerenes have been reported to aid plant growth and can generate multiple responses which can also lead to increased stress tolerance [46]. MWCNTs alone or synergistically with cold plasma exposure can induce the xanthophyll cycle (β-carotene, α carotene, zeaxanthin, and lutein) to protect the photosynthetic machinery and improve the photosynthetic activity; enhance carotenoid content and upregulate the genes associated with terpenoid metabolism [74].
Chitosan nanoparticles have also become quite relevant in the agricultural sector due to their non-toxic, biocompatible properties, prolonged stability, and better delivery of micronutrients in plant tissues thus enhancing their efficacy in metabolic activities. Chitosan nanoparticles have shown promising effects on seed germination and at the same time enhanced plant photosynthetic efficiency and chlorophyll contents. NP priming with chitosan also increased auxin levels by upregulation of auxin synthetic genes [44]. Copper-loaded chitosan nanoparticles also found to have a profound role in promoting germination, seedling length, photosynthesis, and enhanced activity of enzymatic antioxidants in comparison to hydro-primed seeds [25].
Thus, the ability of nano-priming to serve as a propitious growth enhancer is quite evident from the previous discussions. However, most of such advancements are limited to preliminary laboratory findings and require proper validations through field trials for commercialization or agronomic use. Besides, another key determinant is the nutritional profiling of the product obtained through seed priming as well as analyzing its safety levels for consumption.
Abiotic stress management
Abiotic factors such as climate fluctuations, soil conditions, nutrient availability, and heavy metal contamination have inflicted dreadful impacts on the growth, development, and yield of plants. Hence to bridge the gap between population growth and food production, stress tolerance of crop plants must be improved. In this respect, nanotechnology can play a huge role in crop improvement through the alleviation of stresses. Seed priming can ameliorate adverse effects of abiotic (drought, salinity, flooding, heat, cold, heavy metal) stress in plants by triggering antioxidant metabolism, accumulation of non-enzymatic antioxidants, imparting stress memory and synthesis of stress proteins like late embryogenesis abundant (LEA), dehydrin and aquaporin (AQP) [36]. Among several abiotic stresses that can be mitigated through nano-priming, salinity, and heavy metal stress have been highlighted in detail.
Salinity stress
Globally around 20% of cultivated land and 33% of irrigated land have been ruthlessly affected by salinity stress which has raised a lot of concerns regarding global food security [12]. Salinity stress has been characterized by an exorbitant rise in Na+, Cl− ions in the soil which eventually alters soil porosity, and hydraulic conductivity and lowers the osmotic potential of soil. The toxic concentration of ions significantly alters the ionic balance, membrane stability and leads to rapid loss of water from cells [34] This leads to physiological conditions analogous to drought which is also delineated by restricted uptake of essential ions and minerals crucial for growth and development. In addition, salinity stress instigates ROS accumulation which triggers the destruction of proteins, nucleic acids, and lipids. The water deficit intervenes stomatal conductance thus hampering the photosynthetic activities and other associated parameters. Plants have an intrinsic defense mechanism that allows them to amplify the synthesis of osmotic solutes, endogenous hormones (ABA, BR, SA, GA), antioxidative capacity, compartmentalize ions, release secondary metabolites to lessen toxicity, and even interact with plant growth promoting rhizobacteria (PGPR) [22, 59, 82]. Depending upon the concentration of solute and the genetic potential of plants, the expression of stress resistance genes is upregulated to acclimatize themselves to the severity of stress [32].
Nanoparticles have been accredited for their profound role in salinity stress amelioration in plants. Metal oxide NPs, metal NPs, carbon nanomaterials, and nanocomposites have been utilized for the same owing to their ability to maintain ion homeostasis, induce an influx of K+, regulate water absorption as well as upregulate the expression of genes like NHX1 (Na+/H+ antiporter) for sequestration of excess Na+ in the vacuoles [21, 82]. Besides, they increase stomatal conductance, chlorophyll content, activity of enzymes (RuBisCo, carbonic anhydrase) and maintain the ultrastructure of chloroplasts, thus enhancing the photosynthetic efficiency. This subsequently prompted nano-priming with essential micronutrients to boost germination, enhance radical and plumule growth, lateral root formation, and enzymatic antioxidant activities (SOD, POD, CAT) to minimize the detrimental effects of ionic imbalance (Fig. 2) [36]. The mechanism of action of nanopriming in salinity stress extends to balancing reactive oxygen species production and enzymatic antioxidant activity, thereby enhancing the ability of plants to withstand water deficit conditions [60]. Nano-priming promotes α-amylase activity, GA synthesis, aquaporin gene expression, free amino acid, and plasma membrane lipid content as well as restricts the ABA levels to elevate the imbibition of water which eventually supports germination [18]. Therefore, Zn [17], Fe, Se [16], Si [56], and Mn [86] NPs have been utilized successfully to alleviate salt stress. Nanoparticles are often coated with plant extracts, phytohormones, and amino acids which impart stability and biocompatibility to NPs. The capping agents tend to alter the surface properties and pore space which makes them efficient for environmental stress remediation. For example, Vigna radiata L. seeds primed with glutamic acid-coated FeNPs regulated growth under salinity stress by facilitating antioxidant enzyme activities and osmolyte content. Glutamic acid has been speculated to have a crucial role in photosynthesis and nitrogen metabolism which was substantiated by improved chlorophyll content, morphological attributes, and yield [79].
Besides being potential growth regulators, Ag and Ti have been perceived for their response to osmotic stress. Nanopriming wheat and pearl millet with an optimum concentration of AgNPs increases tolerance to stress by increasing photosynthetic efficiency, reprogramming phytohormonal balance, decreasing Na+/K+ ratio, and enhancing secondary metabolite production [2, 41]. Ti2O NPs have also been recognized for their enzymatic antioxidant activity to scavenge ROS, increase K+ and phenolics content as well as reduce MDA and Na+ levels thus maintaining membrane stability [68].
To date, chitosan NPs have been well acclaimed as a bio-stimulant and elicitor as they assist the defense mechanism as well as regulate associated signal transduction pathways under salinity stress. They modulate enzymatic antioxidants, secondary metabolite production, and enzymes corresponding to their synthesis. Nanopriming Cu-chitosan NPs has been effective in wheat seedlings by boosting chlorophyll content, sugar levels, α-amylase, and SOD activity [23]. Chitosan nanoparticles loaded with curcumin too have been ascertained to counteract the toxic effects of salinity stress in wheat owing to the growth-enhancing properties of chitosan and the antioxidative activity of curcumin [33].
The constructive purpose of carbon nanomaterials to combat salinity-induced water deficits has also been verified by their antioxidative potential and enhanced osmolyte levels. This eventually aided nutrient uptake, maintenance of osmotic potential, and membrane integrity. For instance, polyhydroxy fullerene nanoparticles (PHF) and carbon nanotubes, graphene can easily mitigate oxidative stress generated by toxic Na+ concentrations in wheat and tomato respectively [26, 67].
Thus, nano-priming has emerged as a successful and sustainable alternative to bulk chemicals for imparting salinity stress tolerance to plants for efficient agriculture (Table 2). To date, micronutrients and metal NPs have been exploited for the same but the application of macronutrients still needs to be investigated. However, it is obligatory to assess the benefits of nano-priming in salinity stress alleviation through field experiments for implementation on a larger scale.
Heavy Metal Stress
Heavy metal contamination in soil has raised a lot of concerns in the agricultural sector due to its deep-rooted negative impact on the overall growth and development of plants. Non-essential heavy metals like Pb, Cr, Cd, and As are accumulated in the soil owing to several anthropogenic activities and natural disasters which inevitably hamper the productivity and quality of yield of commercially important crops. These metals get easily transported into the plant utilizing the transporters, channel proteins, and pathways responsible for the uptake of vital micronutrients like Fe, Zn, etc. owing to their analogous chemical structure. Their aggravated content not only affects the physiological and metabolic activities of a plant but can also cause potential harm to human beings. Therefore, amelioration of heavy metals in plants and reducing the health risk factors associated with the consumption of heavy metal-contaminated crops becomes the need of the hour. NPs are widely known to reduce heavy metal toxicity by reducing the bioavailable metals in soil, regulating the expression of genes associated with transport, and enhancing the antioxidative machinery as well as secondary metabolic pathways. Foliar spraying and root application of various metal oxide NPs were found to be effective in minimizing the adverse effects of heavy metals by upregulating the transport of essential nutrients and decreasing the translocation of heavy metals by adsorption. Apart from two common approaches, nano-priming has also grabbed a lot of attention for not only reducing heavy metal content in plants but also rescuing the yield-related parameters (represented in Table 3) [8, 23]. It has been established that seed priming with metal oxide NPs can promote the formation of a complex with the metal and the synthesis of metallothionine which eventually forms a complex with the heavy metals to restrict their uptake and translocation (Fig. 2) [45]. They even promote the antioxidative machinery to detoxify ROS produced excessively, maintain the ionic homeostasis of the cells as well as promote photosynthetic activity to induce tolerance against metal toxicity [36]. The reduced rate of germination in toxic metal-contaminated soil can even be retrieved by these NPS as they accelerate α-amylase activity to break down starch, promote germination, and increase seedling vigor.
The antioxidative potential of Zn and Fe (as they serve as co-factors of several antioxidant enzymes) reportedly decreased the oxidative damage induced by Cd, improved the agronomic parameters, enhanced Zn, Fe content, and reduced Cd in vegetative parts as well as grains [30, 62]. It was also found to assist growth and maintain the metabolomic profile of fragrant rice varieties when subjected to Cd stress [45]. Besides, nano-priming reduces Cd accumulation and upregulates the pathways associated with purine, pyrimidine, amino acids, flavonoids, polyunsaturated fatty acids, and other metabolite biosynthesis. Atrocities involved with As stress can also be mitigated by nano-priming with Zn. Nano-priming increases germination percentage and seedling growth, reduces cell damage, and boosts antioxidant levels [6].
Silicon (Si), an essential component of plant cell walls often serves as the first line of defense against several abiotic and biotic stress factors. The role of Si in heavy metal toxicity amelioration has been confirmed by its ability to reduce the soil pH and reduce bioavailable heavy metal uptake into plants. Either independently or in congruence with other essential nutrients, metabolites, and hormones SiNPs can downregulate transporters responsible for their translocation from root to shoot and express genes responsible for heavy metal sequestration in cell vacuoles. These features led to further application of SiNPs in seed priming resulting reduction of Cd content in wheat grains and eventually improved morpho-physiological, photosynthetic, and yield-related parameters [35]. Application of nano-priming with MWCNT has also been established for the amelioration of Cd toxicity in plants [7]. Other NPs that have been used as priming agents against heavy metal stress include TiO2NPs [47], polysuccinimide NPs [83], and aspartic acid-derived CuNPs [80].
Nano-priming in biotic stress amelioration
Biotic stress is an umbrella term that encompasses damage caused to crops by biotic agents such as fungi, viruses, bacteria, parasitic nematodes, insects, and weeds on other plants. Plants have developed sophisticated immune systems to fight against such biotic agents. Physical obstacles like waxes, dense cuticles, hairs, and special trichomes serve as the first line of defense to restrict pathogen entry [85]. However, pathogen invasion triggers varied immune responses in plants such as Hypersensitive Response (HR), Systemic Acquired Resistance (SAR), Induced Systemic Resistance (ISR) via activation of several mediators like Jasmonic Acid (JA), Salicylic Acid (SA), Ethylene (ET), Pathogenesis Related proteins (PR proteins) and secondary metabolites (tannins, alkaloids, phenols). Plants also restrict herbivory or growth of weeds in proximity by either secreting chemicals (allele-chemicals) or through specialized structures (thorns, raphides). Biotic stress leads to massive economic losses every year (20–40% yield loss worldwide). Among several approaches to control biotic agents, the most propitious ones to date have been chemical, biological, genetic, agricultural control, and integrated pest management. Chemical control (pesticide, fungicide, herbicide) is however the most preferred and convenient mode of control but is challenged by the increasing resistance of pathogens and residual effects on animals, humans, and the environment (soil and water contamination). Besides, climate change has confronted locally adapted crop genotypes with migrant pathogens, thereby introducing new biotic stress factors [53].
NPs can either function as nanocide, by killing or restricting the growth of phytopathogens or they might elicit a defense response in treated plants by upregulating responsive genes or triggering anti-oxidant enzyme activities. In-vitro assays demonstrate the efficacy of NPs in killing or inhibiting microbial growth whereas, in vivo, studies highlight the potential of NP-mediated seed priming in imparting enhanced disease resistance to the treated plants. Not necessarily, the NPs follow a particular mode of action, but rather different NPs, with characteristic functions might follow one or more of the above-mentioned routes alone or in combination. The mode of action of NPs as nanocides against seed-borne pathogens might be as follows:
-
The NPs are known to interact directly with the walls of the microbes, resulting in ion uptake and cellular damage. Reports claim that most of the microbes have negative zeta potential compared to the employed NPs that possess a positive potential. These two opposing forces result in strong electrostatic attraction and powerful binding between the NP and the microbial cell. It has also been claimed that Gram-negative bacteria are more susceptible to NPs due to the presence of the external lipopolysaccharide layer imparting a negative charge to the cell [76]. However, other interactions such as receptor ligand, and hydrophobic interactions may serve as a potential cause.
-
The second major cause of NP-mediated stress control could be due to ROS-induced cellular destruction. Uncontrolled ROS leads to cytotoxicity by inducing oxidative damage to the cellular proteins, membranes, and nucleic acid, thereby causing cell death [76].
-
Inactivation of major metabolic enzymes and transporters may be another way of combating biotic agents. Gold NP has been found to disrupt the proton pump in fungi [81].
-
Another plausible mode of action can be NP-induced changes in the cell wall of the microbes. Depolarization of the cell increases cellular permeability leading to the unregulated flow of ions, culminating in cell disintegration and death [76].
Nano-priming has also been effective in eliciting enhanced defense response in planta. Reports reveal that priming with nanomaterials is of greater importance than soil or foliar application as NPs show greater penetration via seed coat leading to enhanced water and nutrient uptake, enzyme activation, and cellular damage repair [75]. Blending bioactive compounds into nanostructures to ensure slow release to target cells is one of the most promising nano-technological tools. Commonly employed NPs in this regard are Chitosan NPs, CuO NPs, SiO2 NPs, ZnO NPs, MgO NPs and TiO2NPs. However, there are major crevices in understanding the mechanism of action of NPs in imparting resistance to several pathogens and thus disease tolerance [61].
Chitosan is known to impart tolerance against several pre- and post-harvest diseases to different crops by inducing phytoalexin synthesis, callose deposition, activities of reactive oxygen species, lignification, enhanced defense enzymes, and PR proteins. The conversion of chitosan to nano-chitosan alters certain characteristics of the molecule. Features like biocompatibility, reduced toxicity, biodegradability, and physiochemical properties like size, cationic nature, and surface area make it a potent elicitor of systemic resistance in plant responses by altering its biological activity. Cu-Chitosan nanoparticle primed seeds were effective in boosting plant growth along with building up a defense system against Curvularia leaf spot (CLS) disease of maize [9] by upregulating antioxidant levels including SOD, POD, PAL, and PPO. Priming of pearl millet seeds with chitosan nanoparticles has also been found beneficial in conferring resistance against downy mildew disease caused by the biotrophic oomycete Sclerospora graminicola. In comparison to bulk chitosan, trace amounts (250 mg/kg) of chitosan nanoparticles (CN) were able to generate a similar response in the plants against pathogens. Besides, CNs being tiny in size could easily enter the tissues, thus reducing the time of resistance induction in the treated plants [74]. Nitric oxide (NO) is responsible for generalized stress response in plants. NO, either by post-translational modification (S-nitrosylation, carbonylation, tyrosine nitration) of target proteins or together with secondary messengers in a signaling cascade (c-GMP) can alter specific defense gene expression [63]. It is also known to induce the synthesis of phytoalexins (an endogenous antimicrobial substance of plants), which lead to the accumulation of secondary metabolites and trigger the activities of antioxidant enzymes [19]. NO molecules are the key mediator of CNs-induced resistance response as evident from the increase in NO content in primed seeds and suppression of defense response on treatment with NO scavenger. CN-mediated defense response had been correlated with the enhanced activity of the defense genes and antioxidant enzymes such as Phenyl Ammonia Lyase (PAL), Polyphenol oxidase (POX), peroxidase, SOD, and catalase. Increased expression of PR proteins (PR-1 & PR-5) also provided an important cue towards augmented defense machinery post-treatment.
Zinc is associated with a plethora of biological processes in plants such as the maintenance of structural and functional integrity of biological membranes, protection of cellular proteins and lipids from oxidative damage, regulates expression of antioxidant enzymes like SOD, POD, GR and also known to induce lignification on pathogen invasion. CLS disease inflicts severe leaf necrosis and ROS accumulation in maize, thus hampering the photosynthetic efficiency, which leads to massive yield loss (60% yield loss yearly). The symptoms and outbreak of the disease are intensified in alkaline soil conditions where the bioavailability of Zn is limited. Zn-chitosan NPs had been found to inhibit spore germination and mycelial growth of C. lunata in-vitro. Pot experiments and field studies with maize seeds primed with Zn-chitosan NPs inoculated with CLS-causing pathogen, delivered improved resistance against CLS. Upregulation of SOD, POD, PAL & PPO expression with enhanced lignification might be the plausible mechanism that abated the microbial infection and its associated atrocities [10].
Silicon is a quasi-essential mineral nutrient for plant growth and development that helps in a better adaptation of plants under environmental stress conditions. Si-NPs besides enhancing the growth & development of plants, also serve as nanocides, herbicides, fertilizers & delivery agents for amino acids, and proteins [27]. Si NPs act as a physical barrier to prevent pathogen entry as they get deposited in a thick layer beneath the cuticle, subcuticular layer, cell wall, and intercellular spaces or develop a biofilm at the epidermal cell wall. This mechanism strengthens the cell wall and restricts pathogen invasion efficiently. Besides, a higher surface-to-volume ratio of Si-NPs increases the reactivity and bioavailability of Si and also spikes up its biological and molecular functions in the cell. Seed priming with SiO2 NPs is more beneficial than foliar application in reducing nematode multiplication, galling, and disease indices in beetroot [42]. Besides, the superior functioning of the antioxidant and defense enzymes contributes to the management of three harmful pathogens causing a disease complex in beetroot viz. Pectobacterium betavasculorum (a pathogenic bacterium causing vascular necrosis, wilting and black streaks on the leaves and petioles, soft rots due to secretion of various extracellular digestive enzymes), Meloidogyne incognita (a root-knot nematode causing severe damage to epidermis, cortex, and stele in root cells, thereby hindering water and nutrient absorption) and Rhizoctonia solani (a pathogenic fungus responsible for crown and root rot diseases). SiO2 NPs priming is also found effective against pathogens (Alternaria dauci and Pectobacterium carotovorum) causing diseases of carrots that cause massive yield losses [75]. However, how the resistance is imparted is yet to be revealed.
FeS-NPs serve as an antifungal agent against Fusarium verticillioides, causing sheath rot and seed discoloration of rice by inhibiting its growth at a much lower concentration (ED50 18 ug/ml) than the standard fungicide Carbendazim (ED50 230 ug/ml) in vitro [5]. Discolored rice seeds primed with such NPs, showed a significant reduction in seedling blight and seed rot symptoms with no morphologically visible signs of treatment-induced toxicity. SEM micrographs revealed that FeS-NPs led to mycelial disintegration, disruption of hyphal cell membrane, and inhibition of macroconidial growth of the fungus thereby restricting fungal proliferation and impeding fungal growth [5].
Cu has been used for ages as a bio-control agent owing to its insecticidal and fungicidal properties [13]. Cu NPs are the most affordable among various transition metals under examination like gold, silver & platinum. Cu-NPs are known to penetrate microbial cells and cause damage due to the inactivation of crucial enzymes by substituting essential ions and blocking functional groups of proteins, generating hydro-peroxide free radicals, modifying membrane integrity, and interacting with P and S-containing macromolecules like DNA, RNA, and lipids [13]. CuO NPs have been used as seed priming agents in forage sorghum (Sorghum bicolour), cowpea (Vigna unguiculata) [49] and wheat [87] and have been effective in improving seedling germination traits. In-vitro assays revealed that CuO could potentially inhibit the mycelial growth and spore germination of Alternaria solani (a fungal pathogen causing leaf blight in wheat) [87] and seed-borne fungal pathogens of sorghum and cowpea [49]. It is believed that CuO NP primed seeds will be more resistant to pathogen attack than unprimed seeds; however, the lack of in-vivo trials fails to validate the hypothesis in these studies.
Advantages and disadvantages of nano-priming
Advantages
Nano-priming indeed possesses the potential to steer a new era of crop productivity and sustainable agriculture. Studies reveal a plethora of advantages bestowed by nano-priming over conventional agricultural practices. Plants are known to provide a potential pathway for NP transportation that closely mimics that of endogenous nutrients. Besides the fact that the binding properties of seeds and nano-priming agents are higher than those of traditional priming agents like water, PEG, results in better seed water uptake in nano-primed seeds, resulting in rapid and synchronized germination ensuring a stable and successful crop stand. Nano-priming assures targeted and smart delivery of the nutrients or desired molecules to the seeds that control and minimize the harmful effect of such chemicals on the environment or the soil microflora. Nano-priming agents possess some unique physical, chemical, and mechanical properties that make them electrochemically more active than bulk materials of the same composition. Due to the presence of a higher proportion of atoms on the surface of nanomaterials, they tend to differ in surface composition and reactivity from their bulk counterparts. Another striking feature of NPs is their requirement in trace amounts and their ability to pass through biological membranes and interact with biomolecules owing to their minute sizes. Nano-priming has been found to influence biochemical pathways, reactive oxygen compound equilibrium, and plant growth hormones, thereby impacting the production of phytochemicals in plants [24]. Research has demonstrated that nano-priming can modulate physiological responses, including antioxidant systems and hormonal pathways, leading to enhanced stress resistance and disease resistance in plants [43]. Additionally, nano-priming has been associated with increased production of secondary phytochemicals, suggesting a potential for enhancing the natural production of these compounds in plants. For instance, priming Momordica Charantia L. seeds with ZnONPs has been speculated to upregulate the expression of genes associated with the shikimate phenylpropanoid pathway which elevates the phenolic and flavonoid content [51]. The positive effects of nano-priming on agriculture have been elucidated by field trials which have demonstrated the nano primed seeds as superior to hydro primed seeds owing to their feasibility in promoting yield as well as nutritional value of grains with negligible effect on soil microbiota [29].
Disadvantages
Nano-priming has been considered to be a prudent alternative to foliar or root application of nanoparticles owing to their minimized chances of environmental exposure. However, it is imperative to assess the optimal concentration and duration of treatment in seeds to ensure maximum growth, development of plants and eliminate chances of interference with beneficial soil microbial community. The adverse effects of nano-priming on soil microbiota have been explained by the significant alterations in nitrogen fixation, phosphate solubilization, downregulation in enzyme activities, and factors associated with soil fertility. The interaction and associated risks of nano priming with human and animal health have been a matter of prime importance and are debatable. It is essential to constantly monitor whether the nano-priming agents can impose any negative implications in future but the lack of any rules or standards has made this task perplexing. Nano-priming has been well acknowledged for its feasibility in augmenting the synthesis of secondary metabolites to defend the pathogens. Nevertheless, this approach may lead to the release of other phytochemicals which may protect the plant against pathogens but simultaneously can have adverse effects on human health [71]. Various priming approaches including UV-B [66], Plant growth promoting rhizobacteria (PGPR) [11], Trichoderma [78], INA [50], etc. have been substantiated to have a profound trans-generational impact on plants which include major transcriptional, post-translational and epigenetic changes. These trans-generational changes create a memory that enables the plants to enhance their growth and vigor as well as act as a defense system against abiotic and biotic stress factors. Priming rice seeds with AgNPs has also been revealed to create a defense memory against salinity stress and rice blast pathogen Magnaporthe oryzae [84]. AgNPs create this defense memory against the pathogen by triggering a significant alteration in the glutathione metabolism, MAPK and phytohormone signaling pathway associated with defense in addition to secondary metabolites like falvones, and flavonol. However, transcriptional changes in these signal transduction pathways persisted for only a few weeks. Lately, carbohydrate-coated CeO2 NPs have been confirmed to have a trans-generational impact on Chenopodium rubrum L. and Sinapis alba L. seeds which were substantiated by elevated seed germination and enhanced morphological as well as physiological characteristics in subsequent generations [52].
Future perspectives and conclusion
The seed priming technique is innovative, cheap, and easy to apply in farmer’s field conditions. Also, many successful case studies have been reported regarding seed priming for the amelioration of biotic and abiotic stress to date. However, we have yet to understand the molecular mechanism of seed priming. Future research must be oriented toward identifying the underlying mechanism of nano seed priming more vividly to popularize its application. Besides, the use of mono-metallic NPs has been taken over by the use of multi-metallic NPs [15] owing to their enhanced benefits. Such NPs can be applied as priming agents to develop multifaceted outcomes. Proper field trials, elucidation of a detailed mode of action for the NP-mediated response, and depiction of the environmental consequence of NPs application are the need of the moment. Thus, an orchestrated approach is required to render nano seed priming as an effective and promising strategy to replace the use of bulk synthetic pesticides to I protect the crop plants against biotic agents.
References
Abdel-Aziz HMM, Rizwan M. Chemically synthesized silver nanoparticles induced physio-chemical and chloroplast ultrastructural changes in broad bean seedlings. Chemosphere. 2019;235:1066–72.
Abou-Zeid H, Ismail G. The role of priming with biosynthesized silver nanoparticles in the response of Triticum aestivum L to salt stress. Egypt J Bot. 2018;58:73–85.
Acharya P, Jayaprakasha GK, Crosby KM, Jifon JL, Patil BS. Green-synthesized nanoparticles enhanced seedling growth, yield, and quality of onion (Allium cepa L.). ACS Sustain Chem Eng. 2019;7:14580–90.
Adetunji AE, Adetunji TL, Varghese B, Sershen PNW. Oxidative stress, ageing and methods of seed invigoration: an overview and perspectives. Agronomy. 2021;11:2369.
Ahuja R, Sidhu A, Bala A. Synthesis and evaluation of iron (ii) sulfide aqua nanoparticles (FeS-NPs) against Fusarium verticillioides causing sheath rot and seed discoloration of rice. Eur J Plant Pathol. 2019;155:163–71.
Banerjee S, Islam J, Mondal S, Saha A, Saha B, Sen A. Proactive attenuation of arsenic-stress by nano-priming: zinc oxide nanoparticles in Vigna mungo (L.) hepper trigger antioxidant defense response and reduce root-shoot arsenic translocation. J Hazard Mater. 2023;446:130735. https://doi.org/10.1016/j.jhazmat.2023.130735.
Chen J, Zeng X, Yang W, Xie H, Ashraf U, Mo Z, Liu J, Li G, Li W. Seed priming with multiwall carbon nanotubes (MWCNTs) modulates seed germination and early growth of maize under cadmium (Cd) toxicity. J Soil Sci Plant Nutr. 2021;21:1793–805.
Chen N, Yao P, Zhang W, Zhang Y, Xin N, Wei H, Zhang T, Zhao C. Selenium nanoparticles: enhanced nutrition and beyond. Crit Rev Food Sci Nutr. 2023;63:12360–71.
Choudhary RC, Kumaraswamy RV, Kumari S, Sharma SS, Pal A, Raliya R, Biswas P, Saharan V. Cu-chitosan nanoparticle boost defense responses and plant growth in maize (Zea mays L.). Sci Rep. 2017;7:9754.
Choudhary RC, Kumaraswamy RV, Kumari S, Sharma SS, Pal A, Raliya R, Biswas P, Saharan V. Zinc encapsulated chitosan nanoparticle to promote maize crop yield. Int J Biol Macromol. 2019;127:126–35.
Devi B, Tiwari M, Yadav N, Singh P. Intergenerational immune priming: harnessing plant growth promoting rhizobacteria (PGPR) for augmented wheat protection against spot blotch. Physiol Mol Plant Pathol. 2023;128: 102164.
Devkota KP, Devkota M, Rezaei M, Oosterbaan R. Managing salinity for sustainable agricultural production in salt-affected soils of irrigated drylands. Agric Syst. 2022;198: 103390.
Dey S, Biswas A, Kundu R, Paul S. Role of Copper in Tolerance Against Different Environmental Stress. In Biology and Biotechnology of Environmental Stress Tolerance in Plants 2024 (pp. 351-386). Apple Academic Press
Dey S, Kundu R, Gopal G, Mukherjee A, Nag A, Paul S. Enhancement of nitrogen assimilation and photosynthetic efficiency by novel iron pulsing technique in Oryza sativa L. var Pankaj. Plant Physiol Biochem. 2019;144:207–21. https://doi.org/10.1016/j.plaphy.2019.09.037.
Dey S, Nath S, Ansari TA, Biswas A, Barman F, Mukherjee S, Gopal G, Bhattacharyya A, Mukherjee A, Kundu R. Application of green synthesized bimetallic nZVI-Cu nanoparticle as a sustainable alternative to chemical fertilizers to enhance growth and photosynthetic efficiency of rice seedlings. Plant Physiol Biochem. 2023;201:107837.
El-Badri AM, Batool M, Mohamed IAA, Wang Z, Wang C, Tabl KM, Khatab A, Kuai J, Wang J, Wang B. Mitigation of the salinity stress in rapeseed (Brassica napus L) productivity by exogenous applications of bio-selenium nanoparticles during the early seedling stage. Environ Pollut. 2022;310:119815.
El-Badri AM, Batool M, Wang C, Hashem AM, Tabl KM, Nishawy E, Kuai J, Zhou G, Wang B. Selenium and zinc oxide nanoparticles modulate the molecular and morpho-physiological processes during seed germination of Brassica napus under salt stress. Ecotoxicol Environ Saf. 2021;225: 112695.
El-Badri AMA, Batool M, Mohamed IAA, Khatab A, Sherif A, Wang Z, Salah A, Nishawy E, Ayaad M, Kuai J. Modulation of salinity impact on early seedling stage via nano-priming application of zinc oxide on rapeseed (Brassica napus L.). Plant Physiol Biochem. 2021;166:376–92.
El-Beltagi HS, Ahmed OK, Hegazy AE. Molecular role of nitric oxide in secondary products production in Ginkgo biloba cell suspension culture. Not Bot Horti Agrobot Cluj-Napoca. 2015;43:12–8.
do Espirito Santo Pereira A, Caixeta Oliveira H, Fernandes Fraceto L, Santaella C. Nanotechnology potential in seed priming for sustainable agriculture. Nanomaterials. 2021 Jan 20;11(2):267.
Etesami H, Fatemi H, Rizwan M. Interactions of nanoparticles and salinity stress at physiological, biochemical and molecular levels in plants: a review. Ecotoxicol Environ Saf. 2021;225: 112769. https://doi.org/10.1016/j.ecoenv.2021.112769.
Fahad S, Hussain S, Matloob A, Khan FA, Khaliq A, Saud S, Hassan S, Shan D, Khan F, Ullah N. Phytohormones and plant responses to salinity stress: a review. Plant Growth Regul. 2015;75:391–404.
Farooq T, Nisa ZU, Hameed A, Ahmed T, Hameed A. Priming with copper-chitosan nanoparticles elicit tolerance against PEG-induced hyperosmotic stress and salinity in wheat. BMC Chem. 2022;16:23.
Garg R, Maheshwari S. Seed priming and fortification of seeds using nanotechnology: a review. EPH-Int J Agric Environ Res. 2023;9:11–7.
Gomes DG, Pelegrino MT, Ferreira AS, Bazzo JHB, Zucareli C, Seabra AB, Oliveira HC. Seed priming with copper-loaded chitosan nanoparticles promotes early growth and enzymatic antioxidant defense of maize (Zea mays L.) seedlings. J Chem Technol Biotechnol. 2021;96:2176–84.
González-García Y, López-Vargas ER, Pérez-Álvarez M, Cadenas-Pliego G, Benavides-Mendoza A, Valdés-Reyna J, Pérez-Labrada F, Juárez-Maldonado A. Seed priming with carbon nanomaterials improves the bioactive compounds of tomato plants under saline stress. Plants. 2022;11:1984.
Goswami P, Mathur J, Srivastava N. Silica nanoparticles as novel sustainable approach for plant growth and crop protection. Heliyon. 2022;8:e09908.
Guha T, Barman S, Mukherjee A, Kundu R. Nano-scale zero valent iron modulates Fe/Cd transporters and immobilizes soil Cd for production of Cd free rice. Chemosphere. 2020;260: 127533. https://doi.org/10.1016/j.chemosphere.2020.127533.
Guha T, Mukherjee A, Kundu R. Nano-scale zero valent iron (nZVI) priming enhances yield, alters mineral distribution and grain nutrient content of Oryza sativa L. cv. Gobindobhog: a field study. J Plant Growth Regul. 2021;41:1–24.
Guha T, Mukherjee A, Kundu R. Nano-scale zero valent iron (nZVI) priming enhances yield, alters mineral distribution and grain nutrient content of Oryza sativa L. cv. Gobindobhog: a field study. J Plant Growth Regul. 2022;41:710–33.
Guha T, Ravikumar KVG, Mukherjee A, Mukherjee A, Kundu R. Nanopriming with zero valent iron (nZVI) enhances germination and growth in aromatic rice cultivar (Oryza sativa cv. Gobindabhog L.). Plant Physiol Biochem. 2018;127:403–13.
Gupta B, Huang B. Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Int J Genomics 2014: 1–18
Hameed A, Maqsood W, Hameed A, Qayyum MA, Ahmed T, Farooq T. Chitosan nanoparticles encapsulating curcumin counteract salt-mediated ionic toxicity in wheat seedlings: an ecofriendly and sustainable approach. Environ Sci Pollut Res. 2024;31(6):1–13.
Hasanuzzaman M, Fujita M. Plant responses and tolerance to salt stress: physiological and molecular interventions. Int J Mol Sci. 2022;23:4810.
Hussain A, Rizwan M, Ali Q, Ali S. Seed priming with silicon nanoparticles improved the biomass and yield while reduced the oxidative stress and cadmium concentration in wheat grains. Environ Sci Pollut Res Int. 2019;26:7579–88. https://doi.org/10.1007/s11356-019-04210-5.
Imtiaz H, Shiraz M, Mir AR, Siddiqui H, Hayat S. Nano-priming techniques for plant physio-biochemistry and stress tolerance. J Plant Growth Regul. 2023;42:1–21.
Jangir H, Bhardwaj A, Das M. Larger root nodules increased Fe, Mo, Mg, P, Ca, Mn, K in the roots and higher yield in chickpea grown from nano FeS2 pre-treated seeds: emulating nitrogenase. Appl Nanosci. 2020;10:445–54.
Kandhol N, Singh VP, Ramawat N, Prasad R, Chauhan DK, Sharma S, Grillo R, Sahi S, Peralta-Videa J, Tripathi DK. Nano-priming: Impression on the beginner of plant life. Plant Stress. 2022;5: 100091.
Karvar M, Azari A, Rahimi A, Maddah-Hosseini S, Ahmadi-Lahijani MJ. Titanium dioxide nanoparticles (TiO2-NPs) enhance drought tolerance and grain yield of sweet corn (Zea mays L.) under deficit irrigation regimes. Acta Physiol Plant. 2022;44:14.
Kasote DM, Lee JHJ, Jayaprakasha GK, Patil BS. Seed priming with iron oxide nanoparticles modulate antioxidant potential and defense-linked hormones in watermelon seedlings. ACS Sustain Chem Eng. 2019;7:5142–51.
Khan I, Raza MA, Awan SA, Shah GA, Rizwan M, Ali B, Tariq R, Hassan MJ, Alyemeni MN, Brestic M. Amelioration of salt induced toxicity in pearl millet by seed priming with silver nanoparticles (AgNPs): the oxidative damage, antioxidant enzymes and ions uptake are major determinants of salt tolerant capacity. Plant Physiol Biochem. 2020;156:221–32.
Khan MR, Siddiqui ZA. Use of silicon dioxide nanoparticles for the management of meloidogyne incognita, Pectobacterium betavasculorum and Rhizoctonia solani disease complex of beetroot (Beta vulgaris L.). Sci Hortic. 2020;265:109211.
Lee JHJ, Kasote DM. Nano-priming for inducing salinity tolerance, disease resistance, yield attributes, and alleviating heavy metal toxicity in plants. Plants. 2024;13:446.
Li R, He J, Xie H, Wang W, Bose SK, Sun Y, Hu J, Yin H. Effects of chitosan nanoparticles on seed germination and seedling growth of wheat (Triticum aestivum L.). Int J Biol Macromol. 2019;126:91–100.
Li Y, Liang L, Li W, Ashraf U, Ma L, Tang X, Pan S, Tian H, Mo Z. ZnO nanoparticle-based seed priming modulates early growth and enhances physio-biochemical and metabolic profiles of fragrant rice against cadmium toxicity. J Nanobiotechnology. 2021;19:1–19.
López-Vargas ER, González-García Y, Pérez-Álvarez M, Cadenas-Pliego G, González-Morales S, Benavides-Mendoza A, Cabrera RI, Juárez-Maldonado A. Seed priming with carbon nanomaterials to modify the germination, growth, and antioxidant status of tomato seedlings. Agronomy. 2020;10:639.
Ma J, Li Y, Chen F, Sun Y, Zhu Y, Wang L. Bacillus mycoides PM35 in combination with titanium dioxide (TiO2)⎯nanoparticles enhanced morpho-physio-biochemical attributes in Barley (Hordeum vulgare L.) under cadmium stress. Chemosphere. 2023;323:138224.
Mahakham W, Sarmah AK, Maensiri S, Theerakulpisut P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosynthesized silver nanoparticles. Sci Rep. 2017;7:8263. https://doi.org/10.1038/s41598-017-08669-5.
Maity A, Natarajan N, Pastor M, Vijay D, Gupta CK, Wasnik VK Nanoparticles influence seed germination traits and seed pathogen infection rate in forage sorghum (Sorghum bicolour) and cowpea (Vigna unguiculata) 2018
Martínez-Aguilar K, Hernández-Chávez JL, Alvarez-Venegas R. Priming of seeds with INA and its transgenerational effect in common bean (Phaseolus vulgaris L.) plants. Plant Sci. 2021;305:110834.
Mazhar MW, Ishtiaq M, Maqbool M, Akram R. Efficacy of zinc oxide nanoparticles on bitter gourd growth, yield, and some phytochemicals through seed priming. Int J Veg Sci. 2023;29:145–55.
Milenković I, Krstić MB, Spasić SZ, Radotić K. Trans-generational effect of cerium oxide-nanoparticles (nCeO2) on Chenopodium rubrum L. and Sinapis alba L. seeds. Funct Plant Biol. 2023;50:303–13.
Moustafa-Farag M, Almoneafy A, Mahmoud A, Elkelish A, Arnao MB, Li L, Ai S. Melatonin and its protective role against biotic stress impacts on plants. Biomolecules. 2019;10:54.
Mukhopadhyay SS. Nanotechnology in agriculture: prospects and constraints. Nanotechnol Sci Appl. 2014;7:63–71.
Nile SH, Thiruvengadam M, Wang Y, Samynathan R, Shariati MA, Rebezov M, Nile A, Sun M, Venkidasamy B, Xiao J. Nano-priming as emerging seed priming technology for sustainable agriculture—recent developments and future perspectives. J Nanobiotechnology. 2022;20:254.
Parveen A, Siddiqui ZA. Impact of silicon dioxide nanoparticles on growth, photosynthetic pigments, proline, activities of defense enzymes and some bacterial and fungal pathogens of tomato. Vegetos. 2022;35:83–93. https://doi.org/10.1007/s42535-021-00280-4.
Paul S, Dey S, Kundu R. Seed priming: an emerging tool towards sustainable agriculture. Plant Growth Regul. 2021;97:1–20.
Prosekov AY, Ivanova SA. Food security: the challenge of the present. Geoforum. 2018;91:73–7.
Qari SH, Tarbiyyah I. The genetic regulation of secondary metabolic pathways in response to salinity and drought as abiotic stresses. Appl Sci. 2021;11:6668.
Rai-Kalal P, Tomar RS, Jajoo A. Seed nanopriming by silicon oxide improves drought stress alleviation potential in wheat plants. Funct Plant Biol. 2021;48:905–15.
Rana K, Luksiene Z, Pudake RN. Recent advances in plant pathogen control by nanocides. Nanoscience for Sustainable Agriculture. 2019:101-37
Rizwan M, Ali S, Ali B, Adrees M, Arshad M, Hussain A, Ur Rehman MZ, Waris AA. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere. 2019;214:269–77.
Santisree P, Bhatnagar-Mathur P, Sharma KK. NO to drought-multifunctional role of nitric oxide in plant drought: do we have all the answers? Plant Sci. 2015;239:44–55.
Savassa SM, Duran NM, Rodrigues ES, De Almeida E, Van Gestel CAM, Bompadre TFV, De Carvalho PHW. Effects of ZnO nanoparticles on Phaseolus vulgaris germination and seedling development determined by X-ray spectroscopy. ACS Appl Nano Mater. 2018;1:6414–26.
Seddighinia FS, Iranbakhsh A, Oraghi Ardebili Z, Nejad Satari T, Soleimanpour S. Seed priming with cold plasma and multi-walled carbon nanotubes modified growth, tissue differentiation, anatomy, and yield in bitter melon (Momordica charantia). J Plant Growth Regul. 2020;39:87–98.
Sen A, Puthur JT, Challabathula D, Brestič M. Transgenerational effect of UV-B priming on photochemistry and associated metabolism in rice seedlings subjected to PEG-induced osmotic stress. Photosynthetica. 2022;60(2):219–29.
Shafiq F, Iqbal M, Ali M, Ashraf MA. Seed pre-treatment with polyhydroxy fullerene nanoparticles confer salt tolerance in wheat through upregulation of H2 O2 neutralizing enzymes and phosphorus uptake. J Soil Sci Plant Nutr. 2019;19:734–42.
Shah T, Latif S, Saeed F, Ali I, Ullah S, Abdullah Alsahli A, Jan S, Ahmad P. Seed priming with titanium dioxide nanoparticles enhances seed vigor, leaf water status, and antioxidant enzyme activities in maize (Zea mays L.) under salinity stress. J King Saud Univ - Sci. 2021;33:101207. https://doi.org/10.1016/j.jksus.2020.10.004.
Sharma A, Kothari S, Kachhwaha S. Effect of multi-walled carbon nanotubes-based seed priming in pennisetum glaucum seedlings affecting the antioxidant activities. J Food Chem Nanotechnol. 2023;9:1–6.
Sharma P, Gautam A, Kumar V, Guleria P. MgO nanoparticles priming promoted the growth of black chickpea. J Agric Food Res. 2022;10: 100435.
Shelar A, Nile SH, Singh AV, Rothenstein D, Bill J, Xiao J, Chaskar M, Kai G, Patil R. Recent advances in nano-enabled seed treatment strategies for sustainable agriculture: challenges, risk assessment, and future perspectives. Nano-Micro Lett. 2023;15:54.
Shelar A, Singh AV, Maharjan RS, Laux P, Luch A, Gemmati D, Tisato V, Singh SP, Santilli MF, Shelar A. Sustainable agriculture through multidisciplinary seed nanopriming: prospects of opportunities and challenges. Cells. 2021;10:2428.
Shukla P, Chaurasia P, Younis K, Qadri OS, Faridi SA, Srivastava G. Nanotechnology in sustainable agriculture: studies from seed priming to post-harvest management. Nanotechnol Environ Eng. 2019;4:11. https://doi.org/10.1007/s41204-019-0058-2.
Siddaiah CN, Prasanth KVH, Satyanarayana NR, Mudili V, Gupta VK, Kalagatur NK, Satyavati T, Dai X-F, Chen J-Y, Mocan A. Chitosan nanoparticles having higher degree of acetylation induce resistance against pearl millet downy mildew through nitric oxide generation. Sci Rep. 2018;8:2485.
Siddiqui ZA, Hashmi A, Khan MR, Parveen A. Management of bacteria Pectobacterium carotovorum, Xanthomonas campestris pv. carotae, and fungi Rhizoctonia solani, Fusarium solani and Alternaria dauci with silicon dioxide nanoparticles on carrot. Int J Veg Sci. 2020;26:547–57.
Slavin YN, Asnis J, Hńfeli UO, Bach H. Metal nanoparticles: understanding the mechanisms behind antibacterial activity. J Nanobiotechnology. 2017;15:1–20.
Sundaria N, Singh M, Upreti P, Chauhan RP, Jaiswal JP, Kumar A. Seed priming with iron oxide nanoparticles triggers iron acquisition and biofortification in wheat (Triticum aestivum L.) grains. J Plant Growth Regul. 2019;38:122–31.
Tiwari M, Singh R, Jha R, Singh P. Heritable priming by Trichoderma: a sustainable approach for wheat protection against Bipolaris sorokiniana. Front Plant Sci. 2022;13:1050765.
Ul Haq T, Ullah R, Khan MN, Nazish M, Almutairi SM, Rasheed RA. Seed priming with glutamic-acid-functionalized iron nanoparticles modulating response of Vigna radiata (L.) R. Wilczek (mung bean) to induce osmotic stress. Micromachines. 2023;14:736.
Ullah R, Ullah Z, Iqbal J, Chalgham W, Ahmad A. Aspartic Acid-based nano-copper induces resilience in zea mays to applied lead stress via conserving photosynthetic pigments and triggering the antioxidant biosystem. Sustainability. 2023;15:12186.
Wani IA, Ahmad T. Size and shape dependant antifungal activity of gold nanoparticles: a case study of Candida. Colloids surfaces B Biointerfaces. 2013;101:162–70.
Xiao F, Zhou H. Plant salt response: perception, signaling, and tolerance. Front Plant Sci. 2023;13:1053699.
Xin X, Zhao F, Rho JY, Goodrich SL, Sumerlin BS, He Z. Use of polymeric nanoparticles to improve seed germination and plant growth under copper stress. Sci Total Environ. 2020;745: 141055.
Yadav N, Bora S, Devi B, Upadhyay C, Singh P. Nanoparticle-mediated defense priming: a review of strategies for enhancing plant resilience against biotic and abiotic stresses. Plant Physiol Biochem. 2024;213:108796.
Yang Z, Zhi P, Chang C. Priming seeds for the future: Plant immune memory and application in crop protection. Front Plant Sci. 2022;13: 961840.
Ye Y, Cota Ruiz K, Hernandez-Viezcas J, Valdés C, Medina Velo I, Turley R, Peralta-Videa J, Gardea-Torresdey J. Manganese nanoparticles control salinity-modulated molecular responses in Capsicum annuum L. through priming: a sustainable approach for agriculture. ACS Sustain Chem Eng. 2020;8(3):1427–36. https://doi.org/10.1021/acssuschemeng.9b05615.
Zakharova O, Kolesnikov E, Shatrova N, Gusev A. The effects of CuO nanoparticles on wheat seeds and seedlings and Alternaria solani fungi: in vitro study. InIOP conference series: earth and environmental science 2019 Feb 19 (Vol. 226, p. 012036). IOP Publishing
Acknowledgements
NG acknowledges SVMCM for her fellowship. SD (CSIR SPM-Sr) acknowledges CSIR for her fellowship and contingency grant. TG acknowledges SERB-NPDF (File no: PDF/2023/003304) for her fellowship. Infrastructural and institutional support from the University of Calcutta is gratefully acknowledged.
Author information
Authors and Affiliations
Contributions
RK conceptualized the study; NG, SD, TG, and SP helped in primary draft preparation. Manuscript review and editing were done by RK, NG and SD. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
All authors have read and approved the manuscript. The authors declare that there is no conflict of interest. RK is the member of the Editorial Board, but was not involved in decision process at any stage.
Additional information
This article is dedicated to Prof. Arun Kumar Sharma to commemorate his Birth Centenary.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Corresponding Editor: Anita Mukherjee; Reviewers: Swarupa Ghosh, Ilika Ghosh.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ghosh, N., Dey, S., Guha, T. et al. Recent advances in nano-priming induced plant growth promotion and environmental stress tolerance. Nucleus (2024). https://doi.org/10.1007/s13237-024-00506-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13237-024-00506-1