
Abstract
The Sipuncula, or peanut worms, are a small taxon of lophotrochozoan worms that live mostly in warm shallow waters, but little is known about the diversity and distribution of this group in the deeper parts of the ocean (>2000 m). To address this point, all existing information from the scientific literature on deep-water sipunculans was retrieved, and a census was organised in several data matrices by species and by geographic and bathymetric distribution. From this data, deep-water sipunculan fauna were found to include representatives of 4 families, 10 genera, 51 species and 3 subspecies. Geographical range maps for all the censused species were created. The most ubiquitous species are Nephasoma diaphanes (Gerould, 1913); Phascolion lutense Selenka, 1885; Golfingia muricaudata (Southern, 1913); Apionsoma murinae (Cutler, 1969), G. margaritacea (Sars, 1851) and Onchnesoma magnibathum Cutler, 1969. By contrast, 18 species of various genera in the dataset are considered to be extremely rare, with only one previous record. No endemic species have been detected on the sea bottoms investigated to date. With regard to vertical distribution, most deep-water sipunculans were collected at depths ranging from 2500 to 3000 m, whereas there were fewer ultra-abyssal and hadal records (>6500 m). This also applies to the deepest depths investigated (>7000 m), where only three isolated records of three different species (N. diaphanes, P. lutense and P. pacificum Murina, 1957) were found. By relating species richness to depth, a significant linear regression was found, with lower richness values at greater depths. Finally, attempts to relate the species richness values to latitudinal or longitudinal gradients have as yet proved unsuccessful. The conclusion drawn from this census is that sipunculan fauna down to a depth of 2000 m is characterised as remarkably sensitive to bathymetry, with the lowest species richness values recorded in the deeper bottoms.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sipunculans (or peanut worms) are soft-bodied, defenceless, sedentary marine worms with a non-segmented body and a highly retractable introvert (Stephen and Edmonds 1972). Approximately 150 species of sipunculans have been described, mostly from warm oceans and shallow waters (Murina 1975a, 1984; Cutler 1994). Sipunculans have been reported in cryptic habitats such as burrows, crevices, tubes and discarded shells of other invertebrates in both sedimentary deposits and rocky bottoms. This enigmatic group has long been considered as a separate phylum, based mainly on the lack of body segmentation and chaetae. However, recent molecular analyses suggest that sipunculans should be placed close to or even nested within the phylum Annelida (Struck et al. 2007, 2011; Dordel et al. 2010; Hickman et al. 2014; Dunn et al. 2014; Ruggiero et al. 2015; Brusca et al. 2016).
The first sipunculans collected from the depths investigated (>2000 m) was by the Norwegian North-Atlantic Expedition 1876–1878 (Danielssen and Koren 1880, 1881), which found a new species named Nephasoma lilljeborgi (Danielssen and Koren, 1880) from depths of 2030 and 2127 m. A few years later, the results of the scientific voyage on board the HMS Challenger were published (Selenka 1885), with sipunculans being reported at abyssal depths, including the new species Nephasoma flagriferum (Selenka, 1885), collected at a depth of 2300 fathoms (4206 m) in the northwestern sector of the Pacific Ocean. More extensive collections of deep-water sipunculans were made during the second part of the twentieth century by well-known scientific expeditions (Murina 1975a, 1984; Cutler 1977; Cutler and Cutler 1980b, 1987). More recently, the Census of Marine Life (CoML), a global network of scientists, set out to assess and quantify the diversity of living organisms throughout the world’s oceans (McIntyre 2010; Snelgrove 2010; Alexander et al. 2011; Crist and O’Dor 2013). One of its field projects was the CeDAMar (Census of Diversity of Abyssal Marine Life), devoted to the study of marine diversity on abyssal plains (Stuart et al. 2008). This large research group was able to compile massive amounts of information on the global species diversity of several invertebrate groups and to investigate their dependence on environmental parameters (Ebbe et al. 2010). Some earlier results dealing with other invertebrate groups collected in waters more than 2000 m deep have been published (Brandt 2005; Paterson et al. 2009; Brandt et al. 2012; Gebruk et al. 2014).
As part of this global initiative, the present study set out (1) to update the global geographical distribution of all sipunculan species at depths in excess of 2000 m, and (2) to detect the (vertical and horizontal) spatial preferences of the species over all the world’s oceans.
Material and methods
The data used here represent a final compilation of all the scientific publications from 1880 to 2015 (Table 1). Only data for depths greater than 2000 m were selected, according to previous studies (Brandt 2005; Paterson et al. 2009; Ramirez-Llodra et al. 2010; Mironov et al. 2013; Gebruk et al. 2014). The compiled information is organized in a spreadsheet that includes details on species, authors, collection date, geographic position of the sampling site, depth, name of the expedition, research vessel name, type of gear used, specimen abundance of the haul, and updated species name of the collected specimens. Classification and coding of the higher taxa of sipunculans follows that of Cutler (1994), with the recent amendments by Kawauchi et al. (2012).
To analyse the diversity of deep–water sipunculans along spatial gradients, several data matrices are constructed with taxa organized in rows and distributional data in columns. A grid of 10 x 10-degree squares, as proposed by Ebbe et al. (2010), is used to estimate and plot the sipunculan species richness across the global geographical distribution. Subgenera and subspecies taxonomic details are omitted from the statistical analyses conducted here.
In order to check for any spatial gradient in the global distribution of deep-water sipunculans, the species richness values have been related to latitude, longitude and depth using linear regression analyses. Species richness was calculated by adding species presence/absence data collected at 30° intervals for both latitudinal and longitudinal analyses. In the case of depth, the species richness value was obtained by adding sipunculan species collected in arbitrary 500-m vertical bands from depths of 2000 to 7500 m. The relationship between the geographic distribution and the eurybathic/stenobathic characteristics of the species was also investigated using various correlation analyses. Regression and correlation analyses were conducted using the MINITAB® 16.1 software package (Minitab 2000). In addition, species were aggregated into higher taxonomic groups such as genera and plotted against depth to visualise different faunal strategies within Sipuncula, particularly in their colonising of the deep bottoms of the world’s oceans.
The geographical and bathymetric variability of the sipunculan assemblages were further investigated by multivariate analysis using the PRIMER 7 (Plymouth Routines in Multivariate Ecological Research) software package (Clarke and Gorley 2015). Two different approaches were adopted in order to (1) display relationships among samples (Clarke 1993) and to (2) address patterns among variables (Somerfield and Clarke 2013). All analyses are based on presence/absence data using Bray–Curtis similarity for analyses among samples and the Index of Association for analyses among spatial variables. The resemblance matrix thus obtained was then grouped using hierarchical agglomerative clustering and group-average linkage routines. The resulting groups were tested for statistical significance using similarities profile (SIMPROF) tests. To visualize possible patterns or trends among samples, non-metric multidimensional scaling ordination (MDS) plots are also shown. For the analysis of spatial variables, coherence plots in combination with shade plots were used to distinguish species groups which covaried coherently across samples.
Results
Geographical distribution
Table 1 lists the 51 species and 3 subspecies of sipunculans recorded in this study. The most ubiquitous species are Nephasoma diaphanes, Phascolion lutense, Golfingia muricaudata, Apionsoma murinae, G. margaritacea and Onchnesoma magnibathum. By contrast, 18 species found in the data collected are considered extremely rare, with only one record (Table 1).
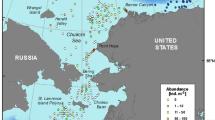
The geographical distribution of the data shows that most of the sipunculan records come from deep areas close to continental margins (Fig. 1). In fact, there are four hot-spot diversity areas that show high numbers of deep-water sipunculan species: the northeastern, northwestern and southeastern sectors of the Atlantic Ocean, and the northwestern part of the Pacific Ocean. However, the percentage of coloured grid cells (Fig. 1) indicating the existence of sipunculans is relatively low (34%) compared to the total number of grid cells greater than 2000 m. This is especially noteworthy in the central part of the Pacific Ocean, which remains largely undersampled.

Global map of sipunculan species recorded at depths in excess of 2000 m, shown as number of species per 10 × 10-degree square. Colours indicate different species number ranges
No significant relationships were found between species richness and their horizontal distribution along either the latitudinal or longitudinal lines of the geographical coordinates. Regarding global latitudinal distribution (Fig. 2a), the lowest species number value was recorded in latitudinal band 1, with only six species, corresponding mainly to the waters of the Arctic Ocean. Similarly, the Southern Ocean, predominantly in band 6 (from 60°S to 90°S), was relatively poor in deep-water sipunculan diversity, with only 12 species recorded. By contrast, a peak of diversity was recorded in latitudinal band 2, which include water masses from 30°N to 60°N, with 35 species counted. The remaining latitudinal bands (3, 4, 5) displayed intermediate diversity values ranging from 23 to 26 species.

Changes in the number of sipunculan species along latitudinal (a) and longitudinal bands (b) arbitrarily defined as 30° intervals. Correlation values are also included
With regard to longitudinal patterns, the diversity recorded in the longitudinal bands in the far western longitudes (bands 1, 2, 3, from 180°W to 90°W) was significantly lower than that in the rest of the areas sampled (Fig. 2b). By contrast, adjacent longitudinal bands from 90°W to 30°E had higher sipunculan species numbers, with a peak of 32 species recorded in longitudinal band 6 (from 30°W to 0°). In the remaining eastern longitudinal bands, species numbers ranged from 9 to 17.
Figure 3 shows the latitudinal variation in deep-water sipunculan species assemblages. SIMPROF analyses conducted on samples highlight the lack of a significant multivariate structure among the species, since all latitudinal bands (except band 1) were grouped in one cluster (Fig. 3a). Moreover, the ordination plot obtained by the MDS analysis (Fig. 3 b) failed to reflect any spatial gradient in faunal composition. Figure 3 c shows ten groups of differentiated species which covary coherently from one latitude to another. Two of these (groups A and C) were restricted to a single latitudinal band (4 and 5, respectively), while the rest were recorded in at least two latitudinal bands. Groups E, G and H are the best represented across all geographical latitudes. The sipunculan Nephasoma diaphanes was recorded in all latitudinal bands, while Phascolion lutense, Golfingia muricaudata, Nephasoma capilleforme (Murina, 1973), G. margaritacea and G. anderssoni (Théel, 1911) were recorded at all latitudes except latitudinal band 1.

Dendrogram (a) and ordination plot by non-metric multidimensional scaling (MDS) (b) obtained using Bray–Curtis similarity measures of species presence/absence data among latitudinal bands (30° intervals from 90°N to 90°S). (c) Shade plot of the data matrix of the 6 latitudinal bands and dendrogram based on the Index of Association among species showing ten ‘coherent groups’ (A–J). White space indicates absence of that species at that range. Solid lines indicate significant groups of samples or variables (p < 0.05); dashed lines indicate groups of samples or variables for which the null hypothesis cannot be rejected (p > 0.05)
The longitudinal variation in species composition is shown in Fig. 4. All samples were grouped in one cluster, as indicated by the SIMPROF analyses, denoting a quite similar multivariate structure of species from one geographical longitude to another (Fig. 4a). The ordination analysis obtained by MDS (Fig. 4b) has a stress value of 0.13, indicating a fairly good representation (Clarke and Warwick 2001). However, the graph does not show any longitudinal gradient in faunal composition. Nevertheless, a total of ten coherent groups of species are significantly obtained across global longitudes (Fig. 4c). Four of these (groups A, B, D and E) comprise species restricted to just one longitudinal band. Group F is the best represented across all longitudes, followed by groups G and H. The remaining groups are recorded in two (group C) or three (groups I and J) longitudinal bands. The sipunculan species Phascolion lutense was recorded in all longitudinal bands, but the rest exhibit noticeable gaps across global geographical longitudes.

Dendrogram (a) and ordination plot by non-metric multidimensional scaling (MDS) (b) obtained using Bray–Curtis similarity measures of species presence/absence data among longitudinal bands (30° intervals from 180°W to 180°E). (c) Shade plot of the data matrix of the 12 longitudinal bands and dendrogram based on the Index of Association among species showing 10 ‘coherent groups’ (A–J). White space indicates the absence of that species at that range. Solid lines indicate significant groups of samples or variables (p < 0.05); dashed lines indicate groups of samples or variables for which the null hypothesis cannot be rejected (p > 0.05)
Bathymetric distribution
A scatter plot of deep-water sipunculan species richness versus depth shows a clear pattern of decreasing diversity values, with the upper parts of the investigated depth spectrum having distinctly higher diversity (Fig. 5). A progressive drop in species richness was observed from 2000 m to 3500 m, followed by a relatively stable stretch to as deep as 5000 m, and a further decrease as the 7000-m isobath was approached. When relating the species number values to depth, a significant relationship was obtained that is well explained by a linear regression model, with highly significant results (p < 0.0001; r = 94.5).

Changes in the number of sipunculan species over 500-m depth bands. The solid line represents the fitted regression line (y = 47.4 – 0.00620 x; p < 0.0001; r = 94.5; n = 11)
Concerning bathymetric distribution, most sipunculan genera are well represented down to the 5500-m isobath, with the exception being Aspidosiphon Diesing, 1851; Themiste Gray, 1828; and Thysanocardia (Fisher, 1950), which showed a preference for upper depths (Fig. 6) throughout the depth spectrum investigated. Moreover, the cases of Themiste and Thysanocardia are rare, since they represent single isolated records in all of the area investigated.

Bathymetric preferences of the ten sipunculan genera recorded in this study. Note that the bathymetric range corresponding to the genus Phascolosoma includes a single record of the genus Themiste in the 4500–4999-m depth band, where Phascolosoma was lacking
With respect to the variation in species composition over the range of depths, the SIMPROF analysis confirms a significant multivariate structure with three distinct main depth zones (Fig 7a). The dendrogram recognized depths from 2000 to 2500 m as being in group I, from 3000 to 5000 m in group II, and greater depths in group III. The ordination plot obtained by the MDS analysis (Fig. 7b) clearly showed a sharp gradient in the sipunculan faunal composition from the upper to the lower depths investigated. Figure 7c indicated the existence of 12 significantly differentiated species groups which covaried coherently with depth. There are two groups of species that are distributed across all depth zones from the upper to the greater depths (groups E and G), and others that are restricted to the 2000–2500-m depth band (group A) or to the 3000–3500-m and 3500–4000-m bands (groups H and I, respectively). Other species were recorded across bathyal and abyssal zones (groups B, C, D and F). There are single groups with species recorded in a single depth range (groups J, K and H).

Dendrogram (a) and ordination by non-metric multidimensional scaling (MDS) (b) obtained using Bray–Curtis similarity measures of species presence/absence data in each depth band. (c) Shade plot of the data matrix of the 11 depth bands and dendrogram based on the Index of Association among species showing 12 ‘coherent groups’ (A–K). White space indicates absence of that species at that depth band. Solid lines indicate significant groups of samples or variables (p < 0.05); dashed lines indicate groups of samples or variables for which the null hypothesis can not be rejected (p > 0.05). I, II and III refer to the groups of samples resulting from the similarity profile (SIMPROF) tests
With regard to bathymetric distribution at the taxonomic specific level, Nephasoma diaphanes and Phascolion lutense were detected throughout the depth ranges investigated. Phascolion pacificum generally follows this pattern, although a clear gap is detected in its progressive bathymetric distribution, between the 6500-m and 7000-m isobaths. Moreover, Golfingia muricaudata, G. anderssoni, Nephasoma abyssorum, Onchnesoma magnibathum and G. vulgaris (De Blainville, 1827) were virtually ubiquitous in all depth zones investigated. A total of 22 sipunculan species were restricted to just one 500-m band, with ten of these inhabiting the upper part of the depth range investigated (2000–2500 m).
Linking geographic and bathymetric distributions
By cross referencing the distributional data of species along the horizontal and vertical axes, a significant positive relationship (r s = 0.887, p < 0.0001) was found between the number of grid cells and the number of depth bands in which each sipunculan species was recorded (Fig. 8). In general, species occurring in restricted geographic areas displayed a narrow vertical distribution, while species broadly distributed on the vast ocean floor also had greater vertical distribution. These contrasting faunal strategies were especially evident in stenobathic species, which were recorded in just a few isolated grid cells. On the contrary, more eurybathic species such as Nephasoma diaphanes, Phascolion lutense and Golfingia muricaudata were also the most widely distributed species worldwide. A few other species (Fig. 8), including Onchnesoma magnibathum, Apionsoma murinae, G. margaritacea and P. pacificum, showed a similar tendency, with larger distributions when both horizontal and vertical distribution axes were considered in a single correlation analysis. However, the species Golfingia anderssoni differed markedly, in that it was found in nine vertical bands but was fairly restricted along the horizontal axis, recorded in only six grid cells (Fig. 8).

Scatter plot showing the relationship between the horizontal and vertical distribution of deep-water sipunculans. Correlation values are also included
Discussion
The deep-water sipunculan fauna consisted of 51 species (plus 3 subspecies) arranged in 10 genera and 4 families (Table 1). No endemic species have been detected to date, based on a detailed comparison of the distribution of sipunculans investigated with previously published data (Cutler 1994). This may indicate the existence of frequent, repeated connections between shallow and deep-water sipunculan faunas and the absence of an effective oceanographic barrier separating the two groups. In terms of species richness, the species encountered make up almost 33 % of sipunculan diversity worldwide. This percentage is relatively low, since 70% of the diversity of the phylum is linked to shallow tropical areas throughout the world’s oceans (Murina 1971a, 1975a, 1977). However, the retrieved figures should be considered with caution. In recent years, several sipunculan species have been identified as complexes of cryptic species through the application of molecular methods (Staton and Rice 1999; Kawauchi and Giribet 2010, 2014; Schulze et al. 2012; Jonhson et al. 2015). The number of cryptic species in Sipuncula was estimated to represent between 10% and 55% of the total diversity (Kawauchi and Giribet 2010; Appeltans et al. 2012; Jonhson et al. 2015). No doubt, the extrapolation of these preliminary results to the overall study of the deep-water sipunculan fauna can represent a substantial increase in the species number obtained in our study.
At the familial level, almost all sipunculan families are represented on the sea bottoms investigated, with the exceptions being the recently erected families Antillesomatidae and Siphonosomatidae, established by Kawauchi et al. (2012). Antillesomatidae is monotypic, with a wide distribution in warm shallow waters (Cutler 1994). Siphonosomatidae shows a higher number of species in tropical and subtropical waters (Murina 1971a, 1975a, 1977; Cutler 1994). The remaining four sipunculan families are better represented in deep waters, with different radiation events. The family Golfingiidae Stephen and Edmonds, 1972, shows the greatest evolutionary diversification, not only in terms of the number of genera and subgenera (6 and 7, respectively), but also in its species numbers (36), compared with Phascolosomatidae Stephen and Edmonds, 1972 (2 genera, 4 subgenera, 8 species), Sipunculidae Rafinesque, 1814 (1 genus, 4 species) and Aspidosiphonidae De Quatrefages, 1865 (1 genus, 2 subgenera, 3 species). These contrasting proportions coincide with previous data from Murina (1975c, 1977, 1984), in which the family Golfingiidae is defined as eurybathic, while the other three families are mainly characterized as thermophilic/coralliophilic and are more dependent on the distribution of warm water masses.
Ten of the 16 genera of sipunculans are represented in the deeper bottoms of the ocean. Some of the genera not found, Antillesoma (Stephen and Edmonds, 1972), Cloeosiphon Grube, 1868, Phascolopsis (Fisher, 1950) and Siphonomecus Fisher, 1947, are monotypic. The first two of these are linked to warm, shallow waters, whereas the last two are very peculiar in their geographical distribution, since they are restricted to shallow waters in the western part of the Atlantic Ocean (Cutler 1994). The remaining two genera, Siphonosoma Spengel, 1912, and Xenosiphon Fisher, 1947, are linked to shallow, warm waters (Murina 1971a, 1975a; Cutler 1994). Of the ten genera found in the depth zones investigated, Themiste and Thysanocardia are rare in the dataset, with only a single record to date. Most records of Themiste retrieved from the scientific literature (Cutler 1994) correspond to the intertidal or upper part of the subtidal zones. Cutler questions the validity of the deep records of Themiste minor (Ikeda, 1904), since they are based on small specimens with anomalies in their anatomy or confusion in collecting station data. The single case of the genus Thysanocardia is to some extent parallel to the aforementioned genus Themiste, since the single record, published by Selenka (1885), represents a single specimen collected from a deep-water (3500 m)-deployed trawl located off the southern Brazilian coast, and no other specimens have ever been collected.
The most speciose genera found in the zone investigated were Nephasoma Pergament, 1946; Phascolion Théel, 1875; and Golfingia Lankester, 1885 (14, 11 and 7 species, respectively), which account for 62% of the total sipunculan census found. Additionally, four of the five subgenera of the genus Phascolion were also recorded in deep bottoms (Cutler 1994), a fact which suggests great potential for speciation in this genus. According to Cutler (1994), only Nephasoma is a predominantly deep-water genus, while Phascolion shows equal numbers of taxa living in shelf waters and deep waters. Phascolionids also have a tendency toward asymmetry in certain internal organs such as nephridia, retractor and gonad placement (Stephen and Edmonds 1972; Cutler 1994), apparently for a better adaptation to life in the helical shells of gastropods (Murina 1975c, 1977; Saiz et al. 2015). This adaptation may have originated in shallow waters, where large accumulations of empty shells are found on the seafloor. In the case of the genus Golfingia, a preference for residing at greater depths in lower latitudes is noted, a feature known as equatorial submergence (Cutler 1994). The evolutionary success of Nephasoma in deep waters may be explained by a rapid speciation in the cold bathyal and abyssal waters of the northern Atlantic and Pacific oceans (Cutler 1994). Thus, Murina (1975c, 1977) noted a process of secondary simplification of anatomical characters in deep-water sipunculans (retractor numbers, body musculature, tentacles, nephridia, etc.) as an adaptation to extreme hyperbaric and low thermal conditions. Finally, the remaining five genera—Phascolosoma Leuckart, 1828; Sipunculus Linnaeus, 1767; Aspidosiphon Diesing, 1851; Apionsoma Sluiter, 1902; and Onchnesoma Koren and Danielssen, 1876—are less diverse (3–5 species) in deeper waters. This is especially evident for Aspidosiphon and Phascolosoma, and to a lesser degree for Sipunculus, compared with the total species numbers for these genera in the world’s oceans (21, 17 and 10, respectively).
The most ubiquitous species are Nephasoma diaphanes, Phascolion lutense, Golfingia muricaudata, Apionsoma murinae, G. margaritacea and Onchnesoma magnibathum. In general, the species are characterised as common at bathyal and abyssal depths (Cutler 1994). It is interesting to note the collection of large and abundant (374 and 600) specimens of P. lutense from bottom trawls (Murina 1957b, 1974, 1977). This result corroborates observations by Rutgers and Lavaleye (1986) of high biomass (up to 28 % of the total macrofaunal biomass) of unidentified sipunculans on an abyssal plain in the northeast sector of the Atlantic Ocean. Moreover, P. lutense can be easily overlooked in dredge samples, since many specimens are enclosed in ‘mud balls’ or ‘cocoons’ produced by the binding of sediment particles with secreted mucus (Murina 1984; Gibbs 1985). Other common species such as Golfingia muricaudata have developed a characteristic long caudal appendage at the end of the body, apparently for anchoring the sipunculan body to the soft fluffy layer of sediment (Murina 1977). This distinctive adaptation is shared by other common deep-water species, namely G. anderssoni, N. bulbosum (Southern, 1913) and N. flagriferum, which are also common in the dataset. High density values (up to 375 individuals/m2) of this last species have been recorded on the abyssal plain from the Celebes Sea (Cutler 1977). N. diaphanes was also abundant (900 individuals) in a haul made on the abyssal plain in the southeastern part of the Atlantic Ocean (Saiz 2007). The species inhabits abandoned tubes of polychaetes for shelter.
By contrast, the data collected include 18 species from genera that are considered extremely rare, with only a single record to date. Some of these are mentioned above in the discussion of monotypic genera. Others are common species in shallow waters; consequently, their presence in deeper bottoms is noteworthy and even confusing in a few cases. This is especially evident for those species identified by Cutler (1977) from the Galathea expedition [S. robustus Keferstein, 1865; Aspidosiphon cristatus (Sluiter, 1902); Apionsoma pectinatum (Keferstein, 1867); P. collare Selenka and De Man, 1883; Phascolosoma nigrescens Keferstein, 1865; P. scolops (Selenka and De Man, 1883)]. This author was not confident in the taxonomic identifications provided. It remains to be demonstrated whether they represent cryptic or pseudo-cryptic complexes of other common species well distributed in shallow waters. The case of the deep-water species Phascolosoma turnerae Rice, 1985, is a good example of difficulties in the identification process (Saiz et al. 2014), due to its morphological similarity to other shallow-water species of Phascolosoma such as the common Phascolosoma granulatum Leuckart, 1828, in the Mediterranean Sea. P. turnerae is a wood-dwelling species patchily distributed in the bathyal Atlantic, Mediterranean and Australian waters (Edmonds 1985; Rice et al. 2012; Saiz et al. 2014).
Spatial analysis of sipunculan diversity (Fig. 1) shows that many areas of the world’s oceans are especially undersampled, such as the far western longitudes of both hemispheres, the Arctic Ocean, and a sector between 30°S and 60°S corresponding to the Atlantic, Pacific and Indian oceans. This is without doubt a source of serious bias in any biogeographical study, and seriously limits the scope of our study. Analysis of areas with deep-water sipunculans reveals no spatial patterns using latitudinal or longitudinal geographical coordinates as independent variables. This failure contrasts with the positive findings by Murina (1971a, 1975a, 1977) when working with shallow-water sipunculan fauna (0–200 m). Peaks of sipunculan diversity are recorded all around tropical areas, with the Indo-West Pacific region as the main center of origin.
Analyses of coherent groups are helpful in gaining a better understanding of the relationships among variables and samples. Interestingly enough, coherent groups of species vary when the species associations are studied across the horizontal and vertical distribution in the world’s oceans. Only two species remain relatively constant in the species assemblages obtained: Golfingia muricaudata and Nephasoma diaphanes. These species always are classified together in the same cluster when the horizontal and vertical distribution of the deep-water sipunculans is analysed. When analyses are restricted to global horizontal axes, further species associations emerge. Golfingia margaritacea, Nephasoma cutleri (Murina, 1975) and Phascolion lutense cluster together. Collectively, they are the most widely distributed in our dataset, except for N. cutleri. This observation is explored in detail by plotting a correlation analysis (Fig. 8) between the horizontal and vertical distribution of all deep-water sipunculan species. The result is statistically significant and reaffirms the earlier assertions by Murina (1971a, 1975a, 1977) concerning the existence of a close correspondence between the distribution abilities of the species over both dimensions in overall ocean distribution. In general, deep-water invertebrates showing a wide vertical range also have an extensive horizontal distribution (Vinogradova 1959, 1979, 1997).
Finally, the bathymetric range of deep-water sipunculans is generally quite large, so this broad depth tolerance was used to investigate the dependence of species on bathymetry. A total of 1348 records beyond the 2000-m contour were retrieved, and nearly half of them (49.3%) corresponded to the bathyal zone investigated here, while the rest were abyssal (50.03%). Records from the hadal zone are much rarer (0.67%)—indeed, there are only nine such records to date. The same is true for the deepest band investigated (>7000 m), where there are only three isolated records corresponding to three different species (N. diaphanes, P. lutense and P. pacificum). These total figures from deep waters can be considered very low compared to the amount of data from shallow warm waters, and they are in accordance with previous results (Murina 1971a, 1975a, 1977, 1984; Cutler 1994) in which bathyal, and especially abyssal, fauna of sipunculans are characterised as extremely poor. This faunal poverty has been explained as the result of a recent migration of some species to greater depths (Murina 1971a, 1975a, 1977).
The impact of depth on species richness is not consistent from one taxonomic group to another (Ellingsen et al. 2007; Brandt et al. 2009; Ebbe et al. 2010). By investigating the latitudinal/depth gradients in the Atlantic sector of the deep Southern Ocean, these authors found a lack of any consistent relationship between the numbers of isopod, polychaete and bivalve species with geographical coordinates of latitude or longitude. Depth, however, is a major environmental factor in organising species richness for isopods and polychaetes, but not for bivalves. Isopods show a unimodal response, but polychaetes are negatively affected by depth similarly to the sipunculans on a global scale (Murina 1984; this study). Much of the large-scale variation in diversity with depth appears to be related to food supply and its collateral effects on population density and growth (Rex and Etter 2010). According to these last authors, diversity at abyssal depths is probably depressed by vulnerability to extreme energy constraints, far below the levels needed for populations to be reproductively viable. Those limitations, linked to food availability, may explain the overall low rate of sipunculan diversification achieved in the deeper waters of the world’s oceans (Murina 1977).
In conclusion, deep-water sipunculan fauna is mainly characterised as remarkably dependent on bathymetry, with species richness greater in the upper part of the depth spectrum investigated and decreasing progressively towards the deepest sea bottoms.
References
Alexander V, Miloslavich P, Yarincik K (2011) The census of marine life—evolution of worldwide marine biodiversity research. Mar Biodivers 41:545–554
Appeltans W, Ahyong ST, Anderson G, Angel MV, Artois T, Bailly N, Bamber RN, Barber A, Bartsch I, Berta A, Blazewicz-Paszkowycz M, Bock P, Boxshall G, Boyko CB, Nunes Brandão S, Bray RA, Bruce NL, Cairns SD, Chan TY, Cheng L, Collins AG, Cribb TH, Curini-Galletti M, Dahdouh-Guebas F, Davie PJF, Dawson MN, De Clerck O, Decock W, De Grave S, de Voogd NJ, Domning DP, Emig CC, Erséus C, Eschmeyer W, Fauchald K, Fautin DG, Feist SW, Fransen CHJM, Furuya H, García-Alvarez O, Gerken S, Gibson D, Gittenberger A, Gofas S, Gómez-Daglio L, Gordon DP, Guiry MD, Hernandez F, Hoeksema BW, Hopcroft RR, Jaume D, Kirk P, Koedam N, Koenemann S, Kolb JB, Kristensen RM, Kroh A, Lambert G, Lazarus DB, Lemaitre R, Longshaw M, Lowry J, MacPherson E, Madin LP, Mah C, Mapstone G, McLaughlin PA, Mees J, Meland K, Messing CG, Mills CE, Molodtsova TN, Mooi R, Neuhaus B, Ng PKL, Nielsen C, Norenburg J, Opresko DM, Osawa M, Paulay G, Perrin W, Pilger JF, Poore GCB, Pugh PR, Read GB, Reimer JD, Rius M, Rocha RM, Saiz Salinas JI, Scarabino V, Schierwater B, Schmidt-Rhaesa A, Schnabel KE, Schotte M, Schuchert P, Schwabe E, Segers H, Self-Sullivan C, Shenkar N, Siegel V, Sterrer W, Stöhr S, Swalla B, Tasker ML, Thuesen EV, Timm T, Todaro MA, Turon X, Tyler S, Uetz P, van der Land J, Vanhoorne B, van Ofwegen LP, Van Soest RWM, Vanaverbeke J, Walker-Smith GK, Walter TC, Warren A, Williams GC, Wilson SP, Costello MJ (2012) The magnitude of global marine species diversity. Curr Biol 22:2189–2202
Brandt A (2005) Isopoda of the European North Atlantic from depths exceeding 2000 m, excluding Epicaridea. Mit Hamb Zool Mus Inst 102:179–190
Brandt A, Linse K, Schüller M (2009) Bathymetric distribution patterns of southern ocean macrofaunal taxa: Bivalvia, Gastropoda, Isopoda and Polychaeta. Deep-Sea Res Pt I 56:2013–2015
Brandt A, Blazewicz-Paszkowycz M, Bamber RN, Mühlenhardt-Siegel U, Malyutina MV, Kaiser S, de Broyer C, Havermans C (2012) Are there widespread peracarid species in the deep sea. Polish Polar Res 33:139–162
Brusca RC, Moore W, Schuster SM (2016) Invertebrates, 3rd edn. Sinauer Associates Inc Publishers, Sunderland
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143
Clarke KR, Gorley RN (2015) PRIMER v7: User Manual/Tutorial. PRIMER-E, Plymouth
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation, 2nd edn. PRIMER-E, Plymouth
Crist DT, O’Dor R (2013) 1: Census of Marine Life. In: Levin S (ed) Encyclopedia of Biodiversity, 2nd edn. Academic Press, Saint Louis MO USA, vol 2:1–22
Cutler EB (1967) The Sipuncula from the Western North Atlantic: their systematics, ecology and distribution. Dissertation, University of Rhode Island, (published by University Microfilms, Inc) no 69–1030, Ann Arbor, Michigan
Cutler EB (1969) New species of Sipuncula from the Western North Atlantic. Proc Biol Soc Wash 82:209–218
Cutler EB (1973) Sipuncula of the Western North Atlantic. Bull Am Mus Nat Hist 152(3):103–204
Cutler EB (1977) The bathyal and abyssal Sipuncula. Galathea Report 14:135–156
Cutler EB (1994) The Sipuncula their systematics, biology, and evolution. Cornell University Press, Ithaca
Cutler EB, Cutler NJ (1979) Madagascar and Indian Ocean Sipuncula. B mus natl hist nat 4ème sér A 1(4):941–990
Cutler EB, Cutler NJ (1980a) Sipuncula collected by the R/V "Vema". J Zool 190:193–209
Cutler EB, Cutler NJ (1980b) Deep-water Sipuncula from the Gulf of Gascogne. J Mar Biol Assoc UK 60:449–459
Cutler NJ, Cutler EB (1986) A revision of the genus Nephasoma (Sipuncula: Golfingiidae). Proc Biol Soc Wash 99(4):547–574
Cutler EB, Cutler NJ (1987) Deep-water Sipuncula from the eastern Atlantic Ocean. Sarsia 72:71–89
Cutler EB, Cutler NJ (1996) Sipuncula from the Indian ocean and New Caledonia. B mus natl hist nat 4ème sér A 18(3–4):341–365
Cutler EB, Duffy NA (1972) A new species of Phascolion (Sipuncula) from the western North Atlantic. Proc Biol Soc Wash 85:71–76
Cutler EB, Cutler NJ, Nishikawa T (1984) The Sipuncula of Japan: their systematics and distribution. Publ Seto Mar Biol Lab 29(4/6):249–322
Cutler EB, Dean HK, Saiz Salinas JI (2001) Sipuncula from Antarctic waters. Proc Biol Soc Wash 114:861–880
Cutler EB, Schulze A, Dean HK (2004) The Sipuncula of sublittoral New Zealand, with a Key to all New Zealand species. Zootaxa 525:1–19
Danielssen DC (1869) Sipunculus norvegicus Danielssen. Forh Skand Naturf Ott Mod 10:541–542
Danielssen DC, Koren JG (1880) New northern Gephyrea. Ann Mag Nat Hist ser 5(6):462–465
Danielssen DC, Koren JG (1881) Zoology. Gephyrea. In: The Norwegian North-Atlantic Expedition. Grondahl and Son, Christiania, pp 1876–1878
De Blainville HMD (1827) Siponcle. In: Dictionnaire des sciences naturelles, dans lequel on traite méthodiquement des diffèrents êtres de la nature, considerés soit en eux-memes, d’après l’état actuel de nos connoissances, soit relativement à l’utilité qu’en peuvent retirer la médicine, l’agriculture, le commerce et les arts (vol 49), Levrault, Paris, pp 305–313
De Quatrefages A (1865) Histoire Naturelle des Annelés marins et d’eau douce. Annélides et Géphyriens (3 vols), Roret, Paris
Diesing CM (1851) Systema Helminthum (vol 1). Braumüller, Vindobonae
Dordel J, Fisse F, Purschke G, Struck TH (2010) Phylogenetic position of Sipuncula derived from multi-gene and phylogenomic data. J Zool Syst Evol Res 48:197–207
Dunn CW, Giribet G, Edgecombe GD, Hejnol A (2014) Animal phylogeny and its evolutionary implications. Annu Rev Ecol Evol Syst 45:371–95
Ebbe B, Billett DSM, Brandt A, Ellingsen K, Glover A, Keller S, Malyutina M, Martínez-Arbizu P, Molodtsova T, Rex M, Smith C, Tselepides A (2010) Chapter 8: Diversity of Abyssal Marine Life. In: McIntyre AD (ed) Life in the World’s Oceans, Blackwell Publishing Ltd Oxford, pp 139–160
Edmonds SJ (1985) A new species of Phascolosoma (Sipuncula) from Australia. T Roy Soc South Aust 109:43–44
Edmonds SJ (1991) Sipunculoidea and Echiuroidea: Sipunculans and Echiurans from the Philippines and New Caledonia (ESTASE 2, BIOCAL, MUSORSTOM 3 and 4). Memoir Mus natl Hist (A) 151:83–90
Ellingsen K, Brandt A, Hilbig B, Linse K (2007) The diversity and spatial distribution of polychaetes, isopodes and bivalves in the Atlantic sector of the deep Southern Ocean. Polar Biol 30:1265–1273
Fischer W (1922) Gephyreen der Deutschen Tiefsee-Expedition. In: Chun C (ed) Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition mit dem Dampfer ‘Valdivia’ 1898–1899 22:1–26, 1–2 pls
Fisher WK (1947) New genera and species of echiuroid and sipunculoid worms. Proc Biol Soc Wash 97:351–372, pls 8–15
Fisher WK (1950) The sipunculid genus Phascolosoma. Ann Mag Nat Hist ser 12(3):547–552
Gebruk AV, Thiel H, Thurston M (eds) (2014) Deep-sea Fauna of European Seas: An annotated species check-list of benthic invertebrates living deeper than 2000 m in the seas bordering Europe. Introduction. Invertebr Zool 11:1–2
Gerould JH (1913) The Sipunculids of the eastern Coast of North America. Proc US Nat Mus 44:373–437, pls 58–62
Gibbs PE (1985) On the genus Phascolion (Sipuncula) with particular reference to the north-east Atlantic species. J Mar Biol Assoc UK 65:311–323
Gibbs PE (1986) The taxonomy of some little-known Sipuncula from the north-east Atlantic Region including new records. J Mar Biol Assoc UK 66:335–341
Gray JE (1828–30) Spicilegia Zoologica or original Figures and short systematic Descriptions of new and unfigured Animals. Treuttel Würtz and Co
Grube E (1868) Loxosiphon, Cloeosiphon, und einige Phascolosomen. Jahres-Bericht der Schlesischen Gesellschaft für vaterländische Cultur 45:47–49
Haldar BP (1975) Sipuncula of the Indian Ocean in the Collection of the Zoological Survey of India. In: Rice ME, Todorovic M (eds) Proceedings of the International Symposium on the Biology of Sipuncula and Echiura. Naucno Delo, Beograd, vol 1:51–92
Hickman C, Roberts LS, Keen SL, Eisenhour DJ, Larson A, l’Anson H (2014) Integrated Principles of Zoology, 16th edn. McGraw-Hill Education, New York
Ikeda I (1904) The Gephyrea of Japan. J Coll Agric Imp Univ Tokyo 20:1–87, 4 pls
Jonhson ND, Sanders C, Maiorova A, Schulze A (2015) Cryptic species in Pacific sipunculans (Sipuncula: Phascolosomatidae): east–west divergence between non-sister taxa. Zool Scr 45:455–463
Kawauchi GY, Giribet G (2010) Are there true cosmopolitan sipunculan worms? A genetic variation study within Phascolosoma perlucens (Sipuncula, Phascolosomatidae). Mar Biol 157:1417–1431
Kawauchi GY, Giribet G (2014) Sipunculus nudus Linnaeus, 1766 (Sipuncula): cosmopolitan or a group of pseudo-cryptic species? An integrated molecular and morphological approach. Mar Ecol 35:478–491
Kawauchi GY, Sharma PP, Giribet G (2012) Sipunculan phylogeny based on six genes, with a new classification and the descriptions of two new families. Zool Scr 41:186–210
Kedra M, Murina GV (2007) The sipunculan fauna of Svalbard. Polar Res 26:37–47
Kedra M, Shields MA (2011) On the deep-sea Nephasoma species in the Eastern North Atlantic a taxonomic guide. Mar Biol Res 7:43–53
Keferstein W (1862) Über die Gattung Phascolosoma. Nach Uni Gesell Wissen Göttingen 1862:65–67
Keferstein W (1865) Beiträge zur anatomischen und systematischen Kenntniss der Sipunculiden. Nach Uni Gesell Wissen Göttingen 1865:189–209
Keferstein W (1866) Untersuchungen über einige amerikanische Sipunculiden. Nach Uni Gesell Wissen Göttingen 1866(14):215–228
Keferstein W (1867) Untersuchungen über einige amerikanische Sipunculiden. Z Wiss Zool 17:44–55, pl 6
Koren J, Danielssen DC (1876) Bidrag til de norske Gephyreers Naturhistorie. Nyt Mag Naturvidensk 21:108–138
Lankester ER (1885) Golfingia MacIntoshii, a new Sipunculid from the Coast of Scotland. Trans Linn Soc Lond 11:469–474, pls 55–56
Leuckart F-S (1828) Breves animalium quorundam maxima ex parte marinorum descriptiones. A Osswaldii, Heidelberg
Linnaeus C (1766/67) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio duodecima, reformata (vol 1: Regnum animale. Pars I et II). Laurantii Salnii, Impensis Direct Holmiae (Stockholm)
Maiorova AS, Adrianov AV (2015) Deep-sea sipunculans from the Kuril-Kamchatka trench and adjacent abyssal plain. Deep-Sea Res Pt II 111:128–134
McIntyre AD (ed) (2010) Life in the World’s oceans; diversity, distribution, and abundance. Blackwell Publishing Ltd, Oxford
Minitab® Statistical Software v16 (2000) Computer software. Minitab, Inc
Mironov AN, Dilman AB, Krylova EM (2013) Global distribution patterns of genera occurring in the Arctic Ocean deeper 2000 m. Invertebr Zool 10(1):167–194
Montagu G (1804) Description of several marine animals found on the south coast of Devonshire. Trans Linn Soc Lond 7:61–85, pls 6–7
Murina VV (1957a) Abyssal sipunculids (genus Phascolion Theel) of the north-western part of the Pacific collected by ‘Vitjaz’ expeditions in 1950–1955. Zool Zh 36:1777–1791
Murina VV (1957b) Sipunculids collected on the first trip of the complex Antarctic expedition on the ‘Ob’ in 1956. Zool Zh 36:992–998
Murina VV (1958) K sistematike dvukh blizkikh vidov glubokovodnykh sipunkulid roda Golfingia po materialam ékspeditsii na "Vityaze" v 1949–1955 gg. Zool Zh 37(11):1624–1633
Murina VV (1961) On the geographic distribution of the abyssal sipunculid Phascolion lutense. Okeanologiya 1:140–142
Murina VV (1964) New and rare species of sipunculids of genus Golfingia. Trudy Instituta Okeanologii 69:216–253
Murina VV (1968) New data concerning the occurence of abyssal sipunculids in the Atlantic Ocean. Zool Zh 47:95–199
Murina VV (1969) Glubokovodnye sipunkulidy i priapulidy Tikhogo okeana. In: Zenkevitch, LA (ed) Tikhii Okean. Biologiya Tikhogo Okeana. Kniga 2. Glubokovodnaya Donnaya Fauna. Pleuston. Nauka, Moscow, pp 84–89
Murina VV (1971a) Geographical distribution of marine worms of the phylum Sipuncula in the world ocean. Zool Zh 50:184–192
Murina VV (1971b) On the occurence of deep-sea sipunculids and priapulids in the Kurile-Kamchatka trench. Trudy Instituta Okeanologii 92:41–45
Murina VV (1972) K faune sipunkulid Yuzhnogo polushchariya. Issledovaniya fauny morei 19:294–314
Murina VV (1973a) The fauna of sipunculids from the Peruvian-Chilean Trench. Vestn Zool 5:66–71
Murina VV (1973b) New species of Pacific sipunculids. Zool Zh 52:942–945
Murina VV (1974) Contributions to the knowledge of the fauna of sipunculid worms from the south Atlantic based on data of the ‘Akademik Kurchatov’ expedition in 1971. Trudy Instituta Okeanologii 98:228–239
Murina VV (1975a) The geographical distribution of marine worms of the phylum Sipuncula of the World Ocean. In: Rice ME, Todorovic M (eds) Proceedings of the International Symposium on the Biology of Sipuncula and Echiura. Naucno Delo, Beograd, vol 1:3–18
Murina VV (1975a) Novye taksony morskikh chervei sipunkulid roda Golfingia. Zool Zh 54(7):1085–1089
Murina VV (1975b) Puti évolyutsii i filogeniya tipa Sipuncula. Zool Zh 54(12):1747–1758
Murina G-VV (1977) Morskie chervi sipunkulidy arkticheskikh i boreal’nykh vod Evrazii. In: Oprediteli po faune SSSR, izdavaemye Zoologicheskim Institutom Akademii Nauk SSSR (vol 111). "Nauka", Leningrad
Murina G-VV (1978) On the sipunculids and priapulids fauna of the southern Pacific. Trudy Instituta Okeanologii 113:120–131
Murina G-VV (1981) New species of the genus Phascolion (Sipuncula) from the shells of Scaphopoda. Zool Zh 60:348–352
Murina G-VV (1982) On the sipunculans and echiurans fauna of Mediterranean and Iberian basin. Trudy Instituta Okeanologii 117:178–191
Murina G-VV (1984) Ecology of Sipuncula. Mar Ecol Prog Ser 17:1–7
Murina G-VV (1993) New data of the fauna of Sipuncula, Echiura and Priapulida from Atlantic ocean. Trudy Instituta Okeanologii 127:107–121
Murina VV (2008) New records of Sipuncula and Echiura in the north Atlantic Ocean and description of a new species of Jacobia. Mar Biol Res 4:1–5
Paterson GLJ, Glover AG, Barrio Froján CRS, Whitaker A, Budaeva N, Chimonides J, Doner S (2009) A census of abyssal polychaetes. Deep-Sea Res Pt II 56:1739–1746
Pergament T (1946) O novom rode i maloizvestnom vide Sipunculida iz Severnogo ledobitogo okeana. In: Buynitskiy VKh (ed) Trudy dreifuiushcheyi ekspeditsii Glavsevmorputi na ledokol’nom parokhode “G. Sedov”, 1937–40 gg. Izdatel’stvo Glavsevmorputi, Moskva 3:189–193
Rafinesque CS (1814) Précis des découvertes somiologiques ou zoologiques et botaniques. Royale Typographie Militaire, Palerme
Ramirez-Llodra E, Brandt A, Danovaro R, Escobar E, German CR, Levin LA, Martinez Arbizu P, Menot L, Buhl-Mortensen P, Narayanaswamy BE, Smith CR, Tittensor DP, Tyler PA, Vanreusel A, Vecchione M (2010) Deep, diverse and definitely different: unique attributes of the world’s largest ecosystem. Biogeosciences 7:2851–2899
Rex MA, Etter RJ (2010) Deep-Sea Biodiversity. Pattern and Scale. Harvard University Press, Cambridge, Massachusetts
Rice ME (1985) Description of a wood dwelling Sipunculan, Phascolosoma turnerae, new species. Proc Biol Soc Wash 98:54–60
Rice ME, Reichardt HG, Piraino J, Young CM (2012) Reproduction, development, growth, and the length of larval life of Phascolosoma turnerae, a wood-dwelling deep-sea sipunculan. Invertebr Biol 131:204–215
Roule L (1898) Notice préliminaire sur les espèces de Géphyriens recueillis dans les explorations sous-marines du Travailleur et du Talisman. B mus hist nat 4:384–387
Roule L (1906) Annélides et Géphyriens. In: Expéditions scientifiques du ‘Travailleur’ et du ‘Talisman’ pendant les années 1880, 1881, 1882, 1883. Masson et Cie, Paris, 8:1–102, pls 1–10
Ruggiero MA, Gordon DP, Orrell TM, Bailly N, Bourgoin T, Brusca RC, Cavalier-Smith T, Guiry MD, Kirk PM (2015) A higher level classification of All living organisms. PLoS ONE 10(4), e0119248
Rutgers van der Loeff MM, Lavaleye MSS (1986) Sediments, Fauna and the Disposal of Radionuclides at the NE Atlantic Dumpsite for Low-level Radioactive Waste. Netherlands Institute for Sea Research, Texel
Saiz Salinas JI (1988) Sipunculiden (Sipuncula) des östlichen Nordatlantik, gesammelt während der Fahrten des FS "Meteor". Mit Hamb Zool Mus Inst 85:7–23
Saiz Salinas JI (1995) Sipuncula of the southeastern Weddell Sea (Antarctica). Polar Biol 15:307–317
Saiz Salinas JI (2007) Sipunculans and echiurans from the deep Angola basin. J Nat Hist 41:2789–2800
Saiz Salinas JI (2016) Further records of sipunculans (Sipuncula) from the Mediterranean Sea. Mar Biodivers. doi:10.1007/s12526-016-0443-z
Saiz JI, Villafranca L (1990) Sipuncula from the Alboran Sea and ibero-moroccan Bay. J Nat Hist 24:1143–1177
Saiz JI, Cartes J, Mamouridis V, Mechó A, Pancucci-Papadopoulou MA (2014) New records of Phascolosoma turnerae (Sipuncula: Phascolosomatidae) from the Balearic Basin, Mediterranean Sea. Mar Biodivers Rec 7, e16
Saiz JI, Bustamante M, Tajadura J, Vijapure T, Sukumaran S (2015) A new subspecies of Phascolion Théel, 1875 (Sipuncula: Golfingiidae) from Indian waters. Zootaxa 3931(3):433–437
Sars M (1851) Beretning om en i Sommeren 1849 foretagen zoologisk Reise i Lofoten og Finmarken. Nyt Mag Naturvidensk 6(2):121–211
Schulze A, Maiorova A, Timm LE, Rice ME (2012) Sipunculan larvae and "cosmopolitan" species. Integr Comp Biol 52:497–510
Selenka E (1885) Report on the Gephyrea collected by H.M.S. Challenger during the years 1873–76 In: Report on the scientific results of the voyage of H.M.S. Challenger, during the years 1873–76. Adam and Charles Black, Edinburgh 13(36):25 pp, 4 pls
Selenka E, De Man JG, Bülow C (1883–84) Die Sipunculiden. Reisen im Archipel der Philippinen von Dr. C. Semper. Zweiter Theil. Wissenschaftliche Resultate (4. Band, 1. Abtheilung). W. Engelmann, Leipzig
Shields MA, Kedra M (2009) A deep burrowing sipunculan of ecological and geochemical importance. Deep-Sea Res Pt I 56:2057–2064
Sluiter CPh (1900) Géphyriens (Sipunculides et Echiurides) provenant des Campagnes de "l’Hirondelle" et de la "Princesse Alice" (1886–1897) In; Résultats des Campagnes scientifiques accomplies sur son yatch, par Albert 1er, publiées sous sa direction avec le concours de M. Jules Richard. Imprimerie de Monaco, Monaco 15:1–30, pls 1–3
Sluiter CPh (1902) Die Sipunculiden und Echiuriden der Siboga-Expedition nebst Zusammenstellung der Ueberdies aus dem Indische Archipel bekannten Arten. In: Weber M (ed) Siboga-Expeditie: uitkomsten op zoologisk, botanisch, oceanographisch en geologisch gebied verzameld in Nederlandsch Oost-Indië 1899–1900 aan boord H.M. Siboga onder commando van luitenant ter zee le kl. G.F. Tydeman, Brill EJ, Leiden 25:1–53, 1–4 pls
Sluiter CPh (1912) Géphyriens (Sipunculides et Echiurides) provenant des campagnes de la Princesse Alice (1898–1910). In: Résultats des Campagnes scientifiques accomplies sur son yatch, par Albert 1er, publiées sous sa direction avec le concours de M. Jules Richard. Imprimerie de Monaco, Monaco 36:1–36, 1 pl
Snelgrove PVR (2010) Discoveries of the Census of Marine Life. Making Ocean Life Count. Cambridge University Press, Cambridge, New York
Somerfield PJ, Clarke KR (2013) Inverse analysis in non-parametric multivariate analyses: distinguishing groups of associated species which covary coherently across samples. J Exp Mar Biol Ecol 449:261–273
Southern R (1913) Gephyrea of the coasts of Ireland. Scientific Investigations. Department of agriculture and technical instruction for Ireland. Fisheries branch 1912 (3):1–46, pls 1–7
Spengel JW (1912) Einige Organisationsverhältnisse von Sipunculusarten und ihre Bedeutung für die Systematik dieser Tiere. Verh Dtsch Zool Ges 22:261–272
Staton JL, Rice ME (1999) Genetic differentiation despite teleplanic larval dispersal: allozyme variation in sipunculans of the Apionsoma misakianum species-complex. B Mar Sc 65:467–480
Stephen AC (1948) Sipunculids. In: Johnston TH (ed). British, Australian and New Zealand (B.A.N.Z.) Antarctic Research Expedition 1929–1931 under the commando of Douglas Mawson. Reports – Series B (Zoology and Botany) (vol 5, part 4). The Hassell Press, Adelaide pp 213–220, pl 1
Stephen AC, Edmonds SJ (1972) The phyla Sipuncula and Echiura. Trustees of the British Museum (Natural History), London
Struck TH, Schult N, Kusen T, Hickman E, Bleidorn C, McHugh D, Halanych KM (2007) Annelid phylogeny and the status of Sipuncula and Echiura. BMC Evol Biol 7:57
Struck TH, Paul C, Hill N, Hartmann S, Hösel C, Kube M, Lieb B, Meyer A, Tiedemann R, Purschke G, Bleidorn C (2011) Phylogenomic analyses unravel annelid evolution. Nature 471:95–98
Stuart CT, Martínez Arbizu P, Smith CR, Molodtsova T, Brandt A, Escobar-Briones E, Fabri MC, Rex MA (2008) CeDAMar global database of abyssal biological sampling. Aquat Biol 4:143–145
Théel H (1875) Etude sur les Géphyriens inermes des mers de la Skandinavie, du Spitzberg et du Groënland. Bih K Sven vetensk akad handl 3 (6):3–30, pls 1–4
Théel H (1911) Priapulids and sipunculids dredged by the Swedish Antarctic Expedition 1901–1903 and the Phenomenon of Bipolarity. K Sven vetensk akad handl 47 (1):3–36, pls 1–5
Thompson BE (1980) A new bathyal Sipunculan from southern California, with ecological notes. Deep-Sea Res 27A:951–957
Vinogradova NG (1959) The zoogeographical distribution of the deep-water bottom fauna in the abyssal zone of the ocean. Deep-Sea Res 5:203–208
Vinogradova NG (1979) The geographical distribution of the abyssal and hadal (ultra-abyssal) fauna in relation to the vertical zonation of the ocean. Sarsia 64:41–50
Vinogradova NG (1997) Zoogeography of the Abyssal and Hadal zones. Adv Mar Biol 32:325–387
Wesenberg-Lund E (1930) Priapulidae and Sipunculidae. In: The Danish Ingolf-Expedition. H. Hagerup, Copenhaguen 4B(7):1–44, 1–6 pls
Wesenberg-Lund E (1955) Sipunculidae In: Reports of the Swedish Deep-Sea Expedition vol 2, Zoology (no 15):197–201
Acknowledgments
This research was supported by the Census of the Diversity of Abyssal Marine Life (CeDAMar). We are indebted to Prof. Dr. Pedro Martínez (Senckenberg am Meer Wilhelmshaven, Germany) and the CeDAMar group for their assistance during the preparation of this work. At the time of this writing, M.B. was the recipient of a postdoctoral research grant (Especialización Personal Investigador) from the University of the Basque Country (UPV/EHU).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by D. Fiege
Rights and permissions
About this article

Cite this article
Saiz, J.I., Bustamante, M. & Tajadura, J. A census of deep-water sipunculans (Sipuncula). Mar Biodiv 48, 449–464 (2018). https://doi.org/10.1007/s12526-016-0568-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12526-016-0568-0