
Abstract
The quarries located to the south of Visé, in southeastern Belgium, correspond to historical type area of the Viséan Stage (Carboniferous, Mississippian) and yielded an extremely rich macrofauna (known as the Visé fauna), including mainly brachiopods and mollusks (bivalves, gastropods, cephalopods), and also corals, trilobites and bryozoans, that are found in institutional collections all over the world. The Visé fauna is of late early Warnantian (late Asbian) age and was essentially recovered from a microbial buildup of Cracoean affinities that was exploited only in the quarries F and G sensu Horion and Gosselet’s (1892) terminology. Only the rugose corals have been revised, and the rest requires a complete taxonomic reassessment. Brachiopods are particularly diverse, and many genera (Productida, Orthida, Rhynchonellida, Terebratulida) were established in the past on the basis of type species from Visé. Productides are by far the most common elements of the brachiopod association, with the development of aberrant morphologies (Proboscidella, Striatifera). Some terebratulides and mollusks display ‘colour’ bandings that have been preserved. Two so-called Carboniferous species described by de Koninck (1887), namely Rhynchonella solitaris and Spirifer capillaris, are middle or late Frasnian in age.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Besides being the historical type area of the Viséan Stage (Hance et al. 2006), the disused quarries situated between Richelle and Souvré, to the south of Visé in southern Belgium (Fig. 1), rank among the most important localities for Carboniferous (Mississippian) palaeontology. Indeed, an incredibly rich macrofauna, known in the literature as the Visé fauna, was recovered from mostly upper Viséan limestones and described notably in a series of monographs mainly published during the second half of the nineteenth century (e.g. de Koninck 1842–1844, 1851, 1872, 1878, 1880, 1881, 1883a, 1885, 1887; de Ryckholt 1851; de Koninck and Le Hon 1854). Due to the inaccessibility of the quarries, it is impossible to collect new specimens nowadays, which is why the former palaeontological collections from Visé, which are curated in many institutions and universities, are of invaluable scientific value.

Tournaisian–Viséan sedimentation areas in the Namur–Dinant Basin (simplified from Poty 2016). Abbreviations: ASA South Avesnois sedimentation area; BVR Booze–Le-Val-Dieu ridge; CB Campine Basin; CSA Condroz sedimentation area; DSA Dinant sedimentation area; G. Germany; HSA Hainaut sedimentation area; N. the Netherlands, NSA Namur sedimentation area; L. Grand-Duchy of Luxembourg; VASA Vesdre–Aachen sedimentation area; VSA Visé–Maastricht sedimentation area
The geological context (e.g. lithostratigraphy, sedimentology, sequence stratigraphy, palaeogeography, palaeoecology, tectonics) of the Visé area was thoroughly documented in the past decades (e.g. Pirlet 1967; Poty 1982, 1991; Muchez and Peeters 1987; Aretz and Chevalier 2007; Poty and Delculée 2011). The most recent studies depicted a very complex history resulting from the conjunction between sea-level oscillations and block faulting that created the conditions favourable to the establishment of microbial buildups during the late early Warnantian (late Asbian). As explained below, this particular reefal environment allowed the development of most of the typical elements of the Visé fauna. The latter is characterised by the profusion of brachiopods, bivalves, gastropods and many other organisms (e.g. cephalopods, echinoids, trilobites); many species are represented by a plethora of specimens, ranging from juveniles to adults, and thus allowing to perform statistics. Due to the incredible richness in fossils (see references above), which are generally relatively well preserved, the quarries of Visé allow to have a broad view of the biodiversity prevailing in microbial reef environments during the late Viséan. For all these reasons and even if soft parts are not preserved, Visé should be considered a conservation Lagerstätte, the second one recognised within the Carboniferous marine succession of Belgium after the Viséan (Moliniacian) ‘black marble’ of Denée (Mottequin 2008).
Contrary to purely geological studies, data specifically related to the brachiopods from Visé are dispersed in a great number of papers but, until now, no overview has been available. Moreover, the re-investigation of the brachiopod faunas from Visé remains long overdue. Although such a revision is well beyond the scope of this paper, it aims at providing (1) detailed information related to the age and origin of the Visé fauna and (2) the first overview of the brachiopods from the historical type area of the Viséan Stage.
Geological context
The small Visé area (Fig. 2) experienced a complex geological history during the Devonian and the Carboniferous as reflected by the numerous studies and models that were proposed to explain the close contact between Upper Devonian (Frasnian) and Mississippian limestones observed in the quarries situated to the south of Visé (e.g. Pirlet 1967; Poty 1982, 1991; Poty and Delculée 2011). It soon appeared that the limestones exposed in the Visé area were partly Devonian as reflected by the presence of some characteristic brachiopod and gastropod species reported by de Koninck (e.g. 1843 in 1842–1844) (see de Ryckholt 1851; Horion 1860, 1863; Gosselet 1861).

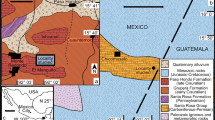
The Visé area is part of the Visé–Maastricht sedimentation area (VSA) (e.g. Poty 1997, 2016) that is located on the easternmost margin of the Brabant Massif (Figs. 1 and 3); Frasnian–Mississippian rocks crop out only in the southern part of the VSA, thus around Visé. According to Poty and Delculée (2011), the VSA is apparently a small pull-apart basin resulting from a strike slip fault related to two north and south half-graben structures, known respectively as the Campine Basin (CB) and the Namur sedimentation area (NSA) (Fig. 1), and was affected by block faulting during Devonian and Carboniferous times (Poty 1991). The VSA has been separated from the NSA from the Ivorian until, at least, the end of the Warnantian by the Booze–Le-Val-Dieu ridge (Poty 1997, 2016) and open to the CB whereas its northern part became a graben (Maastricht Graben) in which carbonate turbidites and debris flows were recognised in boreholes (Poty and Delculée 2011) (Figs. 1 and 3). The southern border of the VSA was subdivided into three different slowly subsiding to rising blocks (Poty 1991; Poty and Delculée 2011) that are each characterised by their own succession (with more or less incomplete sedimentological record), from west to east: the Hermalle-sous-Argenteau, Souvré and Bombaye blocks (Fig. 3). They are bordered to the south by the Booze–Le-Val-Dieu ridge (Figs. 1 and 3) and to the north by the Maastricht block system. The quarries that yielded the typical Visé fauna are located on the eastern margin of the Hermalle-sous-Argenteau block (HSAB), near its contact with the Souvré block, and are known as the quarries F and G following Horion and Gosselet’s (1892) lettering (Fig. 2).

Location map (a) and relative position of the tectonic units (b) on the eastern end of the Brabant Massif during the Late Devonian–Carboniferous with localization of the former quarries F–G following Horion and Gosselet’s (1892) terminology (modified from Poty and Delculée 2011). The reconstruction (b) corresponds to the situation during the late Viséan. The different blocks (Hermalle-sous-Argenteau, Souvré (SB), Bombaye) correspond to the southern border of the Visé–Maastricht sedimentation area (VSA). They were separated from the Namur–Dinant Basin by the Booze–Le-Val-Dieu ridge, and this, without any connection. Blocks situated northwards (Maastricht block system) were highly subsident and formed the Maastricht Graben. They are not differentiated and are suggested on the figure by dashed lines
From the lithostratigraphical viewpoint, the upper Asbian limestone of the disused quarries F and G belongs to the Visé Formation, and the overlying silicified beds (silicified laminar shales and limestones) to the Souvré Formation (Figs. 4 and 5). For its practical use in geological mapping, the Visé Formation was extended to all the pale and grey limestones present in the VSA; therefore, it comprises different facies and different ages ranging from the late Tournaisian to the top of the lower Warnantian (late Asbian) with numerous stratigraphic gaps (see Poty et al. (2002) and Poty and Delculée (2011) for more details). The following distinct facies are recognised within the Visé Formation in the eponymous area: (1) carbonate breccias with centimetric to pluridecametric boulders of Frasnian age; (2) thick-bedded packstones to rudstones, with sedimentary breccias, forming fining-upwards pluridecimetric to metric sequences often laminated in their upper part; (3) thick-bedded to massive packstones to rudstones; and (4) massive algal and bioclastic boundstones forming buildups (Muchez and Peeters 1987) rich in macrofaunas (mainly brachiopods) (Poty et al. 2002). Because of their small thickness (some metres) and their presence only in the quarry F, the thin dolomitic beds of late Warnantian (Brigantian) age resting between the Visé limestone and the Souvré silicified beds were neglected in the lithostratigraphical succession.

Biostratigraphy and lithostratigraphy of the Devonian and Carboniferous deposits recorded in the Visé–Maastricht sedimentation area (modified from Poty and Delculée 2011), and more precisely in the Hermalle-sous-Argenteau (HSA) block where are located the quarries F–G (HAS NE) situated to the south of Visé. Abbreviations: Ar. Arnsbergian; G. Givetian; Fa. Famennian; Fr. Frasnian; L. Lower; Lithostrat. Lithostratigraphy; M. Middle; MFZ Mississippian foraminifer zones (Devuyst and Hance in Poty et al. 2006); P. Pendleian; RCZ Rugose coral zones (Poty in Poty et al. 2006); U. Upper

View of the face of the disused Horion and Gosselet’s (1892) quarry F, south of Visé along the right bank of the Meuse River (NE of Hermalle-sous-Argenteau block) (adapted from Poty and Delculée 2011). The upper Warnantian (corresponding to the British Brigantian) and the Serpukhovian (Namurian according to the traditional subdivision) strata form two bevels, each of them overlying a palaeokarstic surface
The Visé fauna
General remarks
Most of the fossils labelled ‘Visé’ that are encountered in the old collections dispersed in many institutions worldwide (e.g. Natural History Museum (London), Carnegie Museum (Pittsburgh), Harvard Museum of Comparative Zoology (Cambridge, Massachusetts), Museum d’Histoire naturelle (Paris), Lyon 1 University, Royal Belgian Institute of Natural Sciences (Brussels), Smithsonian National Museum of Natural History (Washington D.C.), University of Liège), thanks to exchanges, donations and purchases, are from the quarries once open for the lime production (Delépine 1921) along the Meuse River, between Souvré, to the north and Richelle, to the south (Fig. 2). As indicated by Delépine (1940), the former collections from Visé were gathered without taking care about their exact location and their stratigraphic position in the quarries. Horion and Gosselet (1892) noted that the fossils were difficult to collect and those that were bought to the quarrymen included both Frasnian and Carboniferous taxa.
Provenance and age
The quarries F, G and H (following Horion and Gosselet’s (1892) lettering) (Figs. 2 and 5), also known in the literature as the Andrien’s lime kiln quarry (Horion and Gosselet 1892; Delépine 1911), are the most significant ones for the study of the Visé fauna. According to Demanet (1958), it seems that the massive limestones (reefal level; see Muchez and Peeters 1987) of late early Warnantian (late Asbian) age (RC7β–MFZ14 sensu Poty et al.’s (2006) biozonations) of the Visé Formation (former ‘V3b’) from the quarry G (following Horion and Gosselet’s (1892) lettering) yielded the most important part of the invertebrates described and illustrated in de Koninck’s monographs (see references above). Nevertheless, Pirlet (1967) noted that the quarry F, already disused in the years 1890 (Horion and Gosselet 1892), was the greatest fossil supplier whereas Delépine (1910) reported that the quarry G yielded most of the material at the onset of the twentieth century. We can thus reasonably presume that the fossils described by de Koninck (1842–1844) in his first monograph were collected in the quarry F, and additional ones were then collected in the quarry G, but in the same carbonate buildup and level. Both quarries were poorly developed during the nineteenth century and only the quarry F has kept its original appearance (Fig. 5). Its current face illustrates well what should be that of the quarry G at the beginning of its exploitation. Note that the top of the microbial buildup was locally altered and provided specimens free of matrix and also internal moulds. Demanet (1958) noted that another important part of the RBINS collections comes from the Souvré Formation (‘couches schistoïdes V3c’) of the quarry G on the basis of their state of preservation, their delicacy and their tint. They come from the upper levels that are generally altered and of Brigantian age; many of these specimens are preserved as internal moulds.
The quarry G was also supposed to display the buildup horizon at the onset of its exploitation, but its eastward development has resulted in the limestones now exposed being much less rich. Fossils were collected likewise in the fine-grained, altered dolostones and the silicified beds overlying the buildup horizon.
The buildup horizon rests on early Warnantian breccias with Frasnian limestone blocks, sometimes of cyclopean size. These breccias overlie Tournaisian (upper Ivorian, RC4β1, MFZ8) limestones that are lateral time-equivalent to the oolitic limestones of the Avins Member (Longpré Formation; Avins event (Poty 2007), Fig. 4) but, as they do not crop out in the quarries F and G, they have thus not yielded fossils. Note that before Poty and Delculée’s (2011) overview, they were considered of early Viséan age (‘V1a’).
The Frasnian material is supposed to have been recovered from the blocks included in the breccia levels occurring in the Visé Formation (see above).
In conclusion, the classical Visé fauna sensu L.-G. de Koninck is from both the Visé Formation (e.g. massive limestone rich in productidines) and the Souvré Formation (silicified shales and limestones), and, thus, includes Viséan elements, considering the rocks overlying the reefal levels as Brigantian in age, although part of the Souvré Formation may be of Serpukovian age.
Brief review of the faunas
Besides the brachiopods that are documented below, it is worthwhile to review briefly some significant elements of the Visé macrofauna (except fishes (de Koninck 1878; Derycke et al. 1995)). Mollusks are particularly abundant and diverse as reflected by the long lists of species that flourished in the literature dating back to the nineteenth century (de Koninck 1842–1844, 1878, 1880, 1881, 1883a, 1885; Dewalque 1868; Mourlon 1881). Those provided by de Koninck deserve some discussion although they badly need to be re-assessed in the light of the most recent advances in taxonomy. Furthermore, the number of species is certainly overestimated as this palaeontologist was a splitter. On the 222 bivalve and rostroconch species (all were included under the name ‘lamellibranches’) reported by de Koninck (1885) from the étage de Visé in southern Belgium, most of them are from Visé. A similar statement is also valid for the 283 species of ‘gastropods’ recognised by this author (de Koninck 1881, 1883a) in the étage de Visé. Note that the term ‘gastropods’ sensu de Koninck likewise included polyplacophorans (Hoare 2002), scaphopods (most probably tube worms according to Godefroid et al. (2006)) and conulariids. In addition, mollusks include goniatites (Delépine 1940) and nautiloids (de Koninck 1878, 1880; Maillieux 1925). Among the bivalves, the occurrence of large pinnid representatives (reaching up to 35 cm in length; de Koninck 1885: pl. 25, figs 1–2) is significant because Visé is the only Belgian locality where gigantic brachiopods (see below) and bivalves developed together due to favourable conditions. From the echinoderm viewpoint, crinoids were described by de Koninck and Le Hon (1854) whereas Jackson (1929) documented the echinoids. Hahn et al. (1988) reported nine trilobite species from the upper Warnantian of Visé. Rugose corals were revised by Poty (1981) and some tabulates and chaetetids were illustrated by Tourneur et al. (1989). Bryozoans were illustrated by de Koninck (1842–1844) but remain unrevised.
Material and methods
The bulk of the illustrated material is stored at the Royal Belgian Institute of Natural Sciences (Brussels; prefixed RBINS) and is complemented by specimens curated at the University of Liège (prefixed PA.ULg). A shell selected for scanning electron microscopy was observed using a low-vacuum SEM, an ESEM FEI Quanta 200, but not coated with gold. The larger specimens were coated with ammonium chloride sublimate before being photographed, whereas those displaying ‘colour’ bandings were coated with glycerine or ethanol.
Most of the specimens illustrated here were previously illustrated by drawings (de Koninck 1883a, 1885, 1887) or poor-quality photographs (Demanet 1934). The references to these original illustrations are clearly indicated in the figure legends.
Brachiopods from Visé
History of research
Brachiopods from Visé were documented in a series of monographs by de Koninck (1843 in 1842–1844, 1847a, 1847b, 1851, 1859, 1883b, 1887), who also discussed some former references that may include material from Visé (e.g. Chemnitz 1784) (see de Koninck 1847a). De Koninck died in 1887 while he was working on his last monograph specifically dedicated to the rhynchonellides, terebratulides and spire-bearers (athyridides, spiriferides, spiriferinides). This work was published the same year and includes several plates (de Koninck 1887: pls. 32–37) illustrating numerous unidentified spiriferides and spiriferinides that were ready for publication, but devoid of explanatory text (Demanet 1934). Some of these figured fossils are from Visé (Vandercammen and Vandercammen 1970) (Fig. 10(k)).
Other contributions (descriptions and/or illustrations) were published by Defrance (1826), Hoeninghaus (1828), Davreux (1833), de Verneuil (1840, 1841), Goldfuss (1840), d’Orbigny (1849), de Ryckholt (1851), Quenstedt (1852, 1871 in 1868–1871, 1885) and Fraipont (1888). During the nineteenth century, lists of brachiopod species were provided by Dumont (1832), and most comprehensive ones by Dewalque (1868), Mourlon (1881) and de Koninck (1887). The first half of the twentieth century was particularly poor in studies dedicated to the brachiopods from Visé, except Demanet’s (1931, 1934) papers in which linguliforms, orthotetides, orthides and spiriferinides were described and/or illustrated photographically for the first time. Nonetheless, Gröber (1908), Thomas (1914), Muir-Wood (1928) and Paeckelmann (1930) notably discussed species reported by de Koninck from this area. The second half of the twentieth century is marked by the first photographic illustrations of several de Koninck’s productide species and their revision by Muir-Wood and Cooper (1960), who erected several genera based on type species from Visé (Table 1). This significant work was complemented by the contributions of Stehli (1961), Muir-Wood (1962), Ramsbottom (1963), Grant (1965), Brunton (1971, 1979, 1982a, 1984), Brunton and Mundy (e.g. 1983, 1986, 1988, 1993) and Brunton et al. (1994), and more recently by those of Alvarez and Brunton (2000), Mottequin and Simon (2017a, 2017b) and Mottequin (2021).
Brachiopod succession
The past: Horion (1863) noted that the Carboniferous Limestone of Visé could be subdivided palaeontologically into a lower stage (close of the Tournai Limestone) and an upper one which represents the main part of the Visé Limestone. However, both stages correspond to Viséan rocks. He recognised the following species in the lower stage (the original generic assignments are maintained herein for historical reasons): Productus sublaevis, Chonetes comoides, Spirifer cuspidatus—a Syringothyris species described by Sowerby (1816 in 1815–1818) (not Spirifer aperturatus von Schlotheim var. cuspidata d’Archiac and de Verneuil 1842 (see Paeckelmann 1942))—S. convolutus and Rhynchonella acuminata. The latter species is present in the upper stage but only as isolated specimens according to him. The upper stage, also called upper limestone, is characterised by the abundance of Productus striatus, P. cora, P. giganteus and P. punctatus. Horion (1863) recognised at least three horizons within the upper stage based on the productidines (Fig. 6). He noted that the middle horizon is marked by the predominance of the small species and by the overabundance of P. punctatus while the upper horizon is dominated by the largest ones. According to Horion (1863), the rocks overlaying the limestones are marked by the progressive disappearance of the taxa recognised below and the progressive appearance of species typical of the lower ‘Houiller’ (Namurian according to the traditional subdivision). Horion and Gosselet (1892) did not modify significantly Horion’s (1863) statement (Fig. 6), but they distinctly indicated a late Viséan age for the Carboniferous fossils from Visé. Delépine (1910, 1911) reported that the brachiopods are abundant in the limestones that were exploited in the middle part of the succession exposed in the quarry G. He recognised the following species in these limestones of the Visé Formation: Productus giganteus, P. striatus, P. punctatus, P. latissimus, P. semireticulatus, P. martini, P. scabriculus, P. undatus, Spirifer striatus, S. bisulcatus, S. grandicostatus, etc.

Delépine (1922) identified brachiopods from the limestones of the uppermost part of the Visé Formation that were collected in 1886 by X. Stainier in an outcrop located near the Argenteau bridge, most probably in the hillslope near Richelle (Fig. 2). The following species (or genera) were identified: Productus semireticulatus, P. mesolobus, P. undatus, P. punctatus, P. pyxidiformis, P. fimbriatus, P. spinulosus, P. longispinus, P. aculeatus, P. deshayesianus, P. proboscideus, Chonetes buchiana, Orthotetes, Strophomena, Schizophoria resupinata, Spiriferina octoplicata, Spirifer triangularis, S. ovalis.
Delépine (1921) provided significant data on the brachiopods at the contact between the Visé and Souvré formations at the quarry F (Figs. 2 and 5). He provided a list of brachiopods from the base of the Souvré Formation in which he reported, besides athyridides, Productus latissimus, a variety of Productus striatiferus as well as species and varieties of the Spirifer bisulcatus group. Unfortunately, the material was not illustrated.
In conclusion, the literature data clearly indicate that the productidines are the most common brachiopods collected at Visé as noted by Forir and Destinez (1901) and as reflected by the collections curated in Belgium. Although the other brachiopod orders are represented (see below), they failed to make impression among the different generations of geologists who had the opportunity to collect material in the whole succession, probably because of their lesser abundance.
Additional comments: As indicated by Poty and Delculée (2011), the oldest rocks recognised in the HSAB are of latest Ivorian to early Moliniacian age (see above) but they do not correspond to those ascribed to the lower stage by Horion (1863) and Horion and Gosselet (1892). Among the brachiopods cited by these authors, noticeable is their report of Levitusia sublaevis from these rocks. Do they correspond to the specimens reported as L. humerosa by Pirlet (1967) in the breccias observed in quarries K and L? It is impossible to confirm the identification of the latter author due to the lack of illustration. Firstly, the range of the different species of Levitusia within the Tournaisian–Viséan succession of Belgium is still unclear. Indeed, the re-assessment of Delépine’s (1926) study on Levitusia representatives from the uppermost Ivorian oolitic facies (Avins Member) of the Namur–Dinant Basin is needed in regards of investigations of Brunton (1979, 1982a), who exposed the differences between L. humerosa (and its junior synonym L. christiani) and L. sublaevis (with Visé as type locality). Secondly, Delépine (1926) indicated that at Visé, where the first specimens identified as L. sublaevis by de Koninck (1843 in 1842–1844) were collected, the specimens come from limestones of late Viséan age. The occurrence of Delepinea comoides (Horion 1863) would be also significant, if it is confirmed, as it is one of the most common species in the lower Moliniacian succession of southern Belgium according to Demanet (1958). Nevertheless, the brachiopod succession remains poorly constrained, in both the Ivorian–Moliniacian and Warnantian parts of the Visé Formation, but it is clear that numerous species (e.g. Proboscidella proboscidea, Striatifera striata) occurring in the Warnantian microbial reefal facies seem to be restricted to the Visé area in southern Belgium, which was separated from the Namur–Dinant by the Booze–Le-Val-Dieu ridge at that time.
Brachiopod diversity
The aim here is to briefly discuss and illustrate some specimens of each order present at Visé. Due to a lot of taxonomic issues that will only be resolved by investigation of the rich brachiopod faunas from the Carboniferous of Ireland and the UK described in the pioneering monographs (see references in Brunton 1984) and the revision of de Koninck’s taxa, it is not reasonable to provide here a number of species for each order. Simply to provide a guide, in compiling the data published by Mourlon (1881), de Koninck (1887) and Demanet (1934), c. 125 species were reported at Visé but this number is, in all likelihood, overestimated. Seventeen genera were established on the basis of type species first described in Visé, mainly productidines with nine genera (Table 1).
On the evidence of plates published by de Koninck during his long career, the fauna from Visé seems to be exquisitely preserved, but it is far from being always the case. de Koninck’s (e.g. 1887) drawings of brachiopods from this locality and from other Belgian ones (e.g. Tournai) were generally idealised and reconstructed, sometimes in a significant way, as already noted by Mottequin and Simon (2017a, 2017b), but it is also evident for the other groups investigated by this palaeontologist (e.g. Godefroid et al. 2006). Consequently, the so-called tremendous preservation depicted by these drawings is generally far from the real state of preservation (compare, e.g. de Koninck (1885, pl. 32, fig. 21; Fig. 7a) and Fig. 8(a), and de Koninck (1887, pl. 27, fig. 23; Fig. 7b) and Fig. 11(i)). Material from Visé is frequently incomplete, distorted and decorticated. Despite this pitfall, this locality yielded one of the most impressive Mississippian faunas worldwide. Moreover, alongside Tournai and Waulsort (Fig. 1), Visé is one of the rare Belgian localities where ‘colour’ bandings have been preserved (cephalopods, bivalves (Fig. 8(a)), gastropods (Fig. 8(b–c))) and the only one for brachiopods (de Koninck 1887, pl. 7, figs. 39–40; Fig. 8(d–e)).

a Original de Koninck’s (1885) illustration of ‘Entolium’ coloratum de Koninck 1885, incomplete left valve (RBINS a4952, syntype; de Koninck 1885, pl. 32, fig. 21; compare with Fig. 8a). b Original de Koninck’s (1887) illustration of Spirifer grandicostatus (M‘Coy 1852) (= choristitid? gen. et sp. indet.), articulated specimen in ventral view (RBINS a5590; de Koninck 1887, pl. 27, fig. 23; compare with Fig. 11i)

Examples of bivalve (a), gastropod (b–c) and brachiopod (d–e) with ‘colour’ bandings from the Mississippian of southern Belgium. a ‘Entolium’ coloratum de Koninck 1885, incomplete left valve (RBINS a4952, syntype; de Koninck 1885, pl. 32, fig. 21) in plan view (coated with glycerine), Waulsort Formation (Ivorian), Waulsort (Les Pauquys). b–c Waagenella cf. rownhamensis Weir 1931, complete, distorted shell (RBINS a4120; de Koninck 1883a, pl. 38, figs. 11–14) in umbilical and oblique dorsal views, Visé Formation (Warnantian), Visé. d–e Beecheria radiata (de Koninck 1887), Visé Formation (Warnantian), Visé. d Articulated specimen (RBINS a5106, syntype: de Koninck 1887, pl. 7, figs. 39–40) in ventral view (without and with ethanol coating). e Dorsal valve (RBINS 13463) in plan view
Although it is currently impossible to propose an overview of all the numerous species present in Visé, we provide below a brief survey arranged order by order.
Lingulides: De Koninck (1843 in 1842–1844) mentioned Lingula mytilloides at Visé in contrast to Demanet (1934) who did not report any lingulid there. Discinids are still poorly, but a little better, documented. Noticeable is the presence of large representatives of the genus Lindstroemella (Fig. 9(a–d)) that were specifically identified as L. glebosa by Demanet (1934); however, their general shape is markedly different from de Ryckholt’s (1854) Helcion glebosa, which remains known only by a few drawings. De Ryckholt (1851) described Orbiculoidea dumontiana from the Visé limestone, another poorly known Palaeozoic brachiopod species described by this author (see Mottequin 2019 for the Devonian ones), which was overlooked by Demanet (1934). The latter author just cited O. nitida at Visé (Fig. 9(e)). The species Lingula konincki was introduced by Fraipont (1888) on the basis of an inadequately preserved specimen (not yet traced in the palaeontological collections of the University of Liège), which was considered by Graham (1972) as a possible synonym of Orbiculoidea craigii, but of course, this statement cannot be confirmed currently.

Lingulides (a–e), craniides (f–g), strophomenides (h–i), chonetidines (j) and productidines (k–q) from the Visé Formation (Warnantian), Visé. a–d Lindstroemella sp. indet. a Ventral valve (RBINS a5821; Demanet 1934, pl. 1, fig. 22) in plan view. b Ventral valve (RBINS a13464) in plan view. c–d Dorsal valve partly decorticated (RBINS a5820; Demanet 1934, pl. 1, fig. 20) in plan view and detail of muscle field. e Orbiculoidea sp. indet., poorly preserved dorsal valve (RBINS a13465) in plan view. f–g Petrocrania? sp. indet. (fixed on ventral valve of Acanthoplecta sp. indet.), internal moulds of dorsal valves (RBINS a13466) and enlargement of one of them. h Leptagonia cf. caledonica Brand 1972, incomplete articulated specimen in ventral (perpendicular to the disc) view (RBINS a5915; Demanet 1934, pl. 6, figs. 7, 7a; Mottequin and Simon 2017a, fig. 9.5–9). i Pseudoleptaena distorta (J. de C. Sowerby 1840 in 1840–1846), incomplete articulated specimen (PA.ULg 2020.09.30/1) in ventral (perpendicular to the disc) view. j Plicochonetes buchianus (de Koninck 1843 in 1842–1844), internal mould (RBINS a13467) in ventral view. k Overtonia fimbriata (J. de C. Sowerby 1824 in 1823–1825), complete articulated specimen (RBINS a13468) in ventral view. l Productus productus (Martin 1809), ventral valve (RBINS a13469) in plano-anterior view. m Pustula sp. indet., articulated specimen (RBINS a13470) in ventral view. n Echinoconchus punctatus (J. Sowerby 1822 in 1821–1822), articulated specimen (RBINS a13471) in ventral view. o Striatifera striata (Fischer de Waldheim 1837), two ventral valves (RBINS a13472) in plan view. q Proboscidella proboscidea (de Verneuil 1840), almost complete specimen (RBINS a13473) in ventral view
Craniides: De Koninck (1843 in 1842–1844) erected the species Orbicula? concentrica based on material from Visé (non Orbicula concentrica von Buch 1832, a bivalve species from the Frasnian of the northeastern Rhenish Massif ascribed to the genus Ontaria Clarke 1904 (see references in Nagel 2006)). Although it is only documented by the original illustrations, its ornamentation, including both growth lines and capillae, brings it closer to Deliella? trigonalis as illustrated by Brunton (1968). Few Petrocrania? (Fig. 9(f–g)) specimens have been recovered so far.
Strophomenides: Besides Leptagonia cf. caledonica (Fig. 9(h)) recognised by Mottequin and Simon (2017a) at Visé, de Koninck (1843 in 1842–1844), Demanet (1934) and the former authors reported the presence of the atypical Pseudoleptaena distorta (Fig. 9(i)), which is easily recognisable thanks to its fluting trail. They are the youngest strophomenides recognised in the Carboniferous of Belgium (see discussion in Mottequin and Simon 2017a).
Chonetidines (productides): They are not particularly diverse as only two genera are recognised with certainty, namely Plicochonetes (e.g. P. buchianus) (Table 1; Fig. 9(j)) and Megachonetes (e.g. M. dalmnanianus, M. zimmermani) according to Paeckelmann (1931) and Muir-Wood (1962)). It is highly probable that Delépine’s (1921) report of Daviesiella llangollensis (a possible productidine) from the topmost part of the Visé Formation in the Souvré quarry corresponds to a representative of the genus Globosoproductus.
Productidines (productides): This suborder, alongside the strophalesiidines, includes some of the most emblematic species recognised at Visé (e.g. Muir-Wood and Cooper 1960), which are represented by exquisitely preserved specimens such as Productus productus with long trails (Fig. 9(l)) and others (Fig. 9(k, n)). Productidines are small (Limbifera) to large (Pustula (Fig. 9(m)), Latiproductus (Fig. 10(b)), or even gigantic (Gigantoproductus, Fig. 10(a)). They display an incredible range of morphologies such as the Pinna-shaped Striatifera striata (Fig. 9(o)), the amazing tube-like Proboscidella proboscidea (Fig. 9(q)), and Semenewia concentrica with its unusual strong concentric ornamentation (Brunton et al. 1994).

Productides (a–b), orthotetides (c–d), orthides (e–l) and rhynchonellides (m–q) from the Visé Formation (Warnantian), Visé. a Gigantoproductus sp. indet., external dorsal mould with shelly fragments (RBINS a13474) in plan view. b Latiproductus sp. indet., almost complete ventral valve (RBINS a13475) in plan view. c Schellwienella ornata Demanet 1934, articulated internal mould (RBINS a5922, lectotype; Demanet 1934, pl. 6, fig. 4; Mottequin and Simon 2017a, fig. 15.1–8) in ventral view. d Streptorhynchus anomalus (J. de C. Sowerby 1840 in 1840–1846), internal mould (RBINS a5955; Demanet 1934, pl. 10, fig. 5, 5a–c; Ramsbottom 1963, fig. 1, pl. 3, figs. 4–6) in ventral view. e Aulacophoria keyserlingiana (de Koninck 1843 in 1842–1844), articulated specimen (RBINS a5890; Demanet 1934, pl. 4, fig. 14, 14a–b) in ventral view. f Rhipidomella lyelliana (de Koninck 1851), articulated specimen (RBINS a5872; Demanet 1934, pl. 3, fig. 17) in ventral view. g–h Schizophoria (Schizophoria) connivens (Philips, 1836). g Juvenile specimen (RBINS a5881; Demanet 1934, pl. 4, fig. 5, 5b) in ventral view. h Adult specimen (RBINS a13476) in ventral view. i Schizophoria (Schizophoria) resupinata pinguis Demanet 1934, articulated specimen (RBINS a5885; Demanet 1934, pl. 3, fig. 9, 9a; Mottequin 2021, figs. 14E, 17F–J) in ventral view. j Schizophoria (Schizophoria) resupinata gigantea Demanet 1934, incomplete articulated specimen (RBINS a5888; Demanet 1934, pl. 4, fig. 12; Mottequin 2021, fig. 15K–O) in ventral view. k Schizophoria (Paraschizophoria) woodi Bond, 1941, articulated specimen (RBINS a5877; Demanet 1934, pl. 4, fig. 1, 1a–1b) in ventral view. l Schizophoria (Portlockia) gibbera (Portlock 1843), articulated specimen (RBINS a5880; Demanet 1934, pl. 4, fig. 4, 4a) in ventral view. m–n Pugnax acuminatus (Sowerby 1822 in 1821–1822). m Articulated specimen (RBINS a5190; de Koninck 1887, pl. 9, fig. 5) in ventral view. n Large articulated specimen (RBINS a5189; de Koninck 1887, pl. 9, figs. 1–4) in anterior view. o Pleuropugnoides pleurodon (Philips 1836), distorted articulated specimen (RBINS a5363; de Koninck 1887, pl. 15, figs. 1–4) in ventral view. p Coledium obscurum (de Koninck 1887), articulated specimen (RBINS a5327, syntype; de Koninck 1887: pl. 16, figs. 30–33) in ventral view. qTretorhynchia trilatera (de Koninck 1843 in 1842–1844), articulated specimen (RBINS a5358, neotype; de Koninck 1887, pl. 16, figs. 69–75; Brunton 1971, pl. 11, figs. 1–4) in ventral view
Strophalesiidines (productides): Visé is the type locality of the type species of five genera (Table 1) among which the curious Sinuatella (Brunton and Mundy 1988), which looks like the rafinesquinid Leptagonia, and was sometimes confused with the latter (see discussion in Mottequin and Simon 2017a). Besides these genera, Brunton and Mundy (1988) reported the presence of Retroplexus representatives, perhaps belonging to R. cravenensis.
Orthotetides: This order was discussed by Demanet (1934), Ramsbottom (1963) and Mottequin and Simon (2017a). Seven species were recognised by both former authors, belonging notably to Schellwienella (Fig. 10(c)) and Streptorhynchus (Fig. 10(d)). Visé yielded the largest orthotetides (more than 130 mm in width) known from the Mississippian of Belgium (Demanet 1934). Shells with strong asymmetrically folded anterior commissure are observed in Streptorhynchus anomalus (Fig. 10(d)).
Orthides: Besides the rare Aulacophoria keyserlingiana (Fig. 10(e)) and the long-ranging genus Rhipidomella (R. lyelliana (Fig. 10(f))), the Visé fauna includes small to large, rounded to more transverse representatives of the genus Schizophoria (S.) (Demanet 1934; Mottequin 2021) (Fig. 10(g–j)), which need to be re-investigated. Furthermore, the strongly inflated species S. (Paraschizophoria) woodi (Fig. 10(k)) and S. (Pocockia) gibbera (Fig. 10(l)) (Demanet 1934; Bond 1941; Pocock 1965) are recognised at Visé.
Rhynchonellides: De Koninck (1887) reported 19 species, including Rhynchonella reflexa, which was doubtfully assigned to the terebratulide genus Gacina by Stehli (1961), but tentatively ascribed to Cryptacanthia by Mottequin (2021). Besides costate species (e.g. Pleuropugnoides pleurodon (Fig. 10(o))), a large range of morphology and size can be pointed out among the representatives of this order, from the large and almost smooth Pugnax acuminatus (Fig. 10(m–n)) to the small and nearly smooth Coledium sp. (e.g. Grant 1965) (Fig. 10(p)) or small and costate Tretrorhynchia trilatera (see Brunton 1971) (Fig. 10(q)), in passing by large and strongly costate Yanishewskiella angulata.
Athyridides: Already discussed by de Verneuil (1840), athyridides were described by de Koninck (1887) and partly revised by Alvarez and Brunton (2000) and Mottequin and Simon (2017a). The following genera have been recognised until now: Actinoconchus (Fig. 11(a–b)), Composita, Coveenia, Hustedia and Lamellosathyris (Fig. 11(c)). Some rare specimens of Actinoconchus paradoxus display their long, radially striated flat flanges (de Koninck 1887, pl. 21, figs. 17 (Fig. 11(a–b)), 18–19). One of them (Fig. 10(b)) shows the distal extremity of the flange. This delicate structure seems to be strengthened by the presence of spines, rectangular in section, that extend from the lateral and anterior commissures of the shell.

Athyridides (a–c), spiriferides (d–k), spiriferinides (l–n), and terebratulides (o–p) from the Visé Formation (Warnantian), Visé. a–b Actinoconchus paradoxus M‘Coy, 1844, ventral valve with flange preserved (RBINS a5490; de Koninck 1887, pl. 21, fig. 17) in plan view (with internal mould of a chonetidine) and detail of the distal part of the flange. c Lamellosathyris lamellosa (Léveillé, 1835), incomplete, distorted specimen (RBINS a5471; de Koninck 1887, pl. 21, figs. 9–10; Mottequin and Simon 2017a, fig. 23.1–5). d Podtsheremia sp. indet., articulated, slightly distorted specimen (RBINS a5651; de Koninck 1887, pl. 30, figs. 4–6) in ventral view. e Spirifer (Spirifer) striatus (Martin 1793), incomplete, distorted and partly decorticated specimen (RBINS a4298; de Koninck 1883, pl. 15, figs. 1–2, de Koninck 1887, pl. 23, figs. 1–2) in ventral view. f–g Strophopleurid gen. et sp. indet. f Incomplete, distorted specimen (RBINS a5613; de Koninck 1887, pl. 29, figs. 7–9) in ventral view. g) Incomplete specimen (RBINS a5614; de Koninck 1887, pl. 29, figs. 10–12) in dorsal view. h) Brachythyris sp. indet., articulated specimen (RBINS a5632; de Koninck 1887, pl. 30, figs. 8–11). i Choristitid? gen. et sp. indet., articulated specimen (RBINS a5590; de Koninck 1887, pl. 27, figs. 22–24). j) ‘Verneuilia’ oceani (d’Orbigny 1849), articulated, slightly distorted specimen (RBINS a5629; de Koninck 1887, pl. 28, fig. 14). k) Reticulariid gen. et sp. indet., slightly distorted, articulated specimen (RBINS a5739; de Koninck 1887, pl. 37, figs. 42–45) in ventral view. l Davidsonina septosa transversa Jackson 1922, distorted, articulated specimen (RBINS a5676; de Koninck 1887, pl. 33, figs. 1–3) in ventral view. m Spiriferellina sp. indet., distorted, articulated specimen (RBINS a5518; de Koninck 1887, pl. 22, figs. 51–55) in ventral view. n Cyrtina hibernica Brunton 1984, articulated specimen (RBINS a13097; Mottequin and Simon 2017b, fig. 11l–p) in ventral view (SEM). oAlwynia vesicularis (de Koninck 1851), articulated specimen (RBINS a5185; de Koninck 1887, pl. 8, figs. 30–33) in ventral view.p Beecheria avellana (de Koninck 1887), articulated specimen (RBINS a5073, syntype; de Koninck 1887, pl. 7, figs. 7–10) in ventral view
Spiriferides: Numerous species were reported from Visé by de Koninck (1843 in 1842–1844, 1851, 1887), although many of them were not identified and described in his ultimate contribution (for more information, see Vandercammen and Vandercammen-Goffinet 1970) as is the case of the Reticularioidea (Fig. 11(k)). Pending the investigation of this major order, some more or less emblematic or particular species are illustrated in Fig. 11(d–k). Of these, d’Orbigny (1849)’s species Spirifer oceani (de Koninck 1887; Brunton 1984; Gourvennec 1994) (Fig. 10(j)) is reminiscent of the Mid Devonian spiriferide Verneuilia and the Devonian plicathyridines (Athyridida), which sporadically developed in argillaceous environments during the early–middle Frasnian in the Namur–Dinant Basin (Mottequin et al. 2016), in terms of external morphology. This species was rejected from Verneuilia by Johnson (2006), who considered it as a representative of the Spiriferoidea, but further investigation is needed. D’Archiac and de Verneuil (1842) reported Spirifer cheiropteryx (= Spirifer oceani) from Visé whereas de Koninck (1843 in 1842–1844) recorded it within the calcaire carbonifère friable de Visé. The French adjective friable (as in English) most probably refers to the alteration of the massive algal and bioclastic boundstones of early Warnantian age (late Asbian) that form buildups rich in macrofaunas. Visé and the eponymous formation are the type locality and the type horizon of d’Orbigny’s (1849) species, respectively. Moreover, in Belgium, it is only known from Visé.
Spiriferinides: Besides Cyrtina hibernica (Fig. 11(n)) recently reported by Mottequin and Simon (2017b), the occurrence of the genus Davidsonina must be pointed out. Indeed, the presence of Davidsonina septosa transversa (according to Muir-Wood and Stubblefield 1951) (Fig. 11(l)) was reported in the Visé Limestone and illustrated by de Koninck (1843 in 1842–1844, pl. 12bis, fig. 5a–c (as Spirifer subconicus); 1859, pl. 2, fig. 4; 1887, pl. 33, figs. 1–4 (unidentified (= Syringothyris sp. in Vandercammen and Vandercammen-Goffinet 1970)). In Belgium, Davidsonina septosa is only recorded in the Avins Member by Demanet (1958) and very scarce in the Sovet Formation where it is associated with Davisiella llangollensis according to Mortelmans and Bourguignon (1954). Nonetheless, these reports remain unillustrated until so far. The presence of Syringothyris representatives is not excluded as Horion (1863) reported Spirifer cuspidatus at Visé. Spiriferellina (or Spiriferellina-like) representatives are abundant but still poorly known (Fig. 11(m)) and were photographically illustrated only by Demanet (1931) until now.
Terebratulides: Eleven Dielasma species were reported at Visé by de Koninck (1887), who introduced eight new taxa between 1851 and 1887, but without providing description and/or illustrations of their internal features that are so crucial for generic identification especially in terebratulides in which homeomorphy is common. Besides the erection of the genus Alwynia by Stehli (1961) and the illustration of material of A. vesicularis (Fig. 11(o)) from Visé by Brunton (1982b), Brunton (1984) placed in doubtful synonymy four species described by de Koninck (1887) with Beecheria treakensis Brunton 1984, namely D. tumidum, D. avellana (pars) (Fig. 11(p)) and D. radiatum (pars) (non D. radiatum Termier et al. 1974) (Fig. 8(d–e)), arguing that the latter species should be considered nomina oblita. Let us limit ourselves here to saying that the specimens figured by de Koninck (1887) are curated at the RBINS and, thus, we do not follow Brunton’s (1984) opinion. Finally, this author considered that D. gemmula de Koninck 1887 could be conspecific with Alwynia reidi Brunton 1984. Rare terebratulides display radial ‘colour’ bandings (Fig. 8(d–e)) (see above).
De Koninck’s ( 1887 ) brachiopod species from the Upper Devonian of the Visé area
Several authors (see references above) pointed out the presence of Devonian species among those reported from the calcaire carbonifère de Visé by de Koninck (1842–1844). From the brachiopod viewpoint, this author recorded Terebratula cuboides (non Hypothyridina cuboides (J. de C. Sowerby 1840), see Sartenaer’s (2006) discussion) in the Carboniferous succession, but it is now well known that cuboides-like rhynchonellides are characteristic of the Frasnian succession in southern Belgium, especially in the vicinity of buildups, but are also known from the Lustin and Aisemont formations that crop out in the Visé area (see above).
Among the plethora of brachiopod species described by de Koninck (1887) from the calcaire carbonifère de Visé, it presently appears obvious that at least two species are of Frasnian age based on their preservation (e.g. tint inherited from the matrix that enclosed them) and their strong differences with Mississippian brachiopods. Both species should be added to Mottequin’s (2019) catalogue of Devonian species described in Belgium before 1950, although they should be considered nomina oblita (see below).
Rhynchonella solitaris de Koninck, 1887
(Fig. 12a–e)
1887 Rhynchonella solitaris de Koninck, p. 48, pl. 13, figs. 25–26.
Type material: According to de Koninck (1887), only a single articulated specimen (RBINS a5338, Fig. 12(a–e)) was available for study; it is thus the holotype by virtue of monotypy.

Rhynchonellide (a–e) and spiriferide (f–j) brachiopods from the Frasnian of the Visé area. a–e Rhynchonella solitaris de Koninck 1887, distorted articulated specimen (RBINS a5338, holotype; de Koninck 1887, pl. 14, figs. 25–26) in ventral, dorsal, lateral, posterior and anterior views. f–j Spirifer capillaris de Koninck 1887, distorted articulated specimen (RBINS a5624, syntype) in ventral, dorsal, lateral, posterior and anterior views
Type locality and horizon: Visé area (most probably La Folie hamlet, Fig. 2), Aisemont Formation (late Frasnian).
Description: See de Koninck (1887).
Remarks: This poorly known species is clearly a representative of the Leiorhynchiidae that were so abundant in the mixed carbonate-siliciclastic facies of the Namur–Dinant Basin during the Frasnian. The holotype of Rhynchonella solitaris seems to be closely related to the type species of the genus Navalicria Sartenaer 1989, namely N. compacta Sartenaer 1989, although the ribbing is generally poorly developed in the latter species. N. compacta is particularly abundant in some levels of the Neuville and Aisemont formations in southern Belgium (Mottequin and Poty 2016) and is also known from the Aachen Basin in Germany (Sartenaer and Hartung 1992). If it is proven that both species are conspecific after the examinations of new field collections from La Folie quarry or by the discovery of additional specimens in collections, the former species should be considered a nomen oblitum considering the Article 23.9.1 of the Code (ICZN 1999).
Spirifer capillaris de Koninck, 1887
(Fig. 12f–j)
1887 Spirifer capillaris de Koninck, p. 130, pl. 28, figs. 4–6.
Type material: The specimen illustrated by de Koninck (1887, pl. 28, figs. 4–6) has not been traced so far in de Koninck’s collections curated at the RBINS. Two poorly preserved, articulated specimens identified as such are currently available, but only one (RBINS a5624a, Fig. 12(f–j)) could be possibly ascribed to this species, but it differs from the illustrated specimen in having rounded cardinal extremities. The second specimen (unillustrated here; RBINS a5624b) is characterised by a very high ventral interarea and has thus nothing in common with the species illustrated by de Koninck. For all these reasons, no lectotype is selected among these syntypes.
Type locality and horizon: Visé area, most probably from the Aisemont Formation (late Frasnian).
Description: See de Koninck (1887).
Remarks: The type of ribbing depicted by de Koninck’s drawings on the flanks (simple thin ribs) and the great number of ribs that he mentioned in the sulcus are unusual in Mississippian spiriferides. Moreover, the specimen RBINS a5624 (Fig. 12(f–j)) is clearly a cyrtospiriferid according to these features, but more specimens are needed in order to discuss this species further.
Discussion
Aretz and Chevalier (2007) considered the microbial reefs recognised in the Visé area (Muchez and Peeters 1987) and those known from the Heibaart and Poederlee boreholes in the Campine Basin (Muchez et al. 1987, 1990) very similar to the Viséan Cracoean reefs of North Yorkshire (Mundy 1994), a statement already proposed by Vaughan (1915; see below). The term ‘Cracoean reef’ was introduced by Hudson and Philcox (1965) based on the disused Cracoean stage of Bisat (1928), named after the small village of Cracoe situated at the eastern end of the Craven Reef Belt (e.g. Mundy 1994; Waters et al. 2017). Outside North Yorkshire, these microbial buildups are reported in Ireland, the Isle of Man and England (Mundy 1994) and thus also along the tectonically controlled platform edges developed on the northern margin of the Brabant Massif and bordering the Campine Basin to the south (Aretz and Chevalier 2007).
Close relationships between the faunas from the Warnantian part of the Visé Formation and those occurring in the ‘Brachiopod beds’ (Sibly 1908) of the Midlands in England were soon reported in the literature, not only for the brachiopods but also for the rugose corals (e.g. Delépine 1910). Based on the identical structure and brachiopod and the rugose coral faunas, Vaughan (1915) concluded that the reefs of Visé and Cracoe were contemporaneous and were part of a continuous sea. In the typical Cracoean buildups, benthic organisms are predominant and largely dominated, in order of abundance, by brachiopods, bivalves and gastropods. Brachiopods represent more than 80% of the shelly faunas collected within these reefs and, considering all subfacies, 189 species of brachiopods were reported on the 518 macroinvertebrate taxa (except echinoderms) recognised (Mundy 1978, 1980, 1994). Comparing the lists of brachiopods from Cracoean facies provided by Mundy (1980) for North Yorkshire and that of Treak Cliff in North Derbyshire by Brunton and Tilsley (1991), it is obvious that the brachiopod fauna from Visé is, as a whole, very similar to those from the English localities. Furthermore, it has to be pointed out that the taxonomic revision of the fauna from Visé (except the groups previously documented; see references above) is only in its early stages and some authors (e.g. Brunton and Mundy 1986) reported some genera (Retroplexus) in Visé, but without documenting further their data. Although it is unfortunately not possible to examine the brachiopods in situ due to the inaccessibility of the Souvré–Richelle quarries, it should be not too difficult to try to restore some of them (those which are clearly of reefal type) to their possible original environment thanks to thorough investigations of Mundy (1978, 1980, 1994).
Conclusions
More than 175 years after the publication of de Koninck’s (1842–1844) monograph—the first of a long series—it was necessary to provide a brief account on the Visé fauna, and especially on the brachiopods which rank among the most abundant elements of this particular fauna alongside mollusks (gastropods, bivalves), pending their thorough taxonomic revision. Indeed, palaeontological collections from Visé, i.e. the type locality of the Viséan Stage (Hance et al. 2006), are present in many scientific institutions worldwide and are generally only labelled ‘Visé’ without any more information. Based on literature data and field observations of one of us (EP), it appears that Visé fauna was almost exclusively recovered from the late early Warnantian part (late Asbian) of the Visé Formation (Poty and Delculée 2011), and more specifically from the microbial buildup level (Muchez and Peeters 1987) of Cracoean affinities (Aretz and Chevalier 2007) that was exploited in the quarries F and G (following Horion and Gosselet’s (1892) lettering). Some fossils also came from the overlying levels and are of Brigantian and possibly Serpukhovian age, but their preservation is totally different. These two disused quarries are located in the northeastern part of the Hermalle-sous-Argenteau block (Poty 1991; Poty and Delculée 2011). Besides brachiopods, almost the whole Visé fauna deserves a complete re-assessment in the future.
References
Alvarez, F., & Brunton, C. H. C. (2000). A review of two de Koninck retzioid brachiopod species, and description of a new genus from the Carboniferous of Europe. Palaeontology, 43, 817–832.
Archiac, A. d', & Verneuil, E. de (1842). On the fossils of the older deposits of the Rhenish Provinces. Transactions of the Geological Society of London, series 2, 6(2), 303–410.
Aretz, M., & Chevalier, E. (2007). After the collapse of stromatoporid–coral reefs — the Famennian and Dinantian reefs of Belgium: much more than Waulsortian mounds. Geological Society, London, Special Publications, 275, 163–188.
Barchy, L., & Marion, J.-M. (2000). Carte géologique de Wallonie: Dalhem – Herve, 42/3-4. 1/25 000. Namur: Ministère de la Région wallonne, Direction générale des Ressources naturelles et de l’Environnement.
Barchy, L., & Marion, J.-M. (2017). Carte géologique de la Wallonie: Visé – Sint Maertens Voeren, 34/7-8. 1/25 000. Namur: Service public de Wallonie, Direction générale de l’Agriculture, des Ressources Naturelles et de l’Environnement.
Bisat, W. S. (1928). The Carboniferous goniatite zones of England and their continental equivalents. Congrès pour l’Avancement des Etudes de Stratigraphie carbonifère. Heerlen, 7–11 juin 1927. Compte Rendu, 117–133. Liège: Vaillant-Carmanne.
Bond, G. (1941). Species and variation in British and Belgian Carboniferous Schizophoriidae. Proceedings of the Geologists’ Association, 52, 285–303.
Brand, P. J. (1972). Some British Carboniferous species of the brachiopod genus Leptagonia McCoy. Bulletin of the Geological Survey of Great Britain, 39, 57–79.
Brunton, C. H. C. (1968). Silicified brachiopods from the Viséan of County Fermanagh (II). Bulletin of the British Museum (Natural History), Geology, 16, 1–70.
Brunton, C. H. C. (1971). An endopunctate rhynchonellid brachiopod from the Viséan of Belgium and Britain. Palaeontology, 14, 95–106.
Brunton, C. H. C. (1979). The Lower Carboniferous brachiopod genus Levitusia Muir-Wood, H. M. and Cooper, G. A., from western Europe and the USSR. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 51(10), 1–23.
Brunton, C. H. C. (1982a). The functional morphology and palaeoecology of the Dinantian brachiopod Levitusia. Lethaia, 15, 149–167.
Brunton, C. H. C. (1982b). British Dinantian (Lower Carboniferous) terebratulid brachiopods. Bulletin of the British Museum (Natural History), Geology, 36, 45–57.
Brunton, C. H. C. (1984). Silicified brachiopods from the Viséan of County Fermanagh, Ireland (III). Rhynchonellids, Spiriferids and Terebratulids. Bulletin of the British Museum (Natural History), Geology, 38, 27–130.
Brunton, C. H. C., & Mundy, D. J. C. (1983). The discovery of the Dinantian Productacean Septarinia leuchtenbergensis (de Koninck) in the British Isles. Proceedings of the Yorkshire Geological Society, 44, 333–340.
Brunton, C. H. C., & Mundy, D. J. C. (1986). Some Dinantian Chonopectinid Productaceans (Brachiopoda) from the British Isles. Proceedings of the Yorkshire Geological Society, 46, 1–10.
Brunton, C. H. C., & Mundy, D. J. C. (1988). Strophalosiacean and aulostegacean productoids (Brachiopoda) from the Craven Reef Belt (late Viséan) of North Yorkshire. Proceedings of the Yorkshire Geological Society, 47, 55–88.
Brunton, C. H. C., & Mundy, D. J. C. [with appendix by S. S. Lazarev] (1993). Productellid and plicatiferid (productoid) brachiopods from the Lower Carboniferous of the Craven Reef Belt, North Yorkshire. Bulletin of the British Museum (Natural History), Geology, 49, 99–119.
Brunton, C. H. C., & Tilsley, J. W. (1991). A check list of brachiopods from Treak Cliff, Derbyshire, with reference to other Dinantian (Lower Carboniferous localities). Proceedings of the Yorkshire Geological Society, 48, 287–295.
Brunton, C. H. C., Rachebœuf, P. R., & Mundy, D. J. C. (1994). Reclassification of Semenewia concentrica (de Koninck, 1847) (Brachiopoda, Lower Carboniferous). Geobios, 27, 51–60.
Chemnitz, J. H. (1784). Neues systematisches Conchylien-Cabinet. Band 7. G. N. Raspe.
Clarke, J. M. (1904). Naples Fauna in Western New York. Part 2. New York State Museum, Memoir, 6, 199–454.
Cooper, G. A., & Muir-Wood, H. M. (1967). New names for brachiopod homonyms. Journal of Paleontology, 41, 808.
Davreux, C. J. (1833). Essai sur la constitution géognostique de la province de Liége. Mémoires couronnés de l’Académie royale des Sciences et Belles-Lettres de Bruxelles, 9, 1–297.
Defrance, M. J. L. (1826). Art. Productus. Dictionnaire des Sciences Naturelles, 43, 349–355.
Delépine, G. (1909). Note sur le Calcaire carbonifère de Visé et les couches à brachiopodes du Midland. Bulletin de la Société géologique de France, 4ème série, 9, 307–309.
Delépine, G. (1910). Note sur la position stratigraphique du calcaire carbonifère à Visé. Annales de la Société géologique de Belgique, 37, B238–B246.
Delépine, G. (1911). Recherches sur le Calcaire carbonifère de la Belgique. Mémoires et travaux publiés par des professeurs des facultés catholiques de Lille, 8, 1–419.
Delépine, G. (1921). Les formations supérieures du Calcaire carbonifère de Visé. Annales de la Société scientifique de Bruxelles, 41, 113–123.
Delépine, G. (1922). Notes complémentaires sur la faune du Calcaire carbonifère à Visé et à Florennes (Belgique). Annales de la Société scientifique de Bruxelles, 42, 156–160.
Delépine, G. (1940). Les Goniatites du Dinantien de la Belgique. Mémoires du Musée royal d’Histoire naturelle de Belgique, 91, 1–91.
Demanet, F. (1931). Spiriferina peracuta de Koninck, espèce autonome, distincte de Spiriferina octoplicata Sowerby. Bulletin du Musée royal d’Histoire naturelle de Belgique, 7(18), 1–22.
Demanet, F. (1934). Les brachiopodes du Dinantien de la Belgique. Vol. 1, Atremata, Neotremata, Protremata (pars). Mémoires du Musée royal d’Histoire naturelle de Belgique, 61, 1–116.
Demanet, F. (1958). Contribution à l’étude du Dinantien de la Belgique. Mémoires de l’Institut royal des Sciences naturelles de Belgique, 141, 1–152.
Derycke, C., Cloutier, R., & Candilier, A.-M. (1995). Palaeozoic vertebrates of northern France and Belgium: part II – Chondrichthyes, Acanthodii, Actinopterygii (uppermost Silurian to Carboniferous). Geobios, Mémoire spécial, 19, 343–350.
Dewalque, G. (1868). Prodrome d’une description géologique de la Belgique. Libraire polytechnique De Decq.
Dumont, A. (1832). Mémoire sur la constitution géologique de la province de Liège. Mémoires couronnés de l’Académie royale de Sciences et Belles-Lettres de Bruxelles, 8, 1–374.
Fischer de Waldheim, G. (1837). Oryctographie du gouvernement de Moscou. A. Semen.
Forir, H., & Destinez, P. (1901). Contribution à la détermination de l’âge du massif carboniférien de Visé. Annales de la Société géologique de Belgique, 28, M61–M68.
Fraipont, J. (1888). Une lingule nouvelle du calcaire carbonifère de Visé (Lingula Konincki). Annales de la Société géologique de Belgique, 15(Bulletin), 142–143.
Godefroid, J., Mottequin, B., & Yochelson, E. L. (2006). Restudy of the Lower Carboniferous Scaphopoda described by de Koninck (1843, 1883). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 76, 137–163.
Goldfuss, G. A. (1840). Petrefacta Germaniae tam ea, quae in museo universitatis regiae Borussicae Fridericiae Wilhelmiae Rhenanae servantur, quam alia quaecunque in museis Hoennghusiano, Muensteriano aliisque extant iconibus et descriptionibus illustrata. Zweiter Theil. Düsseldorf: Arnz.
Gosselet, J. (1861). Observations sur les terrains primaires de la Belgique et du nord de la France. Bulletin de la Société géologique de France, série, 2(18), 18–33.
Gourvennec, R. (1994). The genus Verneuilia Hall & Clarke, 1893 (Brachiopoda, Spiriferida). Geobios, 27, 573–581.
Graham, D. K. (1972). A review of the brachiopod genus Orbiculoidea in the Scottish Carboniferous. Bulletin of the Geological Survey of Great Britain, 38, 43–58.
Grant, R. E. (1965). The brachiopod superfamily Stenoscismatacea. Smithsonian Miscellaneous Collections, 148(2), 1–192.
Gröber, P. (1908). Ueber die Faunen des untercarbonischen Transgressionsmeeres des zentralen Tian-schan, die in der Umgebung des Sart-dschol-Passes gefunden worden sind. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 26, 213–248.
Hahn, G., Hahn, R., & Brauckmann, C. (1988). Die Trilobiten des belgischen Kohlenkalkes (Unter-Karbon). 10. Biostratigraphie. Geologica et Palaeontologica, 22, 55–72.
Hance, L., Poty, E., & Devuyst, F.-X. (2006). Viséan. Geologica Belgica, 9, 55–62.
Hoare, R. D. (2002). European Paleozoic Polyplacophora, Multiplacophora, and Turrilepadida in United States repositories. Journal of Paleontology, 76, 95–108.
Hoeninghaus, F. W. (1828). Sur un Productus singulier de Visé. Krefeld: privately printed.
Horion, C. (1860). Note sur le calcaire dévonien des environs de Visé. Bulletin de la Société géologique de France, 2ème série, 17, 58–61.
Horion, C. (1863). Sur les terrains primaires des environs de Visé, Bulletin de la Société géologique de France, 2ème série., 20, 766–770.
Horion, C., & Gosselet, J. (1892). Les calcaires de Visé. 1ère partie : Etude stratigraphique. Annales de la Société géologique du Nord, 20, 194–212.
Hudson, R. G. S., & Philcox, M. E. (1965). The Lower Carboniferous stratigraphy of the Buttevant area, Co. Cork. Proceedings of the Royal Irish Academy, 64, 65–79.
Jackson, J. W. (1922). On the occurrence of Daviesiella llangollensis (Dav.) in Derbyshire. Geological Magazine, 59, 461–468.
Jackson, R. T. (1929). Palaeozoic Echini of Belgium. Mémoires du Musée royal d’Histoire naturelle de Belgique, 38, 1–74.
Johnson, J. G. (2006). Family Verneuiliidae Schuchert, 1929. In R. L. Kaesler (Ed.), Treatise on invertebrate paleontology, Part H (Revised), Brachiopoda, 5 (p. 1746). Geological. Society of America and University of Kansas Press.
Koninck, L.-G. de (1842–1844). Description des animaux fossiles qui se trouvent dans le terrain carbonifère de Belgique. H. Dessain.
Koninck, L.-G. de (1847a). Monographie du genre Productus. Mémoires de la Société royale des Sciences de Liège, 4, 73–278.
Koninck, L.-G. de (1847b). Recherches sur les animaux fossiles, Part I. Monographie des genres Productus et Chonetes. H. Dessain.
Koninck, L.-G. de (1851). Description des animaux fossiles qui se trouvent dans le terrain carbonifère de Belgique. Supplément. H. Dessain.
Koninck, L.-G. de (1859). Liste des Brachiopodes carbonifères de Belgique, munis d’une spire, et des localités dans lesquelles ils se trouvent. In T. Davidson (Ed.), Mémoire sur les genres et les sous-genres des brachiopodes munis d’appendices spiraux et sur leurs espèces découvertes dans les couches carbonifères des Iles britanniques ; traduit et augmenté de notes par L. de Koninck. H. Dessain.
Koninck, L.-G. de (1872). Nouvelles recherches sur les animaux fossiles du terrain carbonifère de la Belgique. Mémoires de l’Académie royale des Sciences, des Lettres et des Beaux-Arts de Belgique, 39(1), 1–178.
Koninck, L.-G. de (1878). Faune du calcaire carbonifère de la Belgique. Première partie. Poissons et genre Nautile. Annales du Musée royal d’Histoire naturelle de Belgique, 2, 1–152.
Koninck, L.-G. de (1880). Faune du calcaire carbonifère de la Belgique. Deuxième partie. Genres: Gyroceras, Cyrtoceras, Gomphoceras, Orthoceras, Subclymenia et Goniatites. Annales du Musée royal d’Histoire naturelle de Belgique, 4, 1–83.
Koninck, L.-G. de (1881). Faune du calcaire carbonifère de la Belgique. Troisième partie. Gastéropodes. Annales du Musée royal d’Histoire naturelle de Belgique, 6, 1–170.
Koninck, L.-G. de (1883a). Faune du calcaire carbonifère de la Belgique. Quatrième partie. Gastéropodes (suite et fin). Annales du Musée royal d’Histoire naturelle de Belgique, 8, 1–240.
Koninck, L.-G. de (1883b). Sur le Spirifer mosquensis et sur ses affinités avec quelques autres espèces du même genre. Bulletin du Musée royal d’Histoire naturelle de Belgique, 2, 371–395.
Koninck, L.-G. de (1885). Faune du calcaire carbonifère de la Belgique. Cinquième partie. Lamellibranches. Annales du Musée royal d’Histoire naturelle de Belgique, 11, 1–283.
Koninck, L.-G. de (1887). Faune du calcaire carbonifère de la Belgique. Sixième partie. Brachiopodes. Annales du Musée royal d’Histoire naturelle de Belgique, 14, 1–154.
Koninck, L.-G. de, & Lehon, H. (1854). Recherches sur les Crinoïdes du terrain carbonifère de la Belgique. Mémoires de l’Académie royale des Sciences, des Lettres et des Beaux-Arts de Belgique, 28(3), 1–215.
Léveillé, C. (1835). Aperçu géologique de quelques localités très riches en coquilles sur les frontières de France et de Belgique. Mémoires de la Société géologique de France, 2, 29–40.
M‘Coy, F. (1844). A synopsis of the characters of the Carboniferous Limestone fossils of Ireland. University Press.
M‘Coy, F. (1852). On some new Brachiopoda from the Carboniferous Limestone. Annals and Magazine of Natural History, 2nd series, 10, 421–429.
Maillieux, E. (1925). Note sur quelques Céphalopodes des terrains paléozoïques de la Belgique. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 34, Procès-verbaux, 34–41.
Martin, W. (1793). Figures and descriptions of petrifications collected in Derbyshire. Lyon & Atkinson.
Martin, W. (1809). Petrificata Derbiensia; or figures and descriptions of petrefactions collected in Derbyshire. D. Lyon.
Mortelmans, G., & Bourguignon, P. (1954). Le Dinantien. In P. Fourmarier (Ed.), Prodrome d’une description géologique de la Belgique (pp. 217–321). Société géologique de Belgique.
Mottequin, B. (2008). The ‘black marble’ of Denée, a fossil conservation deposit from the Lower Carboniferous (Viséan) of southern Belgium. Geological Journal, 43, 197–208.
Mottequin, B. (2019). An annotated catalogue of types of Silurian–Devonian brachiopod species from southern Belgium and northern France in the Royal Belgian Institute of Natural Sciences (1870–1945), with notes on those curated in other Belgian and foreign institutions. Geologica Belgica, 22, 47–89.
Mottequin, B. (2021). Earth science collections of the Centre Grégoire Fournier (Maredsous) with comments on Middle Devonian–Carboniferous brachiopods and trilobites from southern Belgium. Geologica Belgica, 24, 33–68.
Mottequin, B., & Poty, E. (2016). Kellwasser horizons, sea-level changes and brachiopod-coral crises during the late Frasnian in the Namur-Dinant Basin (southern Belgium): a synopsis. Geological Society, London, Special Publications, 423, 235–250.
Mottequin, B., & Simon, E. (2017a). New insights on Tournaisian–Visean (Carboniferous, Mississippian) athyridide, orthotetide, rhynchonellide, and strophomenide brachiopods from southern Belgium. Palaeontologia Electronica, 20.2.28A(2), 1–45.
Mottequin, B., & Simon, E. (2017b). Revision of some spiriferide and spiriferinide brachiopods from the historical type area of the Tournaisian Stage (Carboniferous, southern Belgium). Paläontologische Zeitschrift, 91, 473–496.
Mottequin, B., Brice, D., Marion, J.-M., & Simon, E. (2016). Plicathyridine brachiopods (Athyridida) from the Frasnian (Late Devonian) of Western Europe and Middle East. Geobios, 49, 381–393.
Mourlon, M. (1881). Géologie de la Belgique (Vol. 2). Hayez.
Muchez, P., & Peeters, C. (1987). The occurrence of a cryptalgal reef structure in the upper Viséan of the Visé area. Annales de la Société géologique de Belgique, 109, 573–577.
Muchez, P., Conil, R., Viaene, W., Bouckaert, J., & Poty, E. (1987). Sedimentology and biostratigraphy of the Viséan carbonates of the Heibaart (DzH1) borehole (Northern Belgium). Annales de la Société géologique de Belgique, 110, 199–208.
Muchez, P., Viaene, W., Bouckaert, J., Conil, R., Dusar, M., Poty, E., Soille, P., & Vandenberghe, N. (1990). The occurrence of a microbial buildup at Poederlee (Campine Basin, Belgium): biostratigraphy, sedimentology, early diagenesis and significance for early Warnantian paleogeography. Annales de la Société géologique de Belgique, 113, 329–339.
Muir-Wood, H. M. (1928). The British Carboniferous Producti, part 2. Productus (sensu stricto); semireticulatus and longispinus groups. Memoirs of the Geological Survey of Great Britain, 3(1), 1–217.
Muir-Wood, H. M. (1962). On the morphology and classification of the brachiopod suborder Chonetoidea. British Museum (Natural History).
Muir-Wood, H. M., & Cooper, G. A. (1960). Morphology, classification and life habits of the Productoidea (Brachiopoda). The Geological Society of America, Memoir, 81, 1–447.
Muir-Wood, H. M., & Stubblefield, C. J. (1951). Proposed addition to the ‘Official list of specific trivial names in zoology’ of the trivial names of two species of the class Cephalopoda and of twelve species of the class Brachiopoda hitherto attributed to Martin, 1809, and matters incidental thereto. The Bulletin of zoological nomenclature, 6, 18–30.
Mundy, D. J. C. (1978). Reef communities. In W. S. McKerrow (Ed.), The ecology of fossils (pp. 157–167). Duckworth.
Mundy, D. J. C. (1980). Aspects of the palaeoecology of the Craven Reef Belt (Dinantian) of North Yorkshire. Unpublished PhD thesis, University of Manchester.
Mundy, D. J. C. (1994). Microbialite–sponge–bryozoan–coral framestones in Lower Carboniferous (Late Viséan) buildups of Northern England (UK). Canadian Society of Petroleum Geologists, Memoir, 17, 713–729.
Nagel, J. (2006). Middle and Upper Devonian Cryptodonta (Bivalvia) from the pelagic Hercynian facies – Taxonomy, stratigraphy, and paleoecology. Unpublished PhD thesis, Westfälischen Wilhelms-Universität Münster.
Oehlert, D. P. (1887). Appendice: Brachiopodes; notions sur l’anatomie et la physiologie des brachiopodes. In P. H. Fischer (Ed.), Manuel de Conchyliologie et de Paléontologie conchyliologique ou histoire naturelle des Mollusques vivants et fossiles, fascicule 11 (pp. 1189–1334). F. Savy.
Orbigny, A. d.’. (1849). Prodrome de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés faisant suite au cours élémentaire de paléontologie et de géologie stratigraphiques (Vol. 1). Victor Masson.
Paeckelmann, W. (1930). Die Fauna des deutschen Unterkarbons. 1. Teil. 3. Die Brachiopoden des deutschen Unterkarbons. I. Teil. Die Orthiden, Strophomeniden und Choneten des Mittleren und Oberen Unterkarbons. Abhandlungen der Preußischen Geologischen Landesanstalt, Neue Folge, 122, 143–326.
Paeckelmann, W. (1931). Die Fauna des deutschen Unterkarbons. 2.Teil. Die Brachiopoden des deutschen Unterkarbons. 2. Teil. Die Productinae und Productus-ähnlichen Chonetinae. Abhandlungen der Preußischen Geologischen Landesanstalt, Neue Folge, 136, 1–440.
Paeckelmann, W. (1942). Beiträge zur Kenntnis devonischer Spiriferen. Abhandlungen des Reichsamts für Bodenforschung, Neue Folge, 197, 1–188.
Pander, C. H. (1830). Beiträge zur Geognosie des Russischen Reiches. K. Kray.
Phillips, J. (1836). Illustrations of the geology of Yorkshire, Part 2. The mountain limestone district. J. Murray.
Pirlet, H. (1967). Nouvelle interprétation des carrières de Richelle: le Viséen de Visé (2e note sur les calcaires de Visé). Annales de la Société géologique de Belgique, 90, B299–B328.
Pocock, Y. P. (1965). Devonian and Carboniferous schizophoriid brachiopods from Western Europe. Unpublished PhD thesis, London University.
Pocock, Y. P. (1968). Carboniferous schizophoriid brachiopods from Western Europe. Palaeontology, 11, 64–93.
Portlock, J. E. (1843). Report on the Geology of the County of Londonderry and of parts of Tyrone and Fermanagh. A. Milliken.
Poty, E. (1981). Recherches sur les Tétracoralliaires et les Hétérocoralliaires du Viséen de la Belgique. Mededelingen Rijks Geologische Dienst, 35(1), 1–161.
Poty, E. (1982). Paléokarsts et brèches d’effondrement dans le Frasnien moyen des environs de Visé. Leur influence dans la paléogéographie dinantienne. Annales de la Société géologique de Belgique, 105, 315–337.
Poty, E. (1991). Tectonique de blocs dans le prolongement oriental du Massif du Brabant. Annales de la Société géologique de Belgique, 114, 265–275.
Poty, E. (1997). Devonian and Carboniferous tectonics in the eastern and southern part of the Brabant Massif (Belgium). Aardkundige Mededelingen, 8, 143–144.
Poty, E. (2007). The ‘Avins event’: a remarkable worldwide spread of corals at the end of the Tournaisian (Lower Carboniferous). Schriftenreihe der Erdwissenschaftlichen Kommissionen, Österreichische Akademie der Wissenschaften, 17, 231–249.
Poty, E. (2016). The Dinantian (Mississippian) succession of southern Belgium and surrounding areas: stratigraphy improvement and inferred climate reconstruction. Geologica Belgica, 19, 177–200.
Poty, E., & Delculée, S. (2011). Interaction between eustacy and block-faulting in the Carboniferous of the Visé–Maastricht area (Belgium, The Netherlands). Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 162(2), 117–126.
Poty, E., Hance, L., Lees, A., & Hennebert, M. (2002). Dinantian lithostratigraphic units (Belgium). Geologica Belgica, 4, 69–94.
Poty, E., Devuyst, F.-X., & Hance, L. (2006). Late Devonian and Mississippian foraminiferal and rugose coral zonations of Belgium and Northern France, a tool for Eurasian correlations. Geological Magazine, 143, 829–857.
Quenstedt, F. A. (1852). Handbuch der Petrefaktenkunde. Laupp.
Quenstedt, F. A. (1868–1871). Petrefactenkunde Deutschlands: Abtheilung 1, Band 2. Die Brachiopoden. Fues’s Verlag (R. Reisland).
Quenstedt, F. A. (1885). Handbuch der Petrefaktenkunde. Laupp.
Ramsbottom, W. H. C. (1963). The Carboniferous Limestone brachiopod Streptorhynchus anomalus (J. de C. Sowerby). Transactions of the Leeds Geological Association, 7, 175–179.
Ryckholt, P. de (1851). Mélanges paléontologiques, première partie. Mémoires couronnés des Savants étrangers de l’Académie royale des Sciences, des Lettres et des Beaux-Arts de Belgique, 24, 1–176.
Ryckholt, P. de (1854). Mélanges paléontologiques. Deuxième partie. Privately printed.
Sartenaer, P. (1989). Deux genres rhynchonellides nouveaux d’âge frasnien moyen et supérieur, résultant du brisement de Calvinaria Stainbrook, 1945. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 59, 61–77.
Sartenaer, P. (2006). Parallelepipedorhynchus, a new late Frasnian rhynchonellid (brachiopod) genus from the Dinant Basin, Belgium, and Parallelepipedorhynchidae n. fam. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 76, 1–12.
Sartenaer, P., & Hartung, W. (1992). Die Biozonierung der Oberen Frasnes-Stufe (Frasnes-Knollenkalke und Frasnes-Schiefer) am Nord-Abfall des Hohen Venn smit Hilfe von Rhynchonelliden. Senckenbergiana lethaea, 72, 49–60.
Schuchert, C., & Cooper, G. A. (1931). Synopsis of the brachiopod genera of the suborders Orthoidea and Pentameroidea, with notes on the Telotremata. American Journal of Science, series, 5(22), 241–255.
Sibly, T. F. (1908). On the faunal succession in the Carboniferous Limestone (upper Avonian) of the Midland area (North Derbyshire and North Staffordshire). Quarterly Journal of the Geological Society, 64, 34–82.
Sowerby, J. (1815–1818). The Mineral Conchology of Great Britain, vol. 2. Benjamin Meredith.
Sowerby, J. (1821–1822). The Mineral Conchology of Great Britain, vol. 4. London: privately printed.
Sowerby, J. de C. (1823–1825). The Mineral Conchology of Great Britain, vol. 5. London: privately printed.
Sowerby, J. de C. (1840). Explanation of the plates and woodcuts. In A. Sedgwick, & R. I. Murchison, On the physical structure of Devonshire, and on the subdivisions and geological relations of its older stratified deposits. Transactions of the Geological Society of London, 2nd series, 5, 633–703.
Sowerby, J. de C. (1840–1846). The Mineral Conchology of Great Britain, vol. 7. London: privately printed.
Stehli, F. G. (1961). New genera of Upper Paleozoic terebratuloids. Journal of Paleontology, 35, 457–466.
Termier, G., Termier, H., Lapparent, A. F., & Marin, P. (1974). Monographie du Permo-Carbonifère de Wardak (Afghanistan Central). Documents des Laboratoires de Géologie de la Faculté des Sciences de Lyon, Hors-Série, 2, 1–165.
Thomas, I. (1914). The British Carboniferous Producti, part 1: genera Pustula and Overtonia. Memoirs of the Geological Survey of Great Britain (Palaeontology), 1(4), 197–366.
Tourneur, F., Conil, R., & Poty, E. (1989). Données préliminaires sur les tabulés et les chaetetides du Dinantien de la Belgique. Bulletin de la Société belge de Géologie, 98(3–4), 401–442.
Vandercammen, A., & Vandercammen-Goffinet, I. (1970). Liste des types du sous-ordre des Spiriferidina Waagen, W. 1883 (Brachiopoda) conservés à l’Institut royal des Sciences naturelles de Belgique. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 46(38), 1–87.
Vaughan, A. (1915). Correlation of Dinantian and Avonian. Quaterly Journal of the Geological Society of London, 71, 1–52.
Verneuil, E. de (1840). Sur quelques espèces intéressantes de Brachiopodes des terrains anciens. Bulletin de la Société géologique de France, 1ère série, 11, 257–264.
Verneuil, E. de (1841). Sur une observation que lui a suggérée la 7e livraison du bel ouvrage de M. Goldfuss sur les pétrifications d’Allemagne. Bulletin de la Société géologique de France, 1ère série, 12, 198–200.
von Buch, L. (1832). Über Goniatiten. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin, 1831, 159–187.
Waters, C. N., Haslam, R. B., Cózar, P., Somerville, I. D., Millward, D., & Woods, M. (2017). Mississippian reef development in the Cracoe Limestone Formation of the southern Askrigg Block, North Yorkshire, UK. Proceedings of the Yorkshire Geological Society, 61, 179–196.
Weir, J. (1931). The British and Belgian Carboniferous Bellerophontidae. Transactions of the Royal Society of Edinburgh, 56, 767–861.
Acknowledgements
The SEM images of the brachiopods were taken at the Royal Belgian Institute of Natural Sciences at Brussels by Julien Cillis. The manuscript benefited from the thorough reviews of George Sevastopulo and Xueping Ma.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Mottequin, B., Poty, E. Brachiopods from the historical type area of the Viséan Stage (Carboniferous, Mississippian; Belgium) and the Visé fauna: preliminary remarks. Palaeobio Palaeoenv 102, 351–371 (2022). https://doi.org/10.1007/s12549-021-00498-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-021-00498-9