
Abstract
Dendrolimus pini is one of the most harmful defoliators of Scots pine in Poland and other countries in Europe and Central Asia. The monitoring of this insect needs to be improved and to include the use of pheromone traps. Laboratory studies were conducted to compare changes in the residual amounts of (Z,E)-5,7-dodecadienal and (Z,E)-5,7-dodecadien-1-ol, the major components of the D. pini pheromone, in four types of dispensers. The two most promising dispensers, red rubber cylindrical dispensers (rubber cylinders) and polyethylene vials, were tested in the field for their ability to attract D. pini males. In addition, the effects of trap type (sticky wing, non-sticky cross-vane, bucket, funnel and modified gypsy moth milk carton) and height of trap placement (1.5 m, 5–6 m and tree crown) on trap catches were tested in the field. The vials provided the longest half-lives for both the aldehyde and alcohol under laboratory conditions. These dispensers were also significantly more effective in attracting D. pini males to the traps than the rubber cylinders in the field experiment. The cross-vane and bucket traps were the most effective among the tested traps. The height of trap placement had a significant effect on catches, with more males caught in traps that were hung at 5–6 m above the ground than those hung in the tree crowns. Our results may be useful for developing a monitoring system for D. pini in Europe and other regions where it occurs.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The pine-tree lappet moth, Dendrolimus pini L. (Lepidoptera: Lasiocampidae) is a pest of Scots pine forests in Poland and other countries in Central and Eastern Europe, as well as in Russia, although its natural range is much wider, extending from Western Europe to Central Asia (USDA, APHIS, PPQ 2011). In Poland, it is a univoltine species with the peak of flight activity in mid-July–early August. Larvae hatch approximately two weeks after oviposition. They first feed on egg shells and then on current year needles. After the first autumn frost, the second- through fifth-instar larvae descend to the forest litter for overwintering. In early spring, they climb up to the tree crown and feed until July. The pupal stage lasts approximately 2 weeks.
The main method for population density assessment is based on counting the larvae found in the forest litter in 10 plots 50 × 100 cm in size (Forest protection guidelines [Instrukcja ochrony lasu] 2012). Due to cryptic coloration, the larvae, particularly young instars, may be overlooked, and thus it may lead to population density underestimation and consequently to an unexpected outbreak. The importance of an efficient early warning system regarding population increases has grown in recent years. Although D. pini outbreaks have been observed in Poland since the 1940s, the largest outbreak occurred in 2012–2014, covering over 184 thousand ha (Jabłoński 2014). Moreover, D. pini has also invaded new regions, e.g., Scotland (Ray et al. 2016; Moore et al. 2017) and southern Sweden where it caused an isolated outbreak (Björkman et al. 2013). There is a high risk of D. pini outbreaks in other countries in northern Europe due to climate warming (Ray et al. 2016), which can have direct (e.g., through changes from semi- to univoltinism) and indirect (e.g., through plant quality) effects on this insect species.
Pheromone traps are used in monitoring and as early warning systems for many insect species. However, their effectiveness depends on different factors, including dispenser type, trap design and trap placement (Schmidt and Roland 2003; Laurent and Frérot 2007; Haynes et al. 2007; Zhang et al. 2013; Álvarez et al. 2015). These issues have not received much attention in studies on Dendrolimus spp. Sticky delta or wing traps baited with grey rubber septum lures and suspended at a height of approximately 1.5 m have been used most often (Priesner et al. 1984; Kong et al. 2001, 2007, 2011; Zhang et al. 2003; Ostrauskas and Ivinskis 2011). Other types of dispensers or traps have also been used (Johansson et al. 2002; Baranchikov et al. 2007; Ponomarev et al. 2006; Li et al. 2015), but their effectiveness was not assessed in the same experiment. A single exception was the study by Priesner et al. (1984), who found that the effectiveness of tetra sticky traps was comparable to that of the cross-vane traps with Plexiglas wings, but the authors indicated that there was a saturation issue in the sticky traps. To our knowledge, the effect of trap placement on Dendrolimus catches has never been estimated.
Although the pheromone of D. pini is known to consist of two components, (Z,E)-5,7-dodecadienal (Z5,E7–12:Ald) and (Z,E)-5,7-dodecadien-1-ol (Z5,E7–12:OH) (Bestmann et al. 1982; Priesner et al. 1984; Kovalev et al. 1993), a monitoring system for this species has not been developed. Therefore, we focused on operational factors that could influence the number of moths captured in pheromone-baited traps. The aim of our research was to compare 1) changes with time in the residual amounts of the pheromone components in different dispenser types under laboratory conditions, 2) the effectiveness of red rubber cylindrical dispensers and polyethylene vials under field conditions, 3) the effectiveness of sticky and non-sticky traps under field conditions, and 4) the effectiveness of the traps suspended at different heights under field conditions. The effectiveness is considered to be the number of male moths captured in the traps of different types, hung at different heights or baited with different dispensers.
Materials and methods
Pheromone components and their purity
Z5,E7–12:Ald and Z5,E7–12:OH, the basic components of the D. pini pheromone, were synthesized and purified using procedures based on Stille’s reaction (Stille and Groh 1987; Stille and Simpson 1987).
The chemical and isomeric purity of freshly prepared compounds was determined by gas chromatography (GC) on a low-polar Rxi-5ms capillary column (30 m × 0.25 mm inside diameter, 0.25-μm film thickness, Restek, USA) using an HP 5890 series II gas chromatograph with a flame ionization detector (Agilent Technologies, USA; hereinafter called GC-FID). The injector and detector temperatures were 200 °C and 250 °C, respectively. The oven temperature was programmed from 100 °C (5 min hold) to 280 °C at 10 °C/min and then held constant for 10 min.
To obtain the compounds of a higher chemical and isomeric purity (hereinafter called “purified”), column chromatography on silica gel loaded with 10% silver nitrate was used (Hasdenteufel 2006). Purified compounds were used in one experiment only, i.e., in field testing of two dispenser types: rubber cylinders and polyethylene vials (described below).
Temporal changes in the residual amount of pheromone components in different dispensers under controlled conditions
In this study, we used a similar approach as that described by Butler and McDonough (1979) and Zhang et al. (2013). Four types of pheromone dispensers were studied:
- 1)
red rubber cylindrical dispensers in the form of empty cylinders with an outer diameter of 10 mm, wall thickness of 1.5 mm, length of 15 mm and weight of ca. 0.4 g (hereinafter called rubber cylinders),
- 2)
red precision seal rubber septa (Sigma-Aldrich Sp. z o.o., Poland) formed by two empty 8 mm long cylinders with outer diameters of 6.8 and 5 mm joined by a 2-mm wide layer and with a weight of ca. 0.4 g (hereinafter called septa),
- 3)
polyethylene vials with push-on hinged lids (Kartell, Italy), with an outer diameter of 8 mm, height of 32 mm, wall thickness of 0.98 mm and weight of 0.71 g (hereinafter called vials), and
- 4)
sections of a poly(ethylene-vinyl acetate), 12%, tube with an outer diameter of 11.7 mm, height of 40 mm, wall thickness of 0.75 mm and weight of 0.7 g (hereinafter called tubes).
The pheromone consisting of Z5,E7–12:Ald and Z5,E7–12:OH with a mass ratio of 6:4 (Kovalev et al. 1993) was introduced into the dispensers by the following procedures. The rubber cylinders were immersed for 2.5 min in a solution of 10 mg of pheromone in 1 ml of n-hexane (99% for HPLC; POCH, Poland), hereafter called hexane. The septa, vials and tubes were loaded with a solution of 1 mg of pheromone in 20 μl of hexane. After the evaporation of hexane, the dispensers were ready for use. The ratio of pheromone components and their absolute amount in the freshly prepared dispensers was verified at the beginning of the experiment (time = 0 days) by conducting solvent extractions from 3 dispensers of each type and GC analyses (see below).
Then, 15 dispensers of each type were placed in a fume hood at a temperature of 20–25 °C and air velocity of 1.6 m∙s−1. Three dispensers of each type at a time were removed from the fume hood, cut into pieces, placed into separate 4 ml vials and flooded with a mixture of 3.5 ml hexane and 5 mg dodecanol added as an internal standard for GC analyses. The rubber cylinders, septa and vials were sampled weekly for 35 days, while the tubes were sampled every 3–5 days for 19 days due to a much faster decrease in the residual amount of the pheromone observed in preliminary trials. The analyses of Z5,E7–12:Ald and Z5,E7–12:OH extracted from the dispensers were carried out by GC-FID with the Rxi-5ms capillary column (see above), with the temperature ramp programmed at 100 °C for 5 min, then to 280 °C at 10 °C∙min−1 and held for 5 min. Nitrogen was used as a carrier gas at a pressure of 100 kPa.
The residual amounts of pheromone components in the dispensers were used to calculate their release rates (see Statistical analyses).
Field tests of dispenser types
We tested the effectiveness of the rubber cylinders and vials loaded with hexane solutions of purified 0.6 mg Z5,E7–12:Ald and 0.4 mg Z5,E7–12:OH (isomeric purity ~97% and chemical purity ~99%) using the same procedure as in the laboratory study (see above). The experiment was conducted between 29 July and 6 August 2015 in a 63-year-old Scots pine (Pinus sylvestris L.) stand in the Krucz forest district (52°46′ N, 16°25′ E). The dispensers (8 lures per dispenser type, total N = 16 traps) were tested in white cross-vane IBL-5 traps (R&D Chemipan, Warsaw, Poland), presented in Online Resource 1 and hereinafter called cross-vane traps. The 20 × 20 cm vanes were inserted into a 17-cm diameter funnel. The base of the collectors had a hole covered with metal mesh for water drainage. Each collector contained a 3.5 × 3 cm strip saturated with an insecticide (7% transfluthrin) (Bros sp. z o.o., Poznań, Poland) to kill the trapped moths. The treatments were arranged in a completely randomized design, i.e., in a random order, along two lines. An adapted telescopic pole (folded length – 1.1 m, extended length – 5 m) (Paradox Company, Krakow, Poland) was used to suspend the traps on metal hooks from dead branches on the trees, 4–6 m above the ground. The distance between the traps and lines was at least 20 m.
Testing the effect of trap type
The effect of trap type on catches of D. pini males was evaluated in two experiments. Experiment 1 was conducted between 1 and 19 August 2014 in 37–43-year-old Scots pine stands in the Czlopa forest district (53°08′ N, 16°02′ E). Three types of traps (all presented in Online Resource 1) were tested: 1) a white sticky wing-type PL-2 trap, with the upper and bottom parts 25 × 23 cm in size and the bottom part (internal side) covered with glue (Chemipan R&D Laboratories, Warsaw, Poland), hereinafter called sticky wing trap, 2) the cross-vane trap (described above), and 3) a green bucket-type trap (Agrisense BCS Ltd., Pontypridd, UK), with a height of 22 cm, perimeter (in the widest part) of 53 cm, upper and lower funnel inner diameters of approximately 8.5 and 3 cm, respectively, hereinafter called bucket_green. No insecticide was used in the traps to kill the captured moths so that the ability of non-sticky traps to catch and keep the captured insects inside the traps could be compared. The traps were baited with rubber cylinders loaded with 0.6 mg Z5,E7–12:Ald and 0.4 mg Z5,E7–12:OH (isomeric purity ~94%, chemical purity ~90%) dissolved in 0.05 ml of hexane. The traps were hung at 4–6 m above the ground along two lines as described above. The traps and lines were at least 20 m apart. This trial consisted of 7 replications (traps) of each trap type (total N = 21 traps). The treatments were arranged in a completely randomized design. The traps were checked, cleaned and rotated every 1–7 days. Rotations were not considered as replications.
Experiment 2 was established on 6 August 2015 in a 63-year-old Scots pine stand in the Krucz forest district (52°46′ N, 16°25′ E) and lasted until 13 August. In this experiment, four trap types (all presented in Online Resource 1) were tested: 1) the cross-vane trap, 2) a white funnel trap, which was similar to the cross-vane trap except that the large vanes were replaced with small vanes in a trapezoid shape with 10-cm and 6-cm sides that were 5 cm high and placed in the funnel so that the distance between the lid and the funnel edge was approximately 2–3 cm (both traps were delivered by Chemipan R&D Laboratories, Warsaw, Poland), 3) a bucket-type trap similar in size to the bucket_green trap (see above) with a green lid, yellow funnel and transparent bucket, hereinafter called bucket_colour (Pherobank, Wijk bij Duurstede, the Netherlands), and 4) a green gypsy moth milk carton trap (Alpha Scents, Inc., West Linn, OR, USA) modified by cutting two entry holes 1.0–1.6 × 3.0–3.7 cm in size on each side of the trap, hereinafter called GM_modif. The collectors of each trap type were provided with a strip saturated with an insecticide (as described in the lure testing experiment). The traps were baited with the vials loaded with a hexane solution of 1.2 mg Z5,E7–12:Ald and 0.8 mg Z5,E7–12:OH (isomeric and chemical purity was the same as in Experiment 1). The higher dose of pheromone was used to enhance the moth catches (based on earlier unpublished trials), and the same dose was used in all traps tested in this experiment. The traps (6 traps/trap type, total N = 24 traps) were deployed in a randomized complete block design, with one trap of each type per block, in the distance of at least 20 m between the traps and blocks. The traps were checked, cleaned and rotated twice during the trapping period. Rotations were not considered as replications.
Testing the effect of trap placement
This experiment was conducted between 29 July and 20 August 2015 in an 84-year-old Scots pine stand with an average tree height of 18 m in the Wolsztyn forest district. The bucket_colour traps with the added insecticide strip (described above) were hung at three heights above the ground: 1.5 m (on a plastic thread between the trees), 5–6 m (using the telescopic pole) and in the crown, with 5 replicates (traps) per height level and one trap per tree (N = 15 traps). To hang the traps in the crown, a fishing rod was used to throw a heavy sinker with an attached cord over the branches of any part of the tree crown. Then, the trap was attached to the cord, and both cord ends were joined to create a loop, which was used to lift a trap until it reached any branch in the tree crown. The opposite end of the loop was twisted around the tree trunk at 1.5 m above the ground to prevent the trap from falling down and enable trap inspection. The traps were baited with the vials loaded with the same pheromone as in Experiment 2. The traps were set along two lines with a distance of approximately 20 m between the traps and lines in a randomized complete block design, with one trap at each height per block. All traps were hung at a distance of at least 50 cm from the tree trunk. The traps were checked and cleaned every 7–8 days.
Statistical analyses
The temporal changes in the residual amounts of Z5,E7–12:Ald and Z5,E7–12:OH in the four types of dispensers were compared by calculating their half-lives and ratios. To calculate the half-lives of these compounds, an exponential function describing the first-order kinetic process (e.g., McDonough et al. 1989) was initially used to describe the dynamics of decrease in the residual amounts (results not presented in this manuscript). However, we found that the function y = (a + bx)/(1 + cx), which is the ratio of the first order polynomial functions where y is the residual amount (mg) of the pheromone component in a dispenser; x is the time (days); and a, b and c are function parameters, described these temporal changes slightly better (assessed by a coefficient of determination R2). Therefore, this latter function was used to calculate the half-lives of Z5,E7–12:Ald and Z5,E7–12:OH in each dispenser type.
The release rates of pheromone components from the dispensers were calculated from their residual amounts as the ratio of weight loss to the time of release (each rate value was assigned to the middle point of the release period). The time derivative of the function mentioned above, i.e., (b-ac)/(1 + cx)2, was successfully used to approximate the release rates of the pheromone components from the dispensers.
The analyses were conducted using the Sigma Plot 12.5 software package (Systat Software, USA).
The effect of the dispenser type used in the cross-vane traps on the total catches of D. pini male moths was tested by using a generalized linear model (GLM) with a negative binomial error distribution (NBGLM).
The total numbers of males captured in the different types of traps in experiment 1 were compared with the Kruskal-Wallis test followed by multiple rank comparisons (Siegel and Castellan Jr 1988). The non-parametric test was used because it handled the data with no variance, i.e., zero catches in the sticky wing traps, better than GLM.
Experiment 2 was established using a randomized complete block design. The effect of trap type on the total number of captured moths was estimated by using a generalized linear mixed model with a negative binomial error distribution (NBGLMM) followed by a post hoc test with a Holm correction for multiple mean comparisons. The same approach was used to test the effect of the trap location on the moth catches. In both cases, the blocks were included as a random factor in the models.
All analyses of the data from the field experiments were performed using Statistica 10 software (StatSoft Inc. 2011) or the R environment, version 3.5.1 (R Core Team 2018) with RStudio, version 1.1.463 (R Studio Team 2016). The following packages were used: MASS (Venables and Ripley 2002) for NBGLM, glmmADMB (Fournier et al. 2012; Skaug et al. 2016) for NBGLMM, and multcompView (Graves et al. 2015) and emmeans (Lenth 2018) for multiple mean comparisons. The model’s goodness of fit was estimated by using the ratio of the residual deviance to the residual degrees of freedom, i.e., checking for overdispersion (Mangiafico 2016). The significance level was set at α = 0.05 for all analyses.
Results
Effect of dispenser type on temporal changes in the residual amount and release rates of pheromone components under controlled conditions
The amounts of residual Z5,E7–12:Ald and Z5,E7–12:OH in the four studied dispensers decreased at different rates over the course of 19–35 days (Online Resource 2). The applied function described the dynamics of this process very well (Online Resource 2) as indicated by the high values of R2 (Table 1). The only exceptions were the temporal changes of both pheromone components in the tubes.
The release rates of Z5,E7–12:Ald in all tested dispensers were much higher (Fig. 1) and their half-lives shorter than those of the corresponding alcohol (Table 1). Among tested dispensers, the vials provided the lowest release rates of both the aldehyde and alcohol (Fig. 1), and therefore they had the longest half-lives (11.9 and 18.9 days, respectively). In contrast, the fastest changes in the amount of both components were in the septa, with half-lives equal to 1.4 days for Z5,E7–12:Ald and 7.4 days for Z5,E7–12:OH.

Release rates (mean ± SD) of Z5,E7–12:Ald (aldehyde) and Z5,E7–12:OH (alcohol) from the four types of dispensers in still air
The mass ratio of the residual Z5,E7–12:Ald and Z5,E7–12:OH in all the tested dispensers quickly decreased from the initial values of 1.3–1.7 to less than 1 (Fig. 2). The slowest decrease occurred in the vials that even after 7 days contained more aldehyde than alcohol, and the mass ratio in these vials did not drop below 0.6 by the end of the experiment. In the other dispensers, the minimum ratios observed were 0.05 (in septa) and 0.21 (in rubber cylinders and tubes).

The effect of dispenser type on temporal changes in the mass ratios of residual Z5,E7–12:Ald and Z5,E7–12:OH measured in the laboratory tests
Effect of dispenser type under field conditions
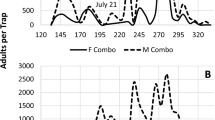
The total numbers of D. pini males caught in the traps with the rubber cylinders over 8 days were very low and varied from 0 to 8 males∙trap−1. In contrast, the numbers of males attracted to the traps with the vials were high, with a much larger variation (from 9 to 110 males∙trap−1) (Fig. 3). The dispenser type had a significant effect on the catches – the vials were significantly more effective than the rubber cylinders in attracting male moths to the traps (NBGLM, z = 5.74, P < 0.0001).

Mean (±SE) total numbers of Dendrolimus pini male moths caught in cross-vane traps baited with the rubber cylinders or vials loaded with the pheromone (0.6 mg of Z5,E7–12:Ald and 0.4 mg of Z5,E7–12:OH) from 29 July – 6 August 2015. The difference was tested using a negative binomial generalized linear model; the different letters indicate means that are significantly different at α = 0.05
Trap type
Surprisingly, in experiment 1, the sticky wing traps did not catch any D. pini moths, whereas the non-sticky cross-vane and bucket_green traps caught up to 5 and 8 males∙trap−1, respectively, over 18 days (Fig. 4a). The differences in the catches were significant (H = 12.3, df = 2, N = 21, P = 0.0021). The numbers of D. pini males in each of the non-sticky traps were significantly higher than those in the sticky traps (post hoc test: cross-vane traps vs sticky wing traps – P = 0.0102; bucket_green traps vs sticky wing traps – P = 0.0095), while the catches in the non-sticky traps were comparable (P > 0.05).

Mean (±SE) total numbers of Dendrolimus pini male moths caught in: (a) three trap types baited with the rubber cylinders loaded with the pheromone (0.6 mg of Z5,E7–12:Ald and 0.4 mg of Z5,E7–12:OH) from 1 to 19 August 2014. The differences were tested with the Kruskal-Wallis test followed by post hoc multiple rank comparisons; (b) four types of non-sticky traps baited with the vials loaded with the pheromone (1.2 mg of Z5,E7–12:Ald and 0.8 mg of Z5,E7–12:OH) from 6 to 13 August 2015. The differences were tested using a negative binomial generalized linear mixed model followed by a post hoc test with a Holm correction for multiple mean comparisons; the different letters indicate means that are significantly different at α = 0.05
Experiment 2 aimed to compare the effectiveness of the non-sticky traps only. The trap type was found to have a significant effect on the D. pini male catches (NBGLMM, χ2 = 13.9, df = 3, P = 0.0031). The highest numbers of male moths were observed in the cross-vane traps (87.8 ± 10.10 males∙trap−1, mean ± SE) and bucket_colour traps (52.3 ± 13.35 males∙trap−1) (Fig. 4b). The catches in the GM_modif traps and the funnel traps were much lower (26.5 ± 11.52 males∙trap−1 and 24.0 ± 2.83 males∙trap−1, respectively), and the difference in the effectiveness between these traps and the cross-vane traps was close to significant (P = 0.0865) and significant (P = 0.0014), respectively.
Trap placement
The numbers of D. pini male moths caught in the traps hung at three height levels differed significantly (NBGLMM, χ2 = 11.2, df = 2, P = 0.0036). The highest catches were in traps located at a height of 5–6 m (62.6 ± 11.84 males∙trap−1), and they were significantly higher (Fig. 5) than those in the traps located in the crowns (22.8 ± 3.80 males∙trap−1, P = 0.0147). The number of moths caught in the traps at 1.5 m (37.2 ± 6.39 males∙trap−1) was higher than in the traps in the crowns, but lower than in the traps at 5–6 m, and in both cases the difference was close to significant (P = 0.0687).

Mean (±SE) total numbers of Dendrolimus pini male moths caught in the bucket_colour traps hung at different heights from 29 July – 20 August 2015 (dispenser and pheromone as in Fig. 4b). The differences were tested using a negative binomial generalized linear mixed model followed by a post hoc test with a Holm correction for multiple mean comparisons; the different letters indicate that the differences are significant at α = 0.05
Discussion
Different types of dispensers are used in traps to attract different insect species. The type of dispenser selected for a certain species depends on the chemical composition of the attractant to be loaded into the dispensers (Butler and McDonough 1979, 1981). Under laboratory conditions, we studied the changes with time in the residual amounts of Z5,E7–12:Ald and Z5,E7–12:OH, the major pheromone components of D. pini (Bestmann et al. 1982; Priesner et al. 1984; Kovalev et al. 1993), D. superans sibiricus Chetverikov (Klun et al. 2000; Pletnev et al. 2000) and D. superans Butler (Kong et al. 2001), in 4 types of dispensers: red rubber cylinders, red precision seal rubber septa, polyethylene vials and sections of poly(ethylene-vinyl acetate) tube.
The release rates of Z5,E7–12:Ald were higher than those of the corresponding alcohol in all tested dispensers. The shorter half-life of Z5,E7–12:Ald may be explained by its higher reactivity and volatility. The lowest release rates of both Z5,E7–12:Ald and Z5,E7–12:OH were observed in the vials. Our studies did not confirm the findings of Li et al. (2015), who suggested that closed vials do not release Z5,E7–12:Ald and Z5,E7–12:OH, although we obtained similar results in the trials without hexane, i.e., with pure pheromone components (not published). Most likely, the hexane we used to dissolve the pheromone components facilitated their migration through the polyethylene walls. Hexane could enter the vial wall and loosen the polymer structure to allow the transfer of larger aliphatic compounds (Harding and Gladden 1998; Makitra et al. 2001). The rubber cylinders and the vials had comparable half-lives for Z5,E7–12:OH, but the rubber cylinders had a much lower half-life for Z5,E7–12:Ald. The septa appeared the least effective because of the fast evaporation of both active pheromone compounds, particularly Z5,E7–12:Ald. This result is consistent with an earlier report by Priesner et al. (1984, ref. to unpublished results of M.D. Chrisholm and D.W. Reed) who observed an exponential decrease in Z5,E7–12:Ald from rubber septa by half every six days. Khrimian et al. (2002) showed a significant decrease in the effectiveness of septa lures saturated with Z5,E7–12:Ald and Z5,E7–12:OH (1:1) after 2 weeks in field tests against the Siberian moth D. superans sibiricus Chetverikov.
The ratio of the residual aldehyde and alcohol decreased with time in all the dispensers, but the rate was lowest in the vials. This ratio along with additional pheromone components serving as repellents for closely related species plays an important role in between-species discrimination of moths during the flight period in the regions shared by more than one species of Dendrolimus spp. (Kovalev et al. 1993; Klun et al. 2000; Pletnev et al. 2000; Kong et al. 2001, 2007, 2011). In the case of D. pini and D. superans sibiricus sharing the two major pheromone components (Z5,E7–12:Ald and Z5,E7–12:OH), their optimal ratio (based on initial loadings of lures in the field tests) slightly differs, i.e., 1.5:1 and 1:1, respectively, and the presence of (Z,E)-5,7-dodecadienyl acetate in the pheromone of D. superans sibiricus (Pletnev et al. 2000) will inhibit attraction of D. pini (Priesner et al. 1984).
In the field tests, the vials were much more effective in attracting D. pini moths than were the rubber cylinders, that can be explained by the difference in the ratio of Z5,E7–12:Ald and Z5,E7–12:OH after 8 days (1.036 and 0.651, respectively, estimated from the laboratory experiments), i.e., still above 1 in the vials and much below 1 in the rubber cylinders. Dispenser exposure was relatively short; therefore, the effect of longer exposure under the field conditions on dispenser effectiveness still needs to be evaluated. However, the lower release rate of the main pheromone components, particularly Z5,E7–12:Ald, in the vials in the laboratory studies indicated that they could be active in the field longer than other dispensers. Similarly, Schmidt and Roland (2003) showed that traps baited with rubber septa loaded with Z5,E7–12:Ald and (Z,Z)-5,7-dodecadienal (100:1) produced significantly lower trap catches of Malacosoma disstria Hübner (Lepidoptera: Lasiocampidae) and higher zero-catch frequencies than polyurethane lures with the same pheromone load. In contrast, Hall et al. (2017) did not find any difference in a response of Dioryctria mendacella (Staudinger) (Lepidoptera: Pyralidae) to the blend of (Z,Z,Z,Z,Z)-3,6,9,12,15-pentacosapentaene and (Z,E)-9,11-tetradecadienyl acetate (10:1) emitted from rubber septa and vials.
Surprisingly, neither moths nor moth signs (scales, legs, etc.) were observed in the sticky wing traps. The shape of these traps is slightly different from that of the wing traps used in other experiments, e.g., Pherocone 1C (Kong et al. 2001, 2007, 2011), and could affect both the odour distribution from the dispensers and the capture probability of moths inside the traps. Sticky traps of any shape were excluded from further studies due to a saturation issue at high insect densities and to the lower or equal effectiveness in comparison to the non-sticky traps at endemic insect densities (Priesner et al. 1984; Schmidt and Roland 2003; Baranchikov et al. 2004). This issue was also the reason for changing the wing traps in 2012 to the modified gypsy moth (GM) milk carton trap (Lance 2006) as the preferred trap for the Dendrolimus species survey in the USA (CAPS Approved Methods 2011). In our tests, the effectiveness of the GM-modified trap was much lower than those of the bucket or cross-vane traps. This result could be caused by entry holes that were too small in our modified GM traps (1.0–1.6 cm high × 3.0–3.7 cm wide) in comparison to the hole size in the GM modified traps used in the USA (3 cm high × 7 cm) (Klun et al. 2000). On the other hand, Baranchikov et al. (2004) concluded that entry holes cannot be too large with the optimal size at approximately 2.5 × 2.7 cm. In our two experiments, the cross-vane and bucket traps were the most effective traps. Similar results for bucket traps were obtained in the trap type tests against other insect species (Gray et al. 1991; Schmidt and Roland 2003; Guerrero et al. 2014), although in some cases, the results were not as obvious (Haynes et al. 2007; Zhang et al. 2013).
To our knowledge, this study was the first to test the effect of the height at which traps are hung on Dendrolimus spp. moth catches. The numbers of moths in the traps hung 5–6 m above the ground were significantly higher than those in the traps suspended in the crowns at approximately 15–18 m above the ground and substantially higher than those in the traps at 1.5 m. Haynes et al. (2007) revealed a similar increase in the catches of Malacosoma americanum (F.), another species of Lasiocampidae, with an increasing height of trap placement from 1.5 m to 7.5 m. In contrast to our results, Zhang et al. (2003) showed that traps placed in the forest canopy of Masson pine trees, Pinus massoniana Lamb., up to 5.5 m caught significantly more D. punctatus (Walker) than traps located higher in the canopy. However, these results demonstrated the effect of tree age (younger or older than 15 years with a respective difference in tree height) rather than the effect of height because traps were hung in the tree crowns in both cases. In other large Lepidoptera, the effect of trap height depends on the insect species, e.g., the catches of Lymantria monacha L. (Erebidae) were significantly higher in traps placed at 2 m above the ground than in those at ground level or at a height of 5 m (Wang et al. 2017), while the numbers of Thaumetopoea pityocampa (Thaumetopoeidae) were the highest at the top of the tree crowns in comparison to traps at breast height and at half the height of the tree crown (Jactel et al. 2006). It appears that the height where traps should be placed to obtain the highest effectiveness depends on the behaviour of pheromone-emitting females and oviposition sites. In the case of D. pini, females usually pupate on tree trunks at different heights and oviposit on tree trunks or dead branches; therefore, the highest catches would be expected in traps located at these sites.
In summary, the results of our study show that cross-vane and bucket traps baited with polyethylene vials are the most promising for monitoring D. pini. We recommend hanging the traps at a height of 5–6 m whenever the highest effectiveness is crucial. However, from practical point of view it will be more feasible to hang them at breast height, where the traps will be less, but still effective. When using the traps for monitoring D. pini, the most important issue would be to keep the height level constant.
Future studies should focus on the effect of dispenser type, protectants, pheromone dose and temperature on dispenser longevity. These studies are particularly important given climate change and the fact that D. pini has a flight period during the hottest periods of the year (mid-July to early August). In addition, a relationship between moth catches and population densities at an egg or larval stage should be studied to make monitoring data more meaningful for forest protection practice.
References
Álvarez, G., Etxebeste, I., Gallego, D., David, G., Bonifacio, L., Jactel, H., Sousa, E., & Pajares, J. A. (2015). Optimization of traps for live trapping of pine wood nematode vector Monochamus galloprovincialis. Journal of Applied Entomology, 139, 618–626.
Baranchikov, Y. N., Pet’ko, V. M., Ragenovich, A. R., Klun, J. A., & Mastro, V. K. (2004). Pheromone trap for monitoring population density of the siberian moth. Lesnoe Khozyaistvo, 3, 46–47 (in Russian).
Baranchikov, Y.N., Pet’ko, V.M., Ponomarev, V.L. (2007). The Russians are coming - aren’t they? Siberian moth in European forests. In K.W. Gottschalk (Ed.), proceedings, 17th U.S. Department of Agriculture interagency research forum on gypsy moth and other invasive species 2006; gen.Tech.Rep. NRS-P-10. (pp. 18–20). Newtown Square, PA: U.S. Department of Agriculture, Forest Service, Northern Research Station.
Bestmann, H. J., Koschatzky, K. H., Platz, H., Süβ, J., Vostrowsky, O., Knauf, W., Burghardt, G., & Schneider, I. (1982). Pheromone, XL. Stereoselektive Synthese des Pheromonkomplexes von Lasiocampidae-Arten (Lepidoptera); ein Sexuallockstoff für den Kiefernspinner Dendrolimus pini. European Journal of Organic Chemistry, 7, 1359–1365.
Björkman, C., Lindelöw, Å., Eklund, K., Kyrk, S., Klapwijk, M. J., Fedderwitz, F., & Nordlander, G. (2013). A rare event – An isolated outbreak of the pine-tree lappet moth (Dendrolimus pini) in the Stockholm archipelago. Entomologisk Tidskrift, 134, 1–9.
Butler, L. I., & McDonough, L. M. (1979). Insect sex pheromones: Evaporation rates of acetates from natural rubber septa. Journal of Chemical Ecology, 5, 825–637.
Butler, L. I., & McDonough, L. M. (1981). Insect sex pheromones: Evaporation rates of alcohols and acetates from natural rubber septa. Journal of Chemical Ecology, 7, 627–633.
CAPS Approved Methods (2011). Pine-tree Lappet – Dendrolimus pini. Last accessed February 5, 2018, http://pest.ceris.purdue.edu/services/approvedmethods/sheet.php?v=682.
Forest protection guidelines [Instrukcja ochrony lasu] (2012). Centrum Informacyjne Lasów Państwowych, Warszawa.
Fournier, D. A., Skaug, H. J., Ancheta, J., Ianelli, J., Magnusson, A., Maunder, M., Nielsen, A., & Sibert, J. (2012). AD model builder: Using automatic differentiation for statistical inference of highly parameterized complex nonlinear models. Optimization Methods and Software, 27, 233–249.
Graves, S., Piepho, H.-P., Selzer, L. with help from Dorai-Raj, S. (2015). multcompView: Visualizations of Paired Comparisons. R package version 0.1–7.
Gray, T. G., Shepherd, R. F., Struble, D. L., Byers, J. B., & Maher, T. F. (1991). Selection of pheromone trap and attractant dispenser load to monitor black army cutworm, Actebia fennica. Journal of Chemical Ecology, 17, 309–316.
Guerrero, S., Brambila, J., & Meagher, R. (2014). Efficacies of four pheromone-baited traps in capturing male Helicoverpa (Lepidoptera: Noctuidae) moths in northern Florida. Florida Entomologist, 97, 1671–1678.
Hall, D. R., Farman, D., Domínguez, J. C., & Pajares, J. A. (2017). Female sex pheromone of the cone moth, Dioryctria mendacella: Investigation of synergism between type I and type II pheromone components. Journal of Chemical Ecology, 43(5), 433–442.
Harding, S. G., & Gladden, L. F. (1998). Diffusion of liquids into semicrystalline polyethylene. Journal of Magnetic Resonance Imaging, 16, 647–649.
Hasdenteufel, F. (2006). Twenty years of separation of cis-trans (Z)-(E) isomers. Separation and Purification Reviews, 35, 193–221.
Haynes, K. F., McLaughlin, J., Stamper, S., Rucker, C., Webster, F. X., Czokajlo, D., & Kirsch, P. (2007). Pheromone trap for the eastern tent caterpillar moth. Environmental Entomology, 36, 1199–1205.
Jabłoński, T. (2014). Barczatka sosnówka – Dendrolimus pini L. In Krótkoterminowa Prognoza Występowania Ważniejszych Szkodników i Chorób Infekcyjnych Drzew Leśnych w Polsce w 2014 r; Analizy i Raporty 22 (pp. 45–49). Instytut Badawczy Leśnictwa: Sękocin Stary, Poland.
Jactel, H., Menassieu, P., Vétillard, F., Barthélémy, B., Piou, D., Frérot, B., Rousselet, J., Goussard, F., Branco, M., & Battisti, A. (2006). Population monitoring of the pine processionary moth (Lepidoptera: Thaumetopoeidae) with pheromone-baited traps. Forest Ecology and Management, 235, 96–106.
Johansson, B. G., Anderbrant, O., & Sierpinski, A. (2002). Multispecies trapping of six pests of scots pine in Sweden and Poland. Journal of Applied Entomology, 126, 212–216.
Khrimian, A., Klun, J. A., Hijji, J., Baranchikov, J. N., Pet’ko, V. M., Mastro, V. C., & Kramer, M. H. (2002). Syntheses of (Z,E)-5,7-dodecadienol and (E,Z)-10,12-hexadecadienol, lepidoptera pheromone components, via zinc reduction of enyne precursors. test of pheromone efficacy against the Siberian moth. Journal of Agricultural and Food Chemistry, 50, 6366–6370.
Klun, J. A., Baranchikov, Y. N., Mastro, V. C., Hijji, Y., Nicholson, J., Ragenovich, I., & Vshivkova, T. A. (2000). A sex attractant for the Siberian moth Dendrolimus superans sibiricus (Lepidoptera: Lasiocampidae). Journal of Entomological Science, 35, 158–166.
Kong, X., Zhao, C., & Gao, W. (2001). Identification of sex pheromones of four economically important species in genus Dendrolimus. Chinese Science Bulletin, 46, 2077–2081.
Kong, X. B., Zhang, Z., Zhao, C. H., & Wang, H. B. (2007). Female sex pheromone of the Yunnan pine caterpillar moth Dendrolimus houi: first (E,Z)-isomers in pheromone components of Dendrolimus spp. Journal of Chemical Ecology, 33, 1316–1327.
Kong, X.-B., Sun, X.-L., Wang, H.-B., Zhang, Z., Zhao, C.-H., & Booij, K. C. J. H. (2011). Identification of components of the female sex pheromone of the Simao pine caterpillar moth, Dendrolimus kikuchii Matsumara. Journal of Chemical Ecology, 37, 412–419.
Kovalev, B. G., Bolgar, T. S., Zubov, P. A., Zharkov, D. G., Golosova, M., Nesterov, E. A., & Tvaradze, M. S. (1993). Identification of additional components of the sex pheromone of Dendrolimus pini. Chemistry of Natural Compounds, 29, 135–136.
Lance, D. (2006). Guidelines for detection trapping of exotic lymantriid and lasiocampid moths. United States Department of Agriculture.
Laurent, P., & Frérot, B. (2007). Monitoring of European corn borer with pheromone-baited traps: Review of trapping system basics and remaining problems. Journal of Economic Entomology, 100, 1797–1807.
Lenth, R. (2018). Emmeans: Estimated marginal means, aka least-squares means. R package version 1.3.1.
Li, X., Kong, X., Zhang, S., Wang, H., Zhang, Z., & Yang, M. (2015). Researches on the release rates of four types of insect semiochemicals from four dispenser types. Scientia Silvae Sinivae, 51, 63–70.
Makitra, R., Pyrih, Y., Sagladko, E., Turovskiy, A., & Zaikov, G. (2001). Generalization of polymer/organic solvent interaction processes by means of linear polyparameter equation. I. Swelling of polyethylene. Journal of Applied Polymer Science, 81, 3133–3140.
Mangiafico, S.S. (2016). Summary and analysis of extension program evaluation in R, version 1.15.0. rcompanion.org/handbook/. (Pdf version: rcompanion.org/documents/RHandbookProgramEvaluation.pdf).
McDonough, L. M., Brown, D. F., & Aller, W. C. (1989). Insect sex pheromone: Effect of temperature on evaporation rates of acetates from rubber septa. Journal of Chemical Ecology, 15, 779–790.
Moore, R., Cottrell, J., A’Hara, S., & Ray, D. (2017). Pine-tree lappet moth (Dendrolimus pini) in Scotland: Discovery, timber movement controls and assessment of risk. Scottish Forestry, 71, 34–43.
Ostrauskas, H., & Ivinskis, P. (2011). Moths caught in pheromone traps during search for Dendrolimus pini and D. sibiricus (Lepidoptera, Lasiocampidae) in Lithuania. Acta Zoologica Lituanica, 21, 238–243.
Pletnev, V. A., Ponomarev, V. L., Vendilo, N. V., Kurbatov, S. A., & Lebedeva, K. V. (2000). Search of the pheromone of Siberian silkworm Dendrolimus superans sibiricus (Lepidoptera: Lasiocampidae). Agrokhimiya, 6, 67–72.
Ponomarev, B. L., Baranchikov, Y. N., Marchenko, Y. I., & Ostrauskas, H. H. (2006). Field testing of Dendrolimus pini L. sex pheromone. Lesnoy Vestnik, 2, 145–151.
Priesner, E., Bogenschütz, H., Albert, R., Reed, D. W., & Chisholm, M. D. (1984). Identification and field evaluation of a sex pheromone of the European pine moth. Zeitschrift für Naturforschung C, 39, 1192–1195.
R Core Team (2018). R: A language and environment for statistical computing, v. 3.5.1. R Foundation for Statistical Computing, Vienna, Austria.
R Studio Team (2016). RStudio: Integrated development for R, v. 1.1.463. RStudio, Inc., Boston, MA.
Ray, D., Peace, A., Moore, R., Petr, M., Grieve, Y., Convery, C., & Ziesche, T. (2016). Improved prediction of the climate-driven outbreaks of Dendrolimus pini in Pinus sylvestris forests. Forestry, 89, 230–244.
Schmidt, B. C., & Roland, J. (2003). Developing techniques for monitoring forest tent caterpillar populations using synthetic pheromones. Canadian Entomologist, 135, 439–448.
Siegel, S., & Castellan Jr., N. J. (1988). Nonparametric statistics for the behavioral sciences (2nd ed.). New York, NY, England: McGraw-Hill Book Company.
Skaug, H., Fournier, D., Bolker, B., Magnusson, A., Nielsen, A. (2016). Generalized linear mixed models using 'AD model Builder'. R package version 0.8.3.3.
StatSoft Inc. (2011). STATISTICA (data analysis software system), version 10.
Stille, J. K., & Groh, B. L. (1987). Stereospecific cross-coupling of vinyl halides with vinyl tin reagents catalyzed by palladium. Journal of the American Chemical Society, 109, 813–817.
Stille, J. K., & Simpson, J. H. (1987). Stereospecific palladium-catalyzed coupling reactions of vinyl iodides with acetylenic tin reagents. Journal of the American Chemical Society, 109, 2138–2152.
USDA, APHIS, PPQ (2011). New Pest Response Guidelines: Dendrolimus Pine Moths. Washington, D.C. Last accessed September 1, 2019,http://www.aphis.usda.gov/import_export/plants/manuals/online_manuals.shtml.
Venables, W. N., & Ripley, B. D. (2002). Modern applied statistics with S (Fourth ed.). New York: Springer.
Wang, P., Chen, G.-F., Zhang, J.-S., Xue, Q., Zhang, J.-H., Chen, C., & Zhang, Q.-H. (2017). Pheromone-trapping the nun moth, Lymantria monacha (Lepidoptera: Lymantriidae) in Inner Mongolia, China. Insect Sci., 24, 631–639.
Zhang, A.-B., Wang, Z.-J., Tan, S.-J., & Li, D.-M. (2003). Monitoring the masson pine moth, Dendrolimus punctatus (Walker) (Lepidoptera: Lasiocampidae) with synthetic sex pheromone-baited traps in Qianshan County, China. Applied Entomology and Zoology, 38, 177–186.
Zhang, A., Leskey, T. C., Bergh, J. C., & Walgenbach, J. F. (2013). Sex pheromone dispenser type and trap design affect capture of dogwood borer. Journal of Chemical Ecology, 39, 390–397.
Acknowledgements
We would like to thank the staff of the Wolsztyn, Krucz and Człopa forest districts and the staff of the Forest Protection Service Units in Szczecinek and Łopuchówko for their support in the field experiments. We are grateful to two anonymous reviewers and Dr. Alejandro Solla from Extremadura University in Spain for valuable comments that allowed to improve the manuscript. This study was financially supported by the National Centre for Research and Development, Poland (Narodowe Centrum Badań i Rozwoju, Agreement No. PBS2/A9/25/2013).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies performed with human participants or animals by any of the authors.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sukovata, L., Asztemborska, M., Rudziński, K.J. et al. Effect of dispenser type, trap design and placement on catches of pine-tree lappet moth, Dendrolimus pini. Phytoparasitica 48, 63–74 (2020). https://doi.org/10.1007/s12600-019-00772-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12600-019-00772-9