
Abstract
Ecologic patterns and community succession are generally controlled by hydrologic mechanisms, especially for plant distributions which are sensitive to habitat conditions. The hydrology characteristics of ecosystems mainly influenced plant ecological processes in water and salinity changes. In this paper, we analyzed the composition and characteristic of natural plant community, divided the plant community classes and discussed the effect of water and salinity gradients on plant species and community classes in Ejina Desert Oasis. The results demonstrated that Populus euphratica, Tamarix chinensis and Phragmites communis were the most important plant species that had the highest important values among forest, shrubs and herbaceous. Six plant community patterns were classified by cluster analysis in Ejina Desert Oasis. Species richness and species diversity were the highest near West River and East River channels of the core oasis area. The distributions of plant community were mainly influenced by the following factors: distance from river channel, groundwater level, soil water content, soil salinity and groundwater salinity. The water and salinity factors, which controlled the distributions of plant, were the main driving forces for ecosystem succession. The plant community succession is becoming toward the type of shrub + herb or low shrub with very drought-tolerant from the type of forest + shrub + herb with tall and high water consumption, when habitat conditions change from good to poor. The water gradients had more significant and more directed effect than salinity gradients on plant species and communities.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Desert oasis is an important ecosystem in arid and semi-arid areas, especially for desert riparian forest which dominates the riparian ecosystems (Yang et al. 2009). Desert riparian forest is quite important for us to maintain the biodiversity of local ecosystem and also is the core component of arid inland basin ecosystems where crucial energy and matter cycles happen at all time (Li et al. 2013). The desert riparian forest provides important habitats for many plants and animals in arid or semi-arid region.
Water is a key factor for restoration of ecosystems in arid regions (Zhou et al. 2010). Changes of hydrological processes could cause the alterations of vegetation pattern (Zhao et al. 2004; Hao et al. 2013). The key components of hydrological process include rainfall (Devoto et al. 2009; Maharjan et al. 2011), surface water (Zenner et al. 2012; Sun et al. 2012), groundwater (Chen et al. 2006; Fan et al. 2011) and soil water content (Laudicina et al. 2012; Kammer et al. 2013), and any changes of them would change current water gradients of habitat and further have significant impact on plant communities. These effects are mainly manifested in the following aspects, such as changes of plant species diversity (Kammer et al. 2013), plant species distribution (Nucci et al. 2012), plant community (Zhang et al. 2013) and community composition (Walbridge 1994). Above research results about the relationships between plant communities and water gradients would help to understand the present habitat conditions and identify future changes (McIlroy and Allen-Diaz 2012).
Generally, salinity shouldn’t be neglected as a factor affecting plant community, especially in the ecology of deserts and wetlands where its importance is obviously given the prevalence of halophytes (Bui 2013). Soil salinity reduces primary production of some natural grass ecosystems and affects allocation of resources to plant reproductive structures (Onkware 2000). The magnitude and rate of plant community response to salinity fluctuations also depended on the structure and composition of the existing plant community (Wetzel et al. 2004). Several studies have been carried out on the effect of salinity on plant species (González-Alcaraz et al. 2014), and the results showed that salinity significantly affected the spatial distribution of plant species in riparian area and desert region. Meanwhile, the study about the relationships between soil salinity and community structure has been carried out (Cardinale 2011), the results revealed that each species had very specific relations to different environmental variables, and these relations were reflected in habitat status, ecological adaptations and stress tolerance degree of individual species (Naz et al. 2010). Groundwater salinity was also identified as an important factor affecting plant species composition (Guha and Panday 2012). On the whole, it can be concluded that salinity influences the distribution of species by changing their habitats.
The environment has changed remarkably in recent decades with cease to flow in the Heihe River, drying-up of lakes, groundwater level declines, vegetation degeneration, atrophy of natural oasis, desertification and frequent sand storms in Ejina desert oasis of northwestern China under the conditions of global climate change and human activities intensified (Zhao et al. 2004). Significant changes have taken place in salinity concentration and availability water resource in Ejina Desert Oasis. The availability water resource and salinity are the key factors that affect plant growth and community succession at arid regions (Chen et al. 2006; Zhang et al. 2013). The ecosystem of Ejina Desert Oasis that has been severely vulnerable will face enormous threatens (Fu et al. 2013). A number of studies reported the impacts of water environmental factors on the ecosystem in Ejina Desert Oasis (Wang et al. 2011). However, the study about plant community composition and succession under water and salinity gradients is very little as we know in this study area. There is also little research on the combined effect of water and salinity on plant community.
The objectives of the current study are to: (1) analyze the major controlling factors on plant community succession; (2) improve understanding of plant community succession along water and salinity gradients in Ejina Desert Oasis; (3) speculate the succession stages in present and future so as to bring the different rehabilitation programs for damaged ecosystem, thus to reverse the succession of phenomena, to accelerate the succession process and to restore the ecosystem. The study is to analyze the composition and characteristic of natural plant community, divide the classes of plant community in study area and discuss the effects of water gradients and salinity gradients on plant species and community classes.
Study area
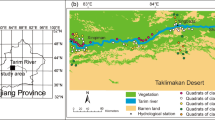
The study area is located in the lower reaches of the Heihe River Basin, northwest of China (Fig. 1), ranges from the Langxin Mountain to the East Juyan Lake and West Juyan Lake (40°00′–42°30′N; 99°30′–103°00′E). The study area belongs to the western Alashan highland, Inner Mongolia, which is made up of a series of mid- or low denudation mountains, proluvial fan dry delta and basin. Its altitude above sea level ranges from 890 m to 1200 m. The site of the lowest altitude is located in East Juyan lake of the study area northeastern. The highest site is on the top of Langxinshan mountain that locates in the southern part of the study area. The streambed is wide and shallow with low and ephemeral runoff. Typical desert riparian forest, shrubbery and pratoherbosa meadow vegetation grow along the Heihe River, and the vegetation patterns mainly include Populus euphratica forest, P. euphratica–Tamarix chinensis mixed forest and T. chinensis forest.

Location map of the study area showing the sampling locations
The region is representative of extreme drought sub-region with sparse precipitation and intense evaporation. It is dry and cold in winter and spring, but becomes extremely hot in summer and autumn. The region has long sunshine duration and frequent sand storms. According to observational data of Ejina weather station, the annual average precipitation is approximately 34 mm (1960–2012), with the maximum and minimum of 101 and 7 mm, respectively. The precipitation mostly occurs during June to September. The average annual pan evaporation is about 3218 mm (1960–2012).
Materials and methods
Collection and analysis of samples
The sampling plots were surveyed from July 5 to July 23, 2011, in Ejina Oasis, the lower reaches of the Heihe River Basin. In total, 309 quadrats were sampled in 61 sampling plots (24 forest sample plots, 26 shrub sample plots and 11 herbaceous sample plots) (Figs. 1, 2). It should be noted that the area of each forest sampling plot was 100 m × 100 m where five 20 m × 20 m quadrats were selected along diagonal of the sampling plot. The area of shrubs sample plot is 50 m × 50 m where five 10 m × 10 m quadrats were sampled same as forest quadrats. The area of herbaceous quadrats is 1 m × 1 m, and total five quadrats were done randomly in each herbaceous sample plot. The details of the sampling methods can be found in Feng et al. (2013).

a Desert riparian forest (P. euphratica + T. chinensis + S. alopecuroides); b desert riparian forest (P. euphratica + P. communis); c desert riparian shrubbery (T. chinensis); d desert riparian meadow (S. alopecuroides); e wet grassland (P. communis); f N. tangutorum shrub island
The vegetation community was divided into forest layer, shrub layer and herbaceous layer so as to facilitate measurements and analysis. For forest layer, the trees with diameter at breast height (DBH) larger than 5 cm were investigated in sampling quadrats, while the shrubs whose basal diameter (BD) is less than 5 cm were sampled in shrub sampling quadrats.
The parameters of diameter at breast height, height and canopy area were investigated for forest layer. The cluster number, canopy area, basal diameter and height were investigated for shrub layer. For herbaceous vegetation, cluster number, average coverage and height were surveyed. Other data including longitude, latitude, elevation, aspect, slope position, gradient, thickness of soil layer, human disturbance conditions were also investigated in each quadrats.
Soil physical and chemical properties were obtained through soil survey. Soil section was divided into eight layers with 20-cm intervals from 0 to 160 cm. Soil samples were sampled from each layer for each sample plot. Laboratory analysis methods were used to obtain soil salinity. The details were shown as follows: Milli-Q water was used for cleaning containers and sample-processing equipment. Samples were diluted in closed polyethylene containers and analyzed for cations (Na+, K+, Ca2+ and Mg2+) and anions (Cl−, NO3 − and SO4 2−) with a Dionex ICS-5000 ion chromatograph. The analyzing columns used for the cations and anions were AS11-HC and CS12A, and protecting columns were AG11-HC and CG12A, respectively. The injected volume was 25 μL according to the concentration of sample. Eluents used for cations and anions were methanesulfonic acid and sodium hydroxide with concentrations of 20 and 30 mM, respectively. Blanks were regularly monitored during the sample analysis, and all blanks were found to be lower than detection limits. Analytical imprecision was calculated (by replicate measurements of standards) to be within 10 % of the average concentration levels found in the samples. Soil hydraulic properties such as field capacity and saturated water content were measured by oven drying method. Groundwater depths were observed through manual measuring and automatic recording by Hobo U20.
Data analysis methods
In this study, important value index was used, and it can be defined as (Giliba et al. 2011):
where Relative dominance (or Relative basal cover) is calculated following as the diameter at breast height (DBH) for tree, the basal diameter (BD) for shrub and herbaceous.
Species richness and diversity
Four kinds of species diversity index were selected, according to the application extent and the reflection ability of species diversity index (Simpson 1949):
where N is total number of individuals observed; S is the species in the community or samples; P i is the ratio of the ith kind number and the total number of samples (in accordance with abundance), or the relative important values for the ith (according to the important value).
Plant community composition and gradients analysis
Species cluster analysis method was used to classify the plant community of study area based on the important values of species for all sample plots (Sun et al. 2011) by PC-ORD 4.0. Detrended canonical correspondence analysis (DCCA) and partial detrended canonical correspondence analysis (partial DCCA) were chosen to study the relationships between the community composition and water and salinity gradient factors by the ordination program CANOCO 4.5 Software (Centre for Biometry, Wageningen, Netherland, Braak and Šmilauer 2002). Monte Carlo test with 1000 permutations was employed to determine the significance of the canonical axes (Lep and Smilauer 2003). The analysis method of spatial interpolation was also performed on groundwater level data obtained from Xi et al. (2010a) with the ArcMap 10.
Results
Analysis of plant community composition
Plant composition of Ejina Desert Oasis
The plant composition of Ejina Desert Oasis was classified as forest, shrub and herbage (Table 1). Thirty-seven species were identified in the samples, including 2 forest species, 9 shrub species and 26 herbaceous species. They belong Asclepiadaceae (1 species), Chenopodiaceae (9 species), Compositae (6 species), Elaeagnaceae (1 species), Ephedraceae (1 species), Gramineae (5 species), Leguminosae (5 species), Polygonaceae (1 species), Salicaceae (1 species), Solanaceae (1 species), Tamaricaceae (2 species) and Zygophyllaceae (4 species). Eight herbaceous species appeared only once when we investigated, so they were eliminated from all species in this study. The results showed that the relative abundance, relative frequency and relative basal cover of P. euphratica were far larger than the Elaeagnus angustifolia in tree layer, and the important value of P. euphratica was 294.718 %. In shrub layer, the important value of T. chinensis was 157.427 %, ranked No. 1 in all shrub species, and its relative abundance, frequency and basal cover were also the largest; the important values of Nitraria tangutorum and Lycium ruthenicum ranked second and third, respectively. The species whose important values ranked top three were Phragmites communis, Sophora alopecuroides and Glycyrrhiza uralensis in herbaceous layer. Although the relative frequency of Peganum harmala ranked third, its relative abundance and relative basal cover ranked ninth, so that the important value of P. harmala occupied the fifth.
The main classification of plant community
Six plant community classes were classified by cluster analysis in Ejina Desert Oasis (Fig. 3; Table 2): Class I: P. euphratica + T. chinensis + S. alopecuroides, G. uralensis, P. harmala (P. communis or Achnatherum splendens). It was shown that P. euphratica (main storey) and T. chinensis (understory shrubs) were the constructive species of the plant community, with the addition of herbaceous layer. Class I included six subclasses with 19 sample plots (99 quadrats) (Table 2) that were mainly distributed in the core oasis area near river channel (Fig. 3). The results of species diversity index for Class I showed that the averages of Shannon–Wiener, Margalef’s and Simpson’s diversity (1-Dominance) were all the largest in all of plant community classes; they were about 0.522 ± 0.301, 0.279 ± 0.148 and 0.300 ± 0.169, respectively. The average of Simpson’s Diversity (1/Dominance) was the second largest in all classes with 1.638 ± 0.473. The Evenness of Class I was 0.679 ± 0.205, the Richness’s was 2.689 ± 1.129, ranked fourth and first respectively in all classes (Fig. 4).

Spatial distribution of sample plots for different plant community classes in Ejina Desert Oasis

Plant community characteristic of different classification (a species diversity index—Shannon–Wiener, b species diversity index—Margalef’s, c species diversity index—Simpson’s diversity (1/Dominance), d species diversity index—Simpson’s diversity (1-dominance), e species evenness, f species richness)
Class II: P. euphratica + S. alopecuroides or P. euphratica as constructive species with the addition of herbaceous layer. It included two subclasses with 6 sample plots (30 quadrats) (Table 2). These sample plots were mainly distributed in the core oasis area near East river (Fig. 3). The average values of Shannon–Wiener, Margalef’s, Simpson’s Diversity (1/Dominance) and Simpson’s Diversity (1-Dominance) for Class II were 0.216 ± 0.125, 0.137 ± 0.080, 1.227 ± 0.122 and 0.135 ± 0.076 respectively, ranked fifth, fourth, fifth and fifth, respectively. The Evenness of Class II was 0.631 ± 0.245, the Richness’s was 2.133 ± 0.873, ranked fifth and fourth in all classes (Fig. 4).
Class III was N. tangutorum, Haloxylon ammodendron, L. ruthenicum (Karelinia caspica) + S. alopecuroides, A. cristatum, Kalidium foliatum, P. harmala. There were in total four subclasses with eight sample plots (40 quadrats) (Table 2). These sample plots were mainly distributed in the lower reaches of East river and West river, as well as Gurinai and Guaizi lakes (Figs. 1, 3). The average values of Shannon–Wiener, Simpson’s Diversity (1/Dominance) and Simpson’s Diversity (1-Dominance) for Class III were 0.263 ± 0.319, 1.307 ± 0.371 and 0.157 ± 0.188 respectively, all of them ranked the fourth largest in all classes. The average of Margalef’s was 0.102 ± 0.128 and ranked fifth in all classes. The Evenness of species was 0.840 ± 0.188, ranked second in all classes. The Richness was 1.75 ± 0.906, ranked fifth (Fig. 4).
Class IV included Calligonum mongolicum, H. ammodendron (Reaumuria soongorica, N. tangutorum) + Zygophyllum fabago. It was shown that shrubs + herbaceous (Z. fabago) was the main pattern in the plant communities. It included four subclasses with five sample plots (25 quadrats) (Table 2). These sample plots were mainly distributed around of Gurinai (Fig. 3). The four kinds of species diversity index for Class IV were all ranked third in six classes. The Evenness of Class IV was 0.546 ± 0.191 that was in last place in six classes (Fig. 4).
Class V was (N. tangutorum, R. soongorica, H. ammodendron, C. mongolicum) + P. communis (Z. fabago). It included four subclasses with nine sample plots (45 quadrats) (Table 2). They were mainly scattered around Gurinai and Guaizi Lake (Fig. 3). The four kinds of species diversity index for Class IV were all the smallest in six classes except Evenness. The Evenness of Class IV was 0.924 ± 0.161 that was the largest in all classes’ (Fig. 4).
Class VI was consisted of (E. angustifolia) + T. chinensis (L. ruthenicum, S. xanthoxylum, R. soongorica, N. tangutorum) + A. sparsifolia, S. alopecuroides, K. caspica (A. splendens, P. harmala, P. communis). It was shown that T. chinensis (understory shrubs) was the constructive species in the plant community with the addition of herbaceous layer. It included eight subclasses with 14 sample plots (70 quadrats) that were mainly distributed in the core oasis area near river channel, East Juyan Lake and Swan Lake (Table 2; Fig. 3). The species diversity index of Class VI showed that the averages of Shannon–Wiener, Margalef’s, and Simpson’s Diversity (1-Dominance) were all the second largest in all of classes’, were about 0.501 ± 0.448, 0.229 ± 0.215 and 0.275 ± 0.238 respectively. The average of Simpson’s Diversity (1/Dominance) was the largest in all classes, with 1.639 ± 0.666. The Evenness of Class VI was 0.733 ± 0.214, and the Richness of Class VI was 2.700 ± 1.588; the Evenness and Richness of Class VI ranked separately third and first in all classes’ (Fig. 4).
These results indicated that the species of plant communities were rich relatively for Class I and Class VI; especially for Class I, the structure of plant community was made up of tree + shrub + herbaceous and its constructive species were P. euphratica and T. chinensis. These plant communities distributed near river where the water conditions were very suitable for plant growth. The Evenness of plant communities of Class V and Class III was higher than other classes, this is because there are less species even single species that grew in those plots under the conditions of either poorer water or higher salinity conditions.
The relationship between water gradients and species
In order to quantitatively research the relationships of water gradients and plant species in Ejina Desert Oasis, DCCA method was performed in this study. Distance from the river channel (DistR), groundwater level (GWL) and soil water content (SWC) were chosen as the water gradient factors. As shown in Fig. 5, hollow triangles represent species, and red arrow lines represent water gradient factors. The angles between the arrow lines could be used to calculate the correlations between the water gradient factors. The correlation coefficient is equal to the cosine value of the angles. The distance between the hollow triangles approximates the Chi-square distance between the species distributions in the biplot scaling, and the larger the distance was between hollow triangles, the more different the species distribution was (Jan leps and Petr Smilauer 2003). Through projecting the species onto the arrow lines, we can get approximate orders of those species’ optima relative to different water gradients.

Species–environment variables (water gradient) biplot ordination diagrams based on detrended canonical correspondence analysis (DCCA) (a) and partial DCCA (b salinity gradients including groundwater salinity and soil salinity variables as the covariables). Species are shown as triangles and labeled with their first three letters of the generic name and the first three letters of the specific name, and environmental variables are shown as arrows [where the DistR is the distance from the river channel to the sample plot (L, km), GWL is the groundwater level in sample plot (L, m), and SWC is the average soil water content of soil layers within 1 m in sample plot (%)]
The results of species–environment variables (i.e., water gradients) biplot ordination are shown in Fig. 5a, and the species–environment correlation was 0.721 in first canonical axis, Monte Carlo test of all canonical axes F-ratio = 2.011, p value = 0.004. The factor of distance from river channel was significantly negatively correlated with the factors of groundwater level depth as well as soil water content. The factor of GWL showed a positive correlation with the factor of SWC. The species of A. splendens, A. sparsifolia and S. alopecuroides had smaller difference of distribution relative to constructive species of P. euphratica and T. chinensis than the species of A. cristatum, P. communis, K. caspica, C. mongolicum, R. soongorica with the constructive species. The difference in distribution between T. chinensis and P. enphratical was smaller than the difference between other kinds of species in shrub layer with P. enphratical.
For species in herbaceous layer, there was smaller difference among A. splendens, S. alopecuroides and A. sparsifolia with P. enphratical than other herbaceous species with P. enphratical. It can be seen that P. enphratical was predicted to have its optimum with respect to environmental variable SWC at higher values than other species based on the results of relationship between species with environmental variables. The species of T. chinensis and P. enphratical were predicted to have their optima with respect to environmental variable GWL at higher values than other species. The species of N. tangutorum, E. angustifolia, G. uralensis and L. ruthenicum would reach their optima as the distance increases away from river channel (Fig. 5a).
In order to eliminate the influence of salinity gradient on species changes, partial DCCA was used to analyze the influence of water gradients on plant communities. The species–environment correlation was 0.691 in first canonical axis, Monte Carlo test of all canonical axes F-ratio = 1.907, p value = 0.01 (seeing Fig. 5b; Table 3). The results showed that the species of A. cristatum, A. splendens and S. alopecuroides were predicted to have their optimums than P. enphratical when the environmental variable SWC reached at higher values, and the species of P. enphratical, S. alopecuroides and T. chinensis were deemed to have optima with larger value of GWL. The species G. uralensis and N. tangutorum had the optima than other species’ with higher value of environmental variable DistR.
The distribution characteristic of plant species along salinity gradients
In order to quantify the influence of salinity gradients on the plant community, groundwater salinity (GW Salt) and soil salinity (Soil Salt) were chosen as the salinity gradient factors in Ejina Desert Oasis. The results of species–environment variables (salinity gradients) biplot ordination are shown in Fig. 6. The species–environment correlation coefficient was 0.612 in first canonical axis, Monte Carlo test of all canonical axes F-ratio = 1.815, p value = 0.004. The relation between two salinity gradient factors was not significant. The species of N. tangutorum was predicted to have its optimum than other species with respect to larger values of environmental variable Soil Salt. P. communis was predicted to have its optimum than other species relative to higher GW Salt (Fig. 6a). When eliminating the influence of water gradient factors on species, the species–environment correlation coefficient was 0.570 in first canonical axis, Monte Carlo test of all canonical axes F-ratio = 1.674, p value = 0.018 (see Fig. 6b; Table 3). The results showed that the species of N. tangutorum, G. uralensis and K. foliatum were predicted to have their optima than other species when environmental variable Soil Salt was at higher values, and the species of H. ammodendron and P. harmala were deemed to have optima than other species relative to higher GW Salt.

Species–salinity gradients biplot ordination diagrams based on DCCA (a) and partial DCCA (b water gradients included DistR, GWL and SWC as the covariables). Species are shown up as triangles and labeled with their first three letters of the generic name and the first three letters of the specific name, and environmental variables are shown as arrows (where the GW Salt is the groundwater salinity, and Soil Salt is the average salinity of soil layers within 1 m in sample plots)
Plant community characteristic under the condition combined water and salinity gradients
There are some overlaps among the spatial distribution of different classes (Fig. 7). However, the environmental variables varied remarkably in different classes. Class III, IV and V were mainly affected by the factor of DistR, Class VI was mainly influenced by GWL, SWC and Soil Salt, while Class I and II were affected by all of the water and salinity gradient factors.

Ordination of plant sampling plots of different classes and impact factors on first two axes of DCCA. Species are shown as solid triangles, sample plots are shown as solid circles, and environmental variables are shown as arrow lines. The samples are divided into six classes (same as Table 2)
Impact factors of the sample classes (test of significance of first canonical axis: F = 4.362, p = 0.002; test of significance of all canonical axes: F = 1.904, p = 0.002, Table 3) were represented by the correlations between the sample classes with Ax1 and Ax2 of DCCA (Fig. 7). The factor that showed a positive correlation with ordination axes was DistR, while the factors that showed negative correlation with ordination axes were GWL, SWC, Soil Salt and GW Salt. Therefore, the GWL, SWC, DistR and Soil Salt were the key factors which controlled and affected the distribution of sample classes.
The positions of samples and species points shown in Fig. 7 allowed us to estimate the relative abundances in the species points. When species are near the positions of samples, it means that the species could occur with the highest relative abundance in the samples, and similarly, if the sample positions are distributed near the positions of species, the species are tended to occur in those samples. It was shown (Fig. 7) that P. euphratica was most likely to occur in Class I and II, and in other words, it was predicted that the P. euphratica would have higher relative abundance in Class I and II than in other classes. The relative abundance of species of T. chinensis was higher near Class I and VI. Similarly, the species of H. ammodendron, R. soongorica, P. communis and K. foliatum were most likely appeared in samples of Class V.
Discussion
Effect of water gradients on plant community
Water is one of the important factors that influence the plant community and ecosystems at arid regions, and the changes of hydrological system strongly affect the structure, dynamics and composition of plant community (Zhou et al. 2010). The water gradients that mainly include precipitation (Brenes-Arguedas et al. 2009), surface runoff (Carling et al. 2013) and groundwater (saturated and unsaturated) (Närhi et al. 2010) could influence the growth and distribution patterns of plant species. Due to the difference and limitation of surface runoff in spatial and temporal distribution, in most cases the plants mainly grow by using precipitation and groundwater in arid region (Lamontagne et al. 2005). The mean annual precipitation is 42 mm, and event precipitation ≥10 mm is rare in Ejina Desert Oasis. But the mean annual potential evapotranspiration is about 3755 mm (Feng et al. 2004; Si et al. 2007). The scarce precipitation for plant growth and development in extremely arid region has no practical significant (Chen et al. 2003). Thus, that was the reason why the groundwater level depth, soil water content and distance from river channel were selected as water gradients to analyze the effect on plant community in this research. The plant species changed from P. euphratica, T. chinesis, S. alopecuroides and A. sparsifolia, to N. tangutorum, L. ruthenicum, A. splendens and G. uralensis to H. ammodendron, R. soongorica, P. communis and K. foliatum as the increase of distance away from river channel (Fig. 5a). The classes of plant communities changed from Class VI, II, I to Class III, IV, V along the arrow direction of DistR. Conversely, the classes of plant communities changed from Class III, IV, V to Class VI, II, I, along the arrow directions of groundwater level and soil water content (Fig. 8a). When the effect of salinity gradients was excluded, the plant species changed from T. chinensis, S. alopecuroides and P. euphratica to S. xanthoxylum, N. tangutorum, L. ruthenicum and Z. fabago to H. ammodendron, K. foliatum and P. communis along the arrow direction of DistR. But the plant species would change from S. xanthoxylum, C. mongolicum and Z. fabago to N. tangutorum, L. ruthenicum and K. caspica to S. alopecuroides, P. euphratica and P. harmala (Fig. 5b), and the plant communities transform from Class IV, III to Class II, I, VI, V along soil water content gradient (Fig. 8b). Thus, it can be seen that the water gradients significantly impact the distribution of species and plant communities, and the trend of plant community succession is from the type of forest + shrub + herb that is tall plants and high water consumption to the type of shrub + herb or low shrub that is very drought-tolerant as the water conditions changing from good to poor. Other studies also found similar results. For example, McIlroy and Allen-Diaz (2012) recorded that the plant communities showed strong connections with water table variables, and they found significant differences among community types for different water table variables. The groundwater level was apparently one of the major attributes to the plant species diversity in arid region (Chen et al. 2006).

Samples–water gradient variables biplot diagrams from DCCA (a) and partial DCCA (b other salinity variables as the covariables) summarizing differences in samples classes along the water gradient (the classes same as Table 2)
Effect of salinity gradients on plant community
The salinity factors (soil salinity and groundwater salinity) were also important factors for plant growth and distribution except the water conditions. The salinity would affect the niche of plant by changing the physical and chemical properties of soil. With the wide application of Niche Theory among species relations, biodiversity, community structure, succession and population evolution, it became one of the very active fields in ecology (Lawton 1985; Cardinale 2011). Due to the little precipitation and strong evaporation of groundwater, the soil salinity and groundwater salinity continuously increased in the Ejina Desert Oasis. An excess of salinity could inhibit the growth of plants and threaten the survival of the vegetation (Kang et al. 2004). Therefore, understanding the relationship between salinity changes with the community of desert riparian forest and population, identifying species response mechanism for salinity changes are essential for protecting the ecological environment in arid areas.
The effect of groundwater salinity on plant communities was different from soil salinity. With increasing groundwater salinity, the plant community classes changed from Class IV, VI, II, I to Class III, V, but along the direction of increasing soil salinity, the plant community classes didn’t show an obvious change trend (Fig. 9a). When only considering the effect of salinity gradients on plant community (eliminating the water gradients effective), the trend of plant community succession was from Class IV, II, VI, I to Class V, III along the arrow direction of groundwater salinity (Fig. 9b). This demonstrated that the effect of groundwater salinity on plant community classes was more evident than the soil salinity. This is mainly because the majority of the desert oasis plants have deeper roots enabling them access to the groundwater table. Hence the effect of groundwater salinity on the plant communities became more significant than the soil salinity’s sampled within 1 m depth. It can be seen in Fig. 9 that Class V of plant community has the greatest tolerance in the gradient of groundwater salinity. Class VI has maximum tolerance in the gradient of soil salinity. Class IV of plant community has minimal tolerance for groundwater salinity and soil salinity. The main reason is that Class VI and Class V have higher biodiversity than Class IV (Fig. 4). This is consistent with the results (Khan et al. 1976) about salt tolerance of species in the similar arid area.

Samples–salinity gradient variables biplot diagrams from DCCA (a) and partial DCCA (b other water variables as the covariables) summarizing differences in samples classes along the water gradient (the classes same as Table 2)
Effect combined water and salinity on plant community
In general, with the runoff reducing, groundwater level declining and salinity gradually increasing in soil root zone, a series of plant community succession would happen, especially in arid regions. The hydrology characteristics of ecosystems mainly show the effect of water and salinity on plant ecological processes simultaneously. The hydrologic mechanisms control the most basic ecologic patterns and processes (Rodriguez-Iturbe 2000), especially for basic pattern of plant distribution. They are the main driving forces of succession of ecosystem. The adjustment of hydrological process is a good way to control the vegetation changes. For example, hydrological processes can be used to adjust and deploy the “flow” within the landscape (including nutrients, pollutions, minerals and organic matter). The deterioration of water quality, the change of water level (especially shallow groundwater), water chemical characteristics and their changes would impact the plant community structure, dynamic, distribution and succession. Glaser et al. (1990) researched the response of vegetation to chemical and hydrological gradients, the results showed that the vascular plant and bryophyte data indicated a close relationship between the vegetation and both moisture and chemical gradients. Baldwin and Mendelssohn (1998) analyzed the effects of salinity and water level on coastal marshes and found that salinity and water level had significant effects on species richness only following disturbance and that similar patterns occurred for aboveground biomass of dominant and subdominant species.
Ejina Desert Oasis is located in the lower reaches of Heihe River basin; in recent decades, water resources have been overexploited for agricultural irrigation and industrial production in the upper and middle reaches of the Heihe River, and the flow entering into the lower reaches has been continuously reduced since the 1990s. The average annual runoff was larger than 1 billion m3 before the 1990s, but the annual runoff fell sharply in recent years, which cause the river and lakes to dry up, and the groundwater level quickly declining (Xi et al. 2010b). The water and salinity environment has changed dramatically, which caused the vegetation degeneration (Xiaohui et al. 2007; Jiang and Liu 2010). This study concluded the processes of plant community succession in Ejina Desert Oasis (Fig. 10), when water and salinity changed in historical period (for example, river migration, groundwater level declining). Figure 10a uncovered the successions from forest of P. euphratica to shrubs of N. tangutorum due to the shift of river channel. Figure 10b revealed the successions from the high shrubs to low shrubs. From Fig. 10c, it can be obviously seen that the succession occurred from shrubs to herbaceous because of the groundwater level declining. To some extent, the changes of water gradients would have more significant and direct effect than salinity gradients on plant species and communities in this study area (Fig. 7).

Plant community succession in Ejina Desert Oasis
Conclusions
This study investigated thoroughly and systematically the distribution of natural vegetation as well as the water and salinity characteristic of natural vegetation habitat in Ejina Desert Oasis. The plant community classes were divided by cluster analysis method. For revealing the quantitative relationships between species and environmental variables, the gradient analysis methods were used to study the effects of water gradients and salinity gradients on plant species. The results indicated that the different species had their optima under different water and salinity characteristic in their habitat. Different plant communities’ succession would happen significantly along the directions of different water gradients and salinity gradients in spatial. In that case, we can speculate the trend of plant communities’ succession, while one or some environmental variables happen to change. This will help to recognize the present classes of plant communities and determine the directions of plant communities’ succession in future. This study can also provide important theoretical basis for ecosystem restoration and protection of Ejina Desert Oasis.
Although the relationships between plant species and water and salinity environment factors have been quantified, there are still other environmental factors that are not being considered in this study, such as topography, soil texture and soil nutrient, which also maybe impact on the plant communities’ characteristic and distribution. For providing technological support to make strategies of ecosystem restoration and protection, next, we hope to predict the possible plant communities’ succession in the future by continuous investigation and model simulation methods under different scenarios that are the changes of one or several environmental variables in Ejina Desert Oasis.
References
Baldwin AH, Mendelssohn IA (1998) Effects of salinity and water level on coastal marshes: an experimental test of disturbance as a catalyst for vegetation change. Aquat Bot 61:255–268. doi:10.1016/s0304-3770(98)00073-4
Brenes-Arguedas T, Coley PD, Kursar TA (2009) Pests vs. drought as determinants of plant distribution along a tropical rainfall gradient. Ecology 90:1751–1761. doi:10.1890/08-1271.1
Bui EN (2013) Soil salinity: a neglected factor in plant ecology and biogeography. J Arid Environ 92:14–25. doi:10.1016/j.jaridenv.2012.12.014
Cardinale BJ (2011) Biodiversity improves water quality through niche partitioning. Nature 472:86-U113. doi:10.1038/nature09904
Carling GT, Richards DC, Hoven H, Miller T, Fernandez DP, Rudd A, Pazmino E, Johnson WP (2013) Relationships of surface water, pore water, and sediment chemistry in wetlands adjacent to Great Salt Lake, Utah, and potential impacts on plant community health. Sci Total Environ 443:798–811. doi:10.1016/j.scitotenv.2012.11.063
Chen YN, Chen YP, Li WH, Zhang HF (2003) Response of the accumulation of proline in the bodies of Populus euphratica to the change of groundwater level at the lower reaches of Tarim River. Chin Sci Bull 48:1995–1999. doi:10.1360/02wd0569
Chen YN, Zilliacus H, Li WH, Zhang HF, Chen YP (2006) Ground-water level affects plant species diversity along the lower reaches of the Tarim river, Western China. J Arid Environ 66:231–246. doi:10.1016/j.jaridenv.2005.11.009
Devoto M, Medan D, Roig-Alsina A, Montaldo NH (2009) Patterns of species turnover in plant-pollinator communities along a precipitation gradient in Patagonia (Argentina). Austral Ecol 34:848–857. doi:10.1111/j.1442-9993.2009.01987.x
Fan XM, Pedroli B, Liu GH, Liu HG, Song CY, Shu LC (2011) Potential plant species distribution in the Yellow River Delta under the influence of groundwater level and soil salinity. Ecohydrology 4:744–756. doi:10.1002/eco.164
Feng Q, Liu W, Su YH, Zhang YW, Si JH (2004) Distribution and evolution of water chemistry in Heihe River basin. Environ Geol 45:947–956. doi:10.1007/s00254-003-0950-7
Feng Q, Su YH, Si JH, Chang ZQ, Xi HY, Guo R, Chen LJ, Huo H, Qin YY (2013) Ecohydrological transect survey of Heihe River Basin. Adv Earth Sci 28:187–196
Fu A, Chen Y, Li W (2013) Water use strategies of the desert riparian forest plant community in the lower reaches of Heihe River Basin, China. Sci China Earth Sci 57:1293–1305. doi:10.1007/s11430-013-4680-8
Giliba RA, Boon EK, Musamba EM, Kashindye AM, Shayo PF (2011) Composition, richness and diversity in Miombo. J Biodivers 2:1–7
Glaser PH, Janssens JA, Siegel DI (1990) The response of vegetation to chemical and hydrological gradients in the Lost River Peatland, Northern Minnesota. J Ecol 78:1021–1048. doi:10.2307/2260950
González-Alcaraz MN, Jiménez-Cárceles FJ, Álvarez Y, Álvarez-Rogel J (2014) Gradients of soil salinity and moisture, and plant distribution, in a Mediterranean semiarid saline watershed: a model of soil–plant relationships for contributing to the management. Catena 115:150–158. doi:10.1016/j.catena.2013.11.011
Guha H, Panday S (2012) Impact of sea level rise on groundwater salinity in a coastal community of South Florida. J Am Water Resour Assoc 48:510–529. doi:10.1111/j.1752-1688.2011.00630.x
Hao XM, Chen YN, Guo B, Ma JX (2013) Hydraulic redistribution of soil water in Populus euphratica Oliv. in a central Asian desert riparian forest. Ecohydrology 6:974–983. doi:10.1002/eco.1338
Jiang XH, Liu CM (2010) The influence of water regulation on vegetation in the lower Heihe River. J Geogr Sci 20:701–711. doi:10.1007/s11442-010-0805-6
Kammer PM, Schob C, Eberhard G, Gallina R, Meyer R, Tschanz C (2013) The relationship between soil water storage capacity and plant species diversity in high alpine vegetation. Plant Ecol Divers 6:457–466. doi:10.1080/17550874.2013.783142
Kang E, Li X, Zhang J, Xinglin H (2004) Water resources relating to desertification in the Hexi area of Gansu Province, China. J Glaciol Geocryol 26:657–667 (In English Abstract)
Khan MI, Khan MA, Khizar T (1976) Plant-growth regulators from species differing in salt tolerance as affected by soil-salinity. Plant Soil 45:267–271. doi:10.1007/bf00011149
Lamontagne S, Cook PG, O’Grady A, Eamus D (2005) Groundwater use by vegetation in a tropical savanna riparian zone (Daly River, Australia). J Hydrol 310:280–293. doi:10.1016/j.jhydrol.2005.01.009
Laudicina VA, De Pasquale C, Conte P, Badalucco L, Alonzo G, Palazzolo E (2012) Effects of afforestation with four unmixed plant species on the soil-water interactions in a semiarid Mediterranean region (Sicily, Italy). J Soils Sediments 12:1222–1230. doi:10.1007/s11368-012-0522-0
Lawton J (1985) Community structure and the niche—giller, Ps. Nature 314:39. doi:10.1038/314039a0
Leps J, Smilauer P (2003) Multivariate analysis of ecological data using CANOCO. United States of America by Cambridge University Press, New York
Li W, Zhou H, Fu A, Chen Y (2013) Ecological response and hydrological mechanism of desert riparian forest in inland river, northwest of China. Ecohydrology 6:949–955. doi:10.1002/eco.1385
Maharjan SK, Poorter L, Holmgren M, Bongers F, Wieringa JJ, Hawthorne WD (2011) Plant functional traits and the distribution of west African rain forest trees along the rainfall gradient. Biotropica 43:552–561. doi:10.1111/j.1744-7429.2010.00747.x
McIlroy SK, Allen-Diaz BH (2012) Plant community distribution along water table and grazing gradients in montane meadows of the Sierra Nevada Range (California, USA). Wetlands Ecol Manage 20:287–296. doi:10.1007/s11273-012-9253-7
Närhi P, Middleton M, Hyvönen E, Piekkari M, Sutinen R (2010) Central boreal mire plant communities along soil nutrient potential and water content gradients. Plant Soil 331:257–264. doi:10.1007/s11104-009-0250-4
Naz N, Hameed M, Ahmad MSA, Ashraf M, Arshad M (2010) Is soil salinity one of the major determinants of community structure under arid environments? Community Ecol 11:84–90. doi:10.1556/comec.11.2010.1.12
Nucci A, Angiolini C, Landi M, Bacchetta G (2012) Influence of bedrock-alluvial transition on plant species distribution along a Mediterranean river corridor. Plant Biosyst 146:564–575. doi:10.1080/11263504.2012.670669
Onkware AO (2000) Effect of soil salinity on plant distribution and production at Loburu Delta, Lake Bogoria National Reserve, Kenya. Austral Ecol 25:140–149. doi:10.1111/j.1442-9993.2000.tb00014.x
Rodriguez-Iturbe I (2000) Ecohydrology: a hydrologic perspective of climate-soil-vegetation dynamics. Water Resour Res 36:3–9. doi:10.1029/1999wr900210
Si JH, Feng Q, Zhang XY, Chang ZQ, Su YH, Xi HY (2007) Sap flow of Populus euphratica in a desert riparian forest in an extreme arid region during the growing season. J Integr Plant Biol 49:425–436. doi:10.1111/j.1672-9072.2006.00388.x
Simpson EH (1949) Measurement of diversity. Nature 163:688. doi:10.1038/163688a0
Sun Y, Li X, He Y, Jia Y, Ma Z, Guo W, Xin Z (2011) Impact factors on distribution and characteristics of natural plant community in reclamation zones of Changjiang River estuary. Chin Geogr Sci 22:154–166. doi:10.1007/s11769-011-0475-z
Sun R, Yuan XZ, Chen ZL, Zhang YW, Liu H (2012) Effect of three georges reservoir (Yangzi River) on the plant species richness in drawdown zone downstream the tributary river (Pengxihe River). Russ J Ecol 43:307–314. doi:10.1134/S1067413612040121
ter Braak CJF and Šmilauer P (eds) (2002) CANOCO Reference Manual and CanoDraw for Windows User's Guide: Software for Canonical Community Ordination (version 4.5). Microcomputer Power, Ithaca NY, USA, pp 500
Walbridge MR (1994) Plant community composition and surface-water chemistry of fen peatlands in West-Virginias appalachian plateau. Water Air Soil Poll 77:247–269. doi:10.1007/bf00478422
Wang YB, Feng Q, Si JH, Su YH, Chang ZQ, Xi HY (2011) The changes of vegetation cover in Ejina Oasis based on water resources redistribution in Heihe River. Environ Earth Sci 64:1965–1973. doi:10.1007/s12665-011-1013-0
Wetzel PR, Kitchens WM, Brush JM, Dusek ML (2004) Use a reciprocal transplant study to measure the rate of plant community change in a tidal marsh along a salinity gradient. Wetlands 24:879–890. doi:10.1672/0277-5212(2004)024[0879:uoarts]2.0.Co;2
Xi HY, Feng Q, Liu W, Si JH, Chang ZQ, Su YH (2010a) The research of groundwater flow model in Ejina Basin, Northwestern China. Environ Earth Sci 60:953–963. doi:10.1007/s12665-009-0231-1
Xi HY, Feng Q, Si JH, Chang ZQ, Cao SK (2010b) Impacts of river recharge on groundwater level and hydrochemistry in the lower reaches of Heihe River Watershed, northwestern China. Hydrogeol J 18:791–801. doi:10.1007/s10040-009-0562-8
Xiaohui J, Mingli S, Zhibing H, Zhen J (2007) Influence of water regulation on the eco-environment in the lower reaches of the Heihe River. In: Proceedings of the 3rd international yellow river forum on sustainable water resources management and delta ecosystem maintenance, vol 46–54
Yang YH, Chen YN, Li WH (2009) Relationship between soil properties and plant diversity in a desert riparian forest in the lower reaches of the Tarim River, Xinjiang, China. Arid Land Res Manag 23:283–296. doi:10.1080/15324980903231991
Zenner EK, Olszewski SL, Palik BJ, Kastendick DN, Peck JE, Blinn CR (2012) Riparian vegetation response to gradients in residual basal area with harvesting treatment and distance to stream. Forest Ecol Manag 283:66–76. doi:10.1016/j.foreco.2012.07.010
Zhang Z, Wan C, Zheng Z, Hu L, Feng K, Chang J, Xie P (2013) Plant community characteristics and their responses to environmental factors in the water level fluctuation zone of the three gorges reservoir in China. Environ Sci Pollut Res Int 20:7080–7091. doi:10.1007/s11356-013-1702-1
Zhao WZ, Chang XL, He ZB (2004) Responses of distribution pattern of desert riparian forests to hydrologic process in Ejina oasis. Sci China D 47:21–31. doi:10.1360/04zd0003
Zhou WL, Yang XQ, Hao P, Liu QW, Cao DC, Baribault T, Li JW (2010) Plant diversity and its maintenance in Populus euphratica riparian forests in the Ejina Oasis, China. For Stud China 12:55–61. doi:10.1007/s11632-010-0011-8
Acknowledgments
This research was supported by the National Natural Science Foundation of China (No. 41101026, 91025002), International Postdoctoral Exchange Fellowship Program (No. 201389), West Light Foundation of The Chinese Academy of Sciences (No. 29Y128881) and China Postdoctoral Science Foundation (No. 20110490863). We appreciate Dr. Xiaoyou Zhang, Dr. Yonghong Su, Mr. Yingsheng Guo and Mrs. Bing Jia for the assistance in field work, and anonymous reviewers for their comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Xi, H., Feng, Q., Zhang, L. et al. Effects of water and salinity on plant species composition and community succession in Ejina Desert Oasis, northwest China. Environ Earth Sci 75, 138 (2016). https://doi.org/10.1007/s12665-015-4823-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12665-015-4823-7