
Abstract
Salinity is one of the major constraints prevailing in environment that affects not only plant growth but also agriculture productivity and soil fertility. Salinity stress causes nutritional and hormonal imbalance, ion toxicity, oxidative and osmotic stress and increased susceptibility of plant to diseases. Increasing use of chemical based fertilizers creates demand for environmental friendly and ecological compatible alternative in agriculture. Plant growth promoting bacteria (PGPB) have tremendous ability to mitigate salinity stress and ameliorate plant growth, playing vital role in food security by increasing agriculture crop productivity. PGPB inoculation under salinity promotes plant growth by several traits including 1-aminocyclopropane 1 carboxylate (ACC) deaminase activity, synthesis of plant hormones (such as indole acetic acid (IAA), gibberellic acid (GA), abscisic acid (ABA), cytokinin) and exopolysaccharides. PGPB alleviates salinity stress in plants by providing nutrients, maintaining high potassium and sodium ratio, increasing accumulation of osmolytes, enhanced photosynthesis and activity of antioxidant enzymes. The present review highlights and discusses current knowledge on effect of salinity stress on plant growth, PGPB mechanisms resulting in mitigation of salinity stress and increasing plant growth. It assesses morphological, biochemical, physiological and molecular changes induced in plants suffering from salinity when supplemented with PGPB.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Stress is any external factor that has negative effects on plant growth and development (Foyer et al. 2016). One of the major abiotic stresses that limit the agricultural yield is salinity stress (Nadeem et al. 2016). The adverse effect of high concentration of minerals such as Na+ and/or Cl− on plant growth is known as salinity stress (Munns 2005). Agriculture field is largest financial source for most of the world’s population and soil salinity is a huge problem in soil under irrigation. The world’s soil in hot and dry region is commonly saline and carries low agriculture potential, in such areas crops grows under irrigation condition, results in secondary soil salinization, affecting worldwide 20% of irrigated land (Glick et al. 2007a). In India salinity affected soil is about 7 billion hectors (Patel et al. 2011), in Maharashtra about 33,200 ha land is suffering from salinity and in case of Kolhapur district the different talukas are adversely affected by salinity (Naik 2014).
Salts are present as electrically charged ions in the soil due to soil weathering and insufficient rainfall (Shrivastava and Kumar 2015). Soil salinity is measured in terms of electric conductivity (EC) which is the capability of the substance to conduct electricity (AE 2001). The saline soil has EC of saturation extract above 4 dS m−1 and 15% of exchangeable sodium. The total yield of many crops is reduced at this EC while many crops shows decrease in yield at the lower EC (Jamil et al. 2011). Around 40% of worlds land has been affected by salinity (Jadhav et al. 2010). The problem of salinity is becoming more severe and by the 2050 an estimated of around 50% of land would be affected by salinity stress (Jamil et al. 2011). In addition every year salinity stressed soil is increasing by 10% because of several primary and secondary reasons. Salinity in soil may be natural that is primary and anthropogenic also known as secondary. The primary causes of salinity are, weathering of rocks, entry of brackish water in soil and ocean salt carried in wind and rains (Manchanda and Garg 2008). The secondary causes include, overuse of chemical fertilizers, industrial effluent entry in agricultural soil, use of low quality ground water for soil irrigation and irrigation without proper drainage system (Parihar et al. 2015). The other factors includes, soil amendments i.e., gypsum that results in increase in salt level of soil (Amacher et al. 2000).
The research on mitigation of salinity stress and soil pollution is prime important to increase crop productivity in order to feed worldwide increasing population. Escalating global hunger and salinity stress creates lucrative interest in sustainable environmental practices (Rashid et al. 2016). Intensive research is going on to ameliorate stress condition, improve plant productivity and protect the plants from phytopathogens using beneficial soil microorganisms known as PGPB (Berg and Martinez 2015; Egamberdieva et al. 2015; Bhise et al. 2017a, 2017b). PGPB promotes the plant growth by nitrogen fixation, phosphate and potassium solubilization as well as synthesis of plant hormones, siderophore, hydrogen cyanide, ammonia and ACC deaminase enzyme (Bhise et al. 2017a; Gontia-Mishra et al. 2017). Soil salinity is scourge for plant growth and crop productivity. Salinity stressed plants suffers from nutritional imbalance, ions toxicity, high osmotic and oxidative stress, reduced photosynthetic rates and decreased productivity. In the view of severity of salinity and PGPB importance in agriculture field the present review discuss the effect of salinity on plant growth status, the key role of PGPB in mitigating salinity stress in plants and promoting its growth and productivity.
2 Effects of salinity stress on plant growth
Salinity stress affects morphological, biochemical and physiological processes of plant growth. Different crops exhibit different response to salinity; mostly all crops shows growth however total yield is reduced (Singh and Chatrath 2001). The overall effect of salinity stress on plant growth is shown in Fig. 1. Excess of salt in the soil leads to accumulation of soluble salt in the root zone which results in osmotic pressure and disturbs the growth of plants by affecting water and ions uptake (Tester and Davenport 2003). The most vital phases of plant growth cycle is seed germination and seedling growth (Kaveh et al. 2011) which determines the total yield of crops. Generally low salt concentration induces dormancy state and reduces germination (Khan and Weber 2006), while high salt increases time required for germination and decreases percentage of germination (Rouhi et al. 2011; Ansari and Zadeh 2012). Salinity stress results in accumulation of high concentration of Na+ and Cl+ which affects other vital elements presence and can decrease plant ability to access as well uptake minerals plus essential nutrients and its distribution in plants (Carmen and Roberto 2011), resulting in nutritional imbalance which reduces physiological activity and plant growth (Moradi and Ismail 2007). The accumulated Na+ in plant tissue increases formation of reactive oxygen species (ROS) such as superoxide (O2−), single oxygen (1O2), hydroxyl radical (OH−), hydrogen peroxide (H2O2) (Ahmad et al. 2011) and inhibit photosynthesis. ROS causes lipid peroxidation, protein oxidation, DNA damage, inactivation of enzymes (Islam et al. 2015) plus chlorophyll degradation (Verma and Mishra 2005). Salinity results in accumulation of sodium and chlorine ions in chlorophyll and chloroplast hence photosynthesis is inhibited as these are important components in photosynthesis (Zhang et al. 2005). In salinity condition the considerably reduced efficiency of photosystem II, CO2 assimilation rate and electron transport chain has been noticed by Stepien and Klobus (2005), Photosystem II is relatively sensitive to salinity stress (Allakhverdiev et al. 2000). Plants exposed to salinity stress produce an increases level of ethylene or stress hormone which actually causes damages to plants (Abeles and Heggestad 1973). Presence of environmental stresses is the major issue that disturbs rhizosphere functioning and influence agriculture productivity (Singh 2015).

Effect of salinity on plant growth
Agriculture soil in arid and semi-arid regions is reduced by 1–2% every year by salinity stress (Kafi and Khan 2008). About 70% reductions in the yield of main crops such as wheat, rice, maize and barley are reported (Acquaah 2007), which is found to be not enough to deal with food demand of worldwide increasing population in 2050 (Ray et al. 2013). The costs related to salinity stress are enormous, estimated at US$12 billion per year and it is increasing (Qadir et al. 2008; Dodd and Pérez-Alfocea 2012). In order to meet food demand of worldwide increasing population, around 50% increase in yield of key crops is crucial (Godfray et al. 2010).
3 Plant growth promoting bacteria
PGPB are present in the soil zone surrounding the plant root known as rhizosphere (Walker et al. 2003). Plant rhizosphere is rich in PGPB due to presence of root exudates, which acts as attractant for number of organisms present in soil (Singh and Strong 2016). Plant rhizosphere is helpful in faciliting nutrients and water to plants and provides continuous benefits to microbes present in rhizosphere, which promotes plant growth. The plant and microbe interaction in rhizosphere determines plant growth and soil fertility. Plant provides nutrients to PGPB in return PGPB endorse growth of plant through several plant growth promoting traits (Patel et al. 2015).
3.1 Role of PGPB in salinity stress amelioration in plants
The main challenge in agriculture field is development of technologies especially for stressed soil which endow agriculture sustainability and increase crop yield (Gepstein and Glick 2013; Hamilton et al. 2016). Currently there has been interest in the use of beneficial organism known as PGPB in the agriculture field, which improves plant growth and performance through several physiological, biochemical and molecular pathway (Palacios and Bashan 2014).
Under non stress condition PGPB enhance the plant growth by common mechanisms however in stress condition some PGPB may be not able to perform efficiently because of the incapability to survive in stress condition. However some PGPB are well known for amelioration of many stresses including salinity stress (Forni et al. 2017). The improved plant growth by PGPB under stress condition is due to the some mechanisms such as lowering stress induced ethylene level by ACC deaminase activity, (Glick et al. 2007a; Saharan and Nehra 2011), improving plant nutrition through nitrogen fixation, phosphate solubilization and siderophore synthesis (Etesami and Beattie 2017; Etesami 2018), by synthesizing exopolysaccharides that decrease accumulation of sodium ions in plant root by binding to excessive sodium ions and preventing their translocation to plant leaves (Qin et al. 2016; Etesami and Beattie 2017), upregulating plant defense enzymes (catalase (CAT), glutathione reductase (GR), superoxide dismutase (SOD), ascorbate peroxidase (APX)) to have protection against oxidative stress (Islam et al. 2016; Qin et al. 2016), maintaining high K+/Na+ ratio by regulating expression of ion transporters hence protects against ion toxicity (Islam et al. 2016; Etesami 2018; Pérez-Alfocea et al. 2010), increasing accumulation of osmolytes such as proline, glycine betain and sugars (Creus et al. 2004), preserving high photosynthetic activity and stomatal conductance (del Amor and Cuadra-Crespo 2012) and altering hormonal status of plant (Arora et al. 2012).
Promotion of salinity stressed plant growth by plant growth promoting traits of PGPB is shown in Fig. 2. Amelioration of salinity stress and alleviation of plant growth has been observed in many plants after PGPB inoculation such as wheat (Egamberdieva 2009), rice (Nautiyal et al. 2013), cotton (Yao et al. 2010), oat (Sapre et al. 2018), chickpea (AbdAllah et al. 2018), chili pepper (Park et al. 2013), licorice (Egamberdieva et al. 2016), mung beans (Patel et al. 2015), mint (Bharti et al. 2014) and chinese cabbage (Lee et al. 2016). Table 1 shows development of salt tolerance in crops inoculated with PGPB with several plant growth promoting traits.

Salinity stress alleviation using PGPB
3.1.1 Reduction in stress ethylene level
Ethylene is gaseous plant hormone, widely known as stress hormone and produced by all plants (Mayak et al. 2004). Ethylene plays a key role in plant growth and causes physiological changes in plant at molecular level at low concentration (Glick et al. 2007b) and is regulated by biotic and abiotic stress factors (Hardoim et al. 2008). An elevated level of ethylene is synthesized when plants are exposed to different stress condition that causes increased damages to plants (Abeles and Heggestad 1973). The ethylene level is elevated under stress condition due to enhanced production of ACC, an immediate precursor of ethylene biosynthesis (Zapata et al. 2007). For maintaining normal plant growth, reduction in stress-induced ethylene is necessary that will alleviate effect of salinity on plant growth (Glick 2004; Etesami and Beattie 2017).
PGPB facilitate the plant growth in saline condition by producing key enzyme known as ACC deaminase which catabolise ACC to ammonia and α-ketobutyrate (Glick et al. 2007a). The reduced level of ACC results in lowering stress ethylene concentration in plant root vicinity and helpful for promoting plant growth. Although salinity stress has been associated with loss of ACC deaminase activity in some PGPB (Upadhyay et al. 2009) however some PGPB maintains ACC deaminase activity even in the presence of salinity and helps the plants to tolerate stress by reducing stress ethylene level (Mayak et al. 2004). Bhise et al. (2017a) isolated plant growth promoting salt tolerant organism Enterobacter cloacae strain KBPD from salt affected soil of Kolhapur, MS, India. Following inoculation of V. radiata by strain KBPD, significant improvement in plant growth was observed even in presence of 50, 100 and 150 mM NaCl stress. Similarly Hartmannibacter diazotrophicus E19T exhibited ACC deaminase activity, isolated from salinity affected rhizosphere of Plantago winteri. When strain E19T added to 200 mM salinity stressed barley seedlings a decreased ethylene emission and significant improved root, shoot dry weight and water contents in root was observed (Suarez et al. 2015). Likewise, P. fluorescens with ACC deaminase activity when added to cucumber plant suffering from salinity stress (7 and 10 dS m−1) alleviation in salinity condition was noticed (Nadeem et al. 2016). Furthermore Klebsiella sp. strain IG 3 with expression of ACC deaminase activity and IAA production in salinity stress, when inoculated to oat (Avena sativa) under NaCl stress (100 mM) considerable improvement in shoot and root length, shoot and root dry weight and relative water content were observed (Sapre et al. 2018). Furthermore stress ethylene level increased under salinity decreases seed germination rate and plant root development (Belimov et al. 2001; Saravanakumar and Samiyappan 2007). Inoculation of salinity affected canola seeds (Brassica napus L.) by ACC deaminase producing P. fluorescens and P. putida significantly increased rate of germination (Jalili et al. 2009) and promoted plant growth. Salinity amelioration in plants by PGPB with ACC deaminase activity has been reported (Gontia-Mishra et al. 2014; Gontia-Mishra et al. 2017; Singh et al. 2015; Siddikee et al. 2011, 2012; Bacilio et al. 2016).
3.1.2 Maintenance of nutitional and ionic balance
Salinity affects plant growth by nutritional imbalance as it increases sodium and decreases calcium and potassium ions uptake. Maintenance of sodium ion exclusion and potassium ion acquisition has been in relation with salt tolerance in plants (Hauser and Horie 2010). Inoculation of salinity suffering plant by B. subtilis can down regulate high affinity potassium ion transporters (HKT1) that reduces excessive absorption of sodium ions by plant root thereby maintains ionic status of plant even in stress (Qin et al. 2016). Furthermore, these PGPB triggers HKT1 induction in inoculated plant shoot thereby facilitating recirculation of sodium ions from shoot to root (Zhang et al. 2008). A reduced sodium ion uptake was noticed in plant tissue when supplied with B. subtilis by regulating HKT1 which controls uptake of sodium ions (Zhang et al. 2008). Bacillus subtilis GB03 produces volatile organic compounds which prevent excessive accumulation of sodium ions in plant over control by regulating HKT1 in plant shoot and root exposed to salinity stress (Yang and Kloepper 2009). The salt tolerance in B. napus is may be due to the, increased activity of plasma membrane Na+/H+ exchanger ensuring enhanced sodium ion extrusion from plant root, activation of H+_ATPase and maintenance of negative membrane potential ensuring increased potassium retention and decreased sensitivity of K+-permeable channels of plant root to ROS (Chakraborty et al. 2016). A very strong correlation between salinity tolerance in plants and K+ retention have been reported for plants such as Brassica species (Chakrabortyetal 2016), Hordeum vulgare (Wu et al. 2015) and Triticum aestivum (Cuin et al. 2012). Supplementation of Azotobacter chroococcum reported to decrease sodium and increase calcium, magnesium and potassium ion level in plants (Rojas-Tapias et al. 2012). Similarly in other reports also decrease in sodium and increase in potassium ion concentration was noticed in plants upon PGPB inoculation (Bano and Fatima 2009; Kohler et al. 2009). The PGPB supplemented salinity stressed plants maintains adequate level of potassium ions to alleviate salinity originated sodium ion toxicity (Wang et al. 2016). A remarkably increased level of P, N, K+, Mg2+ and Ca2 has been noticed in roots and leaves of radish when seeds were pretreated with Pseudomonas fluorescens and Bacillus subtilis (Mohamed and Gomaa 2012). Inoculation of salt affected strawberry plant by Kocuria erythromyxa EY43 alleviates detrimental effects of salinity stress by increasing uptake of micro and macronutrients such as Mg, Mn, S, Cu, Fe, P, K and N plus decreases sodium ion concentration in root and leaves (Karlidag et al. 2013). An increase in phosphate contents of salinity stressed wheat plants was noticed when supplied with B. aquimaris (Upadhyay and Singh 2015). PGPB with nitrogen fixation, phosphate and potassium solubilization ability helps the plants in mobilization of these nutrients from soil and makes accisible form of nutrients available for luxiroius plant growth even under stress condition.
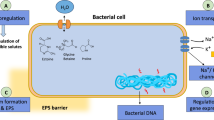
Some PGPB have an ability to assist the plant growing in salt stressed environment from detrimental effects of accumulated sodium ions by producing exopolysaccharides (Upadhyay and Singh 2015; Kasim et al. 2016). The produced exopolysaccharides binds to Na+ and reduces its availability, resulting in low sodium ions uptake hence maintains high K+/Na+ ratio (Upadhyay and Singh 2015). Inoculation of B. subtilis strain GB03 improves K+/Na+ in white clover (Trifolium repens L.) root and shoot in presence of 50, 100 and 150 mM salinity (Han et al. 2014). Similarly the exopolysaccharide synthesis by Enterobacter sp. P23 plays an important role in reducing sodium ion concentration in rice seedlings by binding to excess Na+ thus alleviate salinity stress (Sarkar et al. 2018). These results corroborates with Upadhyay et al. (2011) and Ashraf et al. (2004) for the wheat crops. Furthermore the PGPB produced exopolysaccharide binds to soil particles and forms macroaggregates plus microaggregates and enhance salinity stress resistance because of the improvement in soil structure (Sandhya et al. 2009). Exopolysaccharide production helps the inoculated PGPB to adhere to plant root which helps to exert beneficial effect of PGPB on plant and makes the PGPB as an ideal inoculum to salt suffering soil (Bhise et al. 2017a).
3.1.3 Protection from salinity induced oxidative stress
Salinity affected plants produces ROS which damages nucleic acid, proteins and lipids and are generated by plasma membrane, cell wall, peroxisome, mitochondria and chloroplast (Das and Roychoudhury 2014). The prolonged salinity decreases ROS scavenging enzymes such as CAT, SOD and APX and increases lipid peroxidation (Halo et al. 2015). Salt tolerance can be developed in plants by increasing activity of antioxidant enzymes (Hernández et al. 2003). Bacterization of salinity affected plant by PGPB mitigates stress by increasing ROS scavenging enzyme activity (Bianco and Defez, 2009). Kadmiri et al. (2018) showed increase in activity of POD of wheat under 5% salt stress when bacterized with P. fluorescens Ms-01 and P. fluorescens Ms-01 + A. brasilense DSM1690 in comparison with uninoculated plants, indicating role of PGPB in protecting plants against salinity stimulated oxidative stress. Moreover increase in enzyme activity of CAT and POD were reported by Islam et al. (2016) when 9 dS m−1 salt stressed Vigna radiata supplied with Bacillus cereus Pb25. Similar results of difference in antioxidant enzyme activity of salt stressed plants and bacterized salt stressed plants were reported by Kohler et al. (2010) for lettuce plants supplied with Pseudomonas mendocin, Gururani et al. (2013) for Solanum tuberosum added with Bacillus strains and Egamberdieva et al. (2017) in tomato plants with P. extremorientalis TSAU20. The increase in enzyme activity of ROS scavenging enzyme in salt stressed plants with PGPB inoculation may be because of the PGPB effect on antioxidant enzyme encoding genes (Gururani et al. 2013).
Lipid peroxidation causes increased level of melondialdehyde (MDA), indicates damaged structural integrity of cell membrane however bacterial inoculation to salinity stressed plant reduces MDA accumulation and assist the plant to recover from salinity induced toxic effects (Wu et al. 2014). Furthermore, PGPB supplementation to salinity affected plants enhances level of defensive enzymes and phenol which develops resistance in plants against salinity (Sharma and Sharma 2017). A decrease in MDA level was noticed by Sarkar et al. (2018) in salinity stressed rice seedlings in presence of bacterial inoculation of Enterobacter sp. P23. Similarly, Pseudomonas pseudoalcaligenes promotes growth of salinity affected rice GJ-17 by reducing lipid peroxidation (Jha and Subramanian 2014). These results corroborates with Singh and Jha (2016a) who reported reduced MDA level in wheat plants upon inoculation of Serratia sp. SL-12 and Sapre et al. (2018) in oat plant upon Klebsiella sp. IG 3 inoculation. The lower malondialdehyde content in bacterized salinity affected plants was reported by other studies also (Bharti et al. 2014; Barnawal et al. 2014; Jha and Subramanian 2014).
3.1.4 Protection from salinity induced osmotic stress
In salinity condition soil-water potential changes which decreases water uptake by plant. Under such condition plant accumulates compatible solutes such as proline, trehalose, glycine betaine and some organic solutes, playing vital role in plant protection by osmotic adjustment that enhances water uptake, diluting toxic ion concentration and limiting water loss (Ashraf et al. 2013; Slama et al. 2006). Proline accumulation in salt stress protects the plants by osmotic pressure adjustment and stabilization of plant functional units like proteins, enzymes and electron transport system-complex II (Ashraf and Foolad 2007; Mäkelä et al. 2000). Egamberdieva et al. (2017) observed increased level of proline when salt stressed plants were added with P. extremorientalis TSAU20. The salinity stressed Acacia gerrardii inoculated with Bacillus subtilis enhanced the proline level resulting in salt tolerance by maintaining water balance in plant tissue (Hashem et al. 2016). This result corroborates with Kruasuwan and Thamchaipenet (2018) for sugarcane in presence of Enterobacter sp. EN-21. The Bacillus pumilus and P. pseudoalcaligenes inoculation to rice plants increased concentration of glycine betaine and developes salt tolerance (Jha et al. 2011). Furthermore addition of Dietzia natronolimnaea strain STR1 conferes salt tolerance in wheat plants and exhibited elevated level of proline and antioxidants, enabling wheat plant to survive in stress (Bharti et al. 2016).
3.1.5 Improvement in photosynthesis
Salinity tolerance in plants is positively correlated with photosynthetic contents. Inoculation of salinity affected plants with PGPB increases photosynthetic activity and improves plant growth (Kang et al. 2014a). Salinity suffering plant having decreased synthesis of photosynthetic pigment when supplemented with Bacillus subtilis increased synthesis of chlorophyll a, b and carotenoid was observed (Abeer et al. 2015). Increase in chlorophyll content in salinity stressed plants provided with PGPB inoculum is reported in Arachis hypogaea (Shukla et al. 2012), Mentha arvensis (Bharti et al. 2014), Avena sativa (Sapre et al. 2018), Vigna radiata (Bhise et al. 2017a), Triticum aestivum (Singh and Jha 2016b) and sugarcane (Kruasuwan and Thamchaipenet, 2018). The PGPB inoculation to plants stimulates photosynthetic pigment synthesis might because of the increased uptake of nitrogen, phosphorous and potassium. Dawwam et al. (2013) demonstrated mitigation in salinity induced toxicity on photosynthetic machinery of C. arietinum by inoculation with B. subtilis through promotion of pigment synthesis and its associated components (Abd Allah et al. 2018). Similarly chlorophyll a contents of cotton plant exposed to 0.7% salinity stress was less than non stressed plants while inoculation with K. oxytoca Rs-5 significantly improved chlorophyll a concentration. This increase may be because of the increased plant biomass as dry matter accumulation was significantly increased so the leaf biomass (Wu et al. 2014). Inoculation of 150 mM NaCl stressed chickpea (Cicer arietinum) with ACC deaminase and IAA producing Pantoea dispersa PSB3 ameliorate salinity stress by reducing sodium ion uptake, electrolyte leakage and improving potassium ion uptake and chlorophyll content (Panwar et al. 2016).
3.1.6 Regulation of phytohormones
Salinity stress disturbs plant hormone balance hence hormone homeostasis under salinity condition might be the possible mechanism involved in phytohormone-originated tolerance in plants (Alqarawi et al. 2014; Iqbal and Ashraf 2013). The ability of plant to acclimatize to stress depends on their interaction with microbes that has IAA, GA and cytokinin producing ability (Berg et al. 2013). Phytohormones regulate tolerance in plants against salinity (Ryu and Cho 2015) and develop plant protective response to stress (Raghavan et al. 2006). These phytohormones affect proliferation of cell in root system and increases water and nutrient uptake due to overproduction of root hairs and lateral roots (Arora et al. 2013). Salinity stress reduces IAA content in plants thus causes stomatal closure (Dunlap and Binzel 1996) and distruption in cell wall extension and plasticity (Ribaut and Pilet 1994). PGPB with IAA production ability stimulate endogenous synthesis of IAA hence compensate salinity induced IAA reduction in plants (Liu et al. 2013). Kadmiri et al. (2018) investigated role of IAA producing A. brasilense DSM1696 and P. fluorescens Ms-01 on salinity stress alleviation in wheat crops. Author showed significant improvement in plant height and weight in presence of 5% salinity stress when added with studied organisms indicating role of IAA in plant growth promotion. Furthermore, IAA produced by PGPB stimulates ACC deaminase activity through signalling cascade that hydrolyze ACC into a-ketobutyrate and ammonia (Glick 2005) hence help the plant grow under salinity stress by reducing stress ethylene level. P. aureantiaca TSAU22 and P. extremorientalis TSAU20 with IAA production ability when inoculated to Sylebum marianum mitigates salinity stress (Egamberdieva et al. 2013). Asim et al. (2013) noticed reduced IAA level in salinity affected soyabean seedling while PGPB inoculation improved phytohormone level in plant. IAA changes root morphology resulting in improved plant growth and yield. A. brasilense, a salt tolerant organism isolated from salinity affected soil of Algeria near Mediterranean coast produced IAA even in salt stress. Supplementation of duran wheat plants by A. brasilense ameliorated salinity stress and promoted plant growth (Nabti et al. 2007, 2010). Sarkar et al. (2018) demonstrated rice seed germination of 48% in 150 mM salinity stress and 76% in same salinity stress when inoculated with IAA producing Enterobacter sp. P23, indicating alleviation in salinity stress in rice seed due to bacterial inoculation. The role of IAA in salinity stress tolerance and plant growth promotion has been identified in several studies (Ali and Abbas 2003; Kaya et al. 2013, 2010; Saba et al. 2013).
Salinity stress reduces GA synthesis in plants while PGPB inoculation triggers endogenous level of GA (Shahzad et al. 2016) and activates salinity stress mitigation strategies to prevent damages (Kang et al. 2014b). Gibberellic acids influences many processes of plant growth and development includes cell division, cell elongation, stem growth, meristem size of root and leaf (Guo et al. 2015; Martínez et al. 2016) fruit setting and seed germination (Hedden and Phillips 2000). GA mitigates salinity induced adverse effects on water use efficiency and water plant relationship (Yamaguchi 2008). GA application improved water use efficiency in tomato plants in saline soil (Maggio et al. 2010). In addition, inoculation of salinity affected soyabean by GA producing Pseudomonas putida H-2-3 mitigates salinity stress (Kang et al. 2014c). Under salinity, GA increases magnesssium and nitrogen level in root and leaves (Tuna et al. 2008). GA ameliorates salinity stress in mung beans by increasing protein synthesis, reducing sugars and activity of antioxidant enzymes and by reducing ribonuclease activity (Mohammed 2007). An increased endogenous level of GA in PGPB such as Acinetobacter calcoaceticus SE370, Burkholdera cepacia SE4 and Promicromonospora spp. SE188 inoculated cucumber plants over control plants improved plant growth under salinity (Kang et al. 2014a). IAA and GA producing Bacillus subtilis and Pseudomonas fluorescens when used for pretreatment of radish seed results in increase in salinity tolerance (Mohamed and Gomaa 2012). PGPB with IAA and GA producing ability promotes plant growth and seed germination under salinity (Kang et al. 2014a, 2014b) by enhancing α-amylase activity in seed, resulting in starch solubilization into simple sugars (Kim et al. 2006).
There are few reports on the role of bacterial synthesized ABA on its status in plant under salinity stress. However PGPB changes ABA status of plants and signaling pathways mediated by ABA that may results in enhanced salt tolerance in plants. Bharti et al. (2016) showed salinity tolerance in wheat plants by changing ABA signaling cascade when inoculated with Dietzia natronolimnaea STR1. Inaddition auxin and ABA production by Bacillus amyloliquefaciens RWL-1 induce salinity tolerance in rice plant (Shahzad et al. 2017). ABA mitigates inhibitory effect of salinity on plant growth due to accumulation of calcium and potassium, proline and sugar in root that helps to neutralize inhibitory effects of sodium and chlorine ions (Jaschke et al. 1997; Popova et al. 1995). Halotolerant bacteria with phytohormones such as IAA, GA and ABA producing ability when inoculated to soyabean plants alleviated salinity stress and improved shoot and root length and dry biomass under salinity condition (Naz et al. 2009). Research work carried out on IAA, GA and ABA showed that these hormones mitigated salinity stress and improved plant growth when PGPB P. chlororaphis and P. extremorientalis inoculated in salinity stressed common beans (Egamberdieva 2011; Jha and Subramanian 2013).
PGPB inoculation to salt stressed plant can manipulate cytokinins concentration of plant by shifting cytokinins homeostasis or by producing cytokinins (Glick 2012; Pallai et al. 2012; Kapoor and Kaur 2016). An increased level of cytokinin in plant shoot decreases ethylene level and leaf senescence in tomato plant hence supports plant growth under salinity condition (Ghanem et al. 2010). In plant growth cytokinins acts as master regulator and known to mitigate salinity stress in plants (Fahad et al. 2015). However, the actual role of cytokinin in salinity tolerance in plant is not clear.
3.1.7 Proteogenomic level
Proteomic approach provides information regarding expression of whole proteins and their interaction in cell (Paul et al. 2006). Salt tolerant plants have altered activity of proteins. Studies on molecular mechanism regulating response of plants against salinity mainly focus on transcription changes (Brumós et al. 2009). Kandasamy et al. (2009) showed expression of 23 proteins during study of plant growth promotional property of Pseudomonas fluorescens strain KH-1 in rice leaf sheaths. Salt stressed plants and bacterized salt stressed plants express variation in antioxidative processes, response against pathogens and proteins of photosynthesis (Cheng et al. 2012). Salt stressed plants in presence of bacteria express increase in proteins related to deoxyribonucleic acid (DNA) synthesis hence protects Arabidopsis plants from salinity (Arimura et al. 2004). Furthermore, plants inoculated with PGPB provide protection against biotic and abiotic stress by increasing chitinase and 20S proteasome activity (Cheng et al. 2012).
Gene expression and their role in salt tolerance can be studied by transcriptomic analysis. Genomic and transcriptomic are used to characterize number of transcription factor and genes. Supplementation of Burkholderia phytofirmans PsJN in medium increases genes such as ROS scavenging (APX2), responsive-to-dessication 29B (RD29B) and relative-to-dessication 29A (RD29A) and detoxification (Glyoxalase 7) and reduces lipoxygenase-2 in salinity affected Arabidopsis plants (Pinedo et al. 2015). Similarly Medicago truncatula inoculation by bacteria alleviates salinity stress by reducing ethylene-signaling genes expression (Bianco and Defez, 2009). PGPB inoculation increases gene expression encoding for APX, CAT, DHAR (Dehydroascorbate reductase) and GR (Glutathione reductase) (Habib et al. 2016).
3.2 Adavantages of using PGPB
Agriculture field provides food security to worldwide increasing human population. Now farmers are facing major problems such as increasing soil salinity stress, declined soil fertility and overall crop yield. Improvement in soil fertility and crop yield is main aim of most of the countries. PGPB present in soil acts as powerful agents in maintaining soil fertility, enhancing plant growth and nutrient acquisition, constituting a key resource for sustainable agricultural practices. The term bio-fertilizer mostly refers to use of PGPB to increase the nutrient contents of plants by combination of various activities (Whipps 2001).
Use of PGPB as biofertilizer has several advantages in agriculture field which includes, 1) PGPB as a bioinoculant restore and reclaim wasteland by converting nonfertile soil into fertile soil 2) Reduces the use of chemical based fertilizers that consequently leading to reduce the production cost. The usual annual production cost of nitrogen fertilizer is USD 30 million that would be reduced to 1 million by the replacement of chemical fertilizers by biofertilizers (Herridge 2008). 3) Decreases the pollution problems and land degradation arises because of the use of chemical based fertilizers and constitutes environmentally friendly approach (Stefan et al. 2008). 4) Maintains cation exchange capacity and structure of soil. 5) Significantly improves plant growth, quality and yield by controlling growth of phytopathogens thus ensures food security. Other advantages include safety, targeted activity, effective in small amount and self multiplication.
4 Conclusion
Salinity is one of the major constraints for agriculture yield, economy and food security. Salinity affects plant growth at morphological, biochemical, physiological and molecular level. Now it’s inevitable to have promising agriculture practice that helps plant to withstand salinity stress and also increases agricultural productivity. PGPB can be considered as ecofriendly and cost effective approach to combat salinity stress and improve crop yield. Under salinity condition, ACC deaminase secretion by PGPB reduces stress ethylene accumulation and significantly improves plant growth. PGPB promotes the plant growth by enriching soil with vital nutrients such as nitrogen, phosphorous and potassium. Salinity induced decrease in hormones level is compensated by secretion of plant hormones such as IAA, GA, ABA and cytokinin by PGPB. Exopolysaccharide produced by PGPB maintains ionic balance of plant by preventing transport of accumulated sodium to plant root by binding with the excess sodium ions. Bacterization of salt stressed plants by PGPB enhances antioxidants, which alters redox status of plants thus improving photosynthesis efficiency as well as accumulate compatible solutes such as proline, which avoids stress induced damages by regulating cellular activities. The present reviews focal point is key role of PGPB in salt stress tolerance and promotion of plant growth and development. Considering this scenario future research is needed to develop bacterial consortia to saline soil having multiple plant growth promoting traits expressing PGPB. Understanding of interaction between PGPB present in consortia and consortia plus plant will pave means to exploit much more benefits from PGPB for escalating salinity stress mitigation in plants.
References
Abd Allah EF, Alqarawi AA, Hashem A, Radhakrishnan R, Al-Huqail AA, Al-Otibi FON, Egamberdieva D (2018) Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J Plant Interact 13:37–44
Abeer H, Abd_Allah EF, Alqarawi AA, Al-Huqail AA (2015) Impact of plant growth promoting Bacillus subtilis on growth and physiological parameters of Bassia indica (Indian bassia) grown udder salt stress. Pak J Bot 47:1735–1741
Abeles FB, Heggestad HE (1973) Ethylene: an urban air pollutant. J Air Pollut Control Assoc 23:517–521
Acquaah G (2007). Principles of plant genetics and breeding. Oxford: Blackwell, pp385
AE Government report (2001) Salt contamination assessment and remediation guidelines in: division ES, editor. Albarta Environment, Edonton
Ahmad M, Zahir ZA, Nazli F, Akram F, Arshad M, Khalid M (2013) Effectiveness of halo-tolerant, auxin producing Pseudomonas and Rhizobium strains to improve osmotic stress tolerance in mung bean (Vigna radiata L.). Braz J Microbiol 44:1341–1348
Ahmad P, Nabi G, Ashraf M (2011) Cadmium-induced oxidative damage in mustard [Brassica juncea (L.) Czern. & Coss.] plants can be alleviated by salicylic acid. South Afr J Bot 77:36–44
Ali S, Charles TC, Glick BR (2014) Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol Biochem 80:160–167
Ali RM, Abbas HM (2003) Response of salt stressed barley seedlings to phenylurea. Plant Soil and Environment 49(4):158–162
Allakhverdiev SI, Sakamoto A, Nishiyama Y, Inaba M, Murata N (2000) Ionic and osmotic effects of NaCl-induced inactivation of photosystems I and II in Synechococcus sp. Plant Physiol 123:1047–1056
Alqarawi A, Hashem A, Abd_Allah E, Alshahrani T, Huqail A (2014) Effect of salinity on moisture content, pigment system, and lipid composition in Ephedra alata Decne. Acta Biol Hung 65(1):61–71
Amacher J K, Koenig R, Kitchen B (2000) Salinity and plant tolerance Utah State University, no. AG-S0-03
Ansari O, Zadeh FS (2012) Osmo and hydro priming mediated germination improvement under cold stress conditions in mountain rye (Secale montanum). Cercetări Agron Moldova 45:53–62
Arimura SI, Aida GP, Fujimoto M, Nakazono M, Tsutsumi N (2004) Arabidopsis dynamin-like protein 2a (ADL2a), like ADL2b, is involved in plant mitochondrial division. Plant Cell Physiol 45:236–242
Arora NK, Tewari S, Singh R (2013) Multifaceted plant-associated microbes and their mechanisms diminish the concept of direct and indirect PGPRs. In: Arora NK (ed) plant microbe Symbiosis: fundamentals and advances springer, New Delhi, pp. 411-449
Arora NK, Tewari S, Singh S, Lal N, Maheshwari DK (2012) PGPR for protection of plant health under saline conditions. In: Maheshwari DK (ed) Bacteria in agrobiology: stress management springer. Heidelberg p, Berlin, pp 239–258
Ashraf M, Shahbaz M, Ali Q (2013) Drought-induced modulation in growth and mineral nutrients in canola (Brassica napus L.). Pak J Bot 45:93–98
Ashraf M, Hasnain S, Berge O, Mahmood T (2004) Inoculating wheat seedlings with exopolysaccharide-producing bacteria restricts sodium uptake and stimulates plant growth under salt stress. Biol Fert Soils 40:157–162
Ashraf MFMR, Foolad M (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot 59:206–216
Asim M, Aslam M, Bano A, Munir M, Majeed A, Abbas SH (2013) Role of phytohormones in root nodulation and yield of soybean under salt stress. Am J Res Commun 1:191–208
Bacilio M, Moreno M, Bashan Y (2016) Mitigation of negative effects of progressive soil salinity gradients by application of humic acids and inoculation with Pseudomonas stutzeri in a salt-tolerant and a salt-susceptible pepper. Appl Soil Ecol 107:394–404
Bano A, Fatima M (2009) Salt tolerance in Zea mays (L). Following inoculation with Rhizobium and Pseudomonas. Biol Fert Soils 45:405–413
Barnawal D, Bharti N, Maji D, Chanotiya CS, Kalra A (2014) ACC deaminase-containing Arthrobacter protophormiae induces NaCl stress tolerance through reduced ACC oxidase activity and ethylene production resulting in improved nodulation and mycorrhization in Pisum sativum. J Plant Physiol 171:884–894
Belimov AA, Safronova VI, Sergeyeva TA, Egorova TN, Matveyeva VA, Tsyganov VE, Dietz KJ (2001) Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can J Microbiol 47:642–652
Berg G, Martinez JL (2015) Friends or foes: can we make a distinction between beneficial and harmful strains of the Stenotrophomonas maltophilia complex? Front Microbiol 6:241
Berg G, Alavi M, Schmidt CS, Zachow C, Egamberdieva D, Kamilova F, Lugtenberg BJ (2013) Biocontrol and osmoprotection for plants under salinated conditions. In: de Bruijn Frans J (ed) Molecular microbial ecology of the rhizosphere, Wiley-Blackwell, USA, vol 1, pp 561–573
Bharti N, Barnawal D, Awasthi A, Yadav A, Kalra A (2014) Plant growth promoting rhizobacteria alleviate salinity induced negative effects on growth, oil content and physiological status in Mentha arvensis. Acta Physiol Plant 36:45–60
Bharti N, Pandey SS, Barnawal D, Patel VK, Kalra A (2016) Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci Rep 6:34768
Bhise KK, Bhagwat PK, Dandge PB (2017b) Synergistic effect of Chryseobacterium gleum sp. SUK with ACC deaminase activity in alleviation of salt stress and plant growth promotion in Triticum aestivum L. 3. Biotech 7:105
Bhise KK, Bhagwat PK, Dandge PB (2017a) Plant growth-promoting characteristics of salt tolerant Enterobacter cloacae strain KBPD and its efficacy in amelioration of salt stress in Vigna radiata L. J Plant Growth Regul 36:215–226
Bhise, K. K., & Dandge, P. B. (2019). Alleviation of salinity stress in rice plant by encapsulated salt tolerant plant growth promoting bacteria Pantoea agglomerans strain KL and its root colonization ability. Archives of Agronomy and Soil Science, (just-accepted)
Bianco C, Defez R (2009) Medicago truncatula improves salt tolerance when nodulated by an indole-3-acetic acid-overproducing Sinorhizobium meliloti strain. J Exp Bot 60:3097–3107
Brumós J, Colmenero-Flores JM, Conesa A, Izquierdo P, Sánchez G, Iglesias DJ, Talón M (2009) Membrane transporters and carbon metabolism implicated in chloride homeostasis differentiate salt stress responses in tolerant and sensitive Citrus rootstocks. Funct Integr Genomics 9:293–309
Carmen B, Roberto D (2011) Soil bacteria support and protect plants against abiotic stresses. In:Shanker A (ed) abiotic stress in plants-mechanisms and adaptations. Pub. InTech, pp. 143–170
Chakraborty K, Bose J, Shabala L, Shabala S (2016) Difference in root K+ retention ability and reduced sensitivity of K+-permeable channels to reactive oxygen species confer differential salt tolerance in three Brassica species. J Exp Bot 67:4611–4625
Cheng Z, Woody OZ, McConkey BJ, Glick BR (2012) Combined effects of the plant growth-promoting bacterium Pseudomonas putida UW4 and salinity stress on the Brassica napus proteome. Appl Soil Eco 61:255–263
Creus CM, Sueldo RJ, Barassi CA (2004) Water relations and yield in Azospirillum-inoculated wheat exposed to drought in the field. Can J Bot 82:273–281
Cuin TA, Zhou M, Parsons D, Shabala S (2012) Genetic behaviour of physiological traits conferring cytosolic K+/Na+ homeostasis in wheat. Plant Biol 14:438–446
Das K, Roychoudhury A (2014) Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front Environ sci 2:53
Dawwam GE, Elbeltagy A, Emara HM, Abbas IH, Hassan MM (2013) Beneficial effect of plant growth promoting bacteria isolated from the roots of potato plant. Annal Agri Sci 58:195–201
Del Amor FM, Cuadra-Crespo P (2012) Plant growth-promoting bacteria as a tool to improve salinity tolerance in sweet pepper. Funct Plant Biol 39:82–90
Dodd IC, Pérez-Alfocea F (2012) Microbial amelioration of crop salinity stress. J Exp Bot 63:3415–3428
Dunlap JR, Binzel ML (1996) NaCI reduces indole-3-acetic acid levels in the roots of tomato plants independent of stress-induced abscisic acid. Plant Physiol 112(1):379–384
Egamberdieva D (2009) Alleviation of salt stress by plant growth regulators and IAA producing bacteria in wheat. Acta Physiol Plant 31:861–864
Egamberdieva D, Davranov K, Wirth S, Hashem A, Abd_Allah, E. F (2017) Impact of soil salinity on the plant-growth–promoting and biological control abilities of root associated bacteria. Saudi J Biol Sci 24:1601–1608
Egamberdieva D, Jabborova D, Hashem A (2015) Pseudomonas induces salinity tolerance in cotton (Gossypium hirsutum) and resistance to Fusarium root rot through the modulation of indole-3-acetic acid. Saudi J Biol Sci 22:773–779
Egamberdieva D, Li L, Lindström K, Räsänen LA (2016) A synergistic interaction between salt-tolerant Pseudomonas and Mesorhizobium strains improves growth and symbiotic performance of liquorice (Glycyrrhiza uralensis fish.) under salt stress. Appl Microbiol Biotechnol 100:2829–2841
Egamberdieva D (2011) Survival of Pseudomonas extremorientalis TSAU20 and P. chlororaphis TSAU13 in the rhizosphere of common bean (Phaseolus vulgaris) under saline conditions. Plant Soil Environ 57(3):122–127
Egamberdieva, D., Berg, G., Lindström, K. and Räsänen, L.A. (2013). Alleviation of salt stress of symbiotic Galega officinalis L. (Goat’s rue) by co-inoculation of rhizobium with root colonising Pseudomonas. Plant soil (online first)
Etesami H (2018) Can interaction between silicon and plant growth promoting rhizobacteria benefit in alleviating abiotic and biotic stresses in crop plants? Agric Ecosyst Environ 253:98–112
Etesami H, Beattie GA (2017) Plant-microbe interactions in adaptation of agricultural crops to abiotic stress conditions. In Probiotics and Plant Health Springer, Singapore pp:163–200
Fahad S, Hussain S, Bano A, Saud S, Hassan S, Shan D, Tabassum MA (2015) Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: consequences for changing environment. Environ Sci Pollut Res 22:4907–4921
Forni C, Duca D, Glick BR (2017) Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 410:335–356
Foyer CH, Rasool B, Davey JW, Hancock RD (2016) Cross-tolerance to biotic and abiotic stresses in plants: a focus on resistance to aphid infestation. J Exp Bot 67:2025–2037
Ge H, Zhang F (2019) Growth-promoting ability of Rhodopseudomonas palustris G5 and its effect on induced resistance in cucumber against salt stress. J Plant Growth Regul 38(1):180–188
Gepstein S, Glick BR (2013) Strategies to ameliorate abiotic stress-induced plant senescence. Plant Mol Biol 82:623–633
Ghanem ME, Albacete A, Smigocki AC, Frébort I, Pospisilová H, Martínez-Andújar C et al (2010) Root-synthesized cytokinins improve shoot growth and fruit yield in salinized tomato (Solanum lycopersicum L.) plants. J Exp Bot 62(1):125–140
Glick BR (2004) Bacterial ACC deaminase and the alleviation of plant stress. Adv Appl Microbiol 56:291–312
Glick BR (2005) Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol Lett 251:1–7
Glick BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Hindawi Publishing Corporation, Scientifica
Glick BR, Cheng Z, Czarny J, Duan J (2007a) Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur J Plant Pathol 119:329–339
Glick BR, Todorovic B, Czarny J, Cheng Z, Duan J, McConkey B (2007b) Promotion of plant growth by bacterial ACC deaminase. Crit Rev Plant Sci 26:227–242
Godfray HCJ, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, Toulmin C (2010) Food security: the challenge of feeding 9 billion people. Science 327:812–818
Gontia-Mishra I, Sapre S, Kachare S, Tiwari S (2017) Molecular diversity of 1-aminocyclopropane-1-carboxylate (ACC) deaminase producing PGPR from wheat (Triticum aestivum L.) rhizosphere. Plant Soil 414:213–227
Gontia-Mishra I, Sasidharan S, Tiwari S (2014) Recent developments in use of 1-aminocyclopropane-1-carboxylate (ACC) deaminase for conferring tolerance to biotic and abiotic stress. Biotechnol Lett 36:889–898
Guo H, Wang Y, Liu H, Hu P, Jia Y, Zhang C, Wang C (2015) Exogenous GA3 application enhances xylem development and induces the expression of secondary wall biosynthesis related genes in Betula platyphylla. Int J Mol Sci 16:22960–22975
Gururani MA, Upadhyaya CP, Baskar V, Venkatesh J, Nookaraju A, Park SW (2013) Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J Plant Growth Regul 32:245–258
Habib SH, Kausar H, Saud HM (2016) Plant growth-promoting rhizobacteria enhance salinity stress tolerance in okra through ROS-scavenging enzymes Biomed Res Internat:6284547
Halo BA, Khan AL, Waqas M, Al-Harrasi A, Hussain J, Ali L, Lee IJ (2015) Endophytic bacteria (Sphingomonas sp. LK11) and gibberellin can improve Solanum lycopersicum growth and oxidative stress under salinity. J Plant Interact 10:117–125
Hamilton CE, Bever JD, Labbé J, Yang X, Yin H (2016) Mitigating climate change through managing constructed-microbial communities in agriculture. Agric Ecosyst Environ 216:304–308
Han QQ, Lü XP, Bai JP, Qiao Y, Paré PW, Wang SM, Wang ZL (2014) Beneficial soil bacterium Bacillus subtilis (GB03) augments salt tolerance of white clover. Front Plant Sci 5:525
Hardoim PR, van Overbeek LS, van Elsas JD (2008) Properties of bacterial endophytes and their proposed role in plant growth. Trends microbial 16:463–471
Hashem A, Abd_Allah EF, Alqarawi AA, Al-Huqail AA, Wirth S, Egamberdieva D (2016) The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front Microbiol 7:1089
Hauser F, Horie T (2010) A conserved primary salt tolerance mechanism mediated by HKT transporters: a mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress. Plant Cell Environ 33:552–565
Hedden P, Phillips AL (2000) Gibberellin metabolism: new insights revealed by the genes. Trends Plant Sci 5:523–530
Hernández JA, Aguilar AB, Portillo B, López-Gómez E, Beneyto JM, García-Legaz MF (2003) The effect of calcium on the antioxidant enzymes from salt-treated loquat and anger plants. Funct Plant Biol 30:1127–1137
Herridge DF (2008) Inoculation technology for legumes. In: Dilworth MJ et al (eds) Nitrogen-fixing leguminous symbioses. Springer. Dordrecht, The Netherlands, pp 77–115
Iqbal M, Ashraf M (2013) Gibberellic acid mediated induction of salt tolerance in wheat plants: growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ Exp Bot 86:76–85
Islam F, Yasmeen T, Ali S, Ali B, Farooq MA, Gill RA (2015) Priming-induced antioxidative responses in two wheat cultivars under saline stress. Acta Physiol Plant 37:153
Islam F, Yasmeen T, Arif MS, Ali S, Ali B, Hameed S, Zhou W (2016) Plant growth promoting bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul 80:23–36
Jadhav GG, Salunkhe DS, Nerkar DP, Bhadekar RK (2010) Isolation and characterization of salt-tolerant nitrogen-fixing microorganisms from food. Eurasia J Biosci 4:33–40
Jalili F, Khavazi K, Pazira E, Nejati A, Rahmani HA, Sadaghiani HR, Miransari M (2009) Isolation and characterization of ACC deaminase-producing fluorescent pseudomonads, to alleviate salinity stress on canola (Brassica napus L.) growth. J Plant Physiol 166:667–674
Jamil A, Riaz S, Ashraf M, Foolad MR (2011) Gene expression profiling of plants under salt stress. Crit Rev Plant Sci 30:435–458
Jaschke WD, Peuke AD, Pate JS, Hartung W (1997) Transport, synthesis and catabolism of abscisic acid (ABA) in intact plants of castor bean (Ricinus communis L.) under phosphate deficiency and moderate salinity. J Exp Bot 48(9):1737–1747
Jha Y, Subramanian RB (2014) PGPR regulate caspase-like activity, programmed cell death, and antioxidant enzyme activity in paddy under salinity. Physiol Mol Biol Plant 20:201–207
Jha Y, Subramanian RB, Patel S (2011) Combination of endophytic and rhizospheric plant growth promoting rhizobacteria in Oryza sativa shows higher accumulation of osmoprotectant against saline stress. Acta Physiol Plant 33:797–802
Jha Y, Subramanian RB (2013) Paddy plants inoculated with PGPR show better growth physiology and nutrient content under saline condition. Chilean journal of agricultural research 73(3):213–219
Kadmiri IM, Chaouqui L, Azaroual SE, Sijilmassi B, Yaakoubi K, Wahby I (2018) Phosphate-solubilizing and auxin-producing Rhizobacteria promote plant growth under saline conditions. Arab J Sci Eng 43:3403–3415
Kafi M, Khan MA (2008) Crop and forage production using saline waters. Daya Books
Kandasamy S, Loganathan K, Muthuraj R, Duraisamy S, Seetharaman S, Thiruvengadam R, Ramasamy S (2009) Understanding the molecular basis of plant growth promotional effect of Pseudomonas fluorescens on rice through protein profiling. Prot Sci 7:47
Kang SM, Khan AL, Waqas M, You YH, Kim JH, Kim JG, Lee IJ (2014a) Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J Plant Interact 9:673–682
Kang SM, Khan AL, You YH, Kim JG, Kamran M, Lee IJ (2014b) Gibberellin production by newly isolated strain Leifsonia soli SE134 and its potential to promote plant growth. J Microbioz Biotechnol 24:106–112
Kang SM, Radhakrishnan R, Khan AL, Kim MJ, Park JM, Kim BR, Lee IJ (2014c) Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol Biochem 84:115–124
Kapoor R, Kaur M (2016) Cytokinins production by fluorescent Pseudomonas isolated from rhizospheric soils of Malus and Pyrus. Afr J Microbiol Res 10:1274–1279
Karlidag H, Yildirim E, Turan M, Pehluvan M, Donmez F (2013) Plant growth-promoting rhizobacteria mitigate deleterious effects of salt stress on strawberry plants (Fragaria× ananassa). Hortscience 48(5):563–567
Kasim WA, Gaafar RM, Abou-Ali RM, Omar MN, Hewait HM (2016) Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley. Ann Agri Sci 61:217–227
Kaveh H, Nemati H, Farsi M, Jartoodeh SV (2011) How salinity affect germination and emergence of tomato lines. J Biol Environ Sci 5:159–163
Kaya C, Ashraf M, Dikilitas M, Tuna AL (2013) Alleviation of salt stress-induced adverse effects on maize plants by exogenous application of indoleacetic acid (IAA) and inorganic nutrients-A field trial. Aust J Crop Sci 7(2):249
Kaya C, Tuna AL, Okant AM (2010) Effect of foliar applied kinetin and indole acetic acid on maize plants grown under saline conditions. Turk J Agric For 34(6):529–538
Khan MA, Weber DJ (2006) Ecophysiology of high salinity tolerant plants (Vol. 40). Springer Science & Business Media
Kim SK, Son TK, Park SY, Lee IJ, Lee BH, Kim HY, Lee SC (2006) Influences of gibberellin and auxin on endogenous plant hormone and starch mobilization during rice seed germination under salt stress. J Environ Biol 27:181–186
Kohler J, Hernández JA, Caravaca F, Roldán A (2009) Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ Exp Bot 65:245–252
Kohler J, Knapp BA, Waldhuber S, Caravaca F, Roldán A, Insam H (2010) Effects of elevated CO 2, water stress, and inoculation with Glomus intraradices or Pseudomonas mendocina on lettuce dry matter and rhizosphere microbial and functional diversity under growth chamber conditions. J Soils Sediments 10:1585–1597
Kruasuwan W, Thamchaipenet A (2018) 1-Aminocyclopropane-1-carboxylate (ACC) deaminase-producing endophytic Diazotrophic Enterobacter sp. EN-21 modulates salt–stress response in sugarcane. J Plant Growth Regul 37(3):849–858
Lee GW, Lee KJ, Chae JC (2016) Herbaspirillum sp. strain GW103 alleviates salt stress in Brassica rapa L. ssp. pekinensis. Protoplasma 253:655–661
Liu Y, Shi Z, Yao L, Yue H, Li H, Li C (2013) Effect of IAA produced by Klebsiella oxytoca Rs-5 on cotton growth under salt stress. J Gen Appl Microbiol 59(1):59–65
Maggio A, Barbieri G, Raimondi G, De Pascale S (2010) Contrasting effects of GA 3 treatments on tomato plants exposed to increasing salinity. J Plant Growth Regul 29:63–72
Mäkelä P, Kärkkäinen J, Somersalo S (2000) Effect of glycinebetaine on chloroplast ultrastructure, chlorophyll and protein content, and RuBPCO activities in tomato grown under drought or salinity. Biol Planta 43:471–475
Manchanda G, Garg N (2008) Salinity and its effects on the functional biology of legumes. Acta Physiol Plant 30:595–618
Martínez C, Espinosa-Ruiz A, Prat S (2016) Gibberellins and plant vegetative growth. Annu Plant Rev 49:285–322
Mayak S, Tirosh T, Glick BR (2004) Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol Biochem 42:565–572
Mohamed HI, Gomaa EZ (2012) Effect of plant growth promoting Bacillus subtilis and Pseudomonas fluorescens on growth and pigment composition of radish plants (Raphanus sativus) under NaCl stress. Photosynthetica 50:263–272
Mohammed AHMA (2007) Physiological aspects of mungbean plant (Vigna radiata L. Wilczek) in response to salt stress and gibberellic acid treatment. Res J Agr Biol Sci 3:200–213
Moradi F, Ismail AM (2007) Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann Bot 99:1161–1173
Munns R (2005) Genes and salt tolerance: bringing them together. New Phytol 167:645–663
Nabti E, Sahnoune M, Adjrad S, Van Dommelen A, Ghoul M, Schmid M, Hartmann A (2007) A halophilic and osmotolerant Azospirillum brasilense strain from Algerian soil restores wheat growth under saline conditions. Engineering in Life Sciences 7(4):354–360
Nabti E, Sahnoune M, Ghoul M, Fischer D, Hofmann A, Rothballer M, Schmid M, Hartmann A (2010) Restoration of growth of durum wheat (Triticum durum var. waha) under saline conditions due to inoculation with the rhizosphere bacterium Azospirillum brasilense NH and extracts of the marine alga Ulva lactuca. J Plant Growth Regul 29(1):6–22
Nadeem SM, Ahmad M, Naveed M, Imran M, Zahir ZA, Crowley DE (2016) Relationship between in vitro characterization and comparative efficacy of plant growth-promoting rhizobacteria for improving cucumber salt tolerance. Arch Microbiol 198:379–387
Naik RS (2014) Impact of soil salinity on agriculture in the lift irrigation commands of Shirol taluka. Shivaji University, Department of Economics
Nautiyal CS, Srivastava S, Chauhan PS, Seem K, Mishra A, Sopory SK (2013) Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol Biochem 66:1–9
Naz I, Bano A, Ul-Hassan T (2009) Isolation of phytohormones producing plant growth promoting rhizobacteria from weeds growing in Khewra salt range, Pakistan and their implication in providing salt tolerance to Glycine max L. Afr J Biotechnol 8(21)
Palacios OA, Bashan Y, de-Bashan LE (2014) Proven and potential involvement of vitamins in interactions of plants with plant growth-promoting bacteria- an overview. Biol Fertil Soils 50:415–432
Palaniyandi SA, Damodharan K, Yang SH, Suh JW (2014) Streptomyces sp. strain PGPA39 alleviates salt stress and promotes growth of ‘micro tom’ tomato plants. J Appl Microbiol 117:766–773
Pallai R, Hynes RK, Verma B, Nelson L. M (2012) Phytohormone production and colonization of canola (Brassica napus L.) roots by Pseudomonas fluorescens 6-8 under gnotobiotic conditions. Can J Microbiol 58:170–178
Panwar M, Tewari R, Gulati A, Nayyar H (2016) Indigenous salt-tolerant rhizobacterium Pantoea dispersa (PSB3) reduces sodium uptake and mitigates the effects of salt stress on growth and yield of chickpea. Acta Physiol Plant 38:278
Parihar P, Singh S, Singh R, Singh VP, Prasad SM (2015) Effect of salinity stress on plants and its tolerance strategies: a review. Environ Sci Pollut Res 22:4056–4075
Park JW, Balaraju K, Kim JW, Lee SW, Park K (2013) Systemic resistance and growth promotion of chili pepper induced by an antibiotic producing Bacillus vallismortis strain BS07. Biol Control 65:246–257
Patel BB, Patel BB, Dave RS (2011) Studies on infiltration of saline–alkali soils of several parts of Mehsana and Patan districts of North Gujarat. J Appl Technol Environ Sanitation 1:87–92
Patel RR, Patel DD, Thakor P, Patel B, Thakkar VR (2015) Alleviation of salt stress in germination of Vigna radiata L. by two halotolerant Bacilli sp. isolated from saline habitats of Gujarat. Plant Growth Regul 76:51–60
Pérez-Alfocea F, Albacete A, Ghanem ME, Dodd IC (2010) Hormonal regulation of source–sink relations to maintain crop productivity under salinity: a case study of root-to-shoot signalling in tomato. Funct Plant Biol 37:592–603
Pinedo I, Ledger T, Greve M, Poupin MJ (2015) Burkholderia phytofirmans PsJN induces long-term metabolic and transcriptional changes involved in Arabidopsis thaliana salt tolerance. Front Plant Sci 6:466
Popova, L. P., Stoinova, Z. G., & Maslenkova, L. T. (1995). Involvement of abscisic acid in photosynthetic process in Hordeum vulgare L. during salinity stress. Journal of plant growth regulation, 14(4), 211
Qadir M, Tubeileh A, Akhtar J, Larbi A, Minhas PS, Khan MA (2008) Productivity enhancement of salt-affected environments through crop diversification. Land Degrad Dev 19:429–453
Qin Y, Druzhinina IS, Pan X, Yuan Z (2016) Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture. Biotechnol Adv 34:1245–1259
Raghavan C, Ong EK, Dalling MJ, Stevenson TW (2006) Regulation of genes associated with auxin, ethylene and ABA pathways by 2, 4-dichlorophenoxyacetic acid in Arabidopsis. Funct Integr Genomics 6:60–70
Rashid MI, Mujawar LH, Shahzad T, Almeelbi T, Ismail IM, Oves M (2016) Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol Res 183:26–41
Ray DK, Mueller ND, West PC, Foley JA (2013) Yield trends are insufficient to double global crop production by 2050. PLoS One 8:1–8
Ribaut JM, Pilet PE (1994) Water stress and indol-3yl-acetic acid content of maize roots. Planta 193(4):502–507
Rojas-Tapias D, Moreno-Galván A, Pardo-Díaz S, Obando M, Rivera D, Bonilla R (2012) Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize (Zea mays). Appl Soil Eco 61:264–272
Rouhi HR, Aboutalebian MA, Sharif-Zadeh F (2011) Effects of hydro and osmopriming on drought stress tolerance during germination in four grass species. Int J Agric Sci 1:107–114
Ryu H, Cho YG (2015) Plant hormones in salt stress tolerance. J Plant Biol 58:147–155
Saba K, Muhammad P, Khalid N, Khalid H, Aleena A, Saba S et al (2013) Effect of indole acetic acid (IAA) on morphological, biochemical and chemical attributes of two varieties of maize (Zea mays l.) under salt stress. World Appl Sci J 26(9):1150–1159
Saharan BS, Nehra V (2011) Plant growth promoting rhizobacteria: a critical review. Life Sci Med Res 21:30
Sandhya VZAS, Grover M, Reddy G, Venkateswarlu B (2009) Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol Fertility Soil 46:17–26
Sapre S, Gontia-Mishra I, Tiwari S (2018) Klebsiella sp. confers enhanced tolerance to salinity and plant growth promotion in oat seedlings (Avena sativa). Microbiol Res 206:25–32
Saravanakumar D, Samiyappan R (2007) ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J Appl Microbiol 102:1283–1292
Sarkar A, Ghosh PK, Pramanik K, Mitra S, Soren T, Pandey S, Maiti TK (2018) A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res Microbiol 169:20–32
Shahzad R, Waqas M, Khan AL, Asaf S, Khan MA, Kang SM, Lee IJ (2016) Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol Biochem 106:236–243
Shahzad R, Khan AL, Bilal S, Waqas M, Kang SM, Lee IJ (2017) Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ Exp Bot 136:68–77
Sharma IP, Sharma AK (2017) Physiological and biochemical changes in tomato cultivar PT-3 with dual inoculation of mycorrhiza and PGPR against root-knot nematode. Symbiosis 71:175–183
Shrivastava P, Kumar R (2015) Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J Biol Sci 22:123–131
Shukla PS, Agarwal PK, Jha B (2012) Improved salinity tolerance of Arachis hypogaea (L.) by the interaction of halotolerant plant-growth-promoting rhizobacteria. J Plant Growth Regul 31:195–206
Siddikee MA, Chauhan PS, Sa T (2012) Regulation of ethylene biosynthesis under salt stress in red pepper (Capsicum annuum L.) by 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase-producing halotolerant bacteria. J Plant Growth Regul 31:265–272
Siddikee MA, Glick BR, Chauhan PS, jong Yim W, Sa T (2011) Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing 1-aminocyclopropane-1-carboxylic acid deaminase activity. Plant Physiol Biochem 49:427–434
Singh JS (2015) Microbes: the chief ecological engineers in reinstating equilibrium in degraded ecosystems. Agric Ecosyst Environ 203:80–82
Singh JS, Strong PJ (2016) Biologically derived fertilizer: a multifaceted bio-tool in methane mitigation. Ecotoxicol Environ Saf 124:267–276
Singh KN. Chatrath R (2001) Salinity tolerance. Application of physiology in wheat breeding. CIMMYT. Mexico, DF p. 101-110
Singh RP, Jha P, Jha PN (2015) The plant-growth-promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J Plant Physiol 184:57–67
Singh RP, Jha PN (2016a) Alleviation of salinity-induced damage on wheat plant by an ACC deaminase-producing halophilic bacterium Serratia sp. SL-12 isolated from a salt lake. Symbiosis 69:101–111
Singh RP, Jha PN (2016b) The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLos one 11:e0155026
Singh RP, Jha P, Jha PN (2017) Bio-inoculation of plant growth-promoting rhizobacterium Enterobacter cloacae ZNP-3 increased resistance against salt and temperature stresses in wheat plant (Triticum aestivum L.). J Plant Growth Regul 36(3):783–798
Slama I, Messedi D, Ghnaya T, Savoure A, Abdelly C (2006) Effects of water deficit on growth and proline metabolism in Sesuvium portulacastrum. Environ Exp Bot 56:231–238
Stefan M, Mihasan M, Dunca S (2008) Plant growth promoting rhizobacteria can inhibit the in vitro germination of Glycine max L. seeds. Scientific annals of university “Alexandru Ioan Cuza” Iasi, section genetics and molecular biology. T IX 3:105–110
Stepien P, Klobus G (2005) Antioxidant defense in the leaves of C3 and C4 plants under salinity stress. Physiol Plant 125:31–40
Suarez C, Cardinale M, Ratering S, Steffens D, Jung S, Montoya AMZ, Schnell S (2015) Plant growth-promoting effects of Hartmannibacter diazotrophicus on summer barley (Hordeum vulgare L.) under salt stress. Appl Soil Ecol 95:23–30
Tester M, Davenport R (2003) Na+ tolerance and Na+ transport in higher plants. Ann Bot 91:503–527
Tuna AL, Kaya C, Dikilitas M, Higgs D (2008) The combined effects of gibberellic acid and salinity on some antioxidant enzyme activities, plant growth parameters and nutritional status in maize plants. Environ Exp Bot 62(1):1–9
Upadhyay SK, Singh DP (2015) Effect of salt-tolerant plant growth-promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol 17:288–293
Upadhyay SK, Singh DP, Saikia R (2009) Genetic diversity of plant growth promoting rhizobacteria isolated from rhizospheric soil of wheat under saline condition. Curr Microbiol 59:489–496
Upadhyay SK, Singh JS, Singh DP (2011) Exopolysaccharide-producing plant growth-promoting rhizobacteria under salinity condition. Pedosphere 21:214–222
Verma S, Mishra SN (2005) Putrescine alleviation of growth in salt stressed Brassica juncea by inducing antioxidative defense system. J Plant Physiol 162:669–677
Vimal SR, Patel VK, Singh JS (2018) Plant growth promoting Curtobacterium albidum strain SRV4: an agriculturally important microbe to alleviate salinity stress in paddy plants. Ecol Indic
Walker TS, Bais HP, Grotewold E, Vivanco J. M (2003) Root exudation and rhizosphere biology. Plant Physiol 132:44–51
Wang Q, Dodd IC, Belimov AA, Jiang F (2016) Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase growth and photosynthesis of pea plants under salt stress by limiting Na+ accumulation. Funct Plant Biol 431:161–172
Whipps JM (2001) Microbial interactions and biocontrol in the rhizosphere. J Exp Bot 52:487–511
Wu H, Zhu M, Shabala L, Zhou M, Shabala S (2015) K+ retention in leaf mesophyll, an overlooked component of salinity tolerance mechanism: a case study for barley. J Integr Plant Biol 57:171–185
Wu Z, Peng Y, Guo L, Li C (2014) Root colonization of encapsulated Klebsiella oxytoca Rs-5 on cotton plants and its promoting growth performance under salinity stress. Eur J Soil Biol 60:81–87
Yaish MW, Antony I, Glick BR (2015) Isolation and characterization of endophytic plant growth-promoting bacteria from date palm tree (Phoenix dactylifera L.) and their potential role in salinity tolerance. A Van Leeuw J Microb 107:1519–1532
Yamaguch S (2008) Gibberellin metabolism and its regulation. Annu Rev Plant Biol 59:225–251
Yang J, Kloepper JW, Ryu C. M (2009) Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci 14:1–4
Yao L, Wu Z, Zheng Y, Kaleem I, Li C (2010) Growth promotion and protection against salt stress by Pseudomonas putida Rs-198 on cotton. Eur J Soil Biol 46:49–54
Zapata PJ, Botella MÁ, Pretel MT, Serrano M (2007) Responses of ethylene biosynthesis to saline stress in seedlings of eight plant species. Plant Growth Regul 53:97–106
Zerrouk IZ, Benchabane M, Khelifi L, Yokawa K, Ludwig-Müller J, Baluska F (2016) A Pseudomonas strain isolated from date-palm rhizospheres improves root growth and promotes root formation in maize exposed to salt and aluminum stress. J Plant Physiol 191:111–119
Zhang H, Kim MS, Sun Y, Dowd SE, Shi H, Paré PW (2008) Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol Plant-Microbe Interact 21:737–744
Zhang M, Qin Z, Liu X (2005) Remote sensed spectral imagery to detect late blight in field tomatoes. Precis Agric 6:489–508
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Bhise, K.K., Dandge, P.B. Mitigation of salinity stress in plants using plant growth promoting bacteria. Symbiosis 79, 191–204 (2019). https://doi.org/10.1007/s13199-019-00638-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13199-019-00638-y