
Abstract
Staphylococcal wound infections range from mild to severe with life-threatening complications. The challenge of controlling such infections is related to bacterial biofilm formation, which is a major factor contributing to antibiotic resistance and infection recurrence. In this study, four clinical isolates of staphylococci species; two isolates of methicillin-resistant Staphylococcus aureus (MRSA) and two methicillin-sensitive Staphylococcus aureus (MSSA) isolates. The identification of bacterial species based on cell morphology, initial biochemical tests, and the VITEK2 system were used to confirm the clinical microbiological diagnosis. Antibiotic sensitivity testing showed that the isolated staphylococci were highly resistant to the following antibiotics, amoxicillin, penicillin G, cefotaxime, and methicillin. Combinations of cefotaxime with the cell-free supernatants (CFS) of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895, each one separately showed complementary activity against the tested staphylococci. The co-aggregation capability of the tested bacilli as beneficial bacteria against isolated staphylococci was also evaluated. The data showed a strong co-aggregation with scores (+ 3, + 4) which were reported between the bacilli strains and the isolated staphylococci. Furthermore, the CFS of bacilli strains showed an inhibitory effect against biofilm-associated MRSA and MSSA. These findings confirmed the ability of beneficial bacteria to compete with the pathogens at the site of colonization or for the source of nutrients and, eventually, lead to inhibition of the pathogens’ capability of causing a wound infection. Such beneficial bacteria could play an important role in future pharmaceutical and industrial applications.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wound infections come from three sources, (i) wound contamination by exogenous pathogens, predominantly, Staphylococcus aureus, (ii) the skin’s commensal (endogenous) microbiota, especially Staphylococcus epidermidis, and (iii) endogenous microbial sources such as from the intestinal or urogenital system [1]. In addition to wound infections, Staphylococcus aureus is associated with other health challenges, such as urinary tract infections and toxic shock syndrome [2]. S. aureus–associated wound infections range from mild skin abscesses to severe, life-threatening infections when extending to the surrounding bones and soft tissues [3]. Bacterial pathogenesis is linked to enzymes and toxins such as hemolysin, hyaluronidase, leucocidene, and DNase [3]. In addition, there is an evidence of staphylococcal infections in poultry [4], and with the growing interest in backyard poultry farming, there is a growing threat for the transfer of pathogenic staphylococci from birds to humans, similar to what is observed in the case of salmonella [5].
About 20–25% of staphylococci species are capable of persistent colonization on various surfaces and causing foreign body-related infections, which facilitates the initiation of biofilm formation [6]. Once an S. aureus biofilm is established, it exhibits high antibiotic resistance and transforms the infection from an acute condition to a chronic, and then recurrent infection [7].
The control of biofilm-associated infections is a challenging issue because of the antibiotic resistance and high rates of infection recurrence after treatment [8]. Therefore, an urgent effort is needed to find and develop alternative and effective antimicrobials against biofilm-associated antibiotic-resistant pathogens. In this regard, our study investigates the antimicrobial potential of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 against wound infection–associated staphylococci.
B. subtilis KATMIRA1933 was isolated from a milk product called YoguFarm™, which has been safely consumed by people for several years without any reported harmful side effects [9]. The strain’s capability of producing subtilosin A was reported in several studies [9,10,11]. B. amyloliquefaciens B-1895 is a soil isolate capable of producing numerous proteolytic enzymes and other antimicrobial substances that can inhibit the growth of certain foodborne and human pathogens [12, 13]. The strain has also been shown to have probiotic potential and antimicrobial properties in royal fish [14]. The B. amyloliquefaciens B-1895 solid-state fermented soybean acts as an immunomodulator substance and food additive for improving the muscle mass of poultry [15]. Both Bacillus strains and their metabolites have antioxidant and DNA protective activities [16] and act as probiotics in poultry [17, 18].
Co-aggregation is an interaction of microorganisms aimed at enhancement of their integration into a biofilm [19]. Aggregation and co-aggregation of probiotics play an important role in human health by preventing colonization of pathogens at the site of infection, and competing with them for the nutrients [20]. Auto-aggregation of beneficial bacteria is important for their adhesion onto the body surfaces and the skin epithelium [20]. Biofilm is a complex community of microorganisms embedded in a self-produced matrix, which composed of exopolymeric substance. The process of biofilm formation includes five steps. Briefly, when the correct conditions are provided, the planktonic cells swim toward and adhere to the conditioning surfaces to initiate primary biofilm. The adherent cells start to produce an extracellular polymeric substance (EPS) leading to microbial aggregation and matrix formation. Later, microcolonies and water channel are established while more layers are added to produce a fully mature biofilm with a maximum cell density. Then, microcolonies will be able to migrate and established a new biofilm mass in the other locations [21]. The extracellular matrix of fully mature biofilm is chemically composed of a mixture of proteins, lipids, extracellular DNA, and polysaccharides [22]. Therefore, effective anti-biofilm agents are required to interrupt and damage biofilm-associated pathogens.
The anti-biofilm activity of cell free supernatants (CFS) of the tested bacilli strains was previously evaluated against Proteus mirabilis isolated from humans and sheep having urinary tract infections [13]. In addition, subtilosin A produced by B. subtilis KATMIRA1933 showed anti-biofilm activity against Escherichia coli and Listeria monocytogenes [23]. The goal of this study was to explore whether B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 and their CFS possess anti-microbial and anti-biofilm activity against staphylococci isolated from wound infections. Co-aggregation ability and anti-biofilm activity of B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 against S. aureus may assist with decreasing the likelihood of recurring of staphylococcal infections.
Materials and Methods
Bacterial Growth Conditions
Clinical isolates of staphylococci (MRSA and MSSA) were a gift from the Educational Laboratories, Baqubah Teaching hospital in Diyala, Iraq. The bacterial isolates named as MRSA Th57, Th79 and MSSA Th43, Th85 were maintained on blood agar (HiMedia Laboratories Pvt. Ltd., Mumbai, India) and their ability to ferment mannitol was assessed using mannitol salt agar (HiMedia). The agar plates were incubated under aerobic condition at 37 °C for 24–48 h.
Tryptone Soya Broth (TSB) (Oxoid, ThermoScientific, Hampshire, UK) was used for supporting the growth of staphylococci species and when performing antimicrobial and biofilm inhibition assay. Bacterial isolates were stored in the refrigerator at 4 °C for a short period (7–10 days) on TSA (Oxoid) slants and periodically sub-cultured until they were used.
The sporulated stocks of B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 were maintained in De Man, Rogosa, and Sharpe (MRS) culture medium (Oxoid) and incubated aerobically for 24–36 h at 37 °C.
Bacterial Identification
The identification of staphylococci species was confirmed using VITEK ® 2 COMPACT system (BioMérieux, Marcy-l’Étoile, France). According to the manufacture, the Vitek 2 (30 card capacity) system is based on the fluorogenic methodology and biochemical reactions for organism identification using a 64-well card that is barcoded. The card type, expiration date, lot number, and unique card identification number are clearly mentioned in card. The test kits available for the microbial identification include gram-negative bacillus and gram-positive cocci. The Vitek 2 which is used for gram-positive cocci identifies 124 species of staphylococci, streptococci, enterococci, and a select group of gram-positive organisms within 8 h or less.
Antibiotic Susceptibility of Isolated Staphylococci
The antibiotic susceptibility of isolated staphylococci was evaluated using the Kirby-Bauer method and following current Clinical and Laboratory Standards Institute (CLSI) guidelines [24] with minor modifications. Briefly, 3–5 colonies grown on the TSA plates were transferred by sterile inoculating loop to a tube containing 5 mL of phosphate-buffered saline (PBS) (Bilaney Consultants Ltd., Sevenoaks, UK). Using a spectrophotometer (Molecular Diagnostics, Sunnyvale, CA, USA), the bacterial growth was diluted and adjusted to an optical density (OD600) of 0.1 which correlated with 106 CFU/mL. A 100-μL aliquot of the last bacterial dilution (106 CFU/mL) was inoculated by streaking onto Muller-Hinton (MHA) (Himedia) agar in three directions. The tested antibiotics were chosen based on the recommendation of a local physician, as commonly prescribed antimicrobials for staphylococcal wound infections. The tested antibiotic discs (Mast Group Ltd., Bootle, UK), including amoxicillin (10 μg/disk), penicillin (10 IU/disk), cefotaxime (30 μg/disk), and methicillin (5 μg/disk), were placed on the previously inoculated MHA with staphylococci species. The agar plates were incubated for 24 h at 37 °C. The radius of each zone of inhibition was measured in millimeters (mm). MRSA or MSSA were identified by measuring their sensitivity to the oxacillin (1 μg/disk). Bacterial sensitivity and/or resistance to the tested antibiotics were determined based on the standard chart approved by the CLSI [24] (Table 1).
Antibiotics in Combination with Bacillus CFS
The Kirby-Bauer method, CLSI [24] was applied to identify staphylococci sensitivity to antibiotics in combination with CFS of tested bacilli. The bacterial suspension in PBS was diluted and adjusted to 106 CFU/mL using a spectrophotometer (Molecular Diagnostics), to an OD600 of 0.1. Then, a swab saturated with the bacterial suspension was streaked over the MH agar plate in 3 directions. The entire surface of the dish was covered with the bacterial cells. Each antibiotic disk was saturated with 20 μL of CFS of the tested bacilli. Sterile forceps were used to pick up the antibiotic disks, and three different disks were prepared: Antibiotic disk only, antibiotic disk saturated with bacilli CFS, and a blank disk saturated with Bacillus CFS only. A blank disk was used as a control. All disks were placed on the surface of MH agar previously inoculated with the isolated pathogen. The antibiotic disks were placed 15 mm from the edge of the dish. The agar plates were left for 30 min, until the antibiotic was diffused from the disks into the surrounding agar surface, and then incubated aerobically at 37 °C for 16–18 h. After incubation, the bacterial growth prevention was evaluated by measurement of inhibition zones, and the results were represented as sensitive (S) or resistant (R) according to a standard chart for antibiotic susceptibility testing (Table 1).
Preparation of CFS of the Tested Bacilli
The CFS of bacilli strains was prepared according to Sutyak et al. [9]. The overnight grown B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 on MRS agar were inoculated into MRS broth and incubated under aerobic condition for 24 h at 37 °C. Bacterial cells were precipitated by centrifugation (5000g at 4 °C for 30 min). The CFS was sterilized by filtration using 0.45-μm polytetrafluoroethylene (PTFE) syringe filter, (Fisher Scientific, Ottawa, Ontario, Canada) and kept at 4 °C in the sterile tubes until use.
Antimicrobial Activity of the Bacilli CFS
This assay was performed following Algburi et al. [23] with some modifications. After the CFS of the bacilli was prepared, staphylococci were grown aerobically in TSB for 24 h at 37 °C, and their number was adjusted to an OD600 of 0.1 (106 CFU/mL). The bacterial cells were spread on TSA agar. By using a wide end of a 200-μL micropipette tip, wells were made in the TSA and 150 μL of CFS of B. subtilis KATMIRA1933, and B. amyloliquefaciens B-1895 was added separately into each well, in duplicate. The agar plates were incubated aerobically for 24–36 h at 37 °C, and the diameter of the inhibition zones was measured using a digital caliper.
Auto-aggregation and Co-aggregation Test
This assay was conducted to assess co-aggregation of B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 with the isolated staphylococci following the methods of Cisar et al. [25] with minor modifications. Briefly, bacterial cells were grown and harvested after aerobic incubation for 24 h at 37 °C by centrifugation (5000×g for 15 min at 25 °C). Then, the cells were washed twice with PBS. After the two wash steps, the cells were suspended in PBS again, and the optical density OD600 nm was adjusted to 0.25. Then, 3 mL of each staphylococci species was gently mixed with 3 mL of either Bacillus strain, while 6 mL of each washed bacterial cells suspension in single culture was used as controls. Test and control tubes were incubated at 30 °C, and 20-μL samples were taken from each sample after 2, 4, 24 h for gram staining to observe co-aggregation between bacteria using a microscope. The experiment was performed in triplicates.
Bacterial interaction was visualized after 2 h on the slide using a light microscope after cells had been stained using the gram-staining method. Pictures were taken with a microscope camera (Kopacam, Guangzhou Ostec Electronic Technology Co., Ltd., Guangzhou, China) using the × 100/1.0 oil objective. The images showing bacterial interaction were analyzed using the Kopacam, NIS- Elements D3.0 software. The quantity of co-aggregation was analyzed visually and scored using a scoring system following Algburi et al. [12], with a 0 when there is no co-aggregation and 4 for the highest amount of co-aggregation (Fig. 1). In regard to the calculation of aggregation, the scoring system was based on the strength of microscopic auto-aggregation and co-aggregation observed by the naked eyes.

Auto-aggregation and co-aggregation of the tested bacilli. a Auto-aggregation of B. subtilis KATMIRA1933, b auto-aggregation of B. amyloliquefaciens B-1895, c auto-aggregation of S. aureus, d co-aggregation of B. subtilis KATMIRA1933 with S. aureus, e co-aggregation of B. amyloliquefaciens B-1895 with S. aureus
Biofilm Inhibition by Bacilli CFS
The biofilm inhibition assay with bacilli CFS was performed following the method of Sutyak Noll et al. [10] with some modifications. Briefly, the CFS of B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 (100 μL) were twofold diluted with fresh TSB containing 1% glucose (TSBG1%) in a 96-well tissue culture plate (Greiner Bio-One GmbH, Kremsmünster, Austria). The overnight culture of staphylococci at 3 ± 2 × 109 CFU/mL was diluted and adjusted in TSBG1% to a final concentration of 5 × 106 CFU/mL using spectrophotometer at OD600 nm. An aliquot of 100 μL of bacterial culture (5 × 106 CFU/mL) was inoculated in the well contacting fixed amount of CFS. All plates were incubated aerobically at 37 °C for 24–36 h.
Determination of Biofilm Inhibition by Staining with Crystal Violate
Biofilms inhibition was quantified according to Algburi et al. [23], the non-adsorbed cells (planktonic cells) were aspirated with a micropipette from each well after a 24-h incubation, and the number of cells were adjusted using a spectrophotometer (OD600). Then, each well was gently washed 3 times with 200 μL of TSB. Heat fixation (at 60 °C for 1 h) was used for fixation of the biofilm mass which was then stained with crystal violet (CV) according to Borucki et al. [26]. In each well, 50 μL of 0.2% CV was added to the washed biofilm and kept at room temperature (24–28 °C) for 25 min. Each well was then washed three times with 200 μL of distilled water and left for 10 min to dry inside the incubator at 37 °C. For solubilization of the CV stain attached to the biofilm, 200 μL of ethanol 95% was added into each well. Absorbance was measured using a microplate spectrophotometer at 630 nm (Molecular Diagnostics).
Statistical Analysis
The antibiotic susceptibility test and antimicrobial potential assay of CFS of bacilli strains were performed at least twice in triplicate. Co-aggregation and auto-aggregation assays were performed three times, and their results were represented and scored using visual analysis. Student’s t test with two-tailed distribution (Sigmaplot 11.0, Systat Softwarem Inc., San Jose, CA, USA) was used in antibiotic sensitivity testing and to compare the planktonic growth (%) and biofilm formation (%) by the staphylococci species in the presence of CFS of bacilli.
Results
Antibiotic Susceptibility of Staphylococci Species
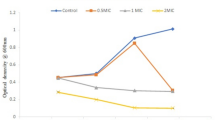
The antibiotic susceptibility of the isolated staphylococci, MRSA and MSSA, was investigated in this study. Our data illustrated that MRSA isolates were resistant to all tested antibiotics while MSSA isolates were resistant to amoxicillin, penicillin, and methicillin with one isolate was sensitive to cefotaxime. (Table 2).
Antibiotic Combination with CFS of the Tested Bacilli
Table 2 shows the susceptibility of the staphylococci isolates to antibiotics alone and in combination with the CFS of B. subtilis KATMIRA1933. The staphylococci strains, which were cefotaxime-resistant, showed sensitivity to cefotaxime in the presence of the CFS of B. subtilis KATMIRA1933. However, neither the CFS of B. subtilis KATMIRA1933 alone, nor cefotaxime individually had an antimicrobial effect on the growth of S. aureus (MRSA or MSSA). Complementary activity was observed when the tested bacilli CFS were added to the cefotaxime disks (Table 2).
In addition, the antimicrobial tolerance of the staphylococci isolates to antibiotics was evaluated when antibiotics were combined with the CFS of B. amyloliquefaciens B-1895. The cefotaxime-resistant isolates showed sensitivity to cefotaxime when it was combined with the CFS of B. amyloliquefaciens B-1895. In addition, the isolates of MRSA and MSSA showed resistance to amoxicillin when it was used alone, but only one isolate of MSSA became sensitive when amoxicillin was combined with the CFS of B. amyloliquefaciens B-1895. Neither the CFS of B. amyloliquefaciens B-1895 alone nor using cefotaxime or amoxicillin individually had an antimicrobial effect on the growth of S. aureus (MRSA or MSSA). Complementary activity was noticed when the antibiotics cefotaxime and amoxicillin were combined with the CFS of the tested bacilli (Table 2).
Auto-aggregation and Co-aggregation of Tested Bacilli
In our study, the auto-aggregation of the tested bacilli and their co-aggregation with the staphylococci isolates were evaluated by the gram-staining technique (Fig. 1). The results were expressed as (0) when no interaction was observed between the tested bacilli and the pathogens, and scored as + 1, + 2, + 3, or + 4 based on the strength of microbial interactions. Auto-aggregation was reported between the same species of the tested bacteria as follows: staphylococci (+ 2), B. subtilis KATMIRA1993 (+ 2), and B. amyloliquefaciens B-1895 (+ 3). Both B. subtilis KATMIRA1993 and B. amyloliquefaciens B-1895 strongly co-aggregated (+ 4 and + 3, respectively) with isolated S. aureus. In Fig. 1, the tested bacilli appeared as gram-positive bacilli aggregated with S. aureus, the gram-positive cocci.
Biofilm Inhibition by the CFS of the Studied Bacilli
The anti-bacterial activity of the CFS of the studied bacilli strains was determined against biofilm-associated S. aureus isolated from wound infection. Compared with the control, 25% and 50% of the CFS of B. subtilis KATMIRA1933 diluted in TSB inhibited 55% and 61%, respectively, of biofilm-associated MRSA. In addition, 57% and 60%, respectively, of MSSA biofilm was inhibited when the same concentrations of CFS (25 and 50%) were used. Regarding bacterial growth, MRSA viability were 95% and 60%, while MSSA viability were 96% and 56% when 25% and 50%, respectively, of CFS of B. subtilis KATMIRA 1933 were applied (Fig. 2). In a separate assay, the anti-biofilm effect of the CFS of B. amyloliquefaciens B-1895 was also assessed against isolated staphylococci. Our findings showed that MRSA biofilm inhibition, compared with the control group, was 48% and 55%, while MSSA biofilm prevention was 45% and 59% when 25% and 50% of the CFS of B. amyloliquefaciens B-1895 were used, respectively. In regard to cell viability, 25% and 50% of the CFS of B. amyloliquefaciens B-1895 caused planktonic growth inhibition (85%, 58%, respectively) of MRSA cells and (91%, 53%, respectively) of MSSA (Fig. 3). Our data showed no significance differences in the biofilm prevention activity of CFS of B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 against isolated MRSA or MSSA.

Anti-biofilm activity of CFS of B. subtilis KATMIRA1933 against isolated MRSA and MSSA. MRSA Biofilm formation % is represented by black bar while MSSA Biofilm formation % is represented by gray bar. MRSA planktonic growth % is represented by black circle (●), while MSSA planktonic growth % is represented by white circle (○)

Anti-biofilm activity of CFS of B. amyloliquefaciens B-1895 against isolated MRSA and MSSA. MRSA Biofilm formation % is represented by black bar while MSSA Biofilm formation % is represented by gray bar MRSA planktonic growth % is represented by black circle (●), while MSSA planktonic growth % is represented by white circle (○)
Discussion
Wound infection is predominantly caused by S. aureus, which is related to several infections [1]. S. aureus–associated wound infection is life threatening when extended to bone and soft tissues [3]. Bacterial pathogenesis is linked to their capability of producing enzymes and toxins in addition to biofilm formation [3]. The capability for β-lactamase production by S. aureus enables them to develop antibiotic resistance to β-lactam antibiotics [27].
In the current study, the sensitivity of staphylococci isolates was investigated for four commonly used antibiotics: amoxicillin, penicillin, cefotaxime, and methicillin. All isolated S. aureus were resistant to amoxicillin, penicillin, and only one isolate of MSSA was also sensitive to cefotaxime (Table 2). Our data were close to the observation of Naimi et al. [28] who reported that 99.05% of S. aureus isolates were resistant to penicillin G and 91.4% of them were multi-drug resistant. A high percentage of S. aureus resistance to penicillin, methicillin, and cephalexin (95.2, 86, and 84.6%) was also noticed by Chinnambedu et al. [29]. The authors referred to the progressive change in the antibiotic resistance profile of the isolated MRSA from 51.8% in 2012 to 86% in 2017.
Alternative strategies are urgently required for controlling the antibiotic resistance of bacterial pathogens. In this work, we evaluated antimicrobial combination of the CFS of B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 with the conventional antibiotics. Complementary activity was noticed when the tested bacilli CFS were combined with cefotaxime against isolated S. aureus. The CFS of various beneficial bacteria has been reported as having antibacterial activity against infections-associated pathogens, including Clostridium difficile, Listeria monocytogenes, Salmonella spp., and S. aureus [30, 31]. Furthermore, antibiotic activity in combination with the CFS of beneficial bacterial against pathogenic microorganisms has been shown in several reports [32, 33]. Aminnezhad et al. [32] noticed a significant antimicrobial activity of probiotic CFS on P. aeruginosa growth. Additionally, synergism was reported when the CFS from L. casei was combined with aminoglycoside antibiotics against P. aeruginosa. The CFS of Lactobacillus strains was reported to contain acetic acid, lactic acid, and hydrogen peroxide (H2O2) using (RP-HPLC) [32]. Compared with using an antibiotic or probiotic individually, the antimicrobial effect of tetracycline against P. aeruginosa was higher when it was combined with probiotic strains [33]. In the same study, probiotic L. plantarum, L. salivarius, and L. routeri demonstrated antimicrobial potential against P. aeruginosa growth that was greater than two commonly used antibiotics, tetracycline, and gentamicin. However, an antagonistic effect was reported for some tested antibiotics, indicating that not all antibiotics will have a synergistic activity with probiotics against bacterial pathogens [33]. Regarding S. aureus, the CFS of L. paraplantarum effectively inhibited bacterial growth [34]. The authors noticed that MRSA, which were resistant to ampicillin and oxacillin, showed a sensitivity to the same antibiotics when they were combined with the CFS of L. paraplantarum, indicating a synergistic activity of these combinations against MRSA strains. The antimicrobial activity of the tested Lactobacillus was related to the presence of organic acids (acetic acid and lactic acid) in CFS of L. paraplantarum which was confirmed by HPLC analysis. The findings of the abovementioned studies suggest that the CFS of beneficial microbes in combination with antibiotics may be viable alternative therapies to the use of conventional antibiotics alone for the treatment of multi-drug-resistant pathogens. We have designated the observed antimicrobial effect in this study as a complementary activity [35]. However, there are some studies reporting the results of antimicrobial disk impregnated CFS antibiotic combination studies as “synergistic” [36, 37]. Still, we are hesitant following these investigators without having the observed synergy confirmed using FICI (fractional inhibitory concentration indices) and/or isobolograms (see Algburi et al. [13] and Algburi et al. [38]). Further studies are required to investigate the nature of each antimicrobial interaction on pathogenic bacteria.
The capability of bacteria for auto-aggregation and co-aggregation with other microbes reflects the desirable properties of the beneficial bacteria. The probiotics interaction with pathogens can prevent their adhesion onto living tissues and help to eliminate them from the body. Moreover, self-aggregation of probiotics plays a role in their survival in a competitive environment.
The antimicrobial capability of probiotics against pathogens linked to aggregation has also been mentioned in some reports. Using atomic force microscopy, Younes et al. [39] noticed a strong adhesion force between the probiotic L. reuteri RC-14 and the virulent toxic shock syndrome toxin producing S. aureus. Significantly large bacterial co-aggregates were identified indicating the rapid anti-pathogenic effects of L. reuteri RC-14 against isolated S. aureus. The two lactobacilli, L. fermentum and L. pentosus isolated from milk showed maximum auto-aggregation and co-aggregation with S. aureus and E. coli [40]. The study of Soleimani et al. [41] reported a strong co-aggregation (88.4 and 76%) between L. plantarum and two S. aureus isolated from mastitis. The adhesive and anti-microbial attributes of probiotics, in addition to their capability of establishing a hostile micro-environment for pathogens, could be utilized in therapeutic formulations for controlling dangerous infection-associated pathogens.
Biofilm formation was previously reported as an important factor of bacterial pathogenesis and a major factor leading to therapeutic failure. Several studies have been conducted to determine the most effective strategies for controlling biofilm formation (see the review of Algburi et al. [8]). One of these methods is the use of probiotics and their metabolites as safe alternatives to traditional antibiotics. The tested probiotics in this study, B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895, were reported to have antimicrobial activity against a wide range of microbial pathogens due to the production of antimicrobial substances, most notably subtilosin and subtilin [12]. In a previous study, Algburi et al. [23], we evaluated the antimicrobial potential of subtilosin, a peptide produced by B. subtilis KATMIRA1933, as a quorum sensing (QS) inhibitor in gram-positive and gram-negative bacteria. The authors reported that subtilosin at sub-MIC concentrations reduced the level of autoinducer-2 (AI-2) produced by the tested gram-positive bacteria. The biofilm-prevention and anti-biofilm activities reported here for the CFS of the studied bacilli probiotics was accompanied by some reduction in the number of viable cells of the targeted pathogen. However, the observed reduction in numbers of staphylococci was less than one log, which is insufficient for the reduction in viable cells to be considered as playing any significant role in the observed effect on the biofilms. Recently, the anti-biofilm activity of the CFS of B. subtilis KATMIRA1933 and B. amyloliquefaciens B-1895 were evaluated against P. mirabilis isolated from humans and sheep having urinary tract infections [13]. The authors noticed that both tested bacilli were strongly co-aggregating with the P. mirabilis isolates. Microbial co-aggregation is an indication of the environmental competition between the tested bacilli and pathogenic bacteria and could possibly interfere with their adhesion to the eukaryotic host and prevent their biofilm formation.
The CFS of L. plantarum species isolated from Siahmazgi cheese, showed an anti-biofilm (reducing and disrupting) activity against multi-drug-resistant S. aureus, E. coli, and P. aeruginosa [42]. In the same report, the authors found that the anti-biofilm potential was stable at high temperatures (80 and 100 °C) and was not influenced by EDTA, SDS, or Tween 80. Biofilms of the pathogens P. aeruginosa and B. cereus were remarkably reduced in the presence of the CFS of the lactobacilli species L. pentosus and L. plantarum [43]. In addition, the anti-biofilm and anti-adhesion activity of cell-free biosurfactant (CFBS) produced by L. acidophilus NCIM 2903 was evaluated against several pathogens including E. coli NCIM 2065, S. aureus NCIM 2079, and P. vulgaris NCIM 2027 [44]. The authors found that CFBS, which was validated through a microfluidic assay, exhibited antimicrobial, anti-adhesive, and anti-biofilm effects against the tested pathogens. Furthermore, the CFS of L. plantarum isolated from fresh vegetables exhibited a strong inhibitory effect on the growth of S. aureus. Based on scanning electron microscopy data, the CFS of lactic acid bacteria are able to attack the bacterial membrane leading to a rough and wrinkled membrane that may lead, eventually, to the inhibition of biofilm formation by S. aureus [45]. Koohestani et al. [46] evaluated the anti-biofilm activity of L. acidophilus and L. casei on 2-day-old biofilm of food borne pathogen, S. aureus. The authors reported that long-acting CFS (for 28 days at 4 °C) showed a significant biofilm removal potential in a concentration-based manner. Fluorescence microscopy confirmed that the CFS of L. acidophilus had more anti-biofilm killing effect compared with L. casei CFS, making them suitable agents for controlling such food-borne pathogens. In a separate study, lactic acid bacteria and bifidobacteria showed an inhibitory effect on the growth of S. aureus, ranging from 0.5 to 34.2%. While a significant reduction in biofilm formation of S. aureus was also noticed when the CFS of Lactobacillus bulgaricus 8611 was used [47]. Based on the abovementioned data, Lactobacillus and spore-forming bacilli are recommended as safe, bio-controlling agents, which are capable of reducing and controlling biofilm formation, and as such can be considered for biomedical applications due to their capability of producing bacteriocins, lactic acid, acetic acid, hydrogen peroxide, and diacetyl.
Conclusion
Antibiotic resistance and infection recurrence are among the greatest treatment challenges for infection-associated biofilm formation. Alternative antimicrobials and therapeutic protocols are urgently required to control such serious infections. The combination of a probiotic and/or its metabolites together with antibiotics is an effective strategy for the inhibition of biofilm formation by wound infection associated S. aureus and for the improvement of antimicrobial activity of conventional antibiotics, especially when targeting MRSA.
References
Brackman G, De Meyer L, Nelis HJ, Coenye T (2013) Biofilm inhibitory and eradicating activity of wound care products against Staphylococcus aureus and Staphylococcus epidermidis biofilms in an in vitro chronic wound model. J Appl Microbiol 114(6):1833–1842. https://doi.org/10.1111/jam.12191
Miller LS, Cho JS (2011) Immunity against Staphylococcus aureus cutaneous infections. Nature Rev Immunol 11(8):505–518. https://doi.org/10.1038/nri3010
Lowy FD (1998) Staphylococcus aureus infections. N Engl J Med 339(8):520–532. https://doi.org/10.1056/NEJM199808203390806
Persoons D, Van Hoorebeke S, Hermans K, Butaye P, de Kruif A, Haesebrouck F, Dewulf J (2009) Methicillin-resistant Staphylococcus aureus in poultry. Emerg Infect Dis 15(3):452–453. https://doi.org/10.3201/eid1503.080696
Tobin MR, Goldshear JL, Price LB, Graham JP, Leibler JH (2015) A framework to reduce infectious disease risk from urban poultry in the United States. Public Health Rep 130(4):380–391. https://doi.org/10.1177/003335491513000417
Dall'Antonia M, Coen PG, Wilks M, Whiley A, Millar M (2005) Competition between methicillin-sensitive and -resistant Staphylococcus aureus in the anterior nares. J Hosp Infect 61:62–67. https://doi.org/10.1016/j.jhin.2005.01.008
Fitzpatrick F, Humphreys H, O'Gara JP (2005) The genetics of staphylococcal biofilm formation—will a greater understanding of pathogenesis lead to better management of device-related infection? Clin Microbiol Infect 11:967–973. https://doi.org/10.1111/j.1469-0691.2005.01274.x
Algburi A, Comito N, Kashtanov D, Dicks LM, Chikindas ML (2017a) Control of biofilm formation: antibiotics and beyond. Appl Environ Microbiol 83(3):e02508–e02516. https://doi.org/10.1128/AEM.02508-16
Sutyak KE, Wirawan R, Aroutcheva A, Chikindas M (2008) Isolation of the Bacillus subtilis antimicrobial peptide subtilosin from the dairy product-derived Bacillus amyloliquefaciens. J Appl Microbiol 104(4):1067–1074. https://doi.org/10.1111/j.1365-2672.2007.03626.x
Noll KS, Prichard MN, Khaykin A, Sinko PJ, Chikindas ML (2012) The natural antimicrobial peptide subtilosin acts synergistically with glycerol monolaurate, lauric arginate and ε-poly-L-lysine against bacterial vaginosis-associated pathogens but not human lactobacilli. Antimicrob agents Chemother 56(4):1756–1761. https://doi.org/10.1128/AAC.05861-11
Van Kuijk S, Noll KS, Chikindas ML (2012) The species-specific mode of action of the antimicrobial peptide subtilosin against Listeria monocytogenes Scott A. Lett App Microbiol 54(1):52–58. https://doi.org/10.1111/j.1472-765X.2011.03170.x
Algburi A, Volski A, Cugini C, Walsh EM, Chistyakov VA, Mazanko MS, Bren AB, Dicks LM, Chikindas ML (2016) Safety properties and probiotic potential of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895. Adv Microbiol 6(06):432–452. https://doi.org/10.4236/aim.2016.66043
Algburi A, Alazzawi SA, Al-Ezzy AIA, Weeks R, Chistyakov V, Chikindas ML (2020) Potential probiotics Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 co-aggregate with clinical isolates of Proteus mirabilis and prevent biofilm formation. Probiotics Antimicrob Prot. https://doi.org/10.1007/s12602-020-09631-0 [ahead of print]
Golovko GV, Zipelt LI, Karpenko GI, Chistyakov VA, Sazykina MA, Kolenko MA (2008) Method for growth of young Azov-Chernomorskaya royal fish in ponds. RU patent no. 2376755. https://patents.google.com/patent/RU2376755C1/en. Accessed on 21 Apr, 2020
Chistyakov V, Melnikov V, Chikindas ML, Khutsishvili M, Chagelishvili A, Bren A, Kostina N, Cavera V, Elisashvili V (2015) Poultry-beneficial solid-state Bacillus amyloliquefaciens B-1895 fermented soybean formulation. Biosci microbiota food health 3:25–28. https://doi.org/10.12938/bmfh.2014-012
Prazdnova EV, Chistyakov VA, Churilov MN, Mazanko MS, Bren AB, Volski A, Chikindas ML (2015) DNA-protection and antioxidant properties of fermentates from Bacillus amyloliquefaciens B-1895 and Bacillus subtilis KATMIRA1933. Lett Appl Microbiol 61:549–554. https://doi.org/10.1111/lam.12491
Mazanko MS, Gorlov IF, Prazdnova EV, Makarenko MS, Usatov AV, Bren AB, Chistyakov VA, Tutelyan AV, Komarova ZB, Mosolova NI, Pilipenko DN, Krotova OE, Struk AN, Lin A, Chikindas ML (2018) Bacillus probiotic supplementations improve laying performance, egg quality, hatching of laying hens, and sperm quality of roosters. Probiotics Antimicrob Proteins 10(2):367–373. https://doi.org/10.1007/s12602-017-9369-4
Prazdnova EV, Mazanko MS, Chistyakov VA, Denisenko YV, Makarenko MS, Usatov AV, Bren AB, Tutelyan AV, Komarova ZB, Gorlov IF, Weeks R, Chikindas ML (2019) Effect of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 on the productivity, reproductive aging, and physiological characteristics of hens and roosters. Benef Microbes 10(4):395–412. https://doi.org/10.3920/BM2018.0149
Ledder RG, Timperley AS, Friswell MK, Macfarlane S, McBain AJ (2008) Coaggregation between and among human intestinal and oral bacteria. FEMS Microbiol Ecol 66:630–636. https://doi.org/10.1111/j.1574-6941.2008.00525.x
Kos B, Suskovic J, Vukovic S, Simpraga M, Frece J, Matosic S (2003) Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J Appl Microbiol 94:981–987. https://doi.org/10.1046/j.1365-2672.2003.01915.x
Schachter B (2003) Slimy business-the biotechnology of biofilms. Nat Biotechnol 21:361–365. https://doi.org/10.1038/nbt0403-361
Annous BA, Fratamico PM, Smith JL (2009) Quorum sensing in biofilms: why bacteria behave the way they do. J Food Sci 74:R24–R37. https://doi.org/10.1111/j.1750-3841.2008.01022.x
Algburi A, Zehm S, Netrebov V, Bren AB, Chistyakov V, Chikindas ML (2017b) Subtilosin prevents biofilm formation by inhibiting bacterial quorum sensing. Probiotics Antimicrob Proteins 9(1):81–90. https://doi.org/10.1007/s12602-016-9242-x
Clinical and Laboratory Standards Institute (2016) Performance standards for antimicrobial susceptibility testing; twenty–second informational supplement. CLSI Document M 100- S26, Wayne
Cisar JO, Kolenbrander PE, McIntire FC (1979) Specificity of coaggregation reactions between human oral streptococci and strains of Actinomyces viscosus or Actinomyces naeslundii. Infect Immun 24(3):742–752
Borucki MK, Peppin JD, White D, Loge F, Call DR (2003) Variation in biofilm formation among strains of Listeria monocytogenes. Appl Environ Microbiol 69(12):7336–7342. https://doi.org/10.1128/AEM.69.12.7336-7342.2003
Qiu J, Wang D, Zhang Y, Dong J, Wang J, Niu X (2013) Molcular modeling reveals the novel inhibition mechanism and binding mode of three natural compounds to staphylococcal alpha-hemolysin. PLoS One 8(11):e80197. https://doi.org/10.1371/journal.pone.0080197
Naimi HM, Rasekh H, Noori AZ, Bahaduri MA (2017) Determination of antimicrobial susceptibility patterns in Staphylococcus aureus strains recovered from patients at two main health facilities in Kabul, Afghanistan. BMC Infect Dis 17(1):737. https://doi.org/10.1186/s12879-017-2844-4
Chinnambedu RS, Marimuthu RR, Sunil SS, Amrose P, Ramachandran V, Pachamuthu B (2020) Changing antibiotic resistance profile of Staphylococcus aureus isolated from HIV patients (2012–2017) in Southern India. J Infect Public Health 31:75–79. https://doi.org/10.1016/j.jiph.2019.06.015
Lee JS, Chung MJ, Seo JG (2013) In vitro evaluation of antimicrobial activity of lactic acid bacteria against Clostridium difficile. Toxicol Res 29(2):99–106. https://doi.org/10.5487/TR.2013.29.2.099
Mariam SH, Zegeye N, Tariku T, Andargie E, Endalafer N, Aseffa A (2014) Potential of cell-free supernatants from cultures of selected lactic acid bacteria and yeast obtained from local fermented foods as inhibitors of Listeria monocytogenes, Salmonella spp. and Staphylococcus aureus. BMC res notes 7:606. https://doi.org/10.1186/1756-0500-7-606
Aminnezhad S, Kermanshahi RK, Ranjbar R (2015) Evaluation of synergistic interactions between cell-free supernatant of Lactobacillus strains and amikacin and genetamicin against Pseudomonas aeruginosa. Jundishapur J Microbiol 8(4):e16592. https://doi.org/10.5812/jjm.8(4)2015.16592
Moghadam SS, Khodaii Z, Zadeh SF, Ghooshchian M, Aghmiyuni ZF, Shabestari TM (2018) Synergistic or antagonistic effects of probiotics and antibiotics-alone or in combination-on antimicrobial-resistant Pseudomonas aeruginosa isolated from burn wounds. Arch Clin Infect Dis 13(3). https://doi.org/10.5812/archcid.6312
Rishi L, Mittal G, Agarwal RK, Sharma T (2017) Melioration in anti-staphylococcal activity of conventional antibiotic (s) by organic acids present in the cell free supernatant of Lactobacillus paraplantarum. Indian J Microbiol 57(3):359–364. https://doi.org/10.1007/s12088-017-0659-z
Kok ET, Jong MC, Gravendeel B, Van Leeuwen WB, Baars EW (2015) Resistance to antibiotics and antifungal medicinal products: can complementary and alternative medicine help solve the problem in common infection diseases? The introduction of a dutch research consortium. Evid Based Complement Alternat Med 2015:521584–521586. https://doi.org/10.1155/2015/521584
Gokarn K, Pal RB (2018) Activity of siderophores against drug-resistant gram-positive and gram-negative bacteria. Infect Drug Resist 1:61–75. https://doi.org/10.2147/IDR.S148602
Abd El-Baky RM, Sandle T, John J, Abuo-Rahma GEA, Hetta HF (2019) A novel mechanism of action of ketoconazole: inhibition of the NorA efflux pump system and biofilm formation in multidrug-resistant Staphylococcus aureus. Infect Drug Resist 12:1703–1718. https://doi.org/10.2147/IDR.S201124
Algburi A, Volski A, Chikindas ML (2015) Natural antimicrobials subtilosin and lauramide arginine ethyl ester synergize with conventional antibiotics clindamycin and metronidazole against biofilms of Gardnerella vaginalis but not against biofilms of healthy vaginal lactobacilli. Pathog dis 73(5). https://doi.org/10.1093/femspd/ftv018
Younes JA, van der Mei HC, van den Heuvel E, Busscher HJ, Reid G (2012) Adhesion forces and coaggregation between vaginal staphylococci and lactobacilli. PLoS One 7(5):e36917. https://doi.org/10.1371/journal.pone.0036917
Jain N, Meta A, Bharti V (2017) Screening, characterization, and in vitro evaluation of probiotic properties of Lactobacillus strains. Asian J pharm and Clin Res 10(8):288. https://doi.org/10.22159/ajpcr.2017.v10i8.14233
Soleimani NA, Kermanshahi RK, Yakhchali B, Sattari TN (2010) Antagonistic activity of probiotic lactobacilli against Staphylococcus aureus isolated from bovine mastitis. African J Microbiol Res 4(20):2169–2173
Zamani H, Rahbar S, Garakoui SR, Afsah Sahebi A, Jafari H (2017) Antibiofilm potential of Lactobacillus plantarum spp. cell free supernatant (CFS) against multidrug resistant bacterial pathogens. Pharm Biomed Res 3(2):39–44. https://doi.org/10.29252/pbr.3.2.39
Khiralla GM, Mohamed EA, Farag AG, Elhariry H (2015) Antibiofilm effect of Lactobacillus pentosus and Lactobacillus plantarum cell-free supernatants against some bacterial pathogens. J Biotech Res 6:86–95
Satpute S, Mone N, Das P, Banpurkar A, Banat I (2018) Lactobacillus acidophilus derived biosurfactant as a biofilm inhibitor: a promising investigation using microfluidic approach. Appl Sci 8(9):1555. https://doi.org/10.3390/app8091555
Wong CB, Khoo BY, Sasidharan S, Piyawattanametha W, Kim SH, Khemthongcharoen N, Ang MY, Chuah LO, Liong MT (2014) Inhibition of Staphylococcus aureus by crude and fractionated extract from lactic acid bacteria. Benef Microbes 6(1):129–139. https://doi.org/10.3920/BM2014.0021
Koohestani M, Moradi M, Tajik H, Badali A (2018) Effects of cell-free supernatant of Lactobacillus acidophilus LA5 and Lactobacillus casei 431 against planktonic form and biofilm of Staphylococcus aureus. Vet res forum 9(4):301–306. https://doi.org/10.30466/vrf.2018.33086
Hor YY, Liong MT (2014) Use of extracellular extracts of lactic acid bacteria and bifidobacteria for the inhibition of dermatological pathogen Staphylococcus aureus. Dermatol Sinica 32(3):141–147. https://doi.org/10.1016/j.dsi.2014.03.001
Acknowledgements
The authors would like to acknowledge the deanery of the Science College, University of Diyala and the General Teaching Hospital in Baqubah, Iraq, for providing free access to their equipment and facilities required for this study and for providing us with clinical isolates of MRSA and MSSA, as a gift.
Funding
AME and MLC were supported by the Ministry of Science and Higher Education of the Russian Federation (Project Number 075-15-2019-1880).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Algburi, A., Al-Hasani, H.M., Ismael, T.K. et al. Antimicrobial Activity of Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 Against Staphylococcus aureus Biofilms Isolated from Wound Infection. Probiotics & Antimicro. Prot. 13, 125–134 (2021). https://doi.org/10.1007/s12602-020-09673-4
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12602-020-09673-4