
Abstract
The Baelen mud mounds in eastern Belgium represent a local member of the mid-Famennian Souverain-Pré Formation (an important carbonate interval within the Condroz Sandstone Group). The lower part of this member contains silty bioclastic wackestones and packstones that are particularly rich in the problematical protists Serrisinella and Dreesenulella. Plurilocular foraminifera (Septabrunsiina and Baelenia) and rare solitary rugose corals (Neaxon? sp.) occur within crinoidal grainstones (tempestites) interfingering with the latter bioclastic wacke-/packstones and with red-stained stromatactoid spiculitic mudstones (carbonate mud mound core facies). Although Serrisinella is quite common in other mid-Famennian limestones of Belgium, Dreesenulella is almost endemic of the Baelen Member. Both genera apparently inhabited the muddy sediment-water interface, constituting meadows probably in zones of intermittently strong bottom currents. The taxonomic similarities between Dreesenulella and Saccamminopsis are discussed, as well as their possible affinities with the Xenophyophora and Kokomiacea. The first stages of the Septabrunsiina–Baelenia foraminiferal lineage are analysed in detail. The taxonomic and palaeoecological positioning of Serrisinella and Dreesenulella adds to the discussion about the palaeobathymetry of the Baelen mud mounds and corroborates sedimentological evidence for their relatively shallow carbonate ramp depositional setting.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The mid-Famennian (late Upper Devonian) Baelen Member represents a lateral mud mound type facies of the Souverain-Pré Formation (Late marginifera conodont Zone). This mud mound complex is a real carbonate plug within an overall siliciclastic (silty-sandy) environment and displays exceptional biosedimentological phenomena (Dreesen et al. 1985, 2013, 2015b). The Souverain-Pré Formation corresponds to a short-term transgressive event and an eustatic rise during the Famennian, interrupting the regressive megasequence on the Condroz shelf in Belgium. The Baelen Member is the only known or at least the only well-documented mid-Famennian carbonate mud mound complex worldwide (Dreesen et al. 1985, 2013) after the end-Frasnian mass extinction that wiped out numerous invertebrate taxa. In contrast with the better-known Belgian upper Frasnian mud mounds, the mid-Famennian Baelen mud mounds are totally devoid of in situ tabulate corals and stromatoporoids. Only poorly preserved solitary rugose corals are present. The Baelen mud mounds differ from the more or less coeval true reefs described from the Alberta (Canada) and Canning (Australia) basins (see Fagerstrom 1994; Webb 2001; Wood 2004).
The main goals of this paper are to provide additional data on the nature and the palaeoecology of the incertae sedis Serrisinella and Dreesenulella (formerly Baculella) and of the plurilocular foraminifera Septabrunsiina and Baelenia and to briefly discuss their value for reconstructing the depositional environment of the Baelen Member.
Geological and lithostratigraphical setting
The red crinoidal Baelen Limestone, also known as the Baelen Red Marble (red-stained crinoidal and stromatactis-bearing limestones), represents the core of the Baelen member mud mound complex. It is a famous historical building stone from the Limbourg area (Vesdre Nappe, eastern Belgium; Fig. 1), where it has been quarried since Roman times (Dreesen et al. 2015a, b). The Baelen Member is a local member of the middle Famennian Souverain-Pré Formation (e.g. Dreesen et al. 1985, 2013; Thorez et al. 2006; Figs. 2, 3, and 4). In Belgium, the latter corresponds to the first major reappearance of marine carbonates in the otherwise siliciclastic Famennian shelf setting, after the disappearance of widespread shelf carbonates that flourished during the previous Frasnian greenhouse episode. The Baelen Member accumulated during one single 4th-order eustatic cycle, the duration of which is possibly less than 0.5 my (the Late marginifera conodont Zone pro parte; Thorez and Dreesen 1997) (Fig. 3). Different carbonate microfacies have been recognised within the Baelen Member. These comprise nodular to lenticular algospongial packstones/bindstones and massive stromatactis-bearing microbialitic mudstones, interfingered with lenticular crinoidal grainstones or rudstones. Silty nodular bioclastic wackestones/packstones, strongly affected by pressure solution (called “peastones”, due to the presence of numerous white circular sections of the crinoid ossicles resembling peas), mark the transition with underlying and overlying micaceous sandstones and occur also as interbeds within the mudmound core (Fig. 4). Dreesen et al. (2013, 2015b) provided a detailed description of the various lithologies and different carbonate microfacies within the Baelen Member.


Simplified geological map of the Limbourg area (modified from Ghysel et al. 1995) highlighting the location of individual mud mound complexes within the Baelen Member. Sampled outcrops: 1, Baelen-Les Forges; 2, Dolhain (La Belle Vue); 3, “Mali” quarry; 4, Limbourg municipal waste deposit; 5, Limbourg La Beverie (playground); 6, Goé; 7, Botterweck (western section); 8, Botterweck (eastern section)

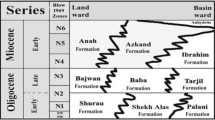
General lithostratigraphical and biostratigraphical framework of the Middle Famennian within the Dinant Synclinorium and the Vesdre Nappe (modified from Dreesen 1989 and Dreesen et al. 2013). CZ conodont zone, E early, L late, L+ latest, Fm formation, Mb member. The oolitic ironstone levels are conspicuous lithostratigraphical event-stratigraphical marker beds within both structural units (e.g. Dreesen 1987, 1989)

It is important to note that all mud mounds occur in a rather restricted area within SW-NE directed fold limbs in between the towns and villages of Limbourg, Baelen and Goé (Fig. 2). The carbonate mounds are preferentially located on a block-faulted basement (Marion 1985; Dreesen et al. 1985) (Fig. 5), and their cluster coincides with the northern extremity of a deep-seated NNW-SSE oriented transversal fault, known also as the Verviers-Trier dislocation (Dvorak 1973).

Cartoon depicting the major depositional environments of the Condroz Sandstone Group (see Fig. 3) in Belgium (after Dreesen and Thorez 1994). The upper part of the figure shows the prograding lithofacies (regressive megasequence) and the stratigraphic succession (from right to left) of the corresponding formations with time (rhomboidea through expansa conodont zones). Arrows point to transport directions of storm surges and tidal currents generating tempestites and resulting in mixing of bioclasts and biota. Note conspicuous development of carbonate mounds (red) during the late marginifera conodont zone and their supposed location at the edge of the carbonate ramp (below SWB) on top of synsedimentary faults. The boxes below refer to the various (subordinated) carbonate deposits within a dominant siliciclastic environment
The Baelen Member biota
The fossil (micro-) biota formerly described by Dreesen et al. (1985, 2015b) and/or newly identified within this paper (observed in outcrop, in thin section or in the dissolution residues after acid etching of limestone), comprise cyanobacteria (Girvanella spp.), microproblematic protists (Serrisinella, Dreesenulella and Disonella), plurilocular foraminifers (Septabrunsiina baeleni and Baelenia gosseleti), rare solitary rugose corals (e.g. Neaxon? sp.), ostracod species and hexactinellid sponge spicules. Several of the microproblematics mentioned above have been partially misinterpreted in the past and these are re-discussed herein. Their palaeoecology might provide additional lines of evidence for the depositional setting of the Baelen mud mounds. The studied material is housed at the Evolution & Diversity Dynamics Lab of Liège University.
Cyanobacteria
Cyanobacteria are quite rare within the Baelen Member (Dreesen et al. 1985, pl. 2, figs. 2–4, pl. 14, figs. 3–6, 8, pl. 18, fig. 1; Fig. 6a) and they are not easy to interpret palaeoecologically. Girvanella sensu lato is a morphotaxon and a taphotaxon, which occurs from intertidal through epibathyal environments and from the lowermost Cambrian through the Recent (Riding 1975; Feldmann and McKenzie 1998). Various girvanellaceans occur in the crinoidal grainstones, either as small filaments (of Girvanella ex gr. staminea Garwood, 1931, misinterpreted as Girvanella wetheredii by Dreesen et al. 1985), encrusting crinoids or as broader filaments (of G. ex gr. wetheredii Chapman, 1908, interpreted as Girvanella problematica by Dreesen et al. 1985). The latter Girvanella colonies appear as intraclasts probably reworked from shallower marine deposits.

Cyanobacteria and incertae sedis algae Girvanella, Serrisinella and Disonella? a Reworked Girvanella spp. G. ex gr. staminea Garwood, 1931 encrusting crinoid ossicles (left), and broader, loose filaments of G. ex gr. wetheredii Chapman, 1908. Sample Baelen-8-1. b, c Packstone with Serrisinella melekessensis (Kulik, 1957) Vachard, 1991 (micrograph b is detail of the left part of micrograph c). Sample Goé-Onstl.19-2. d Silty wackestone/grainstone with Disonella? sp. and oblique sections of Serrisinella melekessensis. Sample Goé-N-10-7
Tubular and globular incertae sedis protists
Serrisinella Vachard, 1991:
The test is small, segmented, cylindrical to high-conical, aspondyle, with numerous thin acrophore laterals or wall perforations. The wall is hyaline-microgranular. The terminal central aperture seems to have been functional (see the illustrations of Vachard 1994). The specimens of Serrisinella from the Baelen Member are probably the oldest representatives of this genus with respect to the stratigraphic distribution given by Vachard (1991). However, nothing is known in the early Famennian about the phylogeny and probable relationships of Serrisinella with Issinella Reitlinger, although this latter genus is present in the classical late Frasnian mud mounds of Belgium and northern France. Serrisinella is a very common and abundant genus in the entire late Famennian, including the latest Famennian, Strunian substage (Herbig and Mamet 2006; Vachard and Cózar 2010). Serrisinella occurs along the northern margins of the Rheic and Tethys oceans from Belgium and northern and southern France to Afghanistan and Tajikistan, as well as in the Urals (Russia). Serrisinella is described as eurytope in the Strunian carbonate platforms of Germany (Herbig and Mamet 2006), but it seems to be more common in the deep-water limestones of western Europe; even if, in the stratotype of La Serre (Montagne Noire, southern France), it is associated with oolitic and bioclastic elements reworked from a shallow carbonate platform (Vachard 1988; Flajs and Feist 1988; Legrand-Blain and Vachard 2005).
The specimens of Serrisinella occurring in the Baelen Member may be identified as Serrisinella melekessensis (Kulik, 1957) emend. Vachard, 1994. Numerous illustrations of this taxon have already been published (Dreesen et al. 1985, pl. 5, fig. 10, pl. 7, fig. 8, 13, pl. 8, figs. 1–2, as Pseudoissinella sp. or Issinella sp.; Vachard 1994, pl. 9, figs. 10, 12–14, 16; and see herein Fig. 6b, c).
Disonella Conil and Lys, 1964:
One single atypical specimen found during this study (Fig. 6d) is analogous to the species Issinella antis Saltovskaya, 1990 (p. 118, pl. 36, figs. 13–14) and to the specimens identified as Disonella lucens Conil and Lys, 1964 by Dreesen et al. (1985, pl. 3, figs. 1–3). For the latter, we agree with their assignment to Disonella, but with slight differences in shape and irregularity from the “true” D. lucens.
Dreesenulella Vachard, 1991 (= Baculella Conil and Dreesen in Dreesen et al., 1985):
The genus Baculella, pre-occupied, was renamed Dreesenulella by Vachard (1991). This nomen novum was used, because it was pre-occupied in the zoological kingdom. However, Baculella was originally described not as a plant or an animal but rather as a microproblematicum; consequently, it is ipso facto not a homonym of the xenophyophora Baculella Tendal and Hessler, 1977. Because Baculella Conil and Dreesen in Dreesen et al., 1985 is interpreted in this paper as a possible xenophyophore, the group to which belongs Baculella Tendal and Hessler, 1977, Dreesenulella is definitively validated herein with regard to the International Code of Zoological Nomenclature. Due to its identical wall microstructure, its contemporaneity, its common fossilisation modes in the Baelen Member and its morphology, Dreesenulella appears relatively closely related to Serrisinella, as evidenced by the illustration below (Fig. 7a). The best argument for the intimate relation between Dreesenulella and Serrisinella is shown by the specimen illustrated by Dreesen et al. (1985, pl. 7, fig. 6) and herein (Fig. 7b) displaying one segment that is of Serrisinella-type and the other of Dreesenulella-type. In contrast to Serrisinella, Dreesenulella is almost endemic of Baelen, where it can be very common. In the studied area, Dreesenulella is even absent from the Haversin section (Souverain-Pré Formation) (Fig. 1), where Serrisinella is very abundant. Except for a possible reference to Saccamminopsis carteri ukrainica Brazhnikova in the Famennian (Kuchelgin and Murzakaev horizons) of the Urals (Grozdilova 1973, p. 91, pl. 6, fig. 7), there are no other references about Dreesenulella in the palaeontological literature.

Incertae sedis algae Dreesenulella. a Silty algal wackestone/packstone affected by pressure solution (“peastone”). Dark stylocummulate separates Dreesenulella-rich facies (above) from Serrisinella-rich facies (below). Sample Goé-N-10-01. b Algal packstone with Dreesenulella showing a transition with Serrisinella (left, centre). Sample Goé-N-10a-04. c, d Micrographs of an algal packstone with Dreesenulella, showing specimens with three succeeding chambers. Samples Goé-Rive droite Falaise-SVP-01-02 (c), Goé-N-10a-13 (d)
Several authors have stressed the similarity in shape between Dreesenulella/Baculella and Saccamminopsis (e.g. Dreesen et al. 1985; Loeblich and Tappan 1987; Mamet 1991) but their walls are different. Moreover, the wall structure of Dreesenulella is in reality identical to that of Serrisinella. The close relationship between Dreesenulella/Baculella and Saccamminopsis is especially obvious when looking at groups of tests. Indeed, the connections between two or three tests of Dreesenulella/Baculella (see Dreesen et al. 1985, pl. 5, figs. 2–5, pl. 6 (pars), pl. 7, figs. 3–5; Fig. 7c, d) are quite identical to those of Saccamminopsis from the latest Viséan of southwestern Spain and the earliest Serpukhovian of southern France (as illustrated by Vachard and Cózar (2003, fig. 8.1–7) and Pille (2008 (unpublished PhD thesis, pl. 7, fig. 10)): these correspond to uniseriate clusters of chambers, the axis of which is not rectilinear. This connection type is known in agglutinated foraminiferal taxa as well, like several species of the so-called Reophax (e.g. Gutschick 1986; Kaminski and Gradstein 2005). Con-sequently, it is possible to establish a morphologic and perhaps even a phylogenetic relationship between the genera Pelosina, Saccamminopsis and Dreesenulella/Baculella. Nevertheless, this relationship implicates important mineralogical changes in the skeleton: agglutinated in Pelosina, calcitic in Dreesenulella/Baculella and aragonitic in Saccamminopsis. The latter shows the typical aspects of aragonitic skeletons recrystallized into calcitic micro-sparite, a phenomenon well known in other groups, such as the bryopsidales, dasycladales, gastropods and the ammonoids.
Unlike the assignment of Saccamminopsis by Vachard and Cózar (2003) to bryopsidale algae, due to its possible aragonitic wall and its aspondyle cortical siphons, we have currently some doubts about the assignment of Dreesenulella/Baculella to the algae, because of the following observations: (a) its original wall was calcitic, when this type of wall is very exceptional among the bryopsidales (Ries 2006); (b) despite all our subsequent studies, we have never found similar fossil examples within the bona fide algal classes; (c) the complete absence of well-preserved specimens showing the medullar filaments of this supposed bryopsidale; and (d) in contrast, Saccamminopsis and Dreesenulella/Baculella display morphologies analogous to those of pelosinid foraminifers, as first indicated by Nestell et al. (2009). Furthermore, the morphological relationships between tubular and globular pelosinids (Avnimelech 1952; Cedhagen 1993; Loeblich and Tappan 1987; Kaminski and Gradstein 2005; Nestell et al. 2009) are strongly reminiscent of those between Dreesenulella/Baculella and Serrisinella. Consequently, Dreesenulella/Baculella might represent a globular form of a pelosinid foraminifer (e.g. Pelosina variabilis sphaeriloculum Höglund), whereas Serrisinella is more similar to a tubular form of this group (e.g. Pelosina arborescens Pearcey). Both taxa, Serrissinella and Dreesenulella, occur together within the Baelen Member, which is analogous to the co-existence of both species of Pelosina in the Swedish coastal waters (Cedhagen 1993).
Concerning Saccamminopsis-like genera mentioned in the literature, it is quite possible that the Silurian microfossils described as Saccamminopsis are similar to Dreesenulella/Baculella, because of their clear hyaline wall. This is especially true for the Silurian Saccamminopsis described by Cummings (1952); Schallreuter (1983), and Niko and Hamada (1986) and for the so-called nodosariates of this age (Cummings 1952; Bykova 1956) as well as for other foraminifers or related Early Palaeozoic groups. We can infer, for example, that the agglutinated foraminifer Oxinoxis Gutschick, 1962 (especially Oxinoxis ligula described by Conkin and Conkin (1964, 1970); currently only known from the USA) also belongs to this group. Other “false” Early Palaeozoic Saccamminopsis were also described by Nestell et al. (2009) as belonging to the recent foraminifer genus Pelosina Brady, 1879. Similarly, the “Saccamminopsis” of the Silurian, as well as some species of the Ordovician foramifer Amphitremoidea (e.g. Nestell and Tolmacheva 2004; Nestell et al. 2009, 2011) could belong to the same protist group. The conspicuous changes in mineralogy of the test, i.e. from agglutinated to low Mg calcite and then into aragonite, allows to reconsider the systematic position of the Issinellales and Beresellales, and to link them to the protists, even if this assignment was questioned by Vachard and Cózar (2010). It is noteworthy that the morphological similarities between the Algospongia and some atypical foraminifera were already stressed by the same authors.
Due to some morphofunctional, diagenetic, ontogenetic and taphonomic observed criteria (e.g. elongate and globular shapes of the test, very high productivity, well preservation of the tests, absence of endolitic microborings, absence of attached epizoans), the microproblematical Serrisinella and Dreesenulella were very abundant in their biotopes and were rapidly accumulated and buried. Consequently, we suggest that Serrisinella and Dreesenulella inhabited the sediment-water interface, forming morphogroups A and B similar to those of the extant foraminifers, prior to be reworked by strong bottom currents and to form bioaccumulations at the origin of the Baelen Member mud mounds.
Other incertae sedis protists
A few kamaenid incertae sedis protists (Kamaena Antropov, 1967 or Subkamaena Berchenko, 1981 (see Dreesen et al. 1985, pl. 7, figs. 1–2)) have been observed within the Baelen Member as well. The specimen identified as Paratikhinella of Dreesen et al. (1985, pl. 3, fig. 28; see below) could represent an internal mould of a kamaenid. The slightly corroded “dasyclad segments” illustrated by Dreesen et al. (1985, pl. 14, fig. 2) remain still enigmatic; they might correspond to incertae sedis protists related to Eouraloporella Berchenko, 1981, that were able to live in disphotic zones (see Vachard and Cózar 2010). The so-called rhodophycean algal fragments illustrated by Dreesen et al. (1985, pl. 2, fig. 1, pl. 12, fig. 1) are probably fragments of bivalve or brachiopod shells.
Foraminifera
Unilocular foraminifera (?):
No true unilocular (= mono-thalamous) foraminifera have been observed in the carbonates of the Baelen Member, only pseudoforaminifera (Vachard 1994) of the class Parathuramminida. The specimens of “Bisphaera sp.” illustrated by Dreesen et al. (1985, pl. 3, figs. 4–5) are atypical and most probably correspond to oblique subaxial sections of Paracaligella sp. (see below). Moreover, the parathuramminids illustrated by Dreesen et al. (1985, pl. 14, fig. 6) belong to the genus Salpingothurammina, the palaeoecology and systematic assignment of which are still debatable (e.g. Vachard et al. 2014; Vachard 2016).
Bilocular foraminifera:
The specimen illustrated and identified as Paratikhinella? sp. or Eonodosaria? sp. by Dreesen et al. (1985, pl. 3, fig. 28) shows many similarities (in size and shape) with Paratikhinella insolita (Gutschick and Treckman, 1959), but it could also represent an internal mould of a kamaenid (e.g. Devonoscala Langer, 1979). On the other hand, numerous Paracaligella spp. seem to be present within the Baelen Member (see Dreesen et al. 1985, pl. 3, figs. 29–34). As indicated above, oblique sections of Paracaligella were probably identified as “Bisphaera sp.” by the same authors. The caligellids represent probably endofaunal biota in coarse-grained sediments (Vachard and Cózar 2004). This is reflected by their curved to sinusoidal shapes, as well-illustrated by Dreesen et al. (1985). Caligellid foraminifera are common since the Middle–Upper Devonian (e.g. Vachard 1994). For this period, two genera can be distinguished based on the type of wall, Caligella and Paracaligella.
Plurilocular foraminifera:
Calcareous plurilocular foraminifers became abundant and relatively diversified from the Middle Devonian onwards, especially after the Givetian revolution (Vachard et al. 2010). After this evolutionary stage, many groups disappeared due to the Frasnian–Famennian boundary extinction crisis. Nevertheless, a streptospiral taxon is probably at the origin of almost all the groups of the Carboniferous–Permian orders (“Tournayellida”, Endothyrida, Palaeo-textulariida and Fusulinida sensu Hance et al. 2011). Their common ancestor is Glomospiranella, the Givetian representatives of which seem to appear and flourish in the Urals and maybe in Siberia (Vachard 2016).
During the Tournaisian, Glomospiranella successively gave rise to all the Lituotubelloidea sensu Hance et al. (2011), and then to the Endothyrida, which, in turn, gave rise to the Fusulinida, which became the dominant foraminiferal order during the Carboniferous–Permian period.
As early as the Famennian, Glomospiranella gave rise to Septaglomospiranella and Brunsiina, with an increase in length of the pseudosepta. At the early–late Famennian transition, both lineages produced taxa with a complete septation at the end of the coiling: these have been called Neosepta-glomospiranella and Septabrunsiina. So far, the outcrop area of the Baelen mud mounds is one of the best places in the world to study these primitive septate forms (including Septabrunsiina baeleni and Baelenia gosseleti). Indeed, the taxon S. baeleni has introduced the double type of coiling (first streptospiral, then planispiral evolute) within the plurilocular taxa, as well as the supplementary deposits, displaying a very rapid development as with Baelenia and the quasiendothyroids. The latter may show some true chomata that will reappear only in the primitive fusulinids during the early Pennsylvanian, 40 million years later. The advantage of these supplementary deposits seems to be a mechanical reinforcement of the endofaunal test against the pressure of the enveloping sediment. In contrast, the presence of pseudochomata–chomata seems more related to an improvement in the attachment system of the pseudopodia and/or the channelizing of the embryos before their release.
Septabrunsiina baeleni Conil in Bouckaert et al., 1967, occurring within the Late marginifera Zone in the Vesdre area of eastern Belgium, is probably the oldest species of the genus Septabrunsiina, whose type species (Endothyra krainica Lipina, 1948) is rather middle to late Tournaisian in age. The species S. baeleni probably gave rise during the late Famennian to Septabrunsiina bertchogurica Reitlinger and Kulagina in Morozov et al., 1987 and then to the well-compressed Septabrunsiina krainica and Septabrunsiina kingirica (larger than S. baeleni). The latter are of Tournaisian age with a first occurrence in the black Avesnelles Limestone (Conil and Lys 1970). The species Septabrunsiina donica Lipina, 1965 belongs probably to the same group and is more regularly evolute. Nevertheless, the species S. aff. donica sensu Dreesen et al. (1985, pl. 3, fig. 16) is most probably a representative of S. baeleni.
Baelenia Conil and Lys, 1977 is a poorly known genus, although it was previously cited from Belgium, Greece, the northern Urals, South China and perhaps also from Canada (Conil and Lys 1977; Durkina 1984; Hance et al. 1993, 2011; Vachard and Clément 1994). The type species of Baelenia is Baelenia gosseleti (Conil in Bouckaert et al. 1967) first described from the Baelen Member. Its FAD in this locality is likely, because of the presence of transitional forms with S. baeleni. However, Eoquasiendothyra bella, a descendent of Baelenia, seems to occur as early as the marginifera Zone (Kulagina 2013, fig. 2, p. 267) indicating that (a) the phylogeny of the first quasiendothyrids is questionable; (b) the foraminifers in the mid-Famennian of Belgium possibly migrated from the southern Urals, as already suggested by Conil et al. (1986, p. 19), and that the southern Urals were the homeland of this migration. The last representatives of Baelenia have been found in the Avesnelles section in northern France (Conil and Lys 1968). As illustrated and discussed by Dreesen et al. (1985, pl. 3, figs. 6–27), a transition exists between S. baeleni and B. gosseleti in the Baelen Member based on a progressive development of the roof crust, grading to bilateral structures resembling chomata (etymologically, the raised banks of a river).
Furthermore, we found specimens of Septaglomospiranella? comblaini Conil and Lys, 1964, associated with numerous Serrisinella in the nodular limestones of the coeval Souverain-Pré Formation, within the Haversin section (Dinant Synclinorium; Fig. 1). The ecological conditions are obviously the same for both foraminiferal taxa.
In the Baelen Member, septabrunsiinoid foraminifers lived probably in situ in the core of mud mounds, infaunally within muddy bottom sediments; they constituted a morphogroup C.
On the other hand, the “silicified foraminifers” illustrated by Dreesen et al. (1985, pl. 4, figs. 1–8) are very interesting due to their apparently trochamminoid coiling and triseriate architecture. As these forms are unknown from the Devonian, they are probably teratogenic forms of Septabrunsiina and Rectoseptaglomospiranella, as already suggested by Conil in Dreesen et al. (1985). Teratogenic forms of foraminifers have been well studied in Mesozoic, Cenozoic and Recent seas, because of their importance with respect to sea pollution and/or the onset of mass extinction crises. Their presence within the Baelen mud mounds remains an enigma and needs further investigation.
We suggest that all the foraminifers described above, are endofaunal and belong to a type C morphogroup (according to Jones and Charnock 1985 and Murray et al. 2011). The micrograph shown in the Fig. 8a–c indicates that the facies with Septabrunsiina and that with Serrisinella are transitional and that the biotopes with foraminifers were adjacent to those of issinellids (see also Dreesen et al. 1985, pl. 15, fig. 3). Both biotopes could be located on either side of the FWWB: the issinellid biotopes in the photic zone and the endofaunal biotopes in the disphotic zone.

Foraminifers (Septabrunsiina baeleni ) with crinoids, Serrisinella and Kamaena, and solitary rugose corals. a Bioclastic wackestone with crinoids, ostracods and rare foraminifers (see top left). Sample Les Forges LF-7-2. b, c Recrystallized (slightly dolomitized) crinoidal grainstone (displaying limpid syntaxial rim cement around the crinoids) with relatively common plurilocular foraminifera, some Serrisinella, and kamaenaceans (Kamaena or Subkamaena sp.). c Samples Les Forges-2-02 (b). Les Forges LF-2-1 (c). d Recrystallized silty bioclastic packstone affected by pressure solution, with an oblique section of Neaxon? sp. Sample Goé-2
Various incertae sedis:
The specimen identified as cf. Rhodophyta (in Dreesen et al. 1985, pl. 1, fig. 1, pl. 12, fig. 1) is most probably a tangential section of a costulate shell, which belongs either to a bivalve or to a brachiopod. The specimens identified as cf. Salebra by Dreesen et al. (1985, pl. 5, fig. 9) represent most probably fragments of bryozoan zoaria. The “vermetid gastropods (serpuloids)” (Dreesen et al. 1985, pl. 7, figs. 10, 14, pl. 15, fig. 5) most probably represent microconchids, the palaeoecology of which seems to be variable, from intertidal to relatively deep subtidal. Small peritidal-lagoonal microconchid buildups have also been described by Dreesen and Jux (1995) from the late Famennian Evieux Formation (Fontin Member) in the Ourthe valley (Belgium).
Solitary rugose corals:
According to Dieter Weyer (personal communication, July 2016), the specimens identified as incertae sedis by Dreesen et al. (1985, pl. 2, figs. 5–8) correspond to isolated, unoriented and corroded sections of solitary rugose corals, which are impossible to identify at the generic level. Another corroded specimen is tentatively assigned to the genus Neaxon Kullmann, 1965 by Dieter Weyer (personal communication, July 2016) (Fig. 8d). Their poor preservation reflects transport, hampering the reconstruction of their primary environment. However, they have been transported over a relatively small distance only but long enough to destroy the wall of the calyx. However, diagenetic phenomena (selective dissolution) might be the cause of their poor preservation as well.
Crinoids:
They are particularly abundant within the Baelen Member (Dreesen et al. 1985, 2013; Fig. 9a–c) producing lenticular deposits of crinoid ossicles (crinoidal grainstones/rudstones) that interfinger with all of the other carbonate facies in the Baelen Member. They frequently display graded, reverse graded, cross bedding and even slumping, and have been interpreted as tempestites (Dreesen et al. 1985, pl. 12, fig. 1, pl. 15, fig. 2, pl. 18, figs. 1–4, pl. 19, figs. 1–2). Some of the crinoid grainstones display also shelter porosities and preferential shell orientation (Dreesen et al. 1985, pl. 12, fig. 1).

Crinoidal grainstones and silty packstones, affected by pressure solution (a–c) and mineralized hardgrounds (d). a Crinoidal grainstone (tempestite) strongly affected by pressure solution. Upper part shows unaffected bioclastic grainstone. Lower part shows corroded crinoid ossicles and dark stylocumulates. Sample Limbourg-Mali-encrinite-02. b–c Micrographs of a silty crinoid-rich packstone/grainstone and even rudstone affected by pressure solution, generating a so-called stylonodular fabric (not affected relicts resemble intraclasts). Samples Les Forges-8-01 (b) and Les Forges-8-02 (c). d Thin brown phosphoritized micro-stromatolitic hardground and intraclasts (top left) separating a bioclastic packstone/grainstone (below) from a very silty bioclastic packstone (above) affected by pressure solution. This sample yielded a relatively rich conodont fauna indicating a Late marginifera age as well as a diversified (silicified) ostracod assemblage (see Dreesen et al. 1985). Sample COM-1-hardground-01
Sponges:
Hexactinellid, tetractinellid and monaxone sponges can be locally abundant within particular carbonate micro-facies of the Baelen Member (Dreesen et al. 1985, pl. 10, figs. 1–17, pl. 11, figs. 1–17), more especially within spiculitic wackestones (Dreesen et al. 1985, pl. 13, figs. 2–3, pl. 14, fig. 1; Fig. 10b, c). Some of the sponge spicules have been tentatively assigned to the genus Arakespongia by K. Rigby (in Dreesen et al. 1985).

Mud mound core (including microbial-spiculitic mudstones and bioclastic wackestones) with reworked Dreesenulella, crinoids, sponge spicules and stromatactis-type fenestrae. a Bioclastic wackestone with crinoids (bottom, left) and Dreesenulella (top, right). Sample Hors-les-Portes-3-1. b, c Bioclastic (spiculitic) mudstone/wackestone with abundant sponge spicules and relatively common crinoid ossicles (especially in micrograph). Samples Hors-les-Portes-1-01 (b) and Hors-les-Portes-1-02 (c). d Red, microbial/bioclastic wackestone with stromatactis structures and bioclasts (including ostracods, crinoids, sponge spicules, reworked algae, etc.). Sample Les Forges (rouge) 01
Brachiopods:
They occur sporadically within the Baelen Member, both in the mud mound core and in the surrounding carbonate facies. Their scarcity could be related to the difficulties of extracting them from the carbonate matrix. The poor preservation of the collected material prevents a systematic study, but small- to medium-sized representatives of the orders Orthida?, Productida (Productina) and Spiriferida (Cyrto-spiriferidae?) have been recognised.
Ostracods:
Bless (in Dreesen et al. 1985) studied a single sample of silicified ostracod shells from the hardground, developed at the base of the Souverain-Pré Formation at Goé (see Figs. 2 and 4). Because of this silicification, the ostracod shells could be removed from the limestone by acid etching. The latter study showed the co-occurrence of both shallow-water- and basinal-type ostracod species in a diversified assemblage (ca. 15 species) dominated by the bairdiaceans. The hardground is a condensed deposit, which shows palaeontological condensation (proved by conodonts) and which might yield biota from different depositional environments that are not necessary coeval. However, until now, there is no detailed information available on the ostracod assemblages present within the Baelen Member sensu stricto.
Carbonate microfacies and diagenetic history
The dominant microfacies consists of “algal”, cryptalgal (stromatactis-bearing) and spiculitic carbonate mudstones or wackestones, with subordinate “algal” bindstones and floatstones. Lenticular crinoidal/foraminiferal packstones, grainstones and even rudstones intermittently interrupt the carbonate mudstones. The latter suggest that the mounds had reached wave base or that they had been affected by storm wave activity (Dreesen et al. 1985; Dreesen and Swennen 1998). The Baelen mound complex displays a conspicuous cyclic aspect, reflecting changes in carbonate microfacies and in detrital content. Both the basalmost and topmost lithological units consist of nodular or lenticular bioclastic wackestones, embedded in calcareous micaceous sandstones. The core of the complex consists of sand-free red-stained cryptalgal-spiculitic mudstones with numerous stromatactis and LF-fabrics. The transitional units to the core are composed of “algal” mudstones and wackestones enclosing numerous crinoidal packstone/grainstone/rudstone lenses. Due to strong pressure solution, the actual grainstone/mudstone ratio is exaggerated with respect to the original mud mound composition. The original thickness of the individual mounds may well have exceeded 150 m (Dreesen et al. 1985; Dreesen and Swennen 1998).
The core of the Baelen Member (cryptalgal-spiculitic mudstones) displays characteristics of rather relatively deeper, open-marine depositional environments (e.g. Dreesen et al. 1985, pl. 12, fig. 3; pl. 13, figs. 1, 3, pl. 14, fig. 4; Fig. 10a–d), in particular the red-stained microbial carbonate mudstones and wackestones rich in stromatactis-type features (Fig. 10d) and so-called zebra structures. The descriptions and interpretations of the genesis of stromatactis in the literature are quite abundant (see for instance Bourque and Boulvain 1993). However, we now agree with the model that stromatactis has no organic origin but that is has rather been generated by taphonomic processes (e.g. remodelling of open spaces left after dissolution of sponges or other biota in incompletely lithified sediments) and that they are of early diagenetic origin (e.g. Wilson 1975; Burchette 1981; Hladil 2005; Olchowy 2011). The red colour of the stromatactis-bearing limestones is due to the presence of finely disseminated hematite and is generally related to bacterial-microbial activities in relative deep marine environments (Monty et al. 1982; Boulvain 1989; Boulvain et al. 2001; Mamet and Préat 2006), or at least in the endostromatoliths (Monty 1982, 1995; Boulvain et al. 2001) mostly associated with these “deep-water” carbonates.
The diagenetic history of the Baelen Member is complex (Dreesen et al. 1985; Belmans 1992 (unpublished MS thesis)) and reveals succeeding marine phreatic, mixed water, meteoric phreatic as well as burial diagenetic conditions, all subsequently overprinted by Variscan tectonics. Stable carbon and oxygen isotope data for different bioclasts and individual diagenetic phases (e.g. radiaxial fibrous marine cements in stromatactis, etc.) all plot within the same field. The carbon signature clearly suggests that the system has been buffered by the host rock, whilst the oxygen signature points to a resetting of the oxygen isotope signal by hot late diagenetic fluids.
Bathymetry
The inferred depositional environments of the various biota described in the previous paragraphs might represent additional lines of evidence for the open marine depositional setting (carbonate ramp) of the mid-Famennian Baelen Member, as suggested earlier by the same authors (Dreesen et al. 1985, 2013, 2015b; Dreesen and Swennen 1998).
The sudden appearance during the mid-Famennian of microbial-spiculitic carbonates containing stromatactis and zebra structures within an overall siliciclastic setting (showing overwhelming sedimentological evidence pointing to a shallow epicontinental shelf with strong longshore and tidal currents; Thorez et al. 1988, 2006) is really puzzling. Moreover, as the location of these mud mounds seems to be strongly related to the presence of block faults (e.g. Marion 1985; Dreesen et al. 2013), a genetic link with synsedimentary tectonic activity is obvious (see Fig. 5). Furthermore, slumps have been described from the upper parts of the Baelen mud mounds, possibly related to synsedimentary tectonic activity as well, along the above-mentioned faults (Dreesen et al. 1985, 2013, 2015b; Aretz and Chevalier 2007).
Because of the sudden “en masse” appearance of so many specimens of only a few species of very particular “incertae sedis”-type protists, Dreesen et al. (2015b) suggested earlier that these Baelen mud mound complexes might possibly be related to the presence of cold (methane) seeps, linked with those block faults, a hypothesis that still needs to be proved. Due to strong tectonic disturbances (faulting), a complex diagenetic history (see Dreesen et al. 1985) and subsequent hydrothermal overprinting, the stable C- and O-isotopic signature of the Baelen microbial carbonates is blurred and needs to be investigated further, in order to test the above hypothesis (Dreesen and Swennen 1998). Furthermore, other groups of fossil organisms, including ostracods, brachiopods, orthoceratids, sponges and crinoids, need still to be studied in more detail in order to provide additional palaeoecological lines of evidence helping to assess the original depositional depths. Hebbeln and Samankassou (2015) recently pointed out that all the commonly used criteria to allocate a shelf origin of ancient carbonate mounds (e.g. high energy environments, micritization phenomena, particular non-biogenic components such as peloids, etc.), except the presence of in situ photoautotrophic organisms, are also applicable for carbonate mounds being formed well below the shelf break, down to water depths of −1000 m along many continental margins of the world. Furthermore, although benthic photoautotrophic organisms would be a clear indicator for shallow-water settings, for most of the organisms found in fossil carbonate mounds and being interpreted to be photoautotrophic, the taxonomic position and the metabolism are controversially discussed. Mud mounds are generally interpreted to have been deposited in deep water (above storm wave base) and still within the photic zone. The lower limits given for the bathymetry for ancient deep-water carbonate mounds mostly range between −30 and −280 m (Hebbeln and Samankassou 2015)
The taxonomic assignment of Serrisinella and Dreesenulella is thus crucial for the interpretation of the depositional depth of the Baelen Member. However, in the absence of an unambiguous statement about their biological affinity, we have to rely on other (indirect) sedimentological and palaeontological criteria. Circumstantial evidence provided by the overall depositional setting of the enveloping and lateral sediments, as well as by the lack of pelagic or hemi-pelagic organisms (e.g. goniatites), points towards a relative shallow shelf setting: the underlying micaceous siltstones and sandstones of the Esneux Formation have been interpreted as distal subtidal deposits enclosing tempestites, whereas the overlying micaceous siltstones and sandstones of the Comblain-la-Tour Formation have been interpreted as open marine subtidal deposits (Thorez and Dreesen 1986; Thorez et al. 1988). The Souverain-Pré Formation (of which the Baelen mounds represent a local member) would correspond to a nearshore subtidal environment, an interpretation also supported by the presence of a conodont fauna characteristic of a mixed palmatolepid-polygnathid and polygnathid-icriodid biofacies (Dreesen and Thorez 1980) as well as by the presence of other shallow-marine biota, such as incertae sedis protists (kamaenids, issinellids), endothyrid-tournayellid foraminifers, nearshore ostracodes (e.g. Cryptophyllus) and small tabulate and rugose corals (e.g. Cladochonus and Catactotoechus; Denayer et al. 2012). With the exception of the autochthonous carbonates of the Baelen Member (Souverain-Pré Formation), the lagoonal-peritidal limestones of the Fontin Member (Evieux Formation) and the biostromal boundstones of the Comblain-au-Pont Formation, all other Famennian carbonates in Belgium are parautochthonous-allochthonous (Dreesen and Thorez 1994). The parautochthonous nodular limestones embedded in a micaceous sandstone matrix of the Souverain-Pré Formation have been interpreted as the result of the mechanical reworking of semi-consolidated shallow-marine carbonate ramp sediments (Thorez et al. 2006).
In the autochthonous carbonates of the Baelen Member, conodonts are extremely rare or even missing with the exception of a poor polygnathid conodont fauna extracted from some of the interfingering subautochtonous crinoidal grainstones (tempestites) (Dreesen et al. 1985).
Thus, typical mid-Famennian biota indicative for hemi-pelagic or pelagic settings (such as goniatites, conodonts of the palmatolepid biofacies and Thuringen-type ostracodes, the latter of which are characteristic of slope and basins), are totally lacking in the Baelen Member. Hence, shallower depths have to be considered for interpreting the depositional environment of the Baelen Member.
Taxonomic assignment of incertae sedis protists and foraminifera
The massive appearance of specimens of Serrisinella in the lower part of the Baelen Member is quite conspicuous and points to a particular sedimentological mechanism. Indeed, these Serrisinella colonise areas subject to strong bottom currents. So, strong current activity might generate “super-packstones” in which 70% of the bioclasts correspond to well-sorted specimens of oligotypic Serrisinella (Fig. 7a). During their life, Serrisinella grew upright in the bottom sediment whilst projecting their pseudopodia above the sediment/sea-water interface. The alternations with Dreesenulella may correspond to different compositions of the muddy bottom, nutrient influx, seasonal alternations or to periods of stronger current activity, where the morphogroups B replace the morphogroups A. Due to their ampulla-like shape and their buoyancy, the dead tests of Dreesenulella and to a lesser extent those of Serrisinella can reach deeper or more distal sediments. This explains their presence within the supposedly relatively “deeper” (open marine–outer ramp) stromatactis-bearing limestones (Dreesen et al. 1985, pl. 8, fig. 1; Fig. 9a).
We are referring to a modern analogue, such as the modern foraminifera genus Pelosina (see Cedhagen 1993), stressing the possible co-existence of both tubular and globular tests. Similarly, this co-occurrence can also be suggested for the following protists: Serrisinella–Dreesenulella, Saccamminopsis and its supposed “sporophyte” of Vachard and Cózar (2003); the Ordovician “Pelosina” and “Hippocrepina” or “Hyperammina” (Nestell and Tolmacheva 2004; Nestell et al. 2009); the Ordovician and Silurian Saccamminopsis and Lagena (Cummings 1952; Bykova 1956; Schallreuter 1983; Niko and Hamada 1986); the Paleogene Pelosina, Caudammina, Kalamopsis and Pelosinella (Kaminski and Gradstein 2005); and the modern P. arborescens Pearcey and P. variabilis spheriloculum Höglund (Cedhagen 1993), this latter being the type species of Globosiphon Avminelech, 1952.
Finally, and as already proposed by Vachard and Cózar (2010), we suggest that Serrisinella and Baculella have many in common with atypical foraminifers such as the Xenophyophorea and Kokomiacea. However, even if the latter are currently assigned to the foraminifers (Pawlowski et al. 2003; Nestell et al. 2009), many authors consider them as belonging to another protist group (e.g. Schröder et al. 1989; Gooday 1994; Levin 1994; Gooday et al. 2007a, b; Lecroq et al. 2009; Kamenskaya et al. 2012). Furthermore, the differences of their evolutionary trends and histories with respect of those of the foraminifers (already stressed by Vachard and Cózar 2010) might indicate that this group could be really different from the foraminifers of the Ordovian (or earlier). This particular group might include all of the algosponges of Vachard and Cózar (2010).
The simultaneous presence of aragonitic, calcitic and agglutinated forms within the Baelen Member might indicate that all these incertae sedis organisms are more closely related to foraminifers than to the algae, because the latter generally display a more stable mineralogy during the geological history.
Due to the morphologic and palaeobiologic similarities between Dreesenulella and Saccamminopsis, the depositional environment of the Baelen Member might be comparable to that of the late Viséan mud mounds containing Saccamminopsis (e.g. Madi et al. 1996; Cózar et al. 2003; Rodríguez-Martínez et al. 2003; Flügel 2004; Vachard et al. 2010).
The bioaccumulations of pelosinids might be considered as “deep” marine because no associated shallow-water microfossils have been observed (Nestell et al. 2009). Furthermore, modern pelosinids are generally encountered in deep seas. However, Cedhagen (1993) mentioned less deep habitats, e.g. below 25 m. The pelosinids, saccamminopsids and dreesenulellids apparently favoured muddy substrates, forming meadows that were frequently affected by bottom currents producing rich bioaccumulations.
The plurilocular foraminifers tend to be preferentially concentrated within the crinoidal grainstones and rudstones, that have been interpreted as storm deposits or tempestites (Dreesen et al. 2013, 2015b). The abundant serrisinellids and dreesenulellids herein were most probably derived from crinoidal and Serrisinella meadows located on the inner part of the ramp (Fig. 5). Subsequently, they have been transported together with crinoid ossicles across the ramp to more offshore (and deeper) settings (middle-outer ramp).
From a foraminifer point of view, the lower–middle Famennian (up to the latest marginifera Zone) representatives might appear quite special because they occur during a recovery period after a major biotic crisis. Moreover, they encompass almost exclusively unilocular genera (Parathurammina, Neoarchaesphaera, Eotuberitina, Diplos-phaerina, Bisphaera; Poyarkov 1969; Hance et al. 1993; Timokhina et al. 2002). Nonetheless, it is noteworthy that some late Devonian carbonate ramps, such as those in Belgium and the Urals, yielded bilocular and pseudoseptate foraminiferal genera including Earlandia, Caligella, Eotournayella, Glomospiranella, Septaglomospiranella ex gr. nana, Brunsiina and the first Septabrunsiina ex gr. baeleni; even the last representatives of the uniseriate Tikhinella subsist during this period (Vachard 1974; Conil in Bouckaert et al. 1967; Dreesen et al. 1985; Kalvoda 1999, 2001, 2002; Timokhina et al. 2002; Vachard et al. 2010).
Despite the fact that modern-looking morphotypes of foraminifera (especially those with trochospiral and triseriate tests) are lacking during the Palaeozoic (Vachard et al. 2010), which might point to a lack or scarcity of epiphyte forms, several modern-type morphogroups of foraminifers are already present during the Palaeozoic. The term “morphogroup” refers here to a general grouping of foraminifers of similar shapes or growth patterns, independently of their exact taxonomy and of the existence of possible homeomorphs. Morphogroups offer a way of overcoming taxonomic differences and thereby making comparisons possible between assemblages of different geological ages (Murray et al. 2011). Consequently, they may been used for reconstructing palaeoenvironments (e.g. Chamney 1976; Jones and Charnock 1985; Murray 1991; Nagy 1992; Kaminski et al. 1995; Nagy et al. 1995; Jones 1999; Preece et al. 1999; Van Den Akker et al. 2000; Mancin 2001; Jones et al. 2005; Kender et al. 2008a, b; Cetean et al. 2011). As the morphology of the foraminiferal test is directly controlled by morphofunctional factors, including nutrient strategy and life position (Tyszka 1994; Coccioni et al. 1995; Reolid et al. 2008), the same morphogroups are present from the Devonian through modern environments (Coccioni et al. 1995, with references therein). In our material, the foraminiferal morphogroups B and C of Jones and Charnock (1985); morphogroups A-1 and A-2 of Coccioni et al. (1995); morphogroups ED1, ED2 and ED4 of Holcová and Slavík (2013); and morphogroups A and B1 of Murray et al. (2011) are present within the Baelen Member. The morphogroup A of Jones and Charnock (1985) was not found, but it is probably replaced by the Serrisinella meadows. Moreover, Dreesenulella may belong to morphogroup B because of its form similar to that of the foraminifer Saccammina. Morphogroup C is represented by Septabrunsiina. Morphogroup D (epiphytic) is uncommon during the Palaeozoic; within the Baelen Member, it is lacking or possibly replaced by some rare Disonella? and/or microconchids.
The biological affinity of Serrisinella and Dreesenulella is still a matter of discussion. This particular group might well be assigned to a protist group that is closely related to the foraminifers. A possible algal origin was initially suggested by Vachard and Cózar (2010). The major argument for this assignment was their huge productivity, especially due to the supposed disphotic (if not aphotic) biotope of these “algospongia”. However, the present study shows that this huge productivity or abundance might also be related to abiotic phenomena or processes, such as the hypothetical presence of cold methane seeps or the effect of bottom currents. Another evidence, used by Vachard and Cózar (2003) to assign Saccamminopsis to the algae, was the aragonitic nature of the wall and the presence of small utricles (i.e. the last order of ramifications of the siphons through the wall). Nevertheless, Saccamminopsis shows analogies with Dreesenulella, displaying a calcitic wall with numerous randomly arranged microperforations that are morphologically different from the utricles of the bryopsidales or the laterals of the dasycladales. We conclude therefore that all evidence for an algal origin of Serrisinella and Dreesenulella is still controversial. In contrast, a protist assignment seems to be more plausible. As foraminifers are excluded as well (according to Vachard and Cózar 2003, 2010), their affiliation to another protist group such as that of the modern Xenophyophora or the Komociacea (enigmatic deep-sea (nowadays) protists related to agglutinated foraminifera, commonly called “giant protists”) is suggested here. It is noteworthy that the modern genus Baculella has also been considered as belonging to the Komokiacea (Tendal and Hessler 1977). Consequently, both the Palaeozoic and modern species of Baculella could be Komokiacea and the name Dreesenulella for the fossil genus is hereby definitively validated.
Conclusions
The Baelen Member of the Souverain-Pré Formation (Late marginifera conodont Zone, middle Famennian) was deposited at the end of a biological recovery period that succeeded the Frasnian–Famennian boundary mass extinction, which is well documented in Belgium (see review in Mottequin and Poty 2016). During this crisis, many builders and benthic groups were eliminated, including colonial corals, stromatoporoids, etc. Rugose and tabulate corals were particularly rare during the lower Famennian (Denayer et al. 2012) in Belgium, and they occur occasionally within the Souverain Formation, including the Baelen Member. The foraminiferal and algal biota found in the Baelen Member are completely renewed with respect to the biota of the Frasnian.
During the mid-Famennian, a carbonate ramp temporarily developed in the Dinant and Vesdre Synclinoria, encompassing large parts of the Famennian epicontinental siliciclastic shelf (essentially composed of micaceous silt- and sandstones constituting the Condroz Group). Exceptionally, coeval autochthonous carbonates originated on the outer ramp, leading to the development of carbonate mounds in a rather restricted geographical area and in a cluster indicating the influence of block faults. The initial triggering mechanism of the Baelen mounds still remains uncertain. Initially, crinoids, issinellids and hexactinellid sponges might have lowered the current velocity by baffling and trapping lime mud. Subsequently, skeletal grains might have been fixed in place (by cyanobacteria?) so that the mud banks developed as a self-propagating system. Sedimentological and palaeoecological lines of evidence point to an open shelf setting (carbonate ramp environment) below or near the wave base but apparently still within the photic zone. Other workers (e.g. Aretz and Chevalier 2007) also suggested that the Baelen mud mounds were located between the base of wave activity and that of storm wave activity. The inner ramp was probably occupied by meadows of crinoids and incertae sedis tubular and globular protists (Serrisinella and Dreesenulella). Plurilocular foraminifera, belonging to the endofaunal morphogroup C, apparently lived in their bottom sediment. Bryozoans, brachiopods, conodonts and ostracods lived in the same environment. Microconchids and girvanellaceans have been derived from more nearshore or restricted marine settings and have been transported by currents across the ramp to become mixed with the other in situ biota. Episodically, inner ramp, bioclastic sediments were reworked by bottom currents or storm surges, producing important accumulations of Serrisinella, Dreesenulella and crinoids, including tests of foraminifera. On the supposedly deepest parts of the outer ramp, relatively pure (siliciclastic-free) red-stained microbial-spiculitic mudstones with Stromatactis-like fenestrae formed below the SWB. In the most distal parts of the shelf, coeval siltstones and silty shales were deposited sporadically enclosing thin lenticular crinoidal grainstones (distal tempestites) the bioclasts of which were derived from the Baelen carbonate mounds. The presence of these massive, “pure” siliciclastic-free microbial-spiculitic mudstones/wackestones (Baelen mounds core) could point towards a temporary deepening, possibly related to subsidence of faulted blocks and/or that the mounding sites have been shifted to settings beyond the reach of currents and/or that the siliciclastic supply was temporarily blocked by the same synsedimentary tectonic activity.
The taxonomical reinterpretation of Serrisinella and Dreesenulella, presented herein, adds to the already existing (sedimentological–palaeoecological) evidence for a rather open marine environment of the Baelen mud mounds, more especially in a supposed mid- and/or outer ramp setting. Both the assignment to algae or to atypical foraminifera would favour the above idea, excluding however pelagic or hemi-pelagic environments.
Change history
09 February 2017
An erratum to this article has been published.
References
Antropov, I. A. (1967). Vodorosli devona i nizhnego karbona (Turne) tsentralnoi chasti vostoka Russkoi Platformy (Algae from the Devonian and Early Carboniferous (Tournaisian) of the centre of the eastern part of the Russian Platform). Akademiya Nauk SSSR, Akademiya Nauk SSSR, Sibirskoe Otdelenie, Trudy Instituta Geologii i Geofiziki, 118–125 (in Russian).
Aretz, M., & Chevalier, E. (2007). After the collapse of stromatoporid–coral reefs—the Famennian and Dinantian reefs of Belgium: much more than Waulsortian mounds. In J. J. Álvaro, M. Aretz, F. Boulvain, A. Munnecke, D. Vachard, & E. Vennin (Eds.), Palaeozoic reefs and bioaccumulations: climatic and evolutionary controls (pp. 163–188). Geological Society, London, Special Publications, 275.
Avnimelech, M. (1952). Revision of the tubular Monothalamia. Contribution from the Cushman Foundation for Foraminiferal Research, 3, 60–68.
Belanger, I., Delaby, S., Delcambre, B., Ghysel, P., Hennebert, M., Laloux, M., Marion, J.-M., Mottequin, B., & Pingot, J.-L. (2012). Redéfinition des unités structurales du front varisque utilisées dans le cadre de la nouvelle Carte géologique de Wallonie (Belgique). Geologica Belgica, 15, 169–175.
Belmans, H. (1992). Sedimentologische en diagenetische studie van het “rode rif van Baelen” in het synclinorium van Verviers. Unpublished master thesis, K.U. Leuven, 90 p.
Berchenko, O.I. (1981). Izvestkovye vodorosli turneiskikh otlozhenii Donbassa (Calcareous algae from Tournaisian deposits of Donbass). Akademiya Nauk Ukrainskoi SSR, Institut Geologicheskikh Nauk “Naukova Dumka”, 1–72 (in Russian).
Béthune, P. De (1954). Carte géologique de Belgique (échelle 1/500.000). Atlas de Belgique, planche 8. Brussels: Académie royale de Belgique.
Bouckaert, J., Conil, R., & Thorez, J. (1967). Position stratigraphique de quelques gîtes famenniens à foraminifères. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 75, 159–184.
Boulvain, F. (1989). Origine microbienne du pigment ferrugineux des monticules du Frasnien de l’Ardenne. Annales de la Société Géologique de Belgique, 112, 79–85.
Boulvain, F., De Ridder, C., Mamet, B., Préat, A., & Gillan, D. (2001). Iron microbial in Belgian Frasnian carbonate mounds. Facies, 44, 47–60.
Bourque, P. A., & Boulvain, F. (1993). A model for the origin and petrogenesis of the red Stromatactis limestone of Paleozoic carbonate. Journal of Sedimentary Petrology, 63, 607–619.
Brady, H. B. (1879). Notes on some of the Reticularian Rhizopoda of the Challenger Expedition. Part 1. On new or little known arenaceous types. Quarterly Journal of Microscopical Science, New Series, 19, 20–63.
Burchette, T.V. (1981). European Devonian reefs: a review of current concepts and models. In D. F. Toomey (Ed.), European fossil reef models (pp. 85–142). Society of Economic Paleontologists and Mineralogists, Special Publication 30.
Bykova, E. V. (1956). Foraminifery Ordovica i Silura Sovetskoy Pribaltiki (Ordovician and Silurian foraminifers from the Soviet Prebaltic). Mikrofauna SSSR, 8(98), 6–37.
Cedhagen, T. (1993). Taxonomy and biology of Pelosina arborescens with comparative notes on Astrorhiza limicola (Foiraminiferida). Ophelia, 37(2), 143–162.
Cetean, C. G., Bălc, R., Kaminski, M. A., & Filipescu, S. (2011). Integrated biostratigraphy and palaeoenvironments of an upper Santonian–upper Campanian succession from the southern part of the Eastern Carpathians, Romania. Cretaceous Research, 32, 575–590.
Chamney, T. P. (1976). Foraminiferal morphogroup symbol for paleoenvironmental interpretation of drill cutting samples: Arctic America. Maritime Sediments Special Publication, 1B, 585–624.
Chapman, F. (1908). On the relationship of the genus Girvanella, and its occurrence in the Silurian Limestones of Victoria. Report of the Australasian Association for the Advancement of Science, 1907, 377–386.
Coccioni, R., Galeotti, S., & Gravili, M. (1995). Latest Albian–earliest Turonian deep–water agglutinated foraminifera in the Botaccione section (Gubbio, Italy)—biostratigraphic and palaeoecologic implications. Revista Española de Paleontología, N° homenaje al Dr. Guillermo Colom, 135–152.
Conil, R., & Lys, M. (1964). Matériaux pour l’étude micropaléontologique du Dinantien de la Belgique et de la France (Avesnois). Pt. 1, Algues et foraminifères; Pt. 2, Foraminifères (suite). Mémoires de l’Institut de Géologie de l’Université de Louvain, 23, 1–372.
Conil, R., & Lys, M. (1968). Utilisation stratigraphique des foraminifères du Dinantien. Annales de la Société Géologique de Belgique, 91, 491–558.
Conil, R., & Lys, M. (1970). Données nouvelles sur les foraminifères du Tournaisien inférieur et des couches de passage du Famennien au Tournaisien dans l’Avesnois. Congrès et Colloques de l’Université de Liège, 55, 241–265.
Conil, R., & Lys, M. (1977). Les transgressions dinantiennes et leur influence sur la dispersion et l’évolution des foraminifères. Mémoires de l’Institut Géologique de l’Université de Louvain, 29, 9–55.
Conil, R., Dreesen, R., Lentz, M. A., Lys, M., & Plodowski, G. (1986). The Devono–Carboniferous transition in the Franco–Belgian basin with reference to Foraminifera and Brachiopods. Annales de la Société Géologique de Belgique, 109, 19–26.
Conkin, J. E., & Conkin, B. M. (1964). Devonian Foraminifera: part I. The Louisiana Limestones of Missouri and Illinois. Bulletin of American Paleontology, 47(213), 49–105.
Conkin, J. E., & Conkin, B. M. (1970). North American Kinderhookian (Lower Mississippian) arenaceous foraminifers. Sixième Congrès International de Stratigraphie et de Géologie du Carbonifère, Sheffield 1967. Compte Rendu, 2, 575–583.
Cózar, P., Rodríguez-Martínez, M., Falces, S., Mas, R., & Rodríguez, S. (2003). Stratigraphic setting in the development of microbial mud mounds of the Lower Carboniferous of the Guadiato area (SW Spain). In W. M.Ahr, P. M. Harris, William A. Morgan, & I. D. Somerville (Eds.), Permo-Carboniferous carbonate platforms and reefs (pp. 57–67). Society of Economic Paleontologists and Mineralogists, Special Publication, 78 and AAPG Memoir, 83.
Cummings, R. H. (1952). Saccamminopsis from the Silurian. Proceedings of the Geologist’s Association, 63, 220–226.
Denayer, J., Poty, E., Marion, J.-M., & Mottequin, B. (2012). Lower and Middle Famennian (Upper Devonian) rugose corals from southern Belgium and northern France. Geologica Belgica, 15, 273–283.
Dreesen, R. (1987). Oolitic ironstones as event-stratigraphical marker beds within the Upper Devonian of the Ardenno-Rhenish Massif. In T. Young & W. Taylor (Eds.), Phanerozoic ironstones (pp. 65–78). Geological Society, London, Special Publications, 46.
Dreesen, R. (1989). Oolitic ironstones as event-stratigraphical marker beds within the Upper Devonian of the Ardenno–Rhenish Massif. Special Publications Geological Society London, 46, 65–78.
Dreesen, R., & Jux, U. (1995). Microconchid buildups from Late Famennian peritidal-lagoonal settings (Evieux Formation, Ourthe valley, Belgium). Neues Jahrbuch für Geologie und Palaeontologie, Abhandlungen, 198, 107–121.
Dreesen, R., & Swennen, R. (1998). Sedimentology and diagenesis of the Mid-Famennian Baelen mud mounds. In De Mol, B. (Ed.), Geosphere-biosphere coupling: carbonate mud mounds and cold water reefs. International conference and sixth post-cruise meeting of the training through research programme, UNESCO (pp. 52–53). Intergovernmental Oceanographic Commission Workshop Report, 143.
Dreesen, R., & Thorez, J. (1980). Sedimentary environments, conodont biofacies and paleoecology of the Belgian Famennian (Upper Devonian)—an approach. Annales de la Société Géologique de Belgique, 103, 97–110.
Dreesen, R., & Thorez, J. (1994). Parautochthonous-allochthonous carbonates and conodont mixing in the Late Famennian (Uppermost Devonian) Condroz Sandstones in Belgium. Courier Forschungsinstitut Senckenberg, 168, 159–182.
Dreesen, R., Bless, M. J. M., Conil, R., Flajs, G., & Laschet, C. (1985). Depositional environment, paleocology and diagenetic history of the “Marbre rouge à crinoïdes de Baelen” (Late Upper Devonian, Verviers Synclinorium, Eastern Belgium). Annales de la Société Géologique de Belgique, 108, 311–359.
Dreesen, R., Marion, J.-M., & Mottequin, B. (2013). The Red Marble of Baelen, a particular historical building stone with global geological importance and local use. Geologica Belgica, 16, 179–190.
Dreesen, R., Marion, J.-M., & Mottequin, B. (2015a). Het Rode marmer van Baelen, een geologisch buitenbeetje. Grondboor en Hamer, 69, 2–13.
Dreesen, R., Vachard, D., Marion, J.-M., & Mottequin, B. (2015b). The Red Marble of Baelen, an exceptional mid-Famennian mud mound complex in a carbonate ramp setting of eastern Belgium. Strata, Série, 1(16), 46–47.
Durkina, A.V. (1984). Foraminifery pogranichnykh otlozhenii devona i karbona Timano–Pechorskoy provintsii (Foraminifers of the Devonian–Carboniferous boundary deposits of the Timan–Pechora province). Nedra, Leningradskoe Otdelenie, 1–138 (in Russian).
Dvorak, J. (1973). Die Quer-Gliederung des Rheinischen Schiefergebirges und die Tektigenese des Siegener Antiklinoriums. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 143, 133–152.
Fagerstrom, J. A. (1994). The history of Devonian-carboniferous reef communities: extinctions, effects, recovery. Facies, 30, 177–192.
Feldmann, M., & McKenzie, J. A. (1998). Stromatolite–thrombolite associations in a modern environment, Lee Stocking Island, Bahamas. Palaios, 13, 201–212.
Flajs, G., & Feist, R. (1988). Index conodonts, trilobites and environment of the Devonian-Carboniferous Boundary beds at La Serre (Montagne Noire, France). Courier Forschungsinstitut Senckenberg, 100, 53–147.
Flügel, E. (2004). Microfacies of carbonate rocks, analysis, interpretation and application. Berlin: Springer.
Garwood, E. J. (1931). The Tuedian Beds of northern Cumberland and Roxburghshire east of the Liddel Water. Geological Society London Quarterly Journal, 87, 97–156.
Ghysel, P., Laloux, M., Dejonghe, L., Hance, L., & Geukens, F. (1995). Carte géologique de la Wallonie 1:25.000: Limbourg–Eupen (n°43/5–6). Ministère de la Région Wallonne. Namur: Direction Générale des Ressources naturelles et de l’Environnement.
Gooday, A. J. (1994). The biology of deep-sea foraminifera: a review of some advances and their applications in paleooceanography. Palaios, 9, 14–31.
Gooday, A. J., Cedhagen, T., & Kamenskaya, O. E. (2007a). New and little-known Komokiacea (Foraminifera) from the bathyal and abyssal Weddell Sea and adjacent areas. Zoological Journal of the Linnean Society, 151, 219–251.
Gooday, A. J., Cedhagen, T., Kamenskaya, O. E., & Cornelius, N. (2007b). The biodiversity and biogeography of kokopmiaceans and other enigmatic foraminiferan-like protists in the deep Southern Ocean. Deep Sea Research II, 54, 1691–1719.
Grozdilova, L.P. (1973). Foraminifers in the study of the Tournaisian sections of the Bashkiria Mountains. In V. M. Posner (Ed.), Foraminifers in the study of the Tournaisian sections of the southern Urals (pp. 68–101). Trudy VNIGRI, Leningrad, 346 [In Russian].
Gutschick, R. C. (1962). Arenaceous foraminifera from oncolites in the Mississippian Sappington Formation. Journal of Paleontology, 36, 1291–1304.
Gutschick, R. C. (1986). Middle Ordovician agglutinated foraminifera including Reophax from the Mifflin Formation, Platteville Group of Illinois. Journal of Paleontology, 60, 233–248.
Gutschick, R. C., & Treckman, J. F. (1959). Arenaceous foraminifera from the Rockford Limestone of northern Indiana. Journal of Paleontology, 23, 229–250.
Hance, L., Muchez, P., Coen, M., Fang, X.–. S., Groessens, E., Hou, H., Poty, E., Steemans, P., Streel, M., Tan, Z., Tourneur, F., van Steenwinkel, M., & Xu, S.-C. (1993). Biostratigraphy and sequence stratigraphy at the Devonian–Carboniferous transition in southern China (Hunan Province). Comparison with southern Belgium. Annales de la Société géologique de Belgique, 116, 359–378.
Hance, L., Hou, H.–F., & Vachard, D. (with the collaboration of Kalvoda, J., Devuyst, F.-X., Poty, E., & Wu, X.) (2011). Upper Famennian to Visean foraminifers and some carbonate microproblematica from South China—Hunan, Guangxi and Guizhou. Beijing: Geological Science Press.
Hebbeln, D., & Samankassou, E. (2015). Where did ancient carbonate mounds grow—in bathyal depths or in shallow shelf seas? Earth-Science Reviews, 145, 56–65.
Herbig, H. G., & Mamet, B. (2006). A muddy to clear carbonate ramp, latest Devonian, Velbert Anticline (Rheinisches Schiefergebirge, Germany). Geologica et Palaeontologica, 40, 1–25.
Hladil, J. (2005). The formation of stromatactis-type fenestral structures during the sedimentation of experimental slurries—a possible clue to a 120-year-old puzzle about stromatactis. Bulletin of Geosciences, 80, 193–211.
Holcová, K., & Slavík, L. (2013). The morphogroups of small agglutinated foraminifera from the Devonian carbonate complex of the Prague Synform (Barrandian area, Czech Republic). Palaeogeography, Palaeoclimatology, Palaeoecology, 386, 210–224.
Jones, R.W. (1999). Forties Field (North Sea) revisited; a demonstration of the value of historical micropalaeontontological data. In R. W. Jones, & M. D. Simmons (Eds.), Biostratigraphy in production and development geology (pp. 185–200). Geological Society, London, Special Publication, 152.
Jones, R. W., & Charnock, M. A. (1985). Morphogroups of agglutinated foraminifera, their life positions and feeding and potential applicability in (paleo)ecological studies. Revue de Paléobiologie, 4(2), 311–320.
Jones, R.W., Pickering, K.T., Bou Dagher-Fadel, M., & Matthews, S. (2005). Preliminary observations on the micropalaeontological characterisation of submarine fan/channel sub-environments, Ainsa system, south-central Pyrenees, Spain. In A. J. Powell, & J. B. Riding (Eds.), Recent developments in applied biostratigraphy (pp. 55–68). The Micropalaeontological Society, Special Publication, 1.
Kalvoda, J. (1999). Tournaisian–Lower Visean calcareous foraminifera. Abhandlungen der geologischen Bundesanstalt, 54, 135–145.
Kalvoda, J. (2001). Upper Devonian–Lower Carboniferous foraminiferal paleobiogeography and Perigondwana terranes at the Baltica-Gondwana interface. Geologica Carpathica, 52(4), 205–215.
Kalvoda, J. (2002). Late Devonian–Early Carboniferous foraminiferal fauna: zonations, evolutionary events, paleobiogeography and tectonic implications. Folia, 39, 1–213.
Kamenskaya, O. E., Melnik, V. F., & Gooday, A. J. (2012). Giant protists (xenophyophores and kokomiaceans) from the Clarion-Clipperton ferromanganese nodule field (eastern Pacific). Zhurnal Obshcheĭ Biologii, 73(5), 377–388.
Kaminski, M.A., & Gradstein, F.M. (2005). Atlas of Paleogene cosmopolitan deep-water agglutinated foraminifera. Grzybowski Foundation.
Kaminski, M.A., Boersma, A., Tyszka, J., & Holbourn, A. (1995). Response of deep-water agglutinated foraminifera to dysoxic conditions in the California borderland basins. In M. A. Kaminski, S., Geroch, & M. A. Gasinski (Eds.), Proceedings of the Fourth International Workshop on Agglutinated Foraminifera (pp. 131–140). Grzybowski Foundation, Special Publication, 3.
Kender, S., Kaminski, M.A., & Jones, R.W. (2008). Oligocene deep-water agglutinated foraminifera from the Congo Fan, offshore Angola: palaeoenvironments and assemblage distributions. In M. A. Kaminski, & R. Coccioni (Eds.), Proceedings of the Seventh International Workshop on Agglutinated Foraminifera (pp. 107–156). Grzybowski Foundation, Special Publication 13.
Kender, S., Kaminski, M. A., & Jones, R. W. (2008b). Early to middle Miocene foraminifera from the deep-sea Congo Fan, offshore Angola. Micropaleontology, 54, 477–568.
Kulagina, E. I. (2013). Taxonomic diversity of foraminifers of the Devonian–Carboniferous boundary interval in the South Urals. Bulletin of Geosciences, 88, 265–282.
Kulik, E. L. (1957). Nakhodka Rhabdoporella v verkhnedevonskikh otlozheniyakh Russkoi Platformy i Kazakhstana (Discovery of Rhabdoporella in the upper Devonian deposits of Russian Platform and Kazakhstan). Doklady Akademii Nauk SSSR, 112, 527–529 (in Russian).
Kullmann, J. (1965). Rugose Korallen der Cephalopodenfazies und ihre Verbreitung im Devon des südöstlichen Kantabrischen Gebirges (Nordspanien). Akademie des Wissenschaften und der Literatur, Abhandlungen der Mathematish-Naturwissenschaftliche Klasse, 1965(2), 595–621.
Langer, W. (1979). Neue karbonatische Microproblematica aus dem westdeutschen Devon. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 1979(12), 723–733.
Lecroq, B., Gooday, A. J., Cedhagen, T., Sabbatini, A., & Pawlowski, J. (2009). Molecular analyses reveal high levels of eukaryotic richness associated with deep-sea protists (Kokomiacea). Marine Biodiversity, 39, 45–55.
Legrand-Blain, M., & Vachard, D. (2005). Carbonifère (Mississippien et Pennsylvanien). Annales de la Société Géologique du Nord, 2ème Série, 11, 151–158.
Levin, L. A. (1994). Paleoecology and ecology of xenophyophores. Palaios, 9, 32–41.
Lipina, O.A. (1948). Foraminifery chernyshinskoi suity turneiskogo yarusa podmoskovnogo nizhnego karbona (Foraminifers of Chernyshin suite of the Tournaisian stage in the Submoscovite Basin). Akademiya Nauk SSSR, Trudy Instituta Geologicheskikh Nauk, 62, geologicheskaya seriya, 19, 251–259 (in Russian).
Lipina, O. A. (1965). Sistematika Turneiellid (Systematics of tournayellids). Akademiya Nauk SSSR, Geologicheskii Institut, 130, 1–115 (in Russian).
Loeblich, A. R., & Tappan, H. (1987). Foraminiferal genera and their classification. New York: Van Nostrand Reinhold.
Madi, A., Bourque, P. A., & Mamet, B. L. (1996). Depth-related ecological zonation of a Carboniferous carbonate ramp, upper Visean of Béchar Basin, western Algeria. Facies, 35, 59–80.
Mamet, B. (1991). Carboniferous calcareous algae. In R. Riding (Ed.), Calcareous algae and stromatolites (pp. 370–451). Berlin: Springer.
Mamet, B., & Préat, A. (2006). Iron-bacterial mediation in Phanerozoic red limestones: state of the art. Sedimentary Geology, 185, 147–157.
Mancin, N. (2001). Agglutinated foraminifera from the Epiligurian succession (Middle Eocene/Lower Miocene, Northern Apennines, Italy): scanning electron microscopic characterization and paleoenvironmental implications. Journal of Foraminiferal Research, 31(4), 294–308.
Marion, J.-M. (1985). La présence de mud mounds famenniens sur un site en block–faulting, à Baelen (Belgique). In FNRS Sédimentologie – Groupe de contact “Sédimentologie”, Liège, 28 mai 1985. Re Sedimentologica, 2, (2 p.).
Monty, C. L. V. (1982). Cavity or fissure-dwelling stromatolites (endostromatolites) from Belgian Devonian mud mounds. Annales de la Société Géologique de Belgique, 105, 305–344.
Monty, C. L. V. (1995). The rise and nature of carbonate mud-mounds: an introductory actualistic approach. In C. L. V. Monty, D. W. J. Bosence, P. H. Bridges, & B. R. Pratt (Eds.), Carbonate mud-mounds, their origin and evolution (pp. 11–48). Special Publications of the International Association of Sedimentologists, 23.
Monty, C., Bernet–Rollande, M. C., & Maurin, A. F. (1982). Re-interpretation of the Frasnian classical “reefs” of the southern Ardennes, Belgium (extended abstract). Annales de la Société Géologique de Belgique, 105, 339–341.
Morozov, S. G., Arkhipova, V. V., & Abramova, V. N. (1987). Fauna i biostratigrafiya pogranichnykh otlozhenii devona i karbona Berchogura (Mugodzhary) (Fauna and biostratigraphy of the Devonian–Carboniferous boundary deposits of Berchogura (Mugodzhary)). Moscow: Nauka Press (in Russian).
Mottequin, B., & Poty, E. (2016). Kellwasser horizons, sea-level changes and brachiopod–coral crises during the late Frasnian in the Namur–Dinant Basin (southern Belgium): a synopsis. In R. T. Becker, P. Königshof, & C. E. Brett (Eds.), Devonian climate, sea level and evolutionary events. Geological Society, London, Special Publications, 423, doi:10.1144/SP423.6.
Murray, J. W. (1991). Ecology and palaeoecology of benthic foraminifera. Hulow: Longman.
Murray, J. W., Alve, E., & Jones, B. W. (2011). A new look at modern agglutinated benthic foraminiferal morphogroups: their value in palaeoecological interpretation. Palaeogeography, Palaeoclimatology, Palaeoecology, 309, 229–241.
Nagy, J. (1992). Environmental significance of foraminiferal morphogroups in Jurassic North Sea deltas. Palaeogeography, Palaeoclimatology, Palaeoecology, 95, 111–134.
Nagy, J., Gradstein, F. M., Kaminski, M. A., & Holbourn, A.E. (1995). Foraminiferal morphogroups, paleoenvironments and new taxa from the Jurassic to Cretaceous strata of Thakkhola, Nepal. In M. A. Kaminski, S. Geroch, & M. A. Gasinski (Eds.), Proceedings of the Fourth International Workshop on Agglutinated Foraminifera (pp. 181–209). Grzybowski Foundation Special Publication, 3.
Nestell, G. P., & Tolmacheva, T. Y. (2004). Early Ordovician foraminifers from the Lava River Section, northwestern Russia. Micropaleontology, 50(3), 253–280.
Nestell, G. P., Mestre, A., & Heredia, S. (2009). First Ordovician Foraminifera from South America: a Darriwilian (Middle Ordovician) fauna from the San Juan Formation, Argentina. Micropaleontology, 55(4), 329–344.
Nestell, G., Heredia, S., Mestre, A., Beresi, M., & González, M. (2011). The oldest Ordovician foraminifers (Oepikodus evae conodont Zone, Floian) from South America. Geobios, 44, 601–608.
Niko, S., & Hamada, T. (1986). The Middle Silurian Foraminifera (Saccamminopsis) from the Gion-yama Formation, Central Kyushu. Scientific Papers of the College of Arts and Sciences, University of Tokyo, 36(2), 173–178.
Olchowy, P. (2011). Possible origin of stromatactis-like cavities in Upper Jurassic sediments from the Wielkanoc quarry near Gołcza (Kraków–Częstochowa Upland, southern Poland)—experimental studies. Facies, 57, 613–625.
Pawlowski, J., Holzmann, M., Fahrni, J., & Richardson, S. L. (2003). Small subunit ribosomal DNA suggests that Xenophyophorean Syringammina corbicula is a foraminiferan. Journal of Eukaryotic Microbiology, 50(6), 483–487.
Pille, L. (2008). Foraminifères et algues calcaires du Mississippien supérieur (Viséen supérieur-Serpukhovien): rôles biostratigraphique, paléoécologique et paléogéographique aux échelles locale, régionale et mondiale. Unpublished PhD thesis, Lille University, 3 vol., 1–226 +1–255+72 pl.
Poyarkov, B. V. (1969). Stratigrafiya i foraminifery devonskikh otlozhenii Tyan-Shanya (Stratigraphy and foraminifers from the Devonian deposits of Tian-Shan). Akademiya Nauk Kirgizskoi SSR, Institut Geologii, Upravlennie Geologii Kirgizskoi SSR, Izdatelstvo “Ilim”, Frunze, 1–186 (in Russian).
Preece, R. C., Kaminski, M. A., & Dignes, T.W. (1999). Miocene benthonic foraminiferal morphogroups in an oxygen minimum zone, offshore Cabinda. In N. R. Cameron, R. H. Bate, & V. S. Clure (Eds.), The oil and gas habitats of the South Atlantic (pp. 267–282). Geological Society, London, Special Publication, 153.
Reolid, M., Nagy, J., Rodríguez-Tovar, F. J., & Olóriz, F. (2008). Foraminiferal assemblages as palaeoenvironmental bioindicators in Late Jurassic epicontinental platforms: relation with trophic conditions. Acta Palaeontologica Polonica, 53(4), 705–722.
Riding, R. (1975). Girvanella and other algae as depth indicators. Lethaia, 8, 173–179.
Ries, J. R. (2006). Aragonitic algae in calcite seas: effect of seawater Mg/Ca ratio on algal sediment production. Journal of Sedimentary Research, 76, 515–523.
Rodríguez-Martínez, M., Cózar, P., Mas, R., & Rodríguez, S. (2003). Upper Viséan Saccamminopsis-sponge microbial mud mounds, Sierra de La Estrella, southwestern Spain. In W. M.Ahr, P. M. Harris, William A. Morgan, & I. D. Somerville (Eds.), Permo-Carboniferous carbonate platforms and reefs (pp. 189–200). Society of Economic Paleontologists and Mineralogists, Special Publication, 78 and AAPG Memoir, 83.
Saltovskaya, V. D. (1990). K morfologii i sistematike roda Issinella Reitlinger, 1954 (On morphology and systematics of the genus Issinella Reitlinger, 1954). In B. S. Sokolov, & I. T. Zhuravleva (Eds.), Iskopaemye problematiki SSSR (Fossil problematics of the USSR) (pp. 113–118). Akademiya Nauk SSR, Sibirskoe Otdelenie, Institut Geologii i Geofiziki, “Nauka”.
Schallreuter, R. E. L. (1983). Calcareous foraminifera from the Ordovician of Baltoscandia. Journal of Micropaleontology, 2, 1–6.
Schröder, C. J. S., Medioli, F. S., & Scott, D. B. (1989). Fragile abyssal foraminifera (including new Komokiacea) from the Nares Abyssal Plain. Micropaleontology, 35(1), 10–48.
Tendal, O. E., & Hessler, R. R. (1977). An introduction to the biology and systematics of Kokomiacea (Textulariina, Foraminiferida). Galathea Report, 14, 165–194.
Thorez, J., & Dreesen, R. (1986). A model of a regressive depositional system around the Old Red Continent as exemplified by a field trip in the Upper Famennian ‘Psammites du Condroz’ in Belgium. Annales de la Société Géologique de Belgique, 109, 285–323.
Thorez, J., & Dreesen, R. (1997). Sequence- and (bio) event-stratigraphy: a tool for global correlation in Euramerica, as exemplified by the late Upper Devonian in Belgium. 1st International Conference on North Gondwana Mid-Paleozoic biodynamics (IGCP 421), Vienna, 17–21 September, Abstract, 56–61.
Thorez, J., Goemaere, E., & Dreesen, R. (1988). Tide- and wave-influenced depositional environments in the Psammites du Condroz (Upper Famennian) in Belgium. In P. L. De Boer et al. (Eds.), Tide-influenced depositional environments and facies (pp. 389–415). Dordrecht: D. Reidel.
Thorez, J., Dreesen, R., & Streel, M. (2006). Famennian. Geologica Belgica, 9, 27–45.
Timokhina, I. G., Klets, A. G., & Bochkarev, V. S. (2002). Foraminifery famennskogo yarusa Zapadno–Sibirskoi plity (Famennian foraminifers from the Western Siberian plate). Novosti Paleontologii i Stratigrafii, 5, 123–135 (in Russian).
Tyszka, J. (1994). Response of Middle Jurassic benthic foraminiferal morphogroups to dysoxic/anoxic conditions in the Pieniny Klippen Basin, Polish Carpathians. Palaeogeography, Palaeoclimatology, Palaeoecology, 110, 55–81.
Vachard, D. (1974). Remarques sur les Foraminifères des calcaires griottes sensu lato (Frasnien inférieur-Tournaisien inférieur) du versant méridional de la Montagne Noire (Aude-Hérault). Comptes Rendus Sommaires de la Société géologique de France, 1973(4), 116–118.
Vachard, D. (1988). Calcareous microfossils (algae, pseudo-algae and foraminifera) from La Serre, Montagne Noire, France. Courier Forschungsinstitut Senckenberg, 100, 139–147.
Vachard, D. (1991). Parathuramminides et moravamminides (Microproblematica) de l’Emsien supérieur de la Formation Moniello (Cordillères Cantabriques, Espagne). Revue de Paléobiologie, 10(2), 255–299.
Vachard, D. (1994). Foraminifères et moravamminides du Givétien et du Frasnien du domaine Ligérien (Massif Armoricain, France). Palaeontographica Abteilung A, 231(1–3), 1–92.
Vachard, D. (2016). Macroevolution and biostratigraphy of the Paleozoic foraminifers. In M. Montenari (Ed.), Stratigraphy and timescales 1. Amsterdam: Elsevier. doi:10.1016/bs.sats.2016.10.005.
Vachard, D., & Clément, B. (1994). L’Hastarien (ex-Tournaisien inférieur et moyen) à algues et foraminifères de la zone Pélagonienne (Attique, Grèce). Revue de Micropaleontologie, 37(4), 238–319.
Vachard, D., & Cózar, P. (2003). Saccamminopsis, Mississippian carbonate microproblematicum, a possible udoteacean algal gametophyte (Chlorophyta, Siphonales). Geobios, 36, 421–436.
Vachard, D., & Cózar, P. (2004). Insolentitheca emend., Protoinsolentitheca n. gen. and Caligellidae emend., permanent cysts of Paleozoic foraminifera? Rivista Italiana di Paleontologia e Stratigrafia, 110(3), 591–603.
Vachard, D., & Cózar, P. (2010). An attempt of classification of the Palaeozoic incertae sedis Algospongia. Revista Española de Micropaleontología, 42, 129–241.
Vachard, D., Pille, L., & Gaillot, J. (2010). Palaeozoic Foraminifera: systematics, palaeoecology and responses to the global changes. Revue de Micropaleontologie, 53, 209–254.
Vachard, D., Haig, D. W., & Mory, A. J. (2014). Lower Carboniferous (middle Visean) foraminifers and algae from an interior sea, Southern Carnarvon Basin, Australia. Geobios, 47, 57–74.
Van Den Akker, T. J. H. A., Kaminski, M. A., Gradstein, F., & Wood, J. (2000). Campanian to Palaeocene biostratigraphy and palaeoenvironments in the Foula Sub-basin, west of the Shetland Islands, UK. Journal of Micropalaeontology, 19, 23–43.
Webb, G. E. (2001). Famennian mud-mounds in the proximal fore-reef slope, Canning Basin, Western Australia. Sedimentary Geology, 145, 295–315.
Wilson, J. L. (1975). Carbonate facies in geologic history. Berlin: Springer.
Wood, R. (2004). Palaeoecology of a post-extinction reef: Famennian (Late Devonian) of the Canning Basin, north-western Australia. Palaeontology, 47(2), 415–445.
Acknowledgements
We are grateful to Dieter Weyer for the identification of the solitary rugose corals from the Baelen Member and to J. Kalvoda and P. Cózar for their constructive reviews.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
This article is a contribution to the special issue “Climate change and biodiversity patterns in the mid-Palaeozoic”.
Rights and permissions
About this article

Cite this article
Vachard, D., Dreesen, R., Marion, JM. et al. New data on the incertae sedis biota and foraminifera of the mid-Famennian Baelen Member (Late Devonian, eastern Belgium). Palaeobio Palaeoenv 97, 565–584 (2017). https://doi.org/10.1007/s12549-016-0263-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-016-0263-y