
Abstract
Matrix games under time constraints represent a natural extension of conventional matrix games. They take into account the additional factor that, apart from the payoff, a pairwise interaction results in a delay for the contestants before they can engage in subsequent interactions. Each matrix game can be associated with a continuous dynamical system, known as the replicator equation, which describes the evolution of phenotype frequencies within the population. One of the fundamental theorems of evolutionary matrix games asserts that the state corresponding to an evolutionarily stable strategy is an asymptotically stable rest point of the replicator equation (Taylor and Jonker in Math Biosci 40:145–156, https://doi.org/10.1016/0025-5564(78)90077-9, 1978; Hofbauer et al. in J Theor Biol 81:609–612, https://doi.org/10.1016/0022-5193(79)90058-4, 1979; Zeeman in Global theory of dynamical systems. Lecture notes in mathematics. Springer, New York, vol 819. https://doi.org/10.1007/BFb0087009, 1980). Garay et al. (J Math Biol 76:1951–1973, https://doi.org/10.1086/681638, 2018) and Varga et al. (J Math Biol 80:743–774, https://doi.org/10.1007/s00285-019-01440-6, 2020) generalized the statement to two-strategy and, in some particular cases, three- or more strategy matrix games under time constraints. However, the general applicability of this implication has remained an open question. In this paper, we present examples that demonstrate the negative answer. Moreover, we illustrate, through the rock-paper-scissors game, that even slight disparities in waiting times can lead to the destabilization of the equilibrium corresponding to an ESS. Additionally, we establish that a stable limit cycle can emerge around the unstable equilibrium in a supercritical Hopf bifurcation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Classic evolutionary matrix games, introduced by Maynard Smith and Price [29], describe populations where individuals compete with each other through pairwise interactions (Maynard Smith [28], Chapters 6-7 in Hofbauer and Sigmund [20], Chapter 2.3 in MestertonGibbons [30], Chapter 6 in Broom and Rychtár̆ [6]) . After an interaction, contestants receive payoffs based on their strategies/phenotypes. The payoffs of the consecutive interactions then determine the (relative) fitness and thereby the evolutionary success of a given phenotype. However, in many situations, the outcome of an interaction involves not only a payoff, but also a time period during which contestants must wait before being ready for subsequent interactions (e.g. to recover from an injury, to handle the payoff (food), to collect energy after a long fight, etc., Holling [22], Charnov [8], Broom et al. [5], Garay et al. [15], Toupo et al. [41]). Therefore, a portion of the population is temporarily unable to interact, leading to no resource intake during the waiting period. This can significantly influence fitness and, consequently, evolutionary outcomes [16, 25]. For instance, in the classical Prisoner’s dilemma, the ESS strategy is the defector strategy. However, if the waiting time associated with interactions among defectors is long enough, the average intake of defectors may become smaller than that of cooperators, resulting in the evolutionary stability of the cooperator strategy in that model. Similarly, if the waiting time after a Hawk-Hawk interaction in the Hawk-Dove game is sufficiently long, the pure Dove strategy can be ESS even at a low cost of fighting. These examples well illustrate how the consideration of time constraints in game-theoretical models can lead to surprising phenomena, underscoring the need for further mathematical investigations.
Maynard Smith and Price [29] studied monomorphic populations, where each individual initially possesses the same (pure or mixed) phenotype. In this scenario, Maynard Smith introduced the following static definition in plain language: “An ESS is a strategy such that, if all the members of a population adopt it, then no mutant strategy could invade the population under the influence of natural selection” [28, p. 10]. This condition implies that the ESS is consistently favoured by natural selection when the frequency of mutants is low.
The concept of ESS aims to encapsulate the intuitive expectation that an evolutionarily stable strategy must be resistant to invasion. This static definition proves invaluable in applied evolutionary game theory, as it allows us to predict evolutionary outcomes without delving into complex models describing temporal population changes. However, from a theoretical perspective in evolutionary game theory, we require dynamics that elucidate the temporal evolution of the population (cf. Introduction in Cressman [10]). To address this, Taylor and Jonker [36] proposed a polymorphic approach, considering populations composed of individuals with N distinct, genetically fixed pure phenotypes. The temporal evolution of the population is then modelled using dynamics, often the replicator equation [20, 36], and a locally asymptotically stable rest point of the dynamics serves as the end point of the evolution.
It is a fundamental result in classic evolutionary game theory that, for matrix games, the corresponding state of an ESS is a locally asymptotically stable rest point of the replicator equation [21, 36, 44]. This relationship holds not only for matrix games but also for some cases where the fitness function is not bilinear in the phenotype of the focal individual and the population’s average strategy. In fact, assuming linearity only in the strategy of the focal individual is sufficient (see, for example, Thomas [37], Chapter 7.2 in Hofbauer and Sigmund [20], or Cressman et al. [11]). However, for other fitness functions, the validity of an analogous theorem remains an open question.
In the case of evolutionary matrix games under time constraints, the fitness function is neither linear in the strategy of the focal individual nor in the average strategy of the population. Similarly to the aforementioned classic result, Garay et al. [17] managed to prove the asymptotic stability of the rest point corresponding to an ESS for two-strategy games (Corollary 4.3 in Garay et al. [17]). The proof relies on the fact that the set of the convex combinations of the strategies for which the replicator equation is considered is one-dimensional. However, the higher-dimensional case remained open, except for some special cases (Theorem 4.1 in Garay et al. [17], Theorem 4.7 and Theorem 4.9 in Varga et al. [42]) where the corresponding state is either asymptotically stable or at least stable. Nevertheless, the authors’ conjecture was that the stability does not hold in general.
In this paper, we confirm that the conjecture is true by providing examples of matrix games under time constraints with an evolutionarily stable strategy such that the corresponding state of the replicator equation is unstable, even in the case of three-dimensional strategies.
One of our examples starts from the rock-paper-scissors game. This game models cyclic dominance [24, 27, 35] that can result in the cyclic coexistence of three phenotypes [20, 31, 32, 41]. In classic matrix games, the replicator dynamics cannot exhibit limit cycles ([3, 44], Chapter 7.5 in Hofbauer and Sigmund [20]). If periodic solutions exist, there are infinitely many, enveloping a (non-asymptotically) stable equilibrium point, much like in the classical rock-paper-scissors game. Moreover, if we replace the zeros along the main diagonal of the payoff matrix of this game with a parameter \(\varepsilon \), the phase portrait corresponds to a degenerate Hopf bifurcation, as the sign of \(\varepsilon \) changes [13, 20]. If \(\varepsilon <0\), the equilibrium point is an asymptotically stable focus. As \(\varepsilon \) reaches zero, periodic solutions emerge, and the equilibrium point turns into a non-asymptotically stable center. For \(\varepsilon >0\), the periodic orbits vanish again, and the equilibrium point becomes an unstable focus. However, the corresponding monomorphic strategy also loses its evolutionary stability. With increasing the mathematical complexity of the model, one can anticipate a more intricate behaviour of the replicator dynamics. Mobilia [31] and Toupo and Strogatz [40] pointed out that even allowing arbitrarily small mutations can induce a limit cycle in a Hopf bifurcation. Here, we demonstrate that arbitrary small differences in waiting times can also have a comparable effect, leading not only to the instability of the equilibrium point but also to the appearance of a limit cycle during a Hopf bifurcation. All this occurs while the strategy corresponding to the equilibrium point maintains its evolutionary stability throughout.
2 Preliminaries
We introduce some functions and notation necessary to our calculations. For detailed background (the heuristic thoughts and the mathematical derivation of the formulas), we refer the reader to Garay et al. [16] and Garay et al. [17].
We examine the relationship between the evolutionary stability of game theory and the stability of the replicator dynamics. From a population biology perspective, the dynamic description is applied to a population in which a finite but arbitrarily large number of pre-defined phenotypes can occur. This is known as the polymorphic approach. In contrast, the concept of evolutionary stability applies to a monomorphic population consisting of individuals with a distinguished resident phenotype, and we investigate what happens when a mutant phenotype appears. Only one type of mutant can appear at a time, but it can be any phenotype that differs from the resident.
2.1 Polymorphic Approach
First, we introduce the toolkit for the polymorphic approach. Consider a well-mixed large population in which individuals compete with each other through pairwise interactions that can be mathematically described with the help of a game. During the game, a player chooses from among N pure strategies with certain probabilities determined by the individual’s (heritable) phenotype, in other words, his strategy. Mathematically, we describe the set of possible strategies (phenotypes) using the N-dimensional simplex:
(where N is a positive integer and \({\mathbb {R}}\) is the set of real numbers). Then, the pure strategies correspond to the vectors \(\textbf{e}_1=(1,0,\dots ,0)\), \(\textbf{e}_2=(0,1,0,\dots ,0)\), ..., \(\textbf{e}_{N-1}=(0,0,\dots ,0,1,0)\), \(\textbf{e}_N=(0,0,\dots ,0,1)\), and the coordinate \(q_i\) of a strategy \(\textbf{q}\) is the probability that an individual with phenotype \(\textbf{q}\) employs the pure strategy \(\textbf{e}_i\) in a game.
Let \(\textbf{q}_1,\textbf{q}_2,\dots ,\textbf{q}_n\) denote the phenotypes occurring in the population, with frequencies \(x_1, x_2, \dots , x_n\) respectively. From an evolutionary perspective, a central question is how the state of the population, represented by the frequency distribution \(\textbf{x}=(x_1, x_2, \dots , x_n)\), changes over time. For a mathematical description of this, Taylor and Jonker [36] introduced the replicator equation or dynamics:
where \(W_i(\textbf{x})\) is the fitness of phenotype \(\textbf{q}_i\) (Taylor and Jonker [36] called it "current growth rate"), and \(\bar{W}(\textbf{x}) = x_1W_1(\textbf{x}) + \dots + x_nW_n(\textbf{x})\) denotes the average fitness of the population. The dynamics assume that the change in frequency of a phenotype is proportional to its relative fitness.
To proceed, we need to specify the mathematical form of the \(W_i\) fitness functions. Let \(a_{ij}\) denote the payoff to a player using pure strategy i while their opponent employs pure strategy j. This yields an \(N\times N\) matrix \(A=(a_{ij})\), which we refer to as the payoff matrix. With this, the expected payoff for an individual of phenotype \(\textbf{q}_i\) in the above population is \(\textbf{q}_i A (x_1 \textbf{q}_1+\dots +x_n\textbf{q}_n)\)Footnote 1 (we assume that the population is large enough to neglect the effect that an individual cannot interact with itself). In the case of evolutionary matrix games, this expected payoff also serves as the fitness function \(W_i(\textbf{x})\).
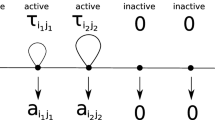
Following the model of Garay et al. [16] or Kr̆ivan and Cressman [25], we also consider that an interaction has another consequence in addition to the payoff. Specifically, if a player uses pure strategy i against an opponent using pure strategy j, then the player must wait for \(\tau _{ij}\) time units before being able to engage in another interaction. (The time of the interaction itself can be considered as 0 or a part of the \(\tau _{ij}\) time.) During this waiting time, the individual is incapable of interaction and is considered inactive from the perspective of the game. One can think of this time as necessary for the individual to process the resources acquired during the interaction or for the individual to recover from any injuries sustained (cf. Holling [22] or the handling time in optimal foraging theory, Charnov [8] and Garay et al. [14]). After the waiting period, the player looks for their next opponent. We consider the time taken for this search to be 1. During the search, the player is capable of interaction and is considered active in terms of the game. If the player encounters an individual who is inactive, no interaction occurs between them, resulting in a payoff of 0, but the search time restarts. It is worth noting that the model of Garay et al. [16] is a Markov model in which both the waiting times \(\tau _{ij}\) and the search time are independent exponentially distributed random variables. However, we will only use derived formulas in which only their expected values (\(\tau _{ij}\) and 1, respectively) appear. We assume that the sequence of interactions and searches occurs on a much faster time scale than the replicator dynamics (cf. Gunawardena [19]), allowing us to assume, from the perspective of replicator dynamics, that the population is in a stationary equilibrium. In other words, on the fast time scale, the proportion of inactive individuals is constant, and the ratio of time spent in an active state for an individual of type \(\textbf{q}_i\) matches the proportion of active individuals in the subpopulation of \(\textbf{q}_i\) individuals. Formally, if we denote this ratio by \(\varrho _i\) and \(T=(\tau _{ij})_{N\times N}\), then:
The denominator represents the expected value of time between two encounters, while the numerator represents the time spent searching (in the active state). Matrix T is referred to as the time constraint matrix. It can be shown that there is only one solution to the system of equations above for which \(0\le \varrho _i\le 1\) for every i (Theorem 1 and Lemma 2 in Garay et al. [16]).
Since there is no interaction in the inactive state, identifying fitness solely on the basis of expected payoffs could give a false picture. Even if the expected payoff is high, it can be misleading if such events rarely occur. To obtain an accurate representation, we must also consider the time between interactions. Therefore, we interpret fitness as the intake per unit of time:
Note that the term in the numerator is just the expected value of payoff in a single interaction, whereas the denominator equals the expected value of time between two consecutive encounters. Theorem 1 in Garay et al. [16] claims that if first the time of observation, then the population size tends to infinity, while the proportion of individuals of type \(\textbf{q}_i\) converges to \(x_i\), then the average intake per unit of time for an individual of type \(\textbf{q}_i\) is described by the above formula. This demonstrates that our formula used for the \(W_i\) fitness can be derived not only intuitively but also rigorously through mathematical means.Footnote 2
If we introduce the notation
and
(the former representing the proportion of active individuals in the population, the latter representing the average strategy of active individuals in the population), then \(W_i\) and \({\bar{W}}\) can be rewritten in the following forms:
and
respectively.
This shows that our model is indeed a generalization of classical evolutionary matrix games, since if all \(\tau _{ij}\) times are zero, then \(\varrho _i(\textbf{x})\textbf{q}_iA\bar{\varrho }(\textbf{x})\textbf{h}(\textbf{x})\) simplifies to the formula \(\textbf{q}_iA\sum _i x_i\textbf{q}_i\) used to express fitness in the classical case.
2.2 Monomorphic Approach and Evolutionarily Stable Strategy
We will need a precise mathematical formulation of evolutionary stability under time constraints and further statements that connect it with the stability of replicator dynamics.
Consider a monomorphic population of individuals with \(\textbf{p}\) phenotype. Assume that some mutant individuals with \(\textbf{q}\) phenotype appear in the population. We should stress that only one type of mutant phenotype can be present in the population at a time. This type is denoted by \(\textbf{q}\). Let \(1-\varepsilon \) and \(\varepsilon \) be the frequencies of the resident and mutant phenotypes, respectively. This scenario mathematically corresponds to the above polymorphic situation when \(n=2\), \(\textbf{q}_1=\textbf{p}\), \(\textbf{q}_2=\textbf{q}\), \(x_1=1-\varepsilon \), and \(x_2=\varepsilon \). However, to emphasize that \(\textbf{p}\) is the resident and \(\textbf{q}\) is the mutant phenotype, we denote the fitness of \(\textbf{p}\) as \(\omega _{\textbf{p}}\) and the fitness of \(\textbf{q}\) as \(\omega _{\textbf{q}}\). Accordingly,
where \(\rho _{\textbf{p}}(\textbf{p},\textbf{q},\varepsilon )\) and \(\rho _{\textbf{q}}(\textbf{p},\textbf{q},\varepsilon )\) are the proportions of active individuals among the \(\textbf{p}\) and \(\textbf{q}\) individuals, respectively, and they constitute the unique solution of the equation system
in the interval [0, 1] (Lemma 2 in Garay et al. [16]).
Definition 2.1
A strategy \(\textbf{p}^*\) is a uniformly evolutionary stable strategy of the matrix game under time constraints (UESS in short) if there is an \(\varepsilon _0>0\) such that
for any strategy \(\textbf{q}\not =\textbf{p}^*\) and \(0<\varepsilon <\varepsilon _0\).
The adverb “uniformly” is used here because \(\varepsilon _0\) does not depend on \(\textbf{q}\). When \(\varepsilon _0\) can depend on \(\textbf{q}\), we refer to \(\textbf{p}^*\) as an ESS without the adverb "uniformly". Clearly, if \(\textbf{p}^*\) is a UESS, it also satisfies the criteria for an ESS. The converse also holds in the case of classic matrix games (without time constraints), as shown in Theorem 6.4.1 in Hofbauer and Sigmund [20]. However, in the context of matrix games under time constraints, this equivalence remains an open question. Although we do not want to deal with this issue in this article, our aim was to explain the rationale behind employing the term "UESS". This distinction between UESS and ESS naturally arises when dealing with non-linear fitness functions [4], and UESS is considered more suitable for describing evolutionary stability in such cases [33].
Our primary objective is to present examples featuring a UESS \(\mathbf {p^*}\) where the corresponding state is an unstable rest point of the replicator equation with respect to pure strategies. Nevertheless, describing this corresponding state is not as straightforward as in the classical scenario. We must reference certain pivotal statements to grasp the connection between the monomorphic and polymorphic models.
2.3 Relationship Between the Polymorphic and the Monomorphic Approach
We introduce the shorter notations \(\omega (\textbf{q})\), \(\omega _{\textbf{p}}(\textbf{q})\), \(\rho (\textbf{q})\) and \(\rho _{\textbf{p}}(\textbf{q})\) for \(\omega _{\textbf{q}}(\textbf{p},\textbf{q},1)\), \(\omega _{\textbf{p}}(\textbf{p},\textbf{q},1)\), \(\rho _{\textbf{q}}(\textbf{p},\textbf{q},1)\) and \(\rho _{\textbf{p}}(\textbf{p},\textbf{q},1)\), respectively. These special quantities describe the initial moment when individuals of type \(\textbf{p}\) start to appear in a population composed exclusively of individuals of type \(\textbf{q}\). \(\rho (\textbf{q})\) can be explicitly expressed as the unique solution in [0, 1] to the equation
(Lemma 2 in Garay et al. [16]). In particular,
This immediately implies that functions
and
all have continuous second-order partial derivatives with respect to the coordinates of \(\textbf{q}=(q_1, q_2, \dots ,q_N) \in S_N\).
It is clear for classical matrix games that if \(\textbf{p}=(p_1,p_2,\dots ,p_N)\) is a strategy then \((x_1,x_2,\dots ,x_N)\) with \(x_i=p_i\) is the corresponding state in the polymorphic system of pure phenotypes \(\textbf{e}_1,\textbf{e}_2,\dots ,\textbf{e}_N\) in the sense that \(\sum _i x_i\textbf{e}_i=\textbf{p}\). Consequently, if \(\textbf{p}\) is an ESS, then it is natural to investigate the stability property of the state \((x_1,x_2,\dots ,x_N)\) with \(x_i=p_i\) for the replicator equation with respect to \(\textbf{e}_1,\textbf{e}_2,\dots ,\textbf{e}_N\). If the replicator equation is considered for arbitrary strategies \(\textbf{q}_1,\textbf{q}_2,\dots ,\textbf{q}_n\) then it is also clear that the corresponding state(s) of \(\textbf{p}\) is (are) the state(s) \((x_1,x_2,\dots ,x_n)\) with \(\textbf{p}=x_1\textbf{q}_1+x_2\textbf{q}_2+\dots +x_n\textbf{q}_n\), that is, the state(s) at which the average strategy of the population is \(\textbf{p}\). However, for matrix games under time constraints, the situation is generally not so obvious. The answer can be found in the next lemma of Garay et al. [17], which asserts that the corresponding state is the state at which the average strategy of active individuals is \(\textbf{p}\). To recall notations \(\textbf{h}(\textbf{x})\) and \({\bar{\varrho }}(\textbf{x})\), see (2.4) and (2.5).
Lemma 2.2
(Garay et al. [17], Corollary 6.3) Consider a population of phenotypes \(\textbf{q}_1\), \(\textbf{q}_2\),...,\(\textbf{q}_n\in S_N\). Assume that \(\textbf{r}=\theta _1\textbf{q}_1+ \theta _2\textbf{q}_2+\dots +\theta _n\textbf{q}_n\) with some \(\varvec{\theta }=(\theta _1,\theta _2,\dots ,\theta _n)\in S_n\). If
then \(\textbf{h}(\textbf{x})=\textbf{r}\) where \(\textbf{x}=\textbf{x} (\varvec{\theta })=\big (x_1(\varvec{\theta }),x_2(\varvec{\theta }), \dots ,x_n(\varvec{\theta })\big )\), that is, the average strategy of active individuals in a population of \(\textbf{q}_1\), \(\textbf{q}_2\),..., \(\textbf{q}_n\) individuals with frequencies \(x_1(\varvec{\theta }),\) \(x_2(\varvec{\theta })\),..., \(x_n(\varvec{\theta })\) is \(\theta _1 \textbf{q}_1+ \theta _2\textbf{q}_2+\dots +\theta _n\textbf{q}_n=\textbf{r}\). Moreover, \(\textbf{x}(\varvec{\theta })\) is the unique state for which \(\theta _i=x_i \varrho (\textbf{x})/\bar{\varrho }(\textbf{x})\) \((i=1,2,\dots ,n)\) holds.
We consider two particular cases. One of them is when \(n=N=3\), \(\textbf{q}_1=\textbf{e}_1\), \(\textbf{q}_2=\textbf{e}_2\) and \(\textbf{q}_3=\textbf{e}_3\). Let \(\textbf{r}=(r_1,r_2,1-r_1-r_2)=r_1\textbf{e}_1+r_2\textbf{e}_2 +(1-r_1-r_2)\textbf{e}_3\) be a strategy from \(S_3\). By the previous lemma, there exists a uniqueFootnote 3 state \(\textbf{x}(\textbf{r})\in S_3\) with \(\textbf{h}\big (\textbf{x}(\textbf{r})\big ) = \textbf{r}\), namely, \(x_i(\textbf{r})=r_i\rho (\textbf{r})/\rho _{\textbf{e}_i}(\textbf{r})\). So \(x_i(\textbf{r})\) has continuous partial derivatives with respect to the variables \(r_1\) and \(r_2\). The notation \(\textbf{x}^R(\textbf{r})\) is used (“R” refers to “restricted”) if necessary to emphasize that \(\textbf{x}(\textbf{r})\) is concerned with this case.
The other case is when \(N=3\), \(n=4\), \(\textbf{q}_1=\textbf{e}_1\), \(\textbf{q}_2=\textbf{e}_2\), \(\textbf{q}_3=\textbf{e}_3\) and \(\textbf{q}_4=\textbf{p}^*:=(1/3,1/3,1/3)\). Then most of the strategies from \(S_3\), namely, the strategies in the interior of \(S_3\) can be expressed in the form \(\theta _1\textbf{e}_1+ \theta _2\textbf{e}_2+\theta _3\textbf{e}_3+\theta _4\textbf{p}^*\) in more than one way. For such a strategy \(\textbf{r}\), there is thus more than one state \(\textbf{x}=(x_1,x_2,x_3,x_4)\) with \(\textbf{h}(\textbf{x})=\textbf{r}\). So it would be ambiguous to use the notation \(\textbf{x}(\textbf{r})\) for a state with \(\textbf{h}(\textbf{x}(\textbf{r}))=\textbf{r}\). Therefore we associate the coefficient-list \(\varvec{\theta }=(\theta _1,\theta _2,\theta _3,\theta _4)\) from the convex combination \(\theta _1\textbf{e}_1+ \theta _2\textbf{e}_2+\theta _3\textbf{e}_3+\theta _4\textbf{p}^*=\textbf{r}\) to the state \(\textbf{x}=\textbf{x}(\varvec{\theta })=\textbf{x}(\theta _1,\theta _2,\theta _3,\theta _4)\) with \(x_i(\varvec{\theta })=\theta _i\rho (\textbf{r})/\rho _{\textbf{r}_i}(\textbf{r})\) rather than associating a strategy \(\textbf{r}\) to such a state. If it is necessary to emphasize that \(\textbf{x}(\varvec{\theta })\) is concerned with this case, then we use the notation \(\textbf{x}^E(\varvec{\theta })\) (“E” refers to “extended”). Observe that if the vector \((x_1,x_2,1-x_1-x_3,0)\in S_4\) is identified with the vector \((x_1,x_2,1-x_1-x_3)\in S_3\), then the map \(\textbf{x}^R\) is just the restriction of the map \(\textbf{x}^E\) to the set \(S_3\).
3 Dynamically Unstable UESS
It is well known in the case of classic evolutionary matrix gamesFootnote 4 that a UESS always implies the asymptotic stability of the corresponding equilibrium point of the associated replicator equation ([21, 36, 44] or Theorem 7.2.4 in Hofbauer and Sigmund [20]).
An analogues implication for matrix games under time constraints up to now was just proved when the the strategy space consists of 2 dimensional strategies. For higher dimensional strategies, however, the question remained open in general [17, 42].
Now we give examples exhibiting UESS with unstable corresponding equilibrium, that is, the accustomed implication of the classical model does not hold in the more general model of matrix games under time constraints. This shows that considering time effects can dramatically alter the outcome of evolution.
The strategy space of the examples is
In each example, strategy \(\textbf{p}^*=(1/3,1/3,1/3)\) is a UESS which is verified in Sections A.1 and A.2. The corresponding state is \(\textbf{x}=\textbf{x}(\textbf{p}^*)= \textbf{x}^R(\textbf{p}^*) =\big (x_1(\textbf{p}^*),x_2(\textbf{p}^*),x_3(\textbf{p}^*)\big )\) for which \(\textbf{h}(\textbf{x})=\textbf{p}^*\) (see Lemma 2.2) (recall the notations \(\textbf{x}^R\) and \(\textbf{x}^E\) in the second paragraph after Lemma 2.2). We analyse the replicator Eq. (2.1) for the polymorphic population consisting of pure phenotypes \(\textbf{e}_1,\) \(\textbf{e}_2\), \(\textbf{e}_3\) with frequencies \(x_1,x_2\) and \(x_3=1-x_1-x_2\):
We also investigate the replicator equation with respect to the population of \(\textbf{q}_1=\textbf{e}_1,\textbf{q}_2=\textbf{e}_2,\textbf{q}_3= \textbf{e}_3\) and \(\textbf{q}_4=\textbf{p}^*=(1/3,1/3,1/3)\) individuals. This means the extension of the previous dynamics (3.11) adding a fourth equation:
where \(x_4\) describes the frequency of \(\textbf{p}^*\) individuals. The phase space of this dynamics is \(S_4\). The state (1, 0, 0, 0) corresponds to pure strategy \(\textbf{e}_1\) in the sense that in this state every individual follows strategy \(\textbf{e}_1\). Similarly, state (0, 1, 0, 0) corresponds to pure strategy \(\textbf{e}_2\), state (0, 0, 1, 0) corresponds to pure strategy \(\textbf{e}_3\) and state (0, 0, 0, 1) corresponds to strategy \(\textbf{p}^*\). Clearly, if the replicator Eq. (3.12) is restricted to the face with vertices (1, 0, 0, 0), (0, 1, 0, 0) and (0, 0, 1, 0) we get back the dynamics (3.11). Accordingly, this face contains the state \(\textbf{x}^E(1/3,1/3,1/3,0)\) (\(\textbf{x}_{\textbf{p}^*}\) for short) corresponding to the UESS \(\textbf{p}^*\) in which each individual follows one of the three pure strategies and the average strategy of active individuals is the UESS \(\textbf{p}^*\). Note that this state is the same as the state \(\textbf{x}^R(\textbf{p}^*)\) for the replicator equation with respect to pure strategies \(\textbf{e}_1,\textbf{e}_2,\textbf{e}_3\).
By Lemma 2.2, the segment (the green segment in Figs. 2 and 5) connecting \(\textbf{x}_{\textbf{p}^*}\) with (0, 0, 0, 1) precisely consists of the states corresponding to the UESS \(\textbf{p}^*\) and the coordinates of these states are
and
respectively, where u runs over the interval [0, 1]. These states are equilibrium points of the replicator Eq. (3.12) (Lemma 3.2 in Garay et al. [17]). Furthermore, state (0, 0, 0, 1) at which every individual of the population follows strategy \(\textbf{p}^*\) is a stable (but not asymptotically stable) rest point of the replicator Eq. (3.12) (Theorem 4.9 in Varga et al. [42]). If the dynamics (3.12) is restricted to any of the faces with vertex (0, 0, 0, 1) then state (0, 0, 0, 1) is asymptotically stable on that face (Theorem 4.7 in Varga et al. [42]).
One of the eigenvalues at every point of the segment is always 0, the other two have zero real part at a single point of the segment which is denoted by \(\textbf{x}_0\) (the upper green point in Figs. 2 and 5). Above the point (the part of the segment falling between the states \(\textbf{x}_0\) and (0, 0, 0, 1)) both non-zero eigenvalues have negative real parts while under the point (the part of the segment falling between the states \(\textbf{x}_{\textbf{p}^*}\) and \(\textbf{x}_0\)) the real parts of the two non-zero eigenvalues are positive. To find \(\textbf{x}_0\) we have analyzed the linearisation of the right hand side of the dynamical system (3.12) along the segment between \(\textbf{x}_{\textbf{p}^*}\) and (0, 0, 0, 1).
We remark that the local property of equilibrium points and the global behaviour of the dynamics on the edges of the phase spaces \(S_3\) and \(S_4\), respectively, are mathematically exact, the eigenvalues were calculated, whereas the global behaviour in the interior of the phase spaces and in the interior of the faces of \(S_4\) are just based on numerical simulations of the phase portrait.
3.1 Example 1: Dynamically Unstable UESS in Rock-Paper-Scissors Game
The popular children’s game, the rock-paper-scissors game, describes a game with three pure strategies where each pure strategy beats precisely another pure strategy (rock beats scissor, scissors beat paper and paper beats rock). The payoff matrix has the following form
The corresponding replicator equation always has a rest point that is asymptotically stable (and the corresponding mixed strategy is a UESS) if and only if the determinant \(b_1b_2b_3-a_1a_2a_3\) is positive (Theorem 7.7.2 in Hofbauer and Sigmund [20]). Otherwise, the rest point is a (non-asymptotically) stable center (if the determinant is zero) or unstable (if the determinant is negative). From evolutionary aspect, it models a cyclic dominance [24, 27, 35] which can result in the coexistence of the three phenotypes [20, 31, 32, 41].
Now we show that arbitrary short waiting times can destabilize a stable coexistence. Let the payoff matrix \(A=A(s)\) and the time constraint matrix \(T=T(s)\) be
where
Then for any \(s\in (0,s_0)\) with some \(s_0\ge 3\) strategy \(\textbf{p}^*=(1/3,1/3,1/3)\) is a UESS of the matrix game under time constraints with payoff matrix A(s) and time constraint matrix T(s) such that the corresponding sate \(\textbf{x}(\textbf{p}^*)\) is an unstable rest point of the replicator Eq. (3.11). The verification can be found in Appendix 1. Note that the determinant of A(s) is positive if \(s\in (0,3]\) so the matrix game with matrix A(s) (\(s\in (0,3]\)) has an interior UESS (which tends to \(\textbf{p}^*\) as \(s\rightarrow 0\)) (see expression (7.34) in Hofbauer and Sigmund [20]) and, consequently, this UESS is an asymptotically stable rest point of the relating replicator equation with respect to pure strategies ([21, 36, 44] or Theorem 7.2.4 in Hofbauer and Sigmund [20]).
The phase portrait of dynamics (3.11) for \(s=1\) is depicted in Fig. 1. The three vertices together with the edges between them form a heteroclinic cycle. The Jacobian matrix of the right hand side at equilibrium point
corresponding to the UESS \(\textbf{p}^*=(1/3,1/3,1/3)\) has eigenvalues with positive real part implying the instability of the rest point. The phase portrait suggests that the solutions tend to the boundary of the simplex.

The phase portrait of the replicator Eq. (3.11) with respect to the pure phenotypes \(\textbf{e}_1,\textbf{e}_2\) and \(\textbf{e}_3\). The payoff matrix A and the time constraint matrix T are given in (3.13) setting s to be 1. \(\textbf{x}(\textbf{p}^*)=\textbf{x}^R(\textbf{p}^*)\) is the state corresponding to strategy \(\textbf{p}^*=(1/3,1/3,1/3)\) through Lemma 2.2: \(x_1(\textbf{p}^*) =x_2(\textbf{p}^*)=(\sqrt{21}-3)/6\approx 0.263\). Although \(\textbf{p}^*\) is a UESS, \(\textbf{x}(\textbf{p}^*)\) is an unstable rest point of the replicator equation. The vertices of the simplex are saddle points and the boundary of the simplex is a heteroclinic cycle which appears to be attractive. For every state \(\textbf{x}\), there is a composition \(Q^{\textbf{p}^*}\hspace{-1mm}(\textbf{x})\) exhibiting strategy \(\textbf{p}^*\) in the population in state \(\textbf{x}\) (see Sect. 3.4 for more explanation). Therefore, a subpopulation in state \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) has higher fitness than that of the whole population. The green segment is the set of states \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\)-s as \(\textbf{x}(t)\) runs over the red orbit (see the enlargement of the green segment in Fig. 6). \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) generally differs from \(\textbf{x}(\textbf{p}^*)\). One can intuitively expect that the population “try” to evolve towards \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) rather than \(\textbf{x}(\textbf{p}^*)\) but \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) varies from moment to moment which can contribute to the instability of \(\textbf{x}(\textbf{p}^*)\) in the example. In the table, we give the real parts of the eigenvalues of the linearisation of the replicator equation at the rest points indicated in the phase portrait
The phase portrait of the extended replicator Eq. (3.12) is shown in Fig. 2. On the face determined by vertices (1, 0, 0, 0), (0, 1, 0, 0), (0, 0, 1, 0), the dynamics agrees with (3.11) so the phase portrait on it is the same as in Fig. 1.
The solutions in the interior of the face determined by vertices (1, 0, 0, 0), (0, 1, 0, 0), (0, 0, 0, 1) start from (1, 0, 0, 0) and end in (0, 0, 0, 1). The situation is similar on the other two faces.
The state \(\textbf{x}_0\) on the segment matching \(\textbf{x}_{\textbf{p} ^*}\) with (0, 0, 0, 1) and corresponding to the UESS \(\textbf{p}^*\) is equal to \(\textbf{x}^E(u_0/3,u_0/3,u_0/3,1-u_0)\) where \(u_0= (71 + 9 \sqrt{21})/334\approx 0.336\). Accordingly, the coordinates of \(\textbf{x}_0\) are \((x_0)_1\approx 0.089\), \((x_0)_2\approx 0.089\), \((x_0)_3\approx 0.159\) and \((x_0)_4=1-u_0\approx 0.664\).

The phase portrait of the replicator equation with respect to phenotypes \(\textbf{e}_1,\textbf{e}_2,\textbf{e}_3\) and \(\textbf{p}^*=(1/3,1/3,1/3)\). The payoff matrix A and the time constraint matrix T are given in (3.13) setting s to 1. \(\textbf{x}_{\textbf{p}^*}=(x_{\textbf{p}^*\hspace{-1mm},1},x_{\textbf{p}^*\hspace{-1mm},2}, x_{\textbf{p}^*\hspace{-1mm},3},x_{\textbf{p}^*\hspace{-1mm},4})\) is the state corresponding to strategy \(\textbf{p}^*=(1/3,1/3,1/3)\) through Lemma 2.2 on the face determined by the vertices (1, 0, 0, 0), (0, 1, 0, 0) and (0, 0, 1, 0): \(x_{\textbf{p}^*\hspace{-1mm},1} =x_{\textbf{p}^*\hspace{-1mm},2}=(\sqrt{21}-3)/6\approx 0.263\), \(x_{\textbf{p}^*\hspace{-1mm},3} =1-x_{\textbf{p}^*\hspace{-1mm},1}-x_{\textbf{p}^*\hspace{-1mm},2}\), \(x_{\textbf{p}^*\hspace{-1mm},4}=0\). It agrees with \(\textbf{x}(\textbf{p}^*)\) in Fig. 1. Every state on the green segment between (0, 0, 0, 1) and \(\textbf{x}_{\textbf{p}^*}\) corresponds to strategy \(\textbf{p}^*\) through Lemma 2.2. Hence, every point of the segment is a rest point of the replicator equation. One of the three eigenvalues of the linearisation of the replicator Eq. (3.12) at these states is zero. At \(\textbf{x}_0\), all of the eigenvalues have zero real part. The states on the segment under \(\textbf{x}_{0}\) have two eigenvalues with positive real part, so they are all unstable though they correspond to the UESS \(\textbf{p}^*\). The states between \(\textbf{x}_0\) and (0, 0, 0, 1) whereas have two eigenvalues with negative real part. The state (0, 0, 0, 1) is stable (but not asymptotically). In the table, we give the real parts of the eigenvalues of the linearisation of the replicator equation at the rest points indicated in the phase portrait
3.2 Limit Cycle, Hopf Bifurcation
Previously, we have found that both the game with payoff matrix A(1) and the game under time constraints with the same payoff matrix and time constraint matrix T(1) have a UESS. Nevertheless, the stability properties of the corresponding rest points are different. While the rest point is asymptotically stable in the former case, it is unstable in the latter case, and the orbits spirally tend to the boundary of the simplex according to the numerical simulation (Fig. 1). The situation can remind us of a supercritical Hopf bifurcation. Indeed, let
where
Here, \({\tilde{a}}_{31}(\sigma )\) has been chosen such that \(\textbf{p}^*=(1/3,1/3,1/3)\) is a UESS for every \(\sigma \in [0,1]\). Note that \({\tilde{A}}(1)=A(1)\). Therefore, the matrix game under time constraints with matrices \({\tilde{A}}(1)\) and \({\tilde{T}}(1)\) has a UESS such that the corresponding rest point of the replicator equation is unstable while the game with matrices \({\tilde{A}}(0)\) and \(\tilde{T}(0)\) has a UESS such that the corresponding rest point is already asymptotically stable. Further analysis reveals that a supercritical Hopf bifurcation occurs at \(\sigma =(1+\sqrt{82})/54\approx 0.186\) (see Appendix A.3). So a limit cycle around the rest point must exist when \(\sigma \) is close to \((1+\sqrt{82})/54\). For instance, for \(\sigma =19/100\), a limit cycle takes shape according to the numerical simulation. However, the orbits tending to the limit cycle wind around themselves so tightly that the limit cycle cannot be perceived easily. Therefore, we give a more spectacular example in Fig. 3.

The phase portrait of the replicator Eq. (3.11) with respect to the pure phenotypes \(\textbf{e}_1,\textbf{e}_2\) and \(\textbf{e}_3\). The payoff matrix A and the time constraint matrix T are given right from the phase portrait. \(\textbf{x}(\textbf{p}^*)=\textbf{x}^R(\textbf{p}^*)\) is the state corresponding to strategy \(\textbf{p}^*=(1/3,1/3,1/3)\) through Lemma 2.2: \(x_1(\textbf{p}^*)=x_2(\textbf{p}^*) = 10/(15+\sqrt{1245})\approx 0.1989\). Although \(\textbf{p}^*\) is a UESS (see Appendix A.2.3 ), \(\textbf{x}(\textbf{p}^*)\) is an unstable rest point of the replicator equation. The vertices of the simplex are saddle points and the boundary of the simplex is a repelling heteroclinic cycle. It seems that there is a stable limit cycle around \(\textbf{x}(\textbf{p}^*)\). In the table, we give the real parts of the eigenvalues of the linearisation of the replicator equation at the rest points indicated in the phase portrait
3.3 Example 2 with a Strict NE and a dynamically Unstable UESS
The payoff matrix A and the time constraint matrix T are the following in this case:
Although a similar phase portrait is possible in the classical case (the last element of the first row in Figure 11 in Zeeman [44], phase portrait 12 in Fig. 6 in Bomze [3]), the unstable interior rest point corresponds to a UESS. In addition to \(\textbf{p}^*=(1/3,1/3,1/3)\), there is a further UESS, moreover, a strict Nash equilibrium which is impossible in classic matrix games (see the first paragraph after the proof of Theorem 6.4.1 on p.64 in Hofbauer and Sigmund [20]). A strategy \(\textbf{r}\) is a strict Nash equilibrium if, for every strategy \(\textbf{q}\not =\textbf{r}\), the strict inequality
holds (Definition 2.2 in Garay et al. [17]). If \(\textbf{r}\) is a strict Nash equilibrium, then it is a pure strategy (Theorem 4.1 in Garay et al. [17]) and a UESS (Theorem 4.1 in Varga et al. [42]). In the present example, strategy \(\textbf{e}_2\) is a strict Nash equilibrium implying that the corresponding state (0, 1, 0) and (0, 1, 0, 0), respectively, is an asymptotically stable rest point of the replicator equation with respect to \(\textbf{e}_1,\textbf{e}_2, \textbf{e}_3\) and the replicator equation with respect to \(\textbf{e}_1,\textbf{e}_2, \textbf{e}_3,\textbf{p}^*\), respectively (Corollary 4.8 in Varga et al. [42]).
To see that \(\textbf{e}_2\) is a strict Nash equilibrium, one should check inequality (3.16) with \(\textbf{r}=\textbf{e}_2\). By (2.6) and (2.7) we get \(\rho (\textbf{e}_2)\) and \(\rho _{\textbf{q}}(\textbf{e}_2)\), respectively. We obtain that
Note that \(\textbf{q}=(q_1,q_2,1-q_1-q_2)\in S_3\) is a strategy. Thus the denominator \(2 + 40 q_1 + 2 q_2\) is positive for every \(\textbf{q}\). Consequently, in order to prove that \(\textbf{e}_2\) is a strict Nash equilibrium, it is enough to validate that the numerator \(3 - 796 q_1 + 38 \sqrt{469} \,q_1 - 3 q_2>0\) for every \(\textbf{q}\not =\textbf{e}_2\) which can easily be seen:
which is positive for every strategy \(\textbf{q}\) different from \(\textbf{e}_2\). Hence \(\textbf{e}_2\) is a strict Nash equilibrium, as claimed.

The phase portrait of the replicator Eq. (3.11) with respect to the pure phenotypes \(\textbf{e}_1,\textbf{e}_2\) and \(\textbf{e}_3\). The payoff matrix A and the time constraint matrix T are the matrices in (3.15). \(\textbf{x}(\textbf{p}^*)=\textbf{x}^R(\textbf{p}^*)\) is the state corresponding to strategy \(\textbf{p}^*=(1/3,1/3,1/3)\) through Lemma 2.2: \(x_1(\textbf{p}^*) = (495 - 4 \sqrt{469})/529\approx 0.772\), \(x_2(\textbf{p}^*)=(409 + 17\sqrt{469})/5290\approx 0.147\). Although \(\textbf{p}^*\) is a UESS, \(\textbf{x}(\textbf{p}^*)\) is an unstable rest point of the replicator equation. Among the vertices, only (1, 0, 0) is a saddle point; (0, 1, 0) is asymptotically stable, moreover, it corresponds to a strict Nash equilibrium while (0, 0, 1) is a source. In addition to the vertices, \(\textbf{x}^{(12)}\) and \(\textbf{x}^ {(13)}\) are two further rest points on the boundary of the simplex. Both of them are saddle points. For every state \(\textbf{x}\), there is a composition \(Q^{\textbf{p}^*}\hspace{-1mm}(\textbf{x})\) exhibiting strategy \(\textbf{p}^*\) in the population in state \(\textbf{x}\) (see Sect. 3.4 for more explanation). Therefore, a subpopulation in state \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) has higher fitness than that of the whole population. The green curve is the set of states \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\)-s as \(\textbf{x}(t)\) runs over the blue orbit (see the enlargement of the green segment in Fig. 6). \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) generally differs from \(\textbf{x}(\textbf{p}^*)\). One can intuitively expect that the population “try” to evolve towards \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) rather than \(\textbf{x}(\textbf{p}^*)\) but \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) varies from moment to moment which can contribute to the instability of \(\textbf{x}(\textbf{p}^*)\) in the example. In the table, we give the real parts of the eigenvalues of the linearisation of the replicator equation at the rest points indicated in the phase portrait
Considering the replicator equation with respect to \(\textbf{e}_1\), \(\textbf{e}_2\) and \(\textbf{e}_3\), we find two additional rest points besides the vertices and the rest point \(\textbf{x}^R(\textbf{p}^*)\) corresponding to the UESS \(\textbf{p}^*\). There is a rest point on the edge between states (1, 0, 0) and (0, 1, 0). We denote it by \(\textbf{x}^{(12)}\approx (0.689,\,0.311,\,0)\). This is an unstable rest point of the dynamics restricted to the edge, and it is a saddle point of the dynamics with respect to the pure strategies.
The other rest point can be found on the edge between (1, 0, 0) and (0, 0, 1). It is denoted by \(\textbf{x}^{(13)}\approx (0.395,\,0,\,0.605)\). This state is asymptotically stable on the edge, but it is a saddle point of the replicator dynamics with respect to \(\textbf{e}_1\), \(\textbf{e}_2\) and \(\textbf{e}_3\). A separatrix starts from it and ends in the state (0, 1, 0).
The (interior) solutions on the (1, 0, 0) side of the separatrix start from state \(\textbf{x}(\textbf{p}^*)=\textbf{x}^R(\textbf{p}^*)\) and end in (0, 1, 0) except for one that ends in \(\textbf{x}^{(12)}\). The (interior) solutions on the (0, 0, 1) side of the separatrix start from (0, 0, 1) and end in (0, 1, 0) (see Fig. 4).
The coordinates of \(\textbf{x}(\textbf{p}^*)\) are
The phase portrait of the extended replicator Eq. (3.12) is illustrated in Fig. 5. The restriction of the dynamics onto the face determined by vertices (1, 0, 0, 0), (0, 1, 0, 0) and (0, 0, 1, 0) is just the replicator equation with respect to pure strategies, so the phase portrait on that face agrees with that in Fig. 4.

The phase portrait of the replicator equation with respect to phenotypes \(\textbf{e}_1,\textbf{e}_2,\textbf{e}_3\) and \(\textbf{p}^*=(1/3,1/3,1/3)\). The payoff matrix A and the time constraint matrix T are given in (3.15). \(\textbf{x}_{\textbf{p}^*}=(x_{\textbf{p}^*\hspace{-1mm},1},x_{\textbf{p}^*\hspace{-1mm},2}, x_{\textbf{p}^*\hspace{-1mm},3},x_{\textbf{p}^*\hspace{-1mm},4})\) is the state corresponding to strategy \(\textbf{p}^*=(1/3,1/3,1/3)\) through Lemma 2.2 on the face determined by the vertices (1, 0, 0, 0), (0, 1, 0, 0) and (0, 0, 1, 0): \(x_{\textbf{p}^*\hspace{-1mm},1}\approx 0.772\), \(x_{\textbf{p}^*\hspace{-1mm},2}\approx 0.147\), \(x_{\textbf{p}^*\hspace{-1mm},3} =1-x_{\textbf{p}^*\hspace{-1mm},1}-x_{\textbf{p}^*\hspace{-1mm},2}\), \(x_{\textbf{p}^*\hspace{-1mm},4} =0\). It agrees with \(\textbf{x}(\textbf{p}^*)\) in Fig. 4. Every state on the green segment between (0, 0, 0, 1) and \(\textbf{x}_{\textbf{p}^*}\) corresponds to strategy \(\textbf{p}^*\) through Lemma 2.2. Hence, every point of the segment is a rest point of the replicator equation. One of the three eigenvalues of the linearisation of the replicator Eq. (3.12) at these states is zero. At \(\textbf{x}_0\), all of the eigenvalues has zero real part. The states on the segment under \(\textbf{x}_{0}\) have two eigenvalues with positive real part, so they are all unstable though they correspond to the UESS \(\textbf{p}^*\). The states between \(\textbf{x}_0\) and (0, 0, 0, 1) whereas have two eigenvalues with negative real part. The state (0, 0, 0, 1) is stable (but not asymptotically). In the table, we give the real parts of the eigenvalues of the linearisation of the replicator equation at the rest points indicated in the phase portrait
There exists a rest point on the edge between (0, 1, 0, 0) and (0, 0, 0, 1). It is denoted by \(\textbf{x}^{(24)}\). Its coordinates are \(x^{(24)}_1=x^{(24)}_3=0\), \(x^{(24)}_2\approx 0.2186\), and \(x^{(24)}_4\approx 0.7814\). The rest point is unstable with respect to the edge.
It appears that there is a separatrix connecting the rest point \(\textbf{x}^{(24)}\) with the rest point \(\textbf{x}^{(12)}\) on the face determined by the vertices (1, 0, 0, 0), (0, 1, 0, 0) and (0, 0, 0, 1). The separatrix divides the face into two parts. The (interior) orbits start from \(\textbf{x}^{(24)}\) and end in the stable (moreover asymptotically stable with respect to the face) rest point (0, 0, 0, 1) on the part falling toward the edge between (1, 0, 0, 0) and (0, 0, 0, 1) while in the asymptotically stable rest point (0, 1, 0, 0) on the other part of the face.
Also, it seems that a separatrix runs from \(\textbf{x}^{(13)}\) to (0, 0, 0, 1) on the face determined by states (1, 0, 0, 0), (0, 0, 1, 0) and (0, 0, 0, 1). The orbits in the interior of the face all end in state (0, 0, 0, 1) but the orbits on the (1, 0, 0, 0) side of the separatrix start from state (1, 0, 0, 0) whereas those falling on the (0, 0, 1, 0) side of the separatrix start from state (0, 0, 1, 0).
It seems that there is also a separatrix on the face determined vertices (0, 1, 0, 0), (0, 0, 1, 0) and (0, 0, 0, 1) connecting state (0, 0, 1, 0) to \(\textbf{x}^{(24)}\). The orbits in the interior of the face start from (0, 0, 1, 0). The orbits located on the (0, 1, 0, 0) side of the separatrix run into state (0, 1, 0, 0) while those on the (0, 0, 0, 1) side of the separatrix end in state (0, 0, 0, 1).
In this case, state \(\textbf{x}_0\) is the state \(\textbf{x}^E(u_0/3,u_0/3,u_0/3,1-u_0)\) where
so its coordinates are \((x_0)_1\approx 0.7596\), \((x_0)_2\approx 0.1446\), \((x_0)_3\approx 0.07981\) and \((x_0)_4=1-u_0\approx 0.01607\).
3.4 Composition Corresponding to a Strategy \(\textbf{q}\) in a Population of State \(\textbf{x}\)
In the context of classic matrix games, if \(\textbf{p}^*\) is a UESS then state this UESS is an asymptotically stable rest point of the replicator equation with respect to pure strategies. The intuitive explanation says that a subpopulation in state \(\textbf{p}^*\) has a higher mean fitness than the total population of state \(\textbf{x}\) and therefore the population evolves to state \(\textbf{p}^*\). In the morning of classical evolutionary matrix games [29], however, the implication was not clear; some years lasted before the discovery of the relation [21, 36, 44].
Our counterexamples show that this classical relation is not self-evident. To help the intuitive comprehension why the implication does not hold in general in the case of matrix games under time constraints, it can be worth analysing the composition corresponding to the UESS in a population of state \(\textbf{x}\).
Lemma 2.2 gives the state \(\textbf{x}\) that corresponds to a strategy \(\textbf{q}\) in the sense that the mean strategy of active individuals is just \(\textbf{q}\) and the proportion of active individuals equals the proportion of active individuals in the monomorphic population of \(\textbf{q}\) individuals, in short, \(\textbf{h}(\textbf{x})=\textbf{q}\) and \({\bar{\varrho }}(\textbf{x})=\rho (\textbf{q})\). Motivated by this lemma, take a population of pure strategists in state \(\textbf{x}\) and consider the following composition. If \(\textbf{q}=q_1\textbf{e}_1+q_2\textbf{e}_2+q_3\textbf{e}_3\) is a strategy, then let
and
Observe that
furthermore,
that is, a subpopulation in state \(Q^{\textbf{q}}(\textbf{x})\) corresponds to strategy \(\textbf{q}\) in the sense that the proportion of active individuals in the subpopulation is the same as the probability that a \(\textbf{q}\) individual playing against the population is active and that the mean fitness of the subpopulation is the same as the fitness of a \(\textbf{q}\) individual playing against the population.
The observation shows that \(Q^{\textbf{q}}(\textbf{x})\) is not a fixed composition in general. It varies with \(\textbf{x}\). In particular, if \(\textbf{q}=\textbf{p}^*\), that is, \(\textbf{q}\) is a UESS, then the subpopulation corresponding to the UESS varies as the state of the population varies (see green curves in Fig. 1, 4, 6 and 7). This is a distinction from the classical case in which the corresponding composition was just the same as the strategy [\(Q^{\textbf{q}}(\textbf{x})=\textbf{q}\)] independently of \(\textbf{x}\). This means that composition \(Q^{\textbf{p}^*}(\textbf{x})\) generally differs from \(\textbf{x}(\textbf{p}^*)\) [in the classical case \(\textbf{x}(\textbf{p}^*)=Q^{\textbf{p}^*}(\textbf{x})=\textbf{p}^*\)]. It seems that this slight deviation can be enough to make \(\textbf{x}(\textbf{p}^*)\) unstable, as shown by our examples.

The neighbourhood of \(\textbf{x}(\textbf{p}^*)\) in Fig. 1 and 4 are enlarged. The left figure corresponds to Example 1 while the right one to Example 2. The green curves consist of the states corresponding to \(\textbf{p}^*\) as \(\textbf{x}(t)\) runs on the orbit around \(\textbf{x}(\textbf{p}^*)\) in Fig. 1, 4 (red orbit in Fig. 1, blue orbit in Fig. 2), in other words, states \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) as defined by (3.17) at \(\textbf{q}=\textbf{p}^*\). If the composition of a subpopulation is \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) then the mean strategy of active individuals of that subpopulation is just \(\textbf{p}^*\). In classic matrix games \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\), \(\textbf{x}(\textbf{p}^*)\) and \(\textbf{p}^*\) coincide, but not in the examples. This can contribute to the instability of \(\textbf{x}(\textbf{p}^*)\)

The initial subarc of the orbit \(\textbf{x}(t)\) around \(\textbf{x}(\textbf{p}^*)\) in Fig. 1, 4 (red orbit in Fig. 1, blue orbit in Fig. 2) and the corresponding curve drawn by composition \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) (green curves). If the composition of a subpopulation is \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) in the population of state \(\textbf{x}(t)\) then the mean strategy of active individuals in the subpopulation is \(\textbf{p}^*\). Therefore, one can intuitively expect that the population evolves towards \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) varying from moment to moment rather than \(\textbf{x}(\textbf{p}^*)\) which leads to the instability of \(\textbf{x}(\textbf{p}^*)\) in the examples. Each segment connects the composition \(Q^{\textbf{p}^*}\hspace{-1mm}\big (\textbf{x}(t)\big )\) with the corresponding state \(\textbf{x}(t)\)
4 Discussion
Garay et al. [16] and Kr̆ivan and Cressman [25] incorporated time constraints into the model of evolutionary matrix games. As they pointed out, time constraints can essentially change evolutionary outcomes. In this article, we have continued the mathematical analyses started in Garay et al. [17], Varga et al. [42] on matrix games under time constraints, and we have demonstrated by two examples that static evolutionary stability does not imply the asymptotic stability of the corresponding state of the replicator equation in three or higher dimensions. This is an essential distinction from the case of classical matrix games, where static evolutionary stability implies dynamic stability [21, 36, 44].
Under the influence of time constraints, individuals in the population move between two states (active and inactive). Since there is no interaction in the inactive state, the duration spent in it can considerably influence how much a particular individual gains from the resources allocated through interactions. Furthermore, the time spent in the inactive state depends on the individual’s strategy. All of this can lead to that individuals of distinct strategies participate in significantly different numbers of interactions over a given period. Thus, in interpreting fitness, it is not enough to simply consider the expected payoff from an interaction, but one must also take into account the process of how interactions unfold. In this regard, matrix games under time constraints share similarities with several recent works in which various mechanisms govern the relevance of game payoffs for evolutionary success. For example, in the model of Foley et al. [12], current payoffs depend on past payoffs, leading to cyclic phenomena in the hierarchy network of the population. Constable et al. [7] introduces stochastic effects into his evolutionary model. Argasinski and Broom [1] and Argasinski and Rudnicki [2] link classical game theory with models where individuals can be in different states (e.g. age-structured population models), so success depends not only on the strategy, but also on the state. In these models, the payoff derived from the games is only a part of the process that, similarly to the time-delay in the time-constrained model, determines the evolutionary dynamics describing the population.
We provided two examples of matrix games under time constraints, wherein the equilibrium point corresponding to an evolutionarily stable strategy proved to be dynamically unstable with respect to the replicator dynamics. The first example pertains to the rock-paper-scissors game, which holds biological relevance in ecology [35], microbiology [24] or biotechnology [27]. Also, it is used for modelling interactions between cancer cells [39]. In addition to proving that arbitrary small distinctions between waiting times can destabilize the rest point corresponding to a UESS, we pointed out that a supercritical Hopf bifurcation can occur, resulting in a stable limit cycle emerging around the unstable rest point. Such a phenomenon is impossible in classic matrix games ([3, 44], Chapter 7.5 in Hofbauer and Sigmund [20]). Mobilia [31] and Toupo and Strogatz [40] obtained a similar result, although their focus was on investigating the effects of mutations rather than time constraints.
The second example exhibited a strict Nash equilibrium in addition to the dynamically unstable interior UESS. This is not possible in the classical case either since the existence of an interior ESS precludes the existence of another Nash equilibrium (see the paragraph after the proof of Theorem 6.4.1 in Hofbauer and Sigmund [20]).
In classical matrix games, if \(\textbf{p}^*\) is an ESS then state \(\textbf{x}^*\) with \(x^*_i=p_i^*\) is an evolutionarily stable state (Theorem 6.4.1 in Hofbauer and Sigmund [20]) in the population of pure phenotypes, that is, for some \(\delta >0\)
where \(W_i\) denotes the fitness of the i-th pure strategy. This condition means that state \(\textbf{x}^*\) exhibits higher fitness in a slightly perturbed state compared to the average fitness of that perturbed state. Therefore, one can expect that the population is steered back to state \(\textbf{x}^*\). Indeed, the above relation implies the asymptotic stability of \(\textbf{x}^*\) with respect to the replicator equation ([21, 44], Theorem 7.2.4 in Hofbauer and Sigmund [20]). Motivated by this, we investigated the composition corresponding to the UESS in a given population of pure phenotypes. Unlike the classical case, this composition depends on the current state of the population as depicted in Figs. 6 and 7. In general, it does not coincide with the state corresponding to the UESS. This discrepancy can potentially contribute to the instability of the state associated with the UESS, as demonstrated in our examples. On the other hand, this provides only a partial explanation since it is easy to give examples such that the composition corresponding to the UESS varies with the current state of the population, yet the state corresponding to the UESS is asymptotically stable. Therefore, future investigations are necessary for a deeper understanding of the stability property of the state corresponding to the UESS.
Finally, analysing the relationship between models on monomorphic and polymorphic populations can primarily appear to be a mathematical problem. However, it is related to the question of diversity in biology. In the theory of classic evolutionary matrix games, for instance, the existence of an interior monomorphic UESS implies stable diversity in polymorphic situations through strong stability [9, 10]. Therefore, making clear the difference between the monomorphic and polymorphic models is an interesting problem from the viewpoint of biology, as well.
Data Availibility
All data generated or analysed during this study are included in this published article.
Notes
If a strategy \(\textbf{q}\in S_N\) is multiplied by an \(N\times N\) matrix M from the right then \(\textbf{q}\) should be considered a row vector while if it is multiplied by M from the left then \(\textbf{q}\) should be considered a column vector.
Garay et al. [16] start from a population consisting of M individuals, where n different phenotypes are present with numbers \(M_1, M_2, \dots , M_n\), respectively. The state of the population is described by an M-dimensional vector \(s = (s_{11} \dots , s_{1M_1} | s_{21}, \dots , s_{2M_2} | \dots | s_{n1}, \dots , s_{nM_n})\), where \(s_{kl}\) represents the state of the l-th individual of type \(\textbf{q}_k\). \(s_{kl}=0\) if the individual is active, while \(s_{kl}=(i,j)\) if he is awaiting after an interaction in which he employed the pure strategy i against the pure strategy j. Since the 0 state corresponds to searching, \(s_{kl}\) takes it for a unit duration, whereas a state value of (i, j) is expected to persist for \(\tau _{ij}\) time units. After \(\tau _{ij}\) time units, \(s_{kl}\) reverts back to 0. If \(Y_k\) denotes the number of active individuals of type \(\textbf{q}_k\) at a given moment, then the intake of an individual of type \(\textbf{q}_k\) during a short time interval \(\Delta t\) is given by \(\left( \frac{M_kY_k-1}{M-1} \textbf{q}_k A \textbf{q}_k + \sum _{z \ne k} \frac{M_zY_z}{M-1} \textbf{q}_k A\textbf{q}_z\right) \Delta t\), since in a random encounter, the opponent is an active individual of type \(\textbf{q}_k\) or \(\textbf{q}_z\) (\(z \ne k\)) with probabilities \(\frac{M_kY_k-1}{M-1}\) and \(\frac{M_zY_z}{M-1}\), respectively. It can be shown that as the observation time and M successively tend to infinity, the time average of the intake tends to the right hand side of (2.3). It can also be verified that \(\lim _{M\rightarrow \infty }{\mathbb {E}}[Y_k(Y_k-1/M_k)]=\varrho _k^2\), \(\lim _{M\rightarrow \infty }{\mathbb {E}}[Y_kY_z]=\varrho _k\varrho _z\), and that the resulting \(\varrho _1,\dots ,\varrho _n\) proportions satisfy Eq. (2.2) (see the proof of Theorem 1 in Garay et al. [16]).
The uniqueness trivially follows from the fact that every \(\textbf{r}=(r_1,r_2,r_3)\in S_3\) is a unique convex combination of the pure strategies: \(\textbf{r}=r_1\textbf{e}_1+ r_2\textbf{e}_2+r_3\textbf{e}_3\).
It corresponds to the case when each entrance of T is the same, say, 0.
A strategy \(\textbf{p}^*\) is called Nash equilibrium for the matrix game under time constraints (NE in short) if the inequality \(\omega _{\textbf{p}^*}(\textbf{p}^*)\ge \omega _{\textbf{q}}(\textbf{p}^*)\) holds for any strategy \(\textbf{q}\in S_N\) (Definition 2.2 in Garay et al. [17]).
This is because every point of the segment is a rest point of the dynamics (3.12).
References
Argasinski K, Broom M (2021) Towards a replicator dynamics model of age structured populations. J Math Biol 22:44. https://doi.org/10.1007/s00285-021-01592-4
Argasinski K, Rudnicki R (2021) Replicator dynamics for the game theoretic selection models based on state. J Theor Biol 526:110540. https://doi.org/10.1016/j.jtbi.2020.110540
Bomze I (1983) Lotka-volterra equation and replicator dynamics: a two-dimensional classification. Biol Cybern 48:201–211. https://doi.org/10.1007/BF00318088
Bomze IM, Weibull J (1995) Does neutral stability imply Lyapunov stability? Games Econ Behav 11:173–192. https://doi.org/10.1006/game.1995.1048
Broom M, Luther RM, Ruxton GD, Rychtář J (2008) A game-theoretic model of kleptoparasitic behavior in polymorphic populations. J Theor Biol 255:81–91. https://doi.org/10.1016/j.jtbi.2008.08.001
Broom M, Rychtář J (2013) Game-theoretical models in biology (1st edition). Chapman & Hall/CRC, New York, Mathematical and computational biology. https://doi.org/10.1201/b14069
Constable GWA, Rogers T, McKane AJ, Tarnita CE (2016) Demographic noise can reverse the direction of deterministic selection. Proc Natl Acad Sci 113:E4745–E4754. https://doi.org/10.1073/pnas.1603693113
Charnov EL (1976) Optimal foraging: attack strategy of a mantid. Am Nat 110:141–151. https://doi.org/10.1086/283054
Cressman R (1990) Strong stability and density-dependent evolutionarily stable strategies. J Theor Biol 145(3):319–330. https://doi.org/10.1016/S0022-5193(05)80112-2
Cressman R (1992) The stability concept of evolutionary game theory. Springer, Berlin. https://doi.org/10.1007/978-3-642-49981-4
Cressman R, Garay J, Varga Z (2003) Evolutionarily stable sets in the single-locus frequency-dependent model of natural selection. J Math Biol 47(5):465–482. https://doi.org/10.1007/s00285-003-0217-7
Foley M, Smead R, Forber P, Riedl C (2021) Avoiding the bullies: the resilience of cooperation among unequals. PLoS Comput Biol 17(4):e1008847. https://doi.org/10.1371/journal.pcbi.1008847
Fudenberg D, Levine DK (1998) The Theory of Learning in Games. MIT Press, Cambridge
Garay J, Varga Z, Cabello T, Gámez M (2012) Optimal nutrient for aging strategy of an omnivore: liebig’s law determining numerical response. J Theor Biol 310:31–42. https://doi.org/10.1016/j.jtbi.2012.06.021
Garay J, Varga Z, Gámez M, Cabello T (2015) Functional response and population dynamics for fighting predator, based on activity distribution. J Theor Biol 368:74–82. https://doi.org/10.1016/j.jtbi.2014.12.012
Garay J, Csiszár V, Móri TF (2017) Evolutionary stability for matrix games under time constraints. J Theor Biol 415:1–12. https://doi.org/10.1016/j.jtbi.2016.11.029
Garay J, Cressman R, Móri TF, Varga T (2018) The ESS and replicator equation in matrix games under time constraints. J Math Biol 76:1951–1973. https://doi.org/10.1007/s00285-018-1207-0
Guckenheimer J, Holmes PJ (1983) Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields. Applied Mathematical Sciences New York: Springer. https://doi.org/10.1007/978-1-4612-1140-2
Gunawardena J (2014) Time-scale separation - Michaelis and Menten’s old idea, still bearing fruit. FEBS J 281(2):473–488. https://doi.org/10.1111/febs.12532
Hofbauer J, Sigmund K (1998) Evolutionary Games and Population Dynamics. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9781139173179
Hofbauer J, Schuster P, Sigmund K (1979) A note on evolutionary stable strategies and game dynamics. J Theor Biol 81:609–612. https://doi.org/10.1016/0022-5193(79)90058-4
Holling CS (1956) The components of predation as revealed by a study of small-mammal predation of the european pine sawfly. Can Entomol 91:293–320. https://doi.org/10.4039/Ent91293-5
John S, Müller J (2023) Age structure, replicator equation, and the prisoner?s dilemma. Math Biosci 365:109076. https://doi.org/10.1016/j.mbs.2023.109076
Kerr B, Riley MA, Feldman MW, Bohannan BJ (2002) Local dispersal promotes biodiversity in a real-life game of rock-paper-scissors. Nature (London) 418:171–174. https://doi.org/10.1038/nature00823
Křivan V, Cressman R (2017) Interaction times change evolutionary outcomes: Two-player matrix games. J Theor Biol 416:199–207. https://doi.org/10.1016/j.jtbi.2017.01.010
Kong Q (2014) A Short Course in Ordinary Differential Equations. Springer, New York. https://doi.org/10.1007/978-3-319-11239-8
Liao MJ, Din MO, Tsimring L, Hasty J (2019) Rock-paper-scissors: engineered population dynamics increase genetic stability. Science 365(6457):1045–1049. https://doi.org/10.1126/science.aaw0542
Maynard Smith J (1982) Evolution and the Theory of Games. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511806292
Maynard Smith J, Price G (1973) The logic of animal conflicts. Nature 246:15–18. https://doi.org/10.1038/246015a0
Mesterton-Gibbons M (2001) An Introduction to Game-Theoretic Modelling (Second edition). Student Mathematical Library, 11. American Mathematical Society. Providence
Mobilia M (2010) Oscillatory dynamics in rock-paper-scissors games with mutations. J Theor Biol 264:1–10. https://doi.org/10.1016/j.jtbi.2010.01.008
Nowak M (2006) Evolutionary Dynamics: Exploring the Equations of Life. The Belknap Press of Harvard University Press, Cambridge. https://doi.org/10.2307/j.ctvjghw98
Pohley HJ, Thomas B (1983) Non-linear ESS-models and frequency dependent selection. Biosystems 16:87–100. https://doi.org/10.1016/0303-2647(83)90030-8
Rudin W (1953) Principles of Mathematical Analysis. McGraw-Hill (New York, Toronto, London)
Sinervo B, Lively CM (1996) The rock-paper-scissors game and the evolution of alternative male strategies. Nature 380:240–243. https://doi.org/10.1038/380240a0
Taylor PD, Jonker LB (1978) Evolutionary stable strategies and game dynamics. Math Biosci 40:145–156. https://doi.org/10.1016/0025-5564(78)90077-9
Thomas B (1985) On evolutionarily stable sets. J Math Biol 22:105–115. https://doi.org/10.1007/BF00276549
Thomas GB (2014) Thomas’ Calculus (13-th edition). Pearson
Tomlinson IPM (1997) Game-theory Models of Interactions Between Tumour Cells. Eur J Cancer 33(9):1495–1500. https://doi.org/10.1016/S0959-8049(97)00170-6
Toupo DFP, Strogatz SH (2015) Nonlinear dynamics of the rock-paper-scissors game with mutations. Phys Rev E 91:052907. https://doi.org/10.1103/PhysRevE.91.052907
Toupo DFP, Strogatz SH, Cohen JD, Rand DG (2015) Evolutionary game dynamics of controlled and automatic decision-making. Chaos 25:073120. https://doi.org/10.1063/1.4927488
Varga T, Móri TF, Garay J (2020) The ESS for evolutionary matrix games under time constraints and its relationship with the asymptotically stable rest point of the replicator equation. J Math Biol 80:743–774. https://doi.org/10.1007/s00285-019-01440-6
Wiggins S (2003) Introduction to applied nonlinear dynamical systems and chaos. Texts in Applied Mathematics New York: Springer. https://doi.org/10.1007/b97481
Zeeman EC (1980) Population dynamics from game theory. In: Global theory of dynamical systems. Lecture Notes in Mathematics 819. New York: Springer-Verlag https://doi.org/10.1007/BFb0087009
Funding
This research was supported by NKFIH, Hungary KKP 129877 (to TV).
Author information
Authors and Affiliations
Contributions
TV and JG designed the study and wrote the main manuscript text, TV constructed and analysed the examples, TV prepared the figures, TV wrote the appendix. Both authors reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no Conflict of interest to declare.
Ethical Approval
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix A
Appendix A
Lemma (2.2) provides the corresponding polymorphic population for the monomorphic population of a given phenotype. The subsequent lemma describes the reverse direction by specifying the phenotype for which the monomorphic population corresponds to a polymorphic population in a given state.
Lemma A.1
( Garay et al. [17], Proposition 3.1) Consider a polymorphic population of phenotypes \(\textbf{q}_1,\textbf{q}_2,\dots ,\textbf{q}_n\) with frequency distribution \((x_1, x_2,\dots ,x_n)\). Then \({\bar{\varrho }}(\textbf{x}) = \rho \big (\textbf{h}(\textbf{x})\big )\) and \(\omega \big (\textbf{h}(\textbf{x})\big ) = {\bar{W}} (\textbf{x})\), that is, the fitness of an \(\textbf{h}(\textbf{x})\) individual is just the average fitness \({\bar{W}}(\textbf{x})\) of the phenotypes \(\textbf{q}_1,\textbf{q}_2,\dots ,\textbf{q}_n\) in the polymorphic model.
Using the previous lemma, Lemma 2.2 and the definitions of the expressions \(\varrho _i\), \({\bar{\varrho }}\), \(\textbf{h}(\textbf{x})\), \(W_i\), \({\bar{W}}\), \(\rho (\textbf{q})\), \(\rho _{\textbf{p}}(\textbf{q})\), \(\omega (\textbf{q})\), \(\omega _{\textbf{p}}(\textbf{q})\) and \(\textbf{x}(\textbf{r})\) (see (2.2), (2.3), (2.4), (2.5), (2.7), (2.8), (2.9) and the second paragraph after Lemma 2.2) we gain the following “exchange rule” between strategies and states.
Corollary A.2
(Exchange rule) Let \(\textbf{r}\) be a strategy in \(S_3\) and consider the polymorphic population of the pure phenotypes \(\textbf{e}_1,\textbf{e}_2,\textbf{e}_3\). Then the following relationships hold:
1.1 A.1 How to Find Dynamically Unstable UESS
One of the main difficulties in the analyses of the replicator Eq. (2.1) is that the right hand side generally cannot be given in explicit form. This is because functions \(\varrho _i\) defined as the solution of the equation system (2.2) cannot be explicitly expressed in general and this problem already arises in three dimensions. We first tried examples in which the explicit calculation was possible, but this way proved ineffective. Therefore, we had to find a more systematic procedure to look for examples with UESS such that the corresponding rest point of the replicator equation is unstable. Here we describe the method used by us. We first quote the following characterization of UESS (Corollary 3.2 in Varga et al. [42]):
A strategy \(\textbf{p}^*\) is a UESS if and only if there is a \(\delta >0\) such that
whenever \(0<||\textbf{p}^*-\textbf{q}||<\delta \).
Accordingly, if
then \(\textbf{p}^*\) is a UESS if and only if it is a strict local minimum of f. Note that since f is defined on \(S_3\) which is a 2 dimensional manifold in \({\mathbb {R}}^3\), it can be considered as a two variable function of \(q_1,q_2\). It is well-known from multivariable calculus that if
-
[C1]
\(f'_{q_1}(p_1^*,p_2^*)=f'_{q_2}(p_1^*,p_2^*)=0,\)
-
[C2]
\(f''_{q_1q_1}(p_1^*,p_2^*)f''_{q_2q_2}(p_1^*,p_2^*)> \big (f''_{q_1q_2}(p_1^*,p_2^*)\big )^2\) and
-
[C3]
\(f''_{q_1q_1}(p_1^*,p_2^*)>0\)
then \(\textbf{p}^*=(p_1^*,p_2^*,1-p_1^*-p_2^*)\) is a strict local minimum of f. (see e.g. Theorem 11 of Chapter 14, p. 838 in Thomas [38]). Consequently, if [C1], [C2], [C3] hold for \(\textbf{p}^*=(1/3,1/3,1/3)\) with respect to the matrices A and T, furthermore, the state \(\textbf{x}(\textbf{p}^*)\) corresponding to \(\textbf{p}^*\) is an unstable rest point of the replicator equation, then we have found a counterexample.
Consider, therefore, the replicator Eq. (3.11) for the polymorphic population consisting of pure phenotypes \(\textbf{q}_1=\textbf{e}_1,\) \(\textbf{q}_2=\textbf{e}_2\), \(\textbf{q}_3=\textbf{e}_3\) with frequencies \(x_1,x_2\) and \(x_3=1-x_1-x_2\), respectively. Since \(x_1+x_2+x_3=1\) also holds, any two equations of the system determine the dynamics. It will thus be sufficient to investigate the differential equation system
If \(\textbf{p}^*\) is a NE,Footnote 5 then the corresponding state \(\textbf{x}(\textbf{p}^*)\) is a rest point of the replicator equation (Lemma 3.2, Garay et al. [17]). Since (1/3, 1/3, 1/3) is a UESS, which implies that it is a NE too (see the paragraph before Definition 2.2 in Garay et al. [17]), it follows that \(\textbf{x}(1/3, 1/3,1/3)\) is a rest point of the replicator equation.
We would like \(\textbf{x}(1/3, 1/3,1/3)\) to be unstable. This holds if the Jacobian matrix of the right hand side of (A.19) has a positive eigenvalue at \(\textbf{x}(1/3, 1/3,1/3)\). It can be easily checked for a function of two variables that if the characteristic polynomials of the Jacobian matrix is \(\lambda ^2 + b\lambda + c\) (where \(\lambda \) is the variable), then the Jacobian matrix has a positive eigenvalue if and only if
-
[C4]
\(b<0\) or \(c<0\)
(see, for example, Chapter 4.3 in Kong [26]).
Conditions [C1]-[C4] provide equations and inequalities for the entrances of the payoff matrix A and the time constraint matrix T. Solving them, one can find appropriate matrices A and T for which there is a UESS \(\textbf{p}^*\) but the corresponding state \(\textbf{x}(\textbf{p}^*)\) is unstable with respect to the replicator equation.
In connection with [C4] we should avoid the explicit calculation of functions \(\varrho _i\) (\(i=1,2,3\)) because, as mentioned, it is generally not possible. Fortunately, we only need the sign of the coefficients of the characteristic polynomial of the Jacobian matrix at the corresponding state \(\textbf{x}(\textbf{p}^*)\) and this can be calculated. Applying Corollary A.2, rewrite (A.19) as follows.
where \(h_1, h_2\) is just the first two components of \(\textbf{h}\), that is, \(\textbf{h}(\textbf{x})=\big (h_1(\textbf{x}), h_2(\textbf{x}),1-h_1(\textbf{x})-h_2(\textbf{x})\big )\). If
then the Jacobian matrix of the right hand side of (A.20) takes the following shape:
We need the Jacobian at state \(\textbf{x}(1/3,1/3,1/3)\). We therefore express \(\textbf{x}\) as a function of \(\textbf{q}\) and then apply the well-known inverse function theorem (see e.g. Theorem 9.22 and (49) in Chapter 9 (p. 181) in Rudin [34]) to the function \(\textbf{h}(\textbf{x})\). We obtain that
where, of course, the determinant in the denominator of the rightmost side should differ from zero. The rightmost expression already can be calculated, and this is enough for our purpose.
As regards dynamics (3.12), one can calculate the linearisation of the right hand side similar to that done in (A.21) for replicator Eq. (3.11). Note that, nevertheless, one should use the maps
respectively, in this case rather than the maps \(\textbf{p}\mapsto \textbf{x}^R(\textbf{p})\) and \(\textbf{x}\mapsto {\textbf{h}}(\textbf{x})\). Accordingly, we rewrite dynamics (3.12) \(\dot{x}_i=x_i[W_i(\textbf{x})-\bar{W}(\textbf{x})]\) (\(i=1,2,3,4\)) as
[Note that \(\textbf{h}(\textbf{x})=\theta _1(\textbf{x}) \textbf{e}_1+\theta _2(\textbf{x}) \textbf{e}_2+ \theta _3(\textbf{x}) \textbf{e}_3+\theta _4(\textbf{x}) \textbf{p}^*\) by (2.5) and (A.22).] Also, the functions \(R_i\) (\(i=1,2,3\)) should be given as the functions of the coefficient-list \(\varvec{\theta }=(\theta _1,\theta _2,\theta _3,\theta _4)\), that is,
where \(\textbf{p}(\varvec{\theta })=\textbf{p}(\theta _1,\theta _2,\theta _3) =\theta _1\textbf{e}_1+\theta _2 \textbf{e}_2+\theta _3\textbf{e}_3+ (1-\theta _1-\theta _2-\theta _3) \textbf{p}^*\) (since \(\theta _1+\theta _2+\theta _3+\theta _4=1\), variable \(\theta _4\) can be dropped). Then, similar to (A.21), we can calculate the Jacobian matrix along the segment \(u\textbf{x}_{\textbf{p}^*}+(1-u)(0,0,0,1)\), \(u\in [0,1]\). After having linearisation along this segment, we compute the related characteristic polynomials \(\lambda ^3+b(u)\lambda ^2+c(u)\lambda +d(u)\). Since one of the eigenvalues is zero at each point of the segment,Footnote 6 it follows that \(d(u)=0\) for any \(u\in [0,1]\). Therefore, the other two eigenvalues should be the zeros of the polynomial \(\lambda ^2+b(u)\lambda +c(u)\). The formulas for b(u) and c(u), respectively, are not too “beautiful” (we, too, have used Wolfram Mathematica 12), but, after analysing them, one can see that \(b(0)=c(0)=0\) and \(c(u)>0\) for \(u\in (0,1]\). Hence, the real part of the zeros can be zero only if \(b(u)=0\). b(u) has the form \((b_{11}-b_{12} u)/(b_{22}u^2-b_{21}u+b_{20})\) or \(u(b_{11}-b_{12} u)/(b_{22}u^2-b_{21}u+b_{20})\) where coefficients \(b_{ij}\) are non-negative, \(b_{12}>b_{11}\) and \(b_{20}>b_{21}\). We infer that the denominator is positive for any \(u\in [0,1]\) so, if \(u\in (0,1]\), the sign of b(u) agrees with the sign of the expression \(b_{11}-b_{12} u\) which is positive if \(0<u<b_{11}/b_{12}\), negative if \(b_{11}/b_{12}<u\le 1\) and 0 if \(u=b_{11}/b_{12}\). It follows (recall condition [C4] too) that the real parts of the two non-zero eigenvalues are positive if \(0<u<b_{11}/b_{12}\), 0 if \(u=b_{11}/b_{12}\) and negative if \(b_{11}/b_{12}<u\le 1\). The state belonging to \(u=b_{11}/b_{12}\) is \(\textbf{x}_0\). The states belonging to u-s between 0 and \(b_{11}/b_{12}\) forms the segment between \(\textbf{x}_{\textbf{p}^*}\) and \(\textbf{x}_0\) while the states belonging to u-s between \(b_{11}/b_{12}\) and 1 form the segment between \(\textbf{x}_{0}\) and (0, 0, 0, 1) (see Figs. 2 and 5).
1.2 A.2 Conditions [C1]-[C4] for the Examples
In the previous section, we identified some conditions that can be used for seeking examples with dynamically unstable UESS. Here, we check the conditions for the examples of the article. Since the expressions that arise in the calculation are rather huge, we used Mathematica 12 to calculate them and we only give rounded values in the most cases.
1.2.1 A.2.1 Conditions [C1]-[C4] for Example 1 with \(s=1\)
First, we consider the special case when \(s=1\). Then one can check that
A straightforward calculation yields condition [C1]. For the second order derivatives we obtain:
from which we get conditions [C2] and [C3]:
Condition [C4] is true with
1.2.2 A.2.2 Conditions [C1]-[C4] for Example 1 with \(s\in (0,3]\)
Note the following simplification. It is clear that \(f(p_1^*,p_2^*)=0\) and
It follows that f has a strict minimum at \(\textbf{p}^*\) if and only if \({\hat{f}}(q_1,q_2):=f(q_1,q_2)/\rho (\textbf{q})\) has a strict minimum at \(\textbf{p}^*\), so it is enough to check [C1]-[C3] for \({\hat{f}}\).
Calculating the first order derivatives, we can see that [C1] is valid. For [C2], we get that
where
with
and
It is easy to see that \(\textrm{den}_1(s)>0\), if \(s\ge 0\). Also, since \(c_5(s)+s\,c_6(s)>s^3\,|c_8(s)|+s^4\,|c_9(s)|\), \(c_4(s)>s^6\,|c_{10}(s)|\), \(c_3(s)>s^8\,|c_{11}(s)|\) and \(c_7(s)>0\) providing that \(s\in (0,3]\), it immediately follows that \(\textrm{num}_1(s)>0\) if \(s\in (0,3]\). Therefore [C2] is valid for \(s\in (0,3]\).
Furthermore,
Since
we infer that \({\hat{f}}''_{q_1,q_1}(1/3,1/3)>0\) for any \(s>0\), so [C3] holds if \(s\in (0,3]\).
To verify [C4], we simplify the calculation again using the fact that \(\rho (\textbf{q})\ge 1/3>0\) and that the orbits of an autonomous differential equation system do not change if each equation is multiplied by the same positive function (Exercise 4.1.3 in Hofbauer and Sigmund [20]). Therefore, we can use differential equation system
instead of differential equation system (A.20). If the characteristic polynomials of (A.23) at \(\textbf{x}(\textbf{p}^*)\) is \(\lambda ^2+{\hat{b}} \lambda +{\hat{c}}\) then [C4] is valid if \({\hat{b}}<0\) or \({\hat{c}}<0\). We obtain that
if \(s\in (0,3]\) and so [C4] is also true.
In summary, [C1]-[C4] hold if \(s\in (0,3]\) which imply that \(\textbf{p}^*\) is a UESS such that the corresponding state \(\textbf{x}(\textbf{p}^*)\) is an unstable rest point of the replicator dynamics with respect to pure strategies for any \(s\in (0,3]\).
1.2.3 A.2.3 Conditions [C1]-[C4] for the Example in Fig. 3
If we calculate the first order derivatives of function f in (A.18) we can see that condition [C1] holds. For the second order derivatives, we get
from which we obtain that conditions [C2] and [C3] also hold:
Condition [C4] holds with
1.2.4 A.2.4 Conditions [C1]-[C4] for Example 2
Calculating the first order derivatives of the function f in (A.18) at \(\textbf{q}=(1/3,1/3)\) we get that both of them are zero so condition [C1] is valid. The second order derivatives are
Hence, we infer that conditions [C2] and [C3] are also satisfied:
Condition [C4] holds with
1.3 A.3 Supercritical Hopf Bifurcation
Here we check that a Hopf bifurcation occurs as \(\sigma \) runs from 0 to 1 in the replicator equation associated to the game with payoff matrix \({\tilde{A}}(\sigma )\) and time constraint matrix \(\tilde{T}(\sigma )\) defined in (3.14).
For this purpose, consider a two dimensional system:
where \(\sigma \in (\sigma _1,\sigma _2)\) is a parameter, \(r_1\) and \(r_2\) are continuously differentiable at least five times. Assume that \((x^*,y^*)\) is an equilibrium point for every \(\sigma \in (\sigma _1,\sigma _2)\) and there is a \(\sigma _0\in (\sigma _1,\sigma _2)\) such that the Jacobian of the right hand side has purely imaginary eigenvalues that is \(r_1(x^*,y^*,\sigma )=r_2(x^*,y^*,\sigma )=0\) and the eigenvalues of
are \(\pm i \beta (\sigma _0)\) with \(\beta (\sigma _0)>0\). Denote by \(B(\sigma )\) the Jacobian
Then the right hand side can be rewritten in the following form:
where \(s_i(x^*,y^*,\sigma )=\partial _xs_i(x^*,y^*,\sigma )=\partial _ys_i(x^*,y^*,\sigma )=0\) (\(i=1,2\)).
If \(\alpha (\sigma )\pm i\beta (\sigma )\) are the eigenvalues of \(B(\sigma )\) and
are the corresponding eigenvectors, then let
Introduce the new variables u, v such that
and let
Using variables u, v, the system (A.24) is transformed into the following form
Consider the so-called Lyapunov coefficient as follows:
where the partial derivatives are taken at \((u,v,\sigma )=(0,0,\sigma _0)\)
The following theorem ensures the occurrence of a supercritical Hopf bifurcation at \(\sigma _0\).
Theorem A.3
(Theorem 3.1.3 in Wiggins [43] or Theorem 3.4.2 in Guckenheimer and Holmes [18]) Using the above notation, assume that the function \(\alpha (\sigma )\) is differentiable, \(\beta (\sigma )\) is continues, \(\alpha (\sigma _0)=0, \alpha '(\sigma _0)\not =0\) and \(\beta (\sigma _0)\not =0\). If \(L<0\) then a supercritical Andronov-Hopf bifurcation occurs at \(\sigma =\sigma _0\), that is,
-
if \(\alpha '(\sigma _0)>0\) then \((x^*,y^*)\) is an asymptotically stable rest point for \(\sigma <\sigma _0\) while unstable for \(\sigma >\sigma _0\), with an asymptotically stable periodic orbit around \((x^*,y^*)\) for \(\sigma >\sigma _0\);
-
if \(\alpha '(\sigma _0)<0\) then \((x^*,y^*)\) is an asymptotically stable rest point for \(\sigma >\sigma _0\) while unstable for \(\sigma <\sigma _0\), with an asymptotically stable periodic orbit around \((x^*,y^*)\) for \(\sigma <\sigma _0\).
We apply the previous theorem for our example. Again, since expressions arising in calculations are huge, we used Mathematica 12. We start from replicator Eq. (3.11) associated with payoff matrix \({\tilde{A}}(\sigma )\) and time constraint matrix \({\tilde{T}}(\sigma )\) defined in (3.14). Since \(x_3=1-x_1-x_2\), the third equation can be neglected. If \(x=x_1\) and \(y=x_2\) then \(r_1(x,y,\sigma )=x[W_1(\textbf{x})-{\bar{W}}(\textbf{x})]\) and \(r_2(x,y,\sigma )=y[W_2(\textbf{x})-{\bar{W}}(\textbf{x})]\). We get that
The characteristic polynomial at \(\sigma _0\) is
It immediately follows that \(\alpha (\sigma _0)=0\) and \(\beta (\sigma _0)\not =0\). In general, the characteristic polynomial is \(\lambda ^2+b(\sigma )\lambda +c(\sigma )\) with
Since
it follows that \(b(\sigma )^2-4c(\sigma )<0\) also holds in a neighbourhood of \(\sigma _0\). For such \(\sigma \)-s we have that \(\alpha (\sigma )=-b(\sigma )/2\) and so we get that
For the Lyapunov coefficient (A.25), we obtain an enormous expression, so here we give a rounded value:
(Note that the precise value is not interesting but the sign which is negative according to Mathematica 12.) In summary, the assumptions of Theorem A.3 are satisfied, implying that supercritical Hopf bifurcation occurs at \(\sigma _0\).
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Varga, T., Garay, J. Dynamically Unstable ESS in Matrix Games Under Time Constraints. Dyn Games Appl (2024). https://doi.org/10.1007/s13235-024-00581-8
Accepted:
Published:
DOI: https://doi.org/10.1007/s13235-024-00581-8