
Abstract
This study aims to characterize the dietary habits of the Barcelona plain (Spain) population during the Early Bronze Age. An analysis of buccal microwear has been performed on 467 samples from 20 individuals buried in the largest cemetery of the plain of Barcelona in this period, the Plaça de la Gardunya, and 1 sample from the site of the Mercat de Santa Caterina. Molars and premolars were examined using a scanning electron microscope (SEM), and their microwear patterns were compared with reference patterns associated with known dietary habits and subsistence strategies. The results suggest that dietary habits differed between age groups and between sexes: males were more carnivorous than females, and subadults ate harder foods than adults. Buccal microwear from the Barcelona plain shows similar values to carnivorous diets of anthropological populations. Based on all the archaeological evidence (building types and zooarchaeology, archaeobotany and stone tools) from this settlement and those in its vicinity, we propose that the individuals from the Barcelona plain had a mixed diet, with large consumption of meat, secondary products and fish. Agricultural products, stored in silos and grill structures and processed with more sophisticated grinding techniques, were also consumed, mainly by subadult individuals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Early Bronze Age in the northeastern Iberian Peninsula is a widely studied period, though it is still poorly characterized because of the paucity of archaeological remains. As a result, most studies have focused on individual sites or specific technologies, such as pottery (Gómez et al. 2015; Maya and Petit 1986, Daura et al. 2017; Soriano 2013), metallurgy (Rovira 2006; Soriano 2018) and ornaments (Soriano 2013; Balaguer et al. 2011; Soriano and Chamón 2012; Daura et al. 2017). During the Early Bronze Age, the population lived in two different types of settlements: in caves at high altitudes and in open-air settlements on the plains. At high altitudes, dwellings were made of durable materials, such as stone, which suggests a high degree of sedentarisation. The occupation and use of caves were common, as shown by the existence of settlements in the caves of Cova de les Pixarelles, Cova del Toll (Maya 1997), Cova del Frare (Martín et al. 1981) or the Bauma del Serrat del Pont (Alcalde et al. 1998). Not all of them were used as dwellings, but also as seasonal livestock shelters (Soriano 2013). Evidence also shows an increase in the number of open-air settlements built in plain areas close to rivers, compared to the Chalcolithic period. These sites used perishable elements such as mud or plants as building materials.
In the Barcelona plain, a total of nine archaeological sites from the Early Bronze Age have been identified: Solar del Rec Comtal 17-19, Solar del Pou de la Figuera, Mercat de Santa Caterina, Plaça de la Gardunya, Illa d’en Robador, Filmoteca de Catalunya, Carrer de la Riereta 37-37 bis, Caserna de Sant Pau del Camp and Conservatori del Liceu. All of them are open-air settlements where the dwellings are usually circular or irregular in shape and are built with perishable materials. The Caserna de Sant Pau del Camp site is an exception to this pattern: a quadrangular building was constructed with stone materials (Granados et al. 1993). The presence of several hearths suggests that social activities took place outside the huts. The oldest radiocarbon samples found in the Plaça de la Gardunya site indicate that this area was first used as a cemetery in the Chalcolithic period (around 2200 cal BC). The oldest dates of occupation can be found in the site of Illa d’en Robador, where a charcoal fragment has been dated to 14C (ca. 2000 cal. BCE) (Gómez et al. 2015; Gómez and Molist 2016a, b; Velasco et al. 2017).
The diet of the Early Bronze Age populations of the Barcelona plain has been reconstructed using archaeobotanical and zooarchaeological analyses, as well as identifying macrolithic artefacts. Research has shown that the subsistence economy was based on a mixed diet, in which agriculture played a very important role. The rearing of livestock to produce primary and secondary products was combined with hunting and fishing. The collection of wild plants was less important. Carpological studies conducted at the Caserna de Sant Pau del Camp have shown that various plant species were processed, including barley (Hordeum vulgare and Hordeum vulgare nudum), soft wheat (Triticum aestivum), durum wheat (Triticum durum), club wheat (Triticum compactum), emmer wheat (Triticum dicoccum) and some legumes (Vicia faba) (Gómez and Molist 2016a, b). The presence of macrolithic artefacts such as grindstones, axes and hoes in the settlements of Illa d’en Robador, the Filmoteca de Catalunya, the Plaça de la Gardunya, the Carrer de la Riereta 37-37 bis and the Caserna de Sant Pau del Camp sites is evidence of agricultural and food processing activities. Carrer de la Riereta 37-37 bis is the most characteristic site, where 16 grindstones have been found. The materials used to make these tools were clastic rocks with large grain sizes, useful for grinding cereals (Bordas 2016; Piera et al. 2005; Suau and Matas 2002; Nadal and Castillo 2009; Velasco et al. 2017; Granados et al. 1993; Gómez and Molist 2016a, b; Bofill 2016). Furthermore, the volumetric capacity and the number of silos present in this period indicate a great increase in agricultural production compared to previous periods. In addition to silos, grill-plan structures have also been found, the first to be documented in the north-east of the Iberian Peninsula. These characteristic structures have a rectangular plan and are made up of several parallel stone plinths. Some of these structures, although not completely preserved in the Barcelona plain, usually have stone slabs or a surface made of perishable materials over the plinths. The main hypothesis is that these grill-plan structures were elevated granaries with a dual function: to keep agricultural products like grain cool and dry and to protect them from moisture so they would not spoil. Thus, the grill plan structures would complement the storage function of silos (Gómez and Molist 2016a, b; Carlús et al. 2010). These engineering structures were first documented in Anatolia during the Neolithic period (ca. 8000–6500 BCE), and it makes sense to find them in the plain of Barcelona since it is a very humid area, and it is very close to water resources, such as the rivers Besòs and Llobregat and several streams coming from the local mountains of Collserola and Montjuïc, as well as the presence of the Mediterranean Sea.
The widespread practice of animal husbandry is evidenced by the analysis of the faunal remains. Ovicaprids (Capra hircus and Ovis aries) were particularly favoured, providing milk and wool. Meat consumption was mainly from suids (Sus domesticus) and bovids (Bos taurus), which provided multiple resources such as meat, brain, offal and marrow. Bovines were also used for traction power. However, there is little evidence of hunting in the Barcelona plain: three bone remains of wild rabbits (Oryctolagus cunniculus) were found at the sites of Sant Pau del Camp and Plaça de la Gardunya, with no specific evidence of human processing (Gómez et al. 2015; Alcàntara 2018). These are specific and scarce remains for which there is insufficient evidence either to suggest their consumption or to consider them intrusive (Alcàntara 2018). The harvesting of mollusks (Patella sp. and Glycymeris glycymeris) was frequent, but there is only evidence of their use as raw materials for making ornaments (Gómez et al. 2015). Evidence of fishing is also scarce: no ichthyofaunal remains have yet been found (Gómez et al. 2014), and only the weight of a fishing net with a triangular morphology has been found in Carrer de la Riereta 37-37 bis (Bofill 2016).
From an anthropological perspective, funerary remains have been found in 6 of the 9 sites. Due to the poor state of dental preservation of most of the archaeological sites considered, only two of them could be included in the present study: Plaça de la Gardunya and Mercat de Santa Caterina. Both are located in the plain of Barcelona between the rivers Besòs and Llobregat. The Plaça de la Gardunya (GCS 2° 10′ 14.8354″ E, 41° 22′ 52.96026″ N; ETRS89 UTM zone 31N, X 430665.185328, Y 4581426.71338) covers an area of at least 600 m2. However, the total area occupied remains unclear, as excavations have only been carried out in a specific area—a car park. Due to the presence of adjacent buildings, the likelihood of an open-area excavation at this site is very low (Velasco et al. 2017). The fieldwork uncovered several types of evidence related to diet: one negative stratum filled with faunal remains, 7 silos, two combustion features with faunal remains and the burial of a cattle (Bos taurus) in anatomical connection. This site comprises the largest Early Bronze Age necropolis in the Barcelona area: a minimum number of 102 individuals have been identified in 12 funerary structures (2 burial chambers with access shafts, 2 burial chambers with access shafts and 5 burial pits). In addition to this, funerary structures from the Middle Neolithic and Chalcolithic periods have also been identified. The 14C analysis of seven individuals revealed the period of use of the necropolis: between ~ 2300 and 1600 cal. BCE (Gómez et al. 2015). The bodies were placed in both primary and secondary positions (the latter being more affected and showing severely degraded bones). The presence of grave goods is very rare: only a few complete ceramic vessels and some shards of Epi-Bell Beaker from the Northeastern Group style (Gómez et al. 2015), V-perforated bone buttons (three of them decorated with oculi), faunal and lithic remains (Velasco et al. 2017) have been recovered.
The Mercat de Santa Caterina (GCS 2° 10′ 43,09432″ E, 41° 23′ 9,90438″ N; ETRS89 UTM zone 31N, X 431326,498046, Y 4581942,99261) is an archaeological site with several periods of occupation. The Early Bronze Age provides little evidence related to diet, as only three silos have been discovered and excavated. The first was refilled with ceramic vessels of the Epi-Bell Beaker style of the Northeastern Group; in the second silo, a young ovicaprine was recovered in anatomic association and without butchery marks, and finally, the last one contained a double inhumation of an adult woman and a child under 3 years-at-death, without any evidence of grave goods (Aguelo et al. 2005; Aguelo and Huertas 2006, 2016). 14C analysis of a faunal and a human bone yielded a chronology of 1880–1720 cal. BCE and 1881–1692 cal. BCE (Gómez et al. 2015).
The present study aims to analyse the buccal microwear pattern to infer the dietary habits of these populations, contextualised with the palaedioetary information currently available, from both the analysis of the material remains and from the archaeozoological and archaeobotanical remains. This study also aims to assess whether differential access to food resources is based on sex or age group. As far as possible social differences are concerned, the Early Bronze Age burial register of the Barcelona plain shows no significant differences in wealth: the majority of the individuals were buried in burial chambers with access shafts in a secondary position and had no or few associated grave goods (Velasco et al. 2017, Gómez and Molist 2016a, b, Carlús and González 2008, Granados et al. 1993, Chambon 2016, Aguelo et al. 2005, Aguelo and Huertas 2006, Aguelo and Huertas 2016, Malgosa 2016). Wealth criteria were discarded from our study because the nature of this type of register does not allow us to observe correspondences between individuals and grave goods.
Materials and methods
Samples included in the study
The initial microwear sample consisted of 442 teeth from 27 different individuals from the Plaça de la Gardunya site, 15 teeth from 2 individuals from the Mercat de Santa Caterina site, 10 teeth from only 1 individual from the Carrer de la Riereta 37-37 bis site and finally 3 teeth from 1 individual from the Solar del Rec Comtal 17-19 (Tables 1 and 2). The sex and age-at-death estimations of the individuals from the Plaça de la Gardunya site were derived from a previous preliminary anthropological study conducted by two of the authors (LM and FE) (Martínez and Estebaranz-Sánchez 2018, unpublished). Sex was determined through standardised osteological traits. Observed skull traits included the morphology and development of the supraorbital ridge, the nuchal crest and the mastoid process (Buikstra and Ubelaker 1994). The pubic portion of the os coxae was studied through qualitative analysis, wherein the sciatic notch and the presence or absence of a sharp edge in the medial aspect of the ischiopubic ramus were observed (Brothwell 1981; Hershkovitz et al. 1993). Age-at-death was determined based on dental eruption and occlusal wear pattern (Brothwell 1981), as well as by epiphyseal fusion in subadult and young adult individuals (Brothwell 1981; Cardoso 2008a,b; Shapland and Lewis 2013). For the site of the Mercat de Santa Caterina, the data was obtained from the original excavation reports (Aguelo and Huertas 2006).
Only one tooth per individual was selected for microwear pattern analysis, preferably in situ left mandibular M2 or M1 (Pérez-Pérez et al. 1994; Lalueza et al. 1996; Estebaranz et al. 2009). Previous studies have excluded that there are significant differences in the buccal MW pattern according to both maxilla and tooth laterality (Pérez-Pérez et al. 1994, Lalueza et al. 1996, Estebaranz et al. 2009). Thus, if neither of these two teeth showed a good microwear pattern, the selection criterion was first the lower left P4, and if this was also absent, then the left mandibular P3 or the left mandibular M3 in its defect. If no lower left tooth had pristine enamel, the selection algorithm continued with the upper left, lower right and finally the upper right dentition. This procedure resulted in an initial sample of 20 permanent teeth/individuals from Plaça de la Gardunya and only 1 permanent tooth/individual from Mercat de Santa Caterina (Tables 1 and 2). The individual from Carrer de la Riereta 37-37 bis was initially excluded because no pristine microwear pattern was preserved in any of the teeth.
To reconstruct the palaeodiet of the population of the Barcelona plain, a large comparative sample of 153 individuals from different populations of hunter-gatherer, pastoralist and agriculturalist groups with different known diets was included (Inuit (n = 20), Fuegians (n = 20), Khoe-Saans (n = 15), Australian Aborigines (n = 18), Andamanese (n = 18), Vancouver Islanders (n = 17), Veddahs (n = 9), Tasmanians (n = 18), Sámi (n = 5) and Hindu (n = 18)) (Lalueza et al. 1996, Jarošová et al. 2006) (Table 3).
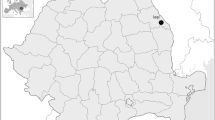
In addition, for comparison and contextualisation within the archaeological context, a second comparative sample from those archaeological populations of the Iberian Peninsula (of variable chronology ranging from the Lower Magdalenian to the Middle Ages, but preferably corresponding to Bronze Age sites) for which buccal microwear data had been published was included (Fig. 1). This archaeological comparison sample included two pre-Neolithic sites: the Lower Magdalenian site of El Mirón Cave (n = 1; only represented by a single lower second premolar; García-González et al. 2015) and the Mesolithic site of El Collado (n = 6; Alrousan et al. 2013). Several Neolithic sites were also included, for which its pattern of buccal microwear was referenced: Can Gambús (n = 8; Alrousan 2009), Caserna de Sant Pau del Camp (n = 11; Estebaranz et al. 2008), Tossal de les Basses (n = 8; Salazar-García et al. 2016) and Cova de la Guineu (n = 15, although the whole sample consists exclusively of child individuals; Hernando et al. 2020). The Chalcolithic comparison sample includes a total of 4 sites: El Molinico (n = 16; Romero and De Juan 2007), El Mirador Cave (n = 2, both infantile; Hernando et al. 2020), El Portalón de la Cueva Mayor (n = 2; García-González et al. 2019) and El Alto de la Huesera (n = 6; García-González et al. 2019). Regarding the Bronze Age, up to 5 sites providing data on buccal microwear have been identified: Cabezo Redondo (n = 16; Romero and De Juan 2007), Cova dels Blaus (n = 6; Polo-Cerdà et al. 2007), Mar i Muntanya (n = 3; Galbany et al. 2008), Cova de Valdavara (n = 2, both children; Hernando et al. 2020) and Cova dels Galls Carboners (n = 9, all children; Hernando et al. 2020). Both the Iron Age site of Son Real (n = 26; Jarošová et al. 2006) and the Late Roman site of Tossal de les Basses (n = 26; Salazar-García et al. 2016) have been included. Finally, four Middle Age sites were also considered: La Olmeda (n = 65; Pérez-Pérez et al. 1994), La Losilla (n = 12; Romero and De Juan 2007), Tossal de les Basses (n = 6; Salazar-García et al. 2016) and L’Esquerda (n = 13; Lalueza and Pérez-Pérez 1989).

Location of the 21 Iberian archaeological sites for which buccal microwear patterns were published
Buccal microwear analysis
Buccal microwear is a dietary proxy (Pérez-Pérez 2004; Davis and Pineda-Munoz 2016) extensively used both in paleoanthropology (Lalueza et al. 1996; Estebaranz et al. 2009, 2012; Martínez et al. 2004, 2015, 2016; Pérez-Pérez et al. 1999, 2003; Pinilla Pérez et al. 2011) and archaeology (Alrousan 2016; Alrousan et al. 2019; Estebaranz et al. 2008; García-González et al. 2015, 2019; Gluitz 2020; Hernando et al. 2020; Jarošová 2007, 2008; Jarošová et al. 2006, 2016; Jarošová and Tvrdý 2017; Kolář et al. 2012; Pérez-Pérez et al. 1994; Polo-Cerdà et al. 2007; Romero and de Juan 2007; Romero et al. 2004; Salazar-García et al. 2016; Domonkošová Tibenská et al. 2010) to infer palaeodietary habits. The buccal dental microwear pattern is formed by the interaction between the abrasive particles harder than enamel contained in the food bolus and the enamel surface during the chewing process, leaving marks exclusively in the form of striations (Puech, Prone and Kraatz 1980). The origin of these abrasive particles can be diverse, as they can be intrinsic elements to the food consumed, such as phytoliths, or extrinsic, such as dust, quartz or silica from the soil (Lucas et al. 2013). In any case, the compressive forces between the cheek and the lateral side of the teeth and the movements of particles in the mouth may be sufficient to form microwear striations on the buccal surface of the tooth (Romero and De Juan 2007; Romero et al. 2009). Thus, buccal dental microwear depicts the abrasiveness of the food consumed by individuals, the physical properties of chewed foods and long-term trends in dietary preferences (Romero et al. 2012, 2013).
In general, the study of the microwear pattern of a given archaeological population is not informative per se, unless it can be contextualised with reference models of buccal microwear of known dietary populations for purposes (Lalueza et al. 1996). Otherwise, we can only make relative statements about the abrasiveness of the diet by comparing it within a population (according to variables such as sex, age and social status) or between populations (by comparing the microwear patterns of different populations or sites). In this sense, without a comparative model of microwear patterns from a known dietary model, a finer reconstruction of the palaeodiet is not possible, even though we may affirm that one population/site, sex, cohort or dietary group has a more abrasive microwear pattern than another. In order to avoid these gaps in palaeodietary research, the present study included a reference model of modern populations with different and distinct diets (Lalueza et al. 1996): 153 individuals from 10 modern groups from the 19th century. They lived in different types of environments and their subsistence economy strategies were based on hunting and gathering, fishing, agriculture or pastoral nomadism. Nevertheless, even when reference models of known diets are available, microwear results should be contextualised with those of other dietary proxies such as stable isotope analysis, faunal and botanical remains, organic residue analysis in ceramic vessels, starch analysis from macrolithic tools or dental calculus analysis. (Pérez-Pérez et al. 2018; Davis and Pineda-Munoz 2016).
Dental casting and SEM imaging
Only teeth with a good preservation of the enamel on the vestibular surface were selected (Estebaranz et al. 2009, 2012). A visual examination was initially done at the Museu d’Història de Barcelona (MUHBA) discard all the teeth that had an evident badly preserved enamel. We made replicas of the selected teeth to further examine them in the laboratory. Human dental crowns were first cleaned with pure acetone using a cotton swab to remove adhering grit, dust or preservatives and then with ethanol, and later left to air dry (Martínez et al. 2020). High-resolution negative moulds were first obtained from the original teeth hosted in the Museu d’Història de Barcelona (MUHBA, Barcelona) using President MicroSystem Regular Body (Coltène’s Affinis) polyvinylsiloxane, an extremely accurate impression material (Beynon 1987; Teaford and Oyen 1989) widely used in odontology and a current standard for microwear studies (Alrousan et al. 2019; Estebaranz et al. 2008, 2009, 2012; Hernando et al. 2020; Jarošová 2007, 2008; Jarošová et al. 2006, 2016; Jarošová and Tvrdý 2017; Martínez et al. 2015, 2016, 2020; Pérez-Pérez et al. 2017). A rigid two-component polyurethane resin (FEROPUR PR55+E5; Feroca®, Spain) was poured into the moulds and allowed to harden in order to obtain high-resolution positive casts (Estebaranz et al. 2012; Martínez et al. 2020). The moulds containing the polyurethane resin were then centrifuged at 1000 rpm for 1 min to prevent the formation of air bubbles on the buccal enamel surfaces (Martínez et al. 2020). Once the material was dry, the replica was removed from the mould and examined under an Olympus SZ61 binocular lens at × 40 magnification. With this process, we were able to discard all the dental pieces that showed postmortem alterations (Pérez-Pérez et al. 2003; Martínez and Pérez-Pérez 2004). Postmortem damage is a reality that affects the correct preservation of the enamel and poses problems for microwear dietary interpretations (Espurz et al. 2004; Estebaranz et al. 2007; King et al. 1999; Martínez and Pérez-Pérez 2004; Rivals and Semprebon 2010; Teaford 1988; Weber et al. 2021a, 2021b). The most immediate consequence of the presence of postmortem signals is the resulting drastic reduction in the size of tooth sample that can be analysed for microwear (Grine et al. 2006a, 2006b; Estebaranz et al. 2009, 2012; Martínez et al. 2016, 2020; Ungar et al. 2008, 2020). The second selection of teeth was mounted on aluminium stubs with term fusible gum and sputter coated for SEM observation with a 5 nm of gold-palladium using an Emitech K550X with a K250 Coater Carbon Attachment at the Servei de Microscopia of the Autonomous University of Barcelona (Gluitz 2020). Following the K550X Sputter Coater manual, we set the duration of each sputtering cycle to 1 min through argon gas at a working distance of 50 mm, a pressure of 0.05 mbar and an intensity of 25 mA.
SEM observation followed the standard methodology for buccal microwear analysis (Pérez-Pérez et al. 1994; Estebaranz et al. 2009; Martínez et al. 2020). SEM micrographs were obtained using a Zeiss EVO ® MA 10 scanning electron microscope in high vacuum mode at the Servei de Microscopia of the Autonomous University of Barcelona. SEM parameters were fixed at 15-kV acceleration voltage and 20 to 30 mm working distance. Each micrograph was taken at × 100 magnification preferably on the middle third of the buccal surface, avoiding both the occlusal and cervical rims. SEM images were cropped to a fixed area of 0.56 mm2 using Adobe Photoshop v.6 (Pérez-Pérez et al. 1994; Jarošová et al. 2016) and contrast-enhanced, following previous recommendations to facilitate manual count of the microstriations (Pérez-Pérez et al. 1994; Estebaranz et al. 2009) (Fig. 2). SigmaScan Pro 5.0 (SPSS™ v.15) was used to semi-automatically count striations longer than 15 µm and with a length-to-width ratio of 4:1. This provided the length and slope of each striation observed (Estebaranz et al. 2009, 2012; Martínez et al. 2020). Both inter- and intra-observer errors are a major problem (Kouchi et al. 1999) and also affect microwear analysis (Galbany et al. 2005; Grine et al. 2002 Mihlbachler et al. 2012). In order to minimize the standard intra-observer measurement error and to obtain a higher precision of the measurements, we followed the recommendations of Pérez-Pérez et al. (1990), which consists in repeating a series of measurements and taking the average of the repetitions as the actual measurement. Therefore, all SEM microwear images were measured 3 times on different days by only one observer (AG).

Cropped images of the dental microwear pattern from different individuals: a LLM2 from individual 4328 (Plaça de la Gardunya), b LRM3 from individual 4319 (Plaça de la Gardunya), c LRM2 from individual 50574-1 (Plaça de la Gardunya), d URM2 from the individual 5516 (Mercat de Santa Caterina), e URM2 from the individual 50616 (Plaça de la Gardunya) and f LLM1 from the individual 50620 (Plaça de la Gardunya)
Using IBM® SPSS® Statistics v.21, the slope of each striation was converted to an angle and was classified into one of four different orientations: horizontal (H), vertical (V), mesio-occlusal to disto-cervical (MD) and disto-occlusal to mesio-cervical (DM) (Pérez-Pérez et al. 1999, 2003). For each orientation (H, V, MD and DM), the number (N), the average (X) and the standard deviation (SD) of the length were calculated, as well as the sum of all striation types (T). From this, the complete buccal microwear pattern was derived, consisting of 15 quantitative variables (NH, XH, SDH, NV, XV, SDV, NMD, XMD, SDMD, NDM, XDM, SDDM, NT, XT, SDT) (Pérez-Pérez et al. 1994, 1999, 2003). In addition to this, three other indices were calculated: the number of horizontal striations divided by the number of vertical striations (NH/NV), the number of horizontal striations divided by the striation density (NH/NT) and the number of vertical striations divided by the striation density (NV/NT) (Pérez-Pérez et al. 1994; Lalueza et al. 1996).
Statistical analysis
All the SEM microwear patterns corresponding to the Plaça de la Gardunya and Mercat de Santa Caterina sites were counted by one single author (AG). All statistical analyses and comparisons were performed using IBM® SPSS® Statistics v.21 at α = 0.05 significance level and Gnuplot v.5.4. When analysing the normality of the Plaça de la Gardunya microwear data, only 7 of the original variables passed the Kolmogórov-Smirnov test (SDH, XV, SV, SDMD, SDDM, NT and SDT). Similar results were obtained using the Shapiro-Wilk test, as only 5 variables (XV, SV, SDMD, NT and SDT) passed the normality test. In addition, six variables (NH, XH, NV, NDM, XDM and XT) did not pass Levene’s test of homoscedasticity (p-value < 0.05) when comparing the homogeneity of variance of the microwear data from Plaça de la Gardunya with those from other nearby Neolithic sites (Can Gambús and Caserna de Sant Pau del Camp). Consequently, non-parametric statistics were applied: group comparisons were made using the Kruskal-Wallis H test (Estebaranz et al. 2012). Post-hoc intergroup pairwise comparisons between archaeological sites (Plaça de la Gardunya, Mercat de Santa Caterina, Caserna de Sant Pau del Camp and Can Gambús) were performed using a Mann-Whitney U test (Teaford and Glander 1991; Estebaranz et al. 2012; DeSantis and Haupt 2014; Louys et al. 2021). Finally, the differences between buccal dental microwear from Plaça de la Gardunya and the hunter-gatherer models of known diet were also evaluated using the Mann-Whitney U test.
A stepwise predictive discriminant function analysis (LDA) was also performed to classify the microwear pattern of the Plaça de la Gardunya archaeological site from the hunter-gatherer comparison samples and to better profile and identify the variables that best explain the differences between the populations in the comparison sample of known diets (Lalueza et al. 1996; Pérez-Pérez et al. 1999; Mainland 2003; Grine et al. 2006a; Mahoney 2006; Romero and De Juan 2007). Prior probabilities of group membership were computed as a function of group size, and separate within-group covariance matrices were used for classification due to significant differences between covariance matrices. Wilk’s Lambda statistic was used to characterize the reliability of the analyses.
Results
The presence of taphonomic alterations has had a significant impact on the size of the original tooth sample (Table 1). Of the total initial sample considered in this study, only 30% of the teeth (141 out of 470 teeth) corresponding to 21 individuals (20 from the Plaça de la Gardunya and 1 from the Mercat de Santa Caterina) were finally included in the study. By site, in the Plaça de la Gardunya site, 187 out of 442 teeth were initially discarded using the binocular magnifying glass. Subsequently, during the SEM observation, of the 255 teeth that were analysed, a further 115 teeth were finally discarded because they did not show completely well-preserved buccal enamel surfaces at a magnification of × 100 (Martínez and Pérez-Pérez 2004). In this sense, SEM observation allowed discarding teeth that did not have a pristine buccal enamel surface but showed slight (n = 57) or moderate taphonomic affectation (n = 58) and were initially considered potentially good by binocular loupe. A similar situation was observed for the other three sites: at the Mercat de Santa Caterina, initially 3 out of the original 15 teeth were initially discarded. Through the use of a binocular lens, we observed that out of the remaining 12 teeth, only one had a good microwear pattern. As a result, the remaining 11 teeth were finally discarded, including all the teeth belonging to the child individual. In Carrer de la Riereta 37-37 bis, only 6 of the 10 original teeth were examined under the SEM microscope, which showed that none of them preserved a pristine buccal enamel surface. Upon visual examination, the 3 teeth from a single individual from Solar del Rec Comtal 17-19 were ultimately discarded due to significant taphonomic alterations (Table 1). The average striation density (NT) of the Plaça de la Gardunya site is 72.53 ± 44.30 and for the Mercat de Santa Caterina 11.33, the average length (XT) is 147 ± 30.93 and 174.31, respectively (Table 4). Individualized data for all individuals included in the study are detailed in Table 4.
Intrapopulation variability
The scatter plot of NT vs. XT for both sexes suggests that male individuals present a certain homogeneity in their MW pattern (XT values ranging from ~ 150 to 190 and XT values from ~ 25 to ~ 75, excluding UE4328), in comparison to females, which show a greater range of distribution (with XT values ranging between ~ 50 and ~ 200 and NT values between ~ 120 and ~ 180) (Figure S1, Supplementary Information). If we look at the index of the relative frequency of NH/NV, females have a mean value of 0.37 while males, 0.15 (Table 5). However, no statistically significant differences were observed for any of the 15 variables (Table 6) for the sex factor (non-parametric Mann-Whitney test; 3 females vs. 8 males) (Table S2, Supplementary Information). The only variable that falls close to the significance value is XV (Mann-Whitney U = 21.00; p-value: 0.066). On the other hand, when analysing MW differences based on age, subadult individuals present statistically higher NT (Mann-Whitney U = 75.00; p-value = 0.018) (Figure S2; Table S3 Supplementary Information), as well as a higher NH/NV ratio than adults (0.21 vs. 0.15, respectively) (Table 5).
Comparison with populations of known diet
With regard to modern populations with known diets, the microwear pattern of the Plaça de la Gardunya also shows a lower NT value than that of agriculturalists (NT = 181.14 ± 47.90), gatherers (NT = 243.15 ± 60.79), hunters (NT = 190.46 ± 64.35) and above all fishermen (NT = 268.56 ± 80.32). Nevertheless, the average length of striations (XT) of the Plaça de la Gardunya population (XT = 147.46 ± 30.93) is similar to that of the agriculturalists (XT = 151.12 ± 20.45), gatherers (XT = 137.91 ± 19.02) and fishermen (XT = 140.09 ± 19.81) and almost double that of the hunters (XT = 71.81 ± 56.74) (Table 4). The boxplots of both variables, total striation density (NT) and average striation length (XT), for the two Bronze Age sites (Plaça de la Gardunya and Mercat de Santa Caterina), as well as for the comparative human populations with known diets, are shown in Fig. 3. As already mentioned, the low density of striations of the Bronze Age sites is highlighted, although the range of dispersion of striation lengths overlaps with that of populations with an agricultural diet, gatherers and fishermen and is clearly superior to hunters with a high incorporation of meat in their diet.

Boxplot showing the density of striations (NT) and the average length of striations (XT) of each known diet (agriculturalists, gatherers, hunters and fishermen) and the archaeological sites of Plaça de la Gardunya, Mercat de Santa Caterina, Can Gambús and Caserna de Sant Pau del Camp
When comparing the microwear pattern of the Plaça de la Gardunya and Mercat de Santa Caterina site with the reference sample (Table 4), statistically significant differences were found for all the variables considered (Table 7). The pairwise comparison between the Plaça de la Gardunya/Mercat de Santa Caterina and the hunter-gatherer samples, using Mann-Whitney U tests, showed statistically significant differences in NH with agriculturalists (U = 0.00, p = 0.000), gatherers (U = 4.000; p = 0.000), hunters (U = 13.500, p = 0.000) and fishermen (U = 6.000; p = 0.000); XH with hunters (U = 44.000, p = 0.000); SDH with agriculturalists (U = 35.000, p = 0.41), hunters (U = 128.000; p = 0.013); NV with agriculturalists (U = 33.000, p = 0.32), gatherers (U = 54.000, p = 0.000), hunters (U = 53.000, p = 0.001) and fishermen (U = 72.000; p = 0.000); XV with hunters (U = 35.000, p = 0.000); SDV with hunters (U = 48.000, p = 0.000); NMD with gatherers (U = 60.000; p = 0.000), hunters (U = 68.000, p = 0.04) and fishermen (U = 149.000; p = 0.000); XMD with hunters (U = 80.000; p = 0.013); NDM with agriculturalists (U = 19.000; p = 0.004), gatherers (U = 29.000, p = 0.000), hunters (U = 45.000; p = 0.000) and fishermen (U = 78.5000; p = 0.000); XDM with agriculturalists (U = 31.000, p = 0.024), gatherers (U = 112.000; p = 0.011), hunters (U = 96.000; p = 0.048) and fishermen (U = 325.000; p = 0.003); SDDM with agriculturalists (U = 24.000; p = 0.009), gatherers (U = 128.000, p = 0.032) and fishermen (U = 329.000; p = 0.004); NT with agriculturalists (U = 7.000; p = 0.000), gatherers (U = 4.000; p = 0.000), hunters (U = 18.000; p = 0.000) and fishermen (U = 9.000; p = 0.000); XT with hunters (U = 49.000; p = 0.000) and SDT with hunters (U = 51.000; p = 0.001) (Table S1, Supplementary Information).
Previous studies have highlighted the importance of plotting the relationship between the percentage of horizontal microstriations relative to the total number of striations (NH/NT) and the percentage of vertical striations (NV/NT) on a bivariate axis (Lalueza et al. 1996; Pérez-Pérez et al. 1994, 1999; García-González et al. 2015) (Fig. 4a, b). The individuals of Plaça de la Gardunya show a great percentage of vertical striations in comparison to the comparison sample grouped according to major dietary categories (hunters, agriculturists, fishermen and gatherers) with a wide dispersion ranging from ~ 0.15 to 0.8 (Fig. 4a). These values are high compared to those observed for both hunters and agricultural populations, while they overlap slightly with those for gatherers and fishermen (although they are clearly separated for the horizontal axis) (Fig. 4a). When comparing the mean NV/NT value of Plaça de la Gardunya with the means of the different original populations (i.e. when the original populations considered are not grouped into general diet categories), the Plaça de la Gardunya site falls close to the Fuegians of Tierra del Fuego (Argentina and Chile) and the Vancouver Islanders (Fig. 4b). The Sámi and Inuit populations also fall close to the Plaça de la Gardunya, although a little more separated for the vertical axis, i.e the Plaça de la Gardunya population (as well as the Fuegians and the Vancouver Islanders) has slightly higher values of vertical striations (mean NV/NT index). In terms of the NH/NT index, the Plaça de la Gardunya microwear pattern has a lower percentage of horizontal striations (ranging from 0.01 to 0.1), which clearly distinguishes the Placa de la Gardunya from the comparative sample of dietary categories (Fig. 4a). Thus, for example, the percentage of horizontal striae ranges from 0.1 to 0.4 for fishermen, 0.08 to 0.4 for gatherers, 0.05 to 0.2 for hunters and 0.15 to 0.35 for agriculturalists (Fig. 4a). When comparing the Plaça de la Gardunya with the original comparison sample by original populations, Plaça de la Gardunya again clusters with the same populations as for the percentage of vertical striations, with an NH/NT index virtually identical to that of the Vancouver Islanders and slightly lower than that of the Fuegians, Inuit and Sami (Fig. 4b). Conversely, the populations of Tasmanians, Australian Aborigines, Andamanese, Veddahs and Khoe-Saans have much higher values (between ~ 0.2 and ~ 0.3), while the Hindus have much more extreme values for NH/NT (~ 0.5). Therefore, the Plaça de la Gardunya microwear pattern differs from that of the known dietary comparison sample for NH, while it is grouped with that of more maritime diets (Fuegins, Vancouver Islanders, Sámi and Inuit) for vertical striae percentage. In fact, by directly comparing the ratio of the total vertical striae to the total horizontal striae (NH/NV) in a boxplot (Fig. 5), the Bronze Age individuals (PG and MSC) are clearly separated from the comparison sample, with much lower values, indicating a very low ratio of horizontal striae to the total number of vertical microstriations.

a NH/NT index plotted against NV/NT index from known diets and archaeological sites. b NH/NT index plotted against NV/NT index. Black-filled dots symbolize the values from reference microwear patterns (Lalueza et al. 1996), and the cross symbolizes the value from Plaça de la Gardunya’s population. Populations with a higher NV/NT index and a lower NH/NT index have a diet based on animal protein (fish and meat), and populations with a lower NV/NT and a higher NH/NT have a mixed or agriculturalist diet

Boxplot of NH/NV index in which we can observe the values from reference microwear patterns (Lalueza et al. 1996) and the values from Plaça de la Gardunya and Mercat de Santa Caterina. Lower values belong to populations with higher animal protein (fish and meat), while higher values belong to populations with a diet based basically on vegetables and processed agricultural products. Boxplot showing the NH/NV index from each known diet and from the archaeological sites sampled for this study (Plaça de la Gardunya and Mercat de Santa Caterina)
In the predictive LDA, the microwear pattern of the individuals from Plaça de la Gardunya and Mercat de Santa Caterina sites was classified according to the comparison sample (hunters, gatherers, agriculturalists and fishermen). Only 3 discriminant factors (DF) were obtained, explaining 100% of the original total variability. The first two DFs, which accounted for 99% of the total variability, showed significant Wilks’ lambda (0.832, p < 0.002). The DF1 (explaining 87.4% of the original variability; canonical correlation: 0.764) was significantly correlated with XV (0.857), XH (0.830) and NT (0.278), whereas DF2 (12.5% of the variability; canonical correlation: 0.408) was correlated with XV (0.022), XH (−0.541) and NT (0.718). In addition to this, differences in matrix covariances between groups were observed (Box’s test F: 3.295; p = 0.000). Figure 6 shows the microwear pattern of the hunter group completely separated from the other groups by DF1 (negative loadings), indicating low horizontal and vertical striation length, as shown in Table 6. The other 3 known dietary groups (agriculturalists, gatherers and fishermen) overlap strongly for both DF1 and DF2. However, there is a tendency for fishermen’s dietary group to have the highest values for both DF1 and DF2. In this sense, fishermen are separated from gatherers by having higher DF1 values (displaced more to the right of the image). On the other hand, individuals with an agriculturalist diet, although they partially overlap with the other groups for DF1, tend to have lower values for DF2, being partially separated from the other groups. In any case, both agriculturalists and hunters are separated by DF2, as the latter has higher values for this discriminating function.

Linear discriminant analysis showing that the buccal microwear patterns from Plaça de la Gardunya and Mercat de Santa Caterina have similar values to the diest of agriculturalists, gatherers and fishermen
The post-hoc probability of correct classification of the original cases was 69.1%. Looking at each dietary group separately, the group with the best percentage of correct classification were the fishermen, with 92.7% (51/55) of the individuals correctly classified individuals (2 individuals −3.6% were classified as hunters and 2 as agricultural −3.6%). The hunter group also showed a clearly differentiated and unique microwear pattern, with 73.3% (11 of 15) of individuals correctly classified, while the remaining 26.7% (4 of 15) were classified as fishermen (and none as gatherers or farmers). Figure 6 shows that the hunters had a different microwear pattern and were clearly separated from the other dietary groups by DF1. For the farmers, 28.6% (2/7) were correctly classified, while 14.3% (1/4) were classified as gatherers and 57.1% (4/7) as fishermen. Finally, only 15% of the gatherer individuals were correctly classified (3 out of 20), while the vast majority was incorrectly classified as fishermen (80%, 16 out of 20). When individuals of archaeological origin were classified according to the microwear patterns of known dietary groups, most were classified as fishermen (66.7%, 14/21). However, it is important to note that although 14 of the 21 individuals were initially classified as fishermen, the second classification for 10 of the individuals was gatherer.
Comparison with other archaeological sites
The second part of the study contextualised the buccal microwear patterns of Plaça de la Gardunya and Mercat de Santa Caterina with those published for other Iberian archaeological sites. In this sense, the microwear density of Plaça de la Gardunya is relatively low (NT = 80.73 ± 41.85) compared to other more ancient archaeological sites in the same region of the Barcelona plain, such as the Neolithic sites of Can Gambús (NT = 156.62 ± 60.43) and Caserna de Sant Pau del Camp (NT = 137.88 ± 43.76). It is similar to that of the contemporary site of Mercat de Santa Caterina (NT = 62.33) (Fig. 3, Table 8).
The microwear pattern was then compared with 17 other Iberian archaeological sites (Fig. 7). For most of these sites, only MW averages have been published. The scatter plot of XT vs. NT shows that both Plaça de la Gardunya and Mercat de Santa Caterina form a distinct cluster with the sites of La Olmeda (Palencia) and Son Real (Mallorca), with values ranging from 55.22 to 87.69 for NT and 136.08 to 174.31 for XT. A second group of sites with lower XT and higher NT values consists of 11 sites with different chronologies: El Mirón cave (Cantabria), El Collado (Alicante), Tossal de les Basses’ Late Roman (Alicante), El Alto de la Huesera (Ávila), El Portalón de la Cueva Mayor (Burgos), Cabezo Redondo (Murcia), La Losilla (Murcia), Mar i Muntanya (Barcelona), Cova dels Blaus (Castellón), Caserna de Sant Pau del Camp (Barcelona) and Can Gambús (Barcelona). This cluster presents a range for NT that oscillates between 80.25 and 156.62 and for XT between 88.50 and 116.86. The last cluster includes the Neolithic and Mesolithic sites of Tossal de les Basses (Alicante) and El Molinico (Murcia). This group ranges from NT = 257.167 to 339.87 and XT = 55.63 to 98.86. Both NT and XT show high variability between sites and over the same time period for children and subadults, as well as for adults (Figure S3, Supplementary). In any case, focusing only on the Bronze Age sites and adult individuals, both the Plaça de la Gardunya and the Mercat de Santa Caterina have a slightly lower NT and a higher XT than the other Bronze Age sites examined (i.e. Cabezo Redondo, Cova dels Blaus, Mar i Muntanya) (Fig. 8). In children, however, these differences in NT and XT are not observed between the children from the Plaça de la Gardunya site (because no child with MW was found in the Mercat de Santa Caterina) and the other two bronze deposits with infantile MW, the Valdavara and Galls Carboners caves (Fig. 8). Children and subadults have a lower density of striations than adults, but overall, the length of striations does not differ greatly between age groups (Figure S3, Supplementary Information).

Density of striations (NT) plotted against mean length of striations (XT in µm) from adult individuals with error bars, which represent the standard deviation of the mean length of striations. Each colour and point type symbolizes a different period: black cross = Lower Madgalenian, purple cross = Mesolithic, brown square = Neolithic, green dot = Chalcolithic, gold inverted triangle = Bronze Age, grey rhombus = Iron Age, pink square = Late Roman and blue triangle = Middle Age

Density of striations (NT) plotted against length of striations (XT in µm) from children (0–12 years old) and subadult (< 20 years old) individuals with error bars, which represent the standard deviation of the mean length of striations. Each colour and point type symbolizes a different period: brown square = Neolithic, green dot = Chalcolithic and gold inverted triangle = Bronze Age
Discussion
The buccal microwear analysis carried out in this project allows us to expand and precise the knowledge about dietary habits of the Bronze Age’s populations from the Northeastern region of the Iberian Peninsula. The consistency of the method of analysis, the abundant references and the contrast with the archaeological record make this research a solid contribution to the subsistence economy strategies of prehistoric populations. The interpretation of the microstriation patterns analysed in this study has been based on the reference patterns of known-diets, on the density of striations (NT) plotted against the mean length of striations (XT) and on three indices of relative frequency (NH/NT, NV/NT and NH/NV) analysed and published by Lalueza et al. 1996 and tested by numerous authors.
The first criterion is to observe the density of striations (NT). A high density means the food consumed was more abrasive, while a low density would mean just the opposite (Hernando et al. 2020; García-González et al. 2019). This criteria would make a difference between the consumption of harder foods or having a softer diet. The second criterion is to look at the indices of relative frequency of striations by its orientation. Lalueza et al. (1996) observed that the ratio of the number of horizontal striations divided by the density of striations (NH/NT) plotted against the ratio of the number of vertical striations divided by the density of striations (NV/NT) showed different results in each modern population they studied. A lower NH/NT ratio and a higher NV/NT ratio were related to the regular consumption of animal protein (meat or fish), while a higher NH/NT ratio and a lower NV/NT ratio were related to the consumption of more agricultural foods (García-González et al. 2015). One more index of relative frequency, the number of horizontal striations divided by the number of vertical striations (NH/NV), has to be considered as well. On the one hand, low NH/NV ratios tend to indicate a low mandibular compression and the chewing of soft foods with low fibre content. It has been observed in populations with an animal protein-based diet (meat or fish) (Lalueza et al. 1996). On the other hand, high NH/NV ratios tend to indicate a high mandibular compression and the chewing of hard, low-refined and highly abrasive foods (Lalueza et al. 1996; Romero et al. 2012, Salazar-García et al. 2016; Jarošová et al. 2016; Radini et al. 2017, García-González et al. 2019; Hernando et al. 2020). It has been observed in populations with a mixed or agricultural diet (Lalueza et al. 1996). The NV/NT and NH/NT ratios of Plaça de la Gardnya are similar to those of populations such as Fuegians, Vancouver Islanders, Sámi and Inuit, all of them being hunter-gatherer and pastoralist populations with a mainly carnivorous diet (Fig. 3).
Data of other archaeological sites with published microwear studies allowed us to compare our results and further refine the diet that Plaça de la Gardunya and Mercat de Santa Caterina’s prehistoric population had in the Early Bronze Age. The plot of density of striations (NT) against mean length of striations (XT) for adult individuals reveals the presence of three clusters. The first one includes the sites Plaça de la Gardunya, Mercat de Santa Caterina, La Olmeda and Son Real. Their range of values show a low density of very long striations, being a possible indicator of a diet based in soft foods. The values observed are consistent with the consumption of meat derived from husbandry practices at the site of Plaça de la Gardunya. Animal remains indicate the exploitation of domestic species such as Bos taurus, Ovis aries and Capra hircus (Alcàntara 2018). Zooarchaeological analysis suggests that bovines were slaughtered at an older age, typically between 3 and 6 years, with one individual older than 8 years old. The youngest specimen was slaughtered at the age of 2 years, when it reached its meat optimum. Cut marks are abundant, suggesting animal processing activities such as skinning, evisceration, dismemberment and butchering. The intensive use of this species is evidenced by the traces of brain and bone marrow consumption. Ovicaprids, on the other hand, were slaughtered when their meat was at its optimum, possibly for secondary products such as milk. There is no evidence of cooking techniques on the animals, such as thermal alterations (Alcàntara 2018), suggesting that only meat and organs may have been cooked, possibly to reduce chewing effort (Hernando et al. 2022). Apart from Bos taurus, Sus domesticus was exploited with similar intensity. The consumption of wild animals may have been possible through hunting, as remains of wild rabbits (Oryctolagus cuniculus) have been found in Plaça de la Gardunya (Alcàntara 2018). Fish consumption is also considered, given the discovery of a fishing net weight at the Carrer de la Riereta 37-37 bis site (Bofill 2016). The significant presence of agricultural evidence in the archaeological register, in contrast to the results from the NV/NT and NH/NT ratios, as well as the resultos of NT and XT results, raises the question of why their buccal microwear pattern is low abrasive if agricultural products are high abrasive. Carpological analyses in the Barcelona plain have recovered several domesticated plant species, such as barley (Hordeum vulgare and Hordeum vulgare nudum), four types of wheat (Triticum aestivum, Triticum durum, Triticum compactum and Triticum dicoccum) and legumes (Vicia faba) (Gómez and Molist 2016a, b). Storage structures for agricultural products are abundant, with the presence of 4 silos in Plaça de la Gardunya, 1 in Filmoteca de Catalunya, 3 in Carrer de la Riereta 37-37 bis, and 3 more in Mercat de Santa Caterina (Velasco et al. 2017; Nadal and Castillo 2009; Carlús and González 2008; Aguelo et al. 2005), all with high volumetric capacities. In addition, grill-plan structures interpreted as elevated granaries have been found in Filmoteca de Catalunya and Carrer de la Riereta 37-37 bis (Nadal and Castillo 2009; Carlús et al. 2010). In Carrer de la Riereta 37-37 bis, large and medium-sized storage jars for cereals were also found (Gómez and Molist 2016a, b). In addition, various lithic tools used for land preparation, harvesting and processing agricultural products, such as axes, hoes and grindstones, have been found at the sites of Illa d’en Robador, Filmoteca de Catalunya, Plaça de la Gardunya, Carrer de la Riereta, Caserna de Sant Pau del Camp and Solar del Pou de la Figuera (Bordas 2016; Piera et al. 2005; Nadal and Castillo 2009; Velasco et al. 2017; Bofill 2016; Granados et al. 1993; Gómez and Molist 2016a, b; Suau and Matas 2002). The discovery of cooking pots in Carrer de la Riereta 37-37 bis (Gómez and Molist 2016a, b) further supports the evidence of a high consumption of agricultural foodstuffs. Dental pathologies, such as periodontal disease, caries, occlusal tooth wear and hypoplasia in 50% of the population of Plaça de la Gardunya (Estebaranz et al., unpublished) suggest a high carbohydrate intake, typical of agricultural societies (Ortner 2003). The presence of abrasive particles in agricultural products could have been minimized by different cooking methods, such as grinding, boiling, fermenting, steaming or roasting. Cereal products could have been consumed in the form of beverages, bread, porridge, semolina or stew, as proposed by Hernando et al. (2022).
For most of the sites included in the second cluster, formed by El Mirón Cave, El Collado, Tossal de les Basses’ Late Roman period, El Alto de la Huesera, El Portalón de la Cueva Mayor, Cabezo Redondo, La Losilla, Mar i Muntanya, Cova dels Blaus, Caserna de Sant Pau del Camp and Can Gambús, diet interpretations have been published. The NH/NT and NV/NT ratios of El Mirón Cave indicate a carnivorous diet with a hunter-gatherer economic subsistence. Additional analyses reveal a mixed but soft diet based on fish, meat and occasionally vegetables (García-González et al. 2015). El Collado’s values and isotope analysis show a marine diet based on dried fish, in which abrasive particles like grit adhered to the fish in the drying process (Alrousan et al. 2013). Tossal de les Basses’ Late Roman period had a soft diet, maybe explained by the presence of wooden mortars that were used for cereal processing instead of stone ones (Salazar-García et al. 2016). For El Alto de la Huesera and El Portalón de la Cueva Mayor, a high meat consumption is observed in the NH/NV ratio (García-González et al. 2019). The hypothesis for Cova dels Blaus’ values is an agricultural and pastoralist economic subsistence, and the main food consumed would be meat and cereals processed with grinding stones (Polo-Cerdá et al. 2007). Cabezo Redondo and La Losilla have been interpreted as a population with a high degree of meat consumption and refined milling techniques which minimized the inclusion of abrasive particles in the resulting food (Romero and De Juan 2007). For Mar i Muntanya, data has shown a very abrasive diet based on processed cereals with grinding stones (Galbany et al. 2008). The interpretation of Neolithic values from Caserna de Sant Pau del Camp and Can Gambús have not been published yet (Alrousan, unpublished dissertation). As we can see, this group is characterized by a mixed diet based on soft foods like meat and fish and more abrasive foods like cereals processed with grinding stones. The last cluster includes the sites Tossal de les Basses’s Neolithic and Middle Age periods and El Molinico. Their values have been interpreted as populations with a mixed diet that had a food intake of low processed foods. These would be very contaminated by abrasive particles like grit and would need a substantial chewing pressure. It is intriguing to see how different populations from the same period have such disparate NT and XT values. We would expect similar processing techniques to be used in the same chronology, but data reveals the high variability of dietary habits in the Iberian Peninsula.
For subadult individuals, a lower density of striations is to be expected because of their young age: the older the individual, the greater the accumulation of striations at the tooth surface. For Cova de Valdavara (Lugo), Hernando et al. (2019, 2020) proposed a mixed diet based on breast milk and processed food with grinding stones, like porridges. For the children of Caserna de Sant Pau del Camp (Barcelona), we interpret the same type of diet because of its similar values. High XT values of El Mirador (Burgos), Cova de la Guineu (Barcelona), Cova dels Galls Carboners (Tarragona) and Plaça de la Gardunya (Barcelona) reveal a very abrasive diet, more focused on the consumption of porridges or other processed agricultural produce with lots of grit or phytoliths. In contrast, lower XT values of Caserna de Sant Pau del Camp’s subadults, El Portalón de la Cueva Mayor (Burgos) and El Alto de la Huesera (Ávila) could originate from a softer diet based on breastfeeding or the consumption of agricultural produce wih low presence of grit or phytoliths.
One of our aims was to investigate social differences in access to food resources. The data provided results that give an insight into possible dynamics within society in the Barcelona plain during the Early Bronze Age. However, this approach should be considered as a preliminary study, as we acknowledge that the number of individuals sampled is small. This comes from a scarce anthropological register in this specific area and chronology. The paucity of remains is a problem that extends to other aspects of the archaeological record, including architectural elements and archaeological artefacts. These results should therefore be interpreted with these limitations in mind. When examining social differences based on sex criteria, we have 11 individuals whose sex has been determined, consisting of 8 males and 3 females. The sample size for each sex is significantly different, which means that the data do not necessarily accurately reflect past dietary habits. Nevertheless, the results of the NH/NV ratio suggest a preliminary trend where females may have consumed foods with a higher presence of abrasive particles compared to males who may have consumed softer foods. A similar pattern has been observed in the settlements of Son Real (Mallorca) (Jarošová et al. 2006) and Caserna de Sant Pau del Camp (Barcelona) (Estebaranz et al. 2008), two Iron Age and Neolithic sites, respectively, in the Iberian Peninsula. In terms of age criteria, we have data on 21 individuals, including 15 adults and 6 subadults. The NH/NV ratio and the relationship between striation density (NT) and mean striation length (XT) indicate the predominance of harder foods in the diet of the subadult population, whereas softer foods would predominate in the diet of the adult population. These results may be due to cooking techniques that reduce the chewability of foods (Hernando et al. 2022), to dental health or to the chewability of each age group, or even to the fact that subadults consume a higher proportion of processed foods contaminated with abrasive inclusions. A similar pattern has been observed in El Portalón de la Cueva Mayor (Burgos) and El Alto de la Huesera (Álava) (García-González et al. 2019), two Chalcolithic sites in the Iberian Peninsula.
Despite all the criteria available to perform an analysis of the dental buccal microstriation pattern, we are aware of its limitations and consider that complementary studies should be performed in order to have more dietary proxies, such as organic residue analysis in ceramic vessels, starch analysis from macrolithic tools, dental calculus analysis (Radini et al. 2017) or stable isotope analysis (Carbon and Nitrogen) of human and animal bones. The analytical method employed can also be highly improved with the use of a confocal microscope to perform microwear texture analysis (Pérez-Pérez et al. 2018; Correia, Foley and Mirazón Lahr 2021), which removes the intra- and inter-researcher error at the counting process because of its automatization.
We would also like to mention the degree of preservation of the samples. As previous studies have stated, postmortem damage greatly affects buccal enamel surfaces (Estebaranz et al. 2007, 2009, 2012; Martínez and Pérez-Pérez 2004; Martínez et al. 2016). In the present study, the percentage of well-preserved teeth stands at 31.7%, a value similar to that published in other studies. Buccal-microwear studies in Plio-Pleistocene hominins tend to agree that the percentage of teeth that preserve good enamel on the buccal face ranges from 15 to 25% (Estebaranz et al. 2009, 2012, Martínez et al. 2016; Pérez-Pérez et al. 2003). The slightly higher values reported in the present study may be due precisely to the fact that are much more recent sites and the individuals were conscientiously buried, which could favour a lower taphonomic impact and, therefore, a higher percentage of teeth well preserved. The percentage of taphonomic impact for the site of the plain of Barcelona should be contextualized with that of other contemporary sites. Unfortunately, in most published buccal microwear studies on archaeological populations, the percentage of well-preserved teeth is not indicated (Alrousan et al. 2013, 2019; García-González et al. 2015, 2019; Hernando et al. 2020; Jarošová 2007, 2008; Jarošová et al. 2006; Jarošová and Tvrdý 2017; Polo-Cerdá et al. 2007). Most articles only include a reference to the absolute number of teeth with good buccal-microwear pattern, but without any reference to the initial number of teeth. In the only Bronze Age site in which the percentage of well-preserved teeth has been estimated, the Mar i Muntanya site (Barcelona area) has a value very similar to that reported for the plain of Barcelona (37.5%—3 out of a total of 10 molars vs. 30.2%, respectively).
Conclusion
The buccal microwear pattern from Plaça de la Gardunya and Mercat de Santa Caterina indicates a predominantly soft diet, a finding that could not be inferred solely from the archaeological data previously analysed. The presence of abundant plant species, lithic tools used for land preparation, harvesting and processing of agricultural produce, as well as silos with high volumetric capacities, storing jars for cereal, and grill-plan structures in close vecinty, along with a high percentage of dental diseases associated with an increase in carbohydrate intake (periodontal disease, caries and occlusal tooth wear) (Martínez and Estebaranz-Sánchez 2018, unpublished) led to the conclusion that agriculture played a significant role in the Early Bronze Age diet in the Barcelona plain. The low-abrasive buccal microwear pattern could be attributed to refined cooking techniques that would reduce chewing effort and thus minimized the abrasiveness (or even prevented its increase) of the agricultural foodstuff. It is likely that milling, steaming, boiling, fermenting or roasting were commonly employed. Food would probably have been consumed in various forms such as stew, bread, porridge, beverages or semolina (Hernando et al. 2022). Alternatively, a portion of the agricultural produce might have been used as feed for livestock. Nonetheless, animal protein also played a significant role in the diet of the population from the Barcelona plain. The evidence of husbandry and the slaughter patterns suggest the consumption of animal products, inclusing meat, organs, bone marrow, milk or cheese, which are low-abrasive for dental enamel. The application of cooking techniques could further reduce its abrasiveness. Fish should also be considered as part of their diet, given the discovery of a fishing net’s ceramic weight with triangular morphology. This study underscores the importance of the water sources, such as the Llobregat and Besòs rivers and the Mediterranean Sea, in shaping the subsistence economy strategies of these communities. For the communities of the Early Bronze Age in the plain of Barcelona, we propose a mixed diet consisting of highly refined and cooked agricultural products, animal protein from cooked meat and fish and the consumption of secondary products. Their subsistence economy strategies likely revolved around a diverse range of activities, with a strong emphasis on agriculture, animal husbandry and fishing, while hunting and gathering had a lesser impact.
We have conducted a preliminary study to determine whether social inequalities existed concerning differential access to food. However, we acknowledge the limitations of the samples and archaeological remains, and therefore, our interpretation should be approached with caution, as results may not precisely reflect past dietary habits. Based on our observations, adult male individuals probably consumed softer foods, such as cooked agricultural produce or primary and secondary animal products. In contrast, adult female individuals might have had a greater intake of harder and higher abrasive foods, such as processed agricultural produce. On the other hand, children and subadults likely had a harder diet than adults, possibly relying on less refined agricultural products and a lower intake of animal protein. This could be explained by their chewing capacity of healthy teeth and the lower level of teeth erosion at that age.
Data availability
Not applicable
Code availability
Not applicable
References
Aguelo J, Huertas J (2006) Memòria de la intervenció arqueológica al solar del Mercat de Santa Caterina, Ciutat Vella (Barcelona, Barcelonès). Unpublished. http://cartaarqueologica.bcn.cat/Docs/340/2015_03_05_11_04_17_Mem%C3%B2ria%20068-99.pdf
Aguelo J, Huertas J (2016) Les restes prehistòriques del subsòl del Mercat de Santa Caterina. In: Gómez A & Molist M (eds) La prehistòria al pla de Barcelona. Documents per a una nova síntesi. Ajuntament de Barcelona, Institut de Cultura, Museu d’Història de Barcelona, Barcelona, pp 182-183
Aguelo J, Huertas J, Puig F (2005) Santa Caterina de Barcelona: assaig d’ocupació i evolució. Quarhis: Quaderns d’Arqueologia i Història de la Ciutat de Barcelona 1:12-43
Alcalde G, Molist M, Montero I, Planagumà L, Saña M, Toledo A (1998) Producciones metalúrgicas en el nordeste de la Península Ibérica durante el III milenio cal. AC: el taller de la Bauma del Serrat del Pont (Tortellá, Girona). Trab Prehist 55(1):81–100. https://doi.org/10.3989/tp.1998.v55.i1.318
Alcàntara R (2018, unpublished). 4. Estudi Arqueozoològic. In: Estudi del jaciment arqueològi de la Plaá de la Gardunya (Fase III). Anàlisi de la formació dels dipòsits arqueològics. Coordination: Dr. Miquel Molist. Universitat Autònoma de Barcelona, 148-183.
Alrousan MF (2009) The Mesolithic-Neolithic transition in the Near East: biological implications of the shift in subsistency strategies through the analysis of dental morphology and dietary habits of human populations in the Mediterranean area 12.000-5.000 BP. Dissertation, University of Barcelona. https://portalrecerca.csuc.cat/74250011
Alrousan M (2016) Human dental buccal microwear and paleodiet reconstruction. Anthropologie (1962-) 54(3):305–315
Alrousan M, Al-Shorman A, Galbany J, Pérez-Pérez A (2013) Buccal dental microwear and stable isotopes of El Collado: a mesolithic site from Spain. Bull Int Assoc Paleodontol 7(1):4–11
Alrousan M, Estebaranz-Sánchez F, Al-Shorman A, Martínez LM, Gharaibeh N, Otum K, Pérez-Pérez A (2019) Buccal dental microwear as an indicator of dietary habits and dietary adaptation of the Byzantine people of Jordan. Anthropol Anz 76(4):353–362. https://doi.org/10.1127/anthranz/2019/0971
Balaguer P, Hinojo E, Oliart C, Soriano I (2011) Tecnología metalúrgica prehistórica en el nordeste de la Península Ibérica: el caso de las “hachas planas”. In: Mata-Perelló JM, Torró i Abat L, & M. Fuentes Prieto MN (eds) Actas del V Congreso Internacional sobre minería y metalurgia históricas en el suroeste europeo (León - 2008). Libro en homenaje a Claude Domergue. Sociedad Española para la Defensa del Patrimonio Geológico y Minero, pp 159-168.
Beynon AD (1987) Replication technique for studying microstructure in fossil enamel. Scanning Microsc 1(2):663–669
Bofill M (2016) Les indústries macrolítiques. In: Gómez A & Molist M (eds) La prehistòria al pla de Barcelona. Documents per a una nova síntesi. Ajuntament de Barcelona, Institut de Cultura, Museu d’Història de Barcelona, Barcelona, pp 117-123
Bordas A (2016) El jaciment de Nou de la Rambla 82. In: Gómez A & Molist M (eds) La prehistòria al pla de Barcelona. Documents per a una nova síntesi. Ajuntament de Barcelona, Institut de Cultura, Museu d’Història de Barcelona, Barcelona, pp 196-198
Brothwell DR (1981) Digging up bones. Pergamon Press, Oxford
Buikstra JE, & Ubelaker DH (1994) Standards for data collection from human skeletal remains. Arkansas archaeological survey research series 44:18
Cardoso HFV (2008) Epiphyseal union at the innominate and lower limb in a modern Portuguese skeletal sample, and age estimation in adolescent and young adult male and female skeletons. Am J Phys Antopol 135(2):161–170. https://doi.org/10.1002/ajpa.20717
Cardoso HFV (2008) Age estimation of adolescent and young adult male and female skeletons II, epiphyseal union at the upper limb and scapular girdle in a modern Portuguese skeletal sample. Am J Phys Antopol 137(1):97–105. https://doi.org/10.1002/ajpa.20850
Carlús X, González J (2008) Carrer de la Riereta, 37-37 bis: un nou assentament prehistòric al pla de Barcelona. Primers resultats. Cypsela: revista de prehistòria i protohistòria 17:91-114
Carlús X, González J, Nadal E. (2010) Estructures de l’Edat del Bronze tipus grill plan al litoral de Barcelona. Cypsela: revista de prehistòria i protohistòria 18:157-169
Chambon P (2016) Les practiques funéraires : évolution du rituel. In: Gómez A & Molist M (eds) La prehistòria al pla de Barcelona. Documents per a una nova síntesi. Ajuntament de Barcelona, Institut de Cultura, Museu d’Història de Barcelona, Barcelona, pp 141-150
Correia MA, Foley R, Mirazón Lahr M (2021) Applying dental microwear texture analysis to the living: challenges and prospects. Am J Phys Anthropol 174:542–554. https://doi.org/10.1002/ajpa.24133
Daura J, Sanz M, Soriano I, Pedro M, Rubio Á, Oliva M, Gibaja JF, Queralt I, Álvarez R, López-Cachero FJ (2017) Objetos de oro y epicampaniforme en la Cova del Gegant. Relaciones en la costa mediterránea de la Península Ibérica durante la Edad del Bronce. Trab Prehist 74(1):149–167. https://doi.org/10.3989/tp.2017.12188
Davis M, Pineda-Munoz S (2016) The temporal scale of diet and dietary proxies. Ecol Evol. 6(6):1883–1897. https://doi.org/10.1002/ece3.2054
DeSantis LRG, Haupt RJ (2014) Cougars’ key to survival through the Late Pleistocene extinction: insights from dental microwear texture analysis. Biol Lett 10(4):20140203. https://doi.org/10.1098/rsbl.2014.0203
Domonkošová Tibenská K, Bodoriková S, Katina S, Kovácsová V, Kubová J, Takács M (2010) Reconstruction of dietary habits on the basis of dental microwear and trace elements analysis of individuals from Gáň cemetery (District Galanta, Slovakia). Anthropol Anz 68(1):67–84
Espurz V, Pérez-Pérez A, Turbón D (2004) An approach to the study of post-depositional processes affecting inter-proximal wear facets and buccal enamel surfaces in hominid teeth. Anthropologie (1962-) 42(1):43–47
Estebaranz F, Galbany J, Martínez LM, Pérez-Pérez A (2007) 3-D interferometric microscopy applied to the study of buccal enamel microwear. In: Bailey SE, Hublin J-J (eds) Dental Perspectives on Human Evolution: State of the Art Research in Dental Paleoanthropology. Springer, Dordrecht, pp 391–403
Estebaranz F, Martínez LM, Galbany J, Turbón D, Pérez-Pérez A (2009) Testing hypotheses of dietary reconstruction from buccal dental microwear in Australopithecus afarensis. J Hum Evol. 57(6):739–750
Estebaranz F, Galbany J, Martínez LM, Turbón D, Pérez-Pérez A (2012) Buccal dental microwear analyses support greater specialization in consumption of hard foodstuffs for Australopithecus anamensis. J Anthropol Sci 90:1–24. https://doi.org/10.4436/jass.90006
Estebaranz F, Fernández E, Martínez L, Gamba C, Alrousan M, Turbón D, Arroyo-Pardo E, Oms JI, Pérez-Pérez A, Anfruns J (2008) Anàlisi antropològica de les restes neolítiques de la Caserna de Sant Pau (biometria, dentició, ADNA i microestriació dentària). Quarhis: Quaderns d’Arqueologia i Història de la Ciutat de Barcelona 4:76-82
Galbany J, Martínez LM, López-Amor HM, Espurz V, Hiraldo O, Romero A, De Juan J, Pérez-Pérez A (2005) Error rates in buccal-dental microwear quantification using scanning electron microscopy. Scanning 27:23–29. https://doi.org/10.1002/sca.4950270105
Galbany J, Garriga N, Majoral M, Coll R, Fluxà J (2008) Microdesgaste y patología dental en la población de la Edad de Bronce de «Mar i Muntanya» (Alella, Barcelona). Rev Esp Antrop Fís 28:25–36
García-González R, Carretero JM, Richards MP, Rodríguez L, Quam R (2015) Dietary inferences through dental microwear and isotope analyses of the Lower Magdalenian individual from El Mirón Cave (Cantabria, Spain). J Archaeol Sci 60:28–38. https://doi.org/10.1016/j.jas.2015.03.020
García-González R, Sánchez-Puente Z, Arsuaga JL, Carretero JM (2019) Dietary inferences from dental microwear patterns in Chalcolithic populations from the Iberian Peninsula: the case of El Portalón de Cueva Mayor (Sierra de Atapuerca, Burgos, Spain) and El Alto de la Huesera (Álava, Spain). Archaeol Anthropol Sci 11(8):3811–3823. https://doi.org/10.1007/s12520-018-0711-x
Gluitz A (2020) Análisis del patrón de microestriación dental: los hábitos alimenticios de las sociedades del llano de Barcelona en el Bronce Inicial. Master’s dissertation, Autonomous University of Barcelona
Gómez A, Nadal J, Molist M (2014) El mar i les comunitats humanes al llarg de la prehistòria al Pla de Barcelona. Barcelona Quaderns d’Història 21:63–76
Gómez A, Molist M (2016a) La seqüència temporal dels hàbitats. In: Gómez A & Molist M (eds) La prehistòria al pla de Barcelona. Documents per a una nova síntesi. Ajuntament de Barcelona, Institut de Cultura, Museu d’Història de Barcelona, Barcelona, pp 75-90.
Gómez A, Molist M (2016b) Les ocupacions del III i II mil·lennis: consolidació i desenvolupament del poblament. In: Gómez A & Molist M (eds) La prehistòria al pla de Barcelona. Documents per a una nova síntesi. Ajuntament de Barcelona, Institut de Cultura, Museu d’Història de Barcelona, Barcelona, pp 51-64
Gómez A, Bordas A, Arroyo S, Huertas J, Aguelo J, Velasco A, González J, Nadal J, Saña M, Molist M (2015) Evidències arqueològiques de les comunitats humanes en la transició del III al II mil·lenni cal BC al Pla de Barcelona. Quarhis: Quaderns d’Arqueologia i Història de la Ciutat de Barcelona 11:82-96
Granados O, Puig F, Farré R (1993) La intervenció arqueològica a Sant Pau del Camp: un nou jaciment prehistòric al Pla de Barcelona. Tribuna d’arqueologia 1991–1992:27–38
Grine F, Ungar PS, Teaford MF (2002) Error rates in dental microwear quantification using scanning electron microscopy. Scanning 24:144–153. https://doi.org/10.1002/sca.4950240307
Grine F, Ungar PS, Teaford MF (2006) Was the Early Pliocene hominin ‘Australopithecus’ anamensis a hard object feeder? S Afr J Sci 102(7):301–310
Grine F, Ungar PS, Teaford MF, El-Zaatari S (2006) Molar microwear in Preanthropus afarensis: evidence for dietary stasis through time and under diverse palaeoecological conditions. J Hum Evol 51(3):297–319. https://doi.org/10.1016/j.jhevol.2006.04.004
Hernando R, Lozano M, Ceperuelo D, Alonso S, Vaquero M (2019) Los niños de La Cova de Valdavara (Becerreá, Lugo): Análisis morfológico, patológico y del microdesgaste dental. Munibe 70:207–218. https://doi.org/10.21630/maa.2019.70.17
Hernando R, Willman JC, Vergès JM, Vaquero M, Alonso S, Oms X, Cebrià A, Morales JI, Lozano M (2020) Inferring childhood dietary maturation using buccal and occlusal deciduous molar microwear: a case study from the recent prehistory of the Iberian Peninsula. Archaeol Anthropol Sci 12(1):30. https://doi.org/10.1007/s12520-019-00997-z
Hernando R, Cano-Cano N, Martín P, Üstünkaya MC, Allué E, Lozano M (2022) Human diet at El Mirador Cave. In: Allué E, Martín P, Vergès JM (eds) Prehistoric herders and farmers. A transdisciplinary overwier to the archaeological record from El Mirador Cave. Springer, Cham, pp 295-326. https://doi.org/10.1007/978-3-031-12278-1_15
Hershkovitz I, Yakar R, Taitz C, Wish-Baratz S, Pinhasov A, Ring B (1993) The human remains from a Byzantine monastery at Khan el-Ahmar. Liber Annus 43:373–385
Jarošová I (2007) Dental buccal microwear of the medieval population from Dolní Věstonice, Czech Republic. Anthropologie (Brno) 45(1):71–80
Jarošová I (2008) Dietary inferences using buccal microwear analysis on the LBK population from Vedrovice, Czech Republich. Anthropologie (1962-) 46(2–3):101–110
Jarošová I, Tvrdý Z (2017) Diet and diversity of early farmers in Neolithic period (LBK): buccal dental microwear and stable isotopic analysis at Vedrovice (Czech Republic) and Nitra – Horné Krškany (Slovakia). Anthropologie (Brno) 55(3):353–384
Jarošová I, Pérez-Pérez A, Dočkalová M, Drozdová E, Turbón D (2006) Buccal dental microwear as a dietary indicator in the Iron Age human population from Son Real Spain. Anthropologie (1962-) 44(2):139–150
Jarošová I, Vávra J, Jiřík J, Horáková M (2016) Buccal dental microwear of a barbarian population from Prague-Zličín – a study of the Migration period in the Czech Republic. Interdiscip Archaeol 7(1):55–70. https://doi.org/10.24916/iansa.2016.1.3
King T, Andrews P, Boz B (1999) Effect of taphonomic processes on dental microwear. Am J Phys Anthropol 180:359–373
Kolář J, Jarošová I, Dreslerová G, Drozdová E, Dobisíková M (2012) Food strategies in Central Moravia (Czech Republic) during Final Eneolithic. A case study of Corded Ware culture communities. Archeologické Rozhl 64:237–264
Kouchi M, Mochimaru M, Tsuzuki K, Yokoi T (1999) Interobserver errors in anthopometry. J Human Ergol. 28(1–2):15–24
Lalueza C, Pérez-Pérez A (1989) Estudio nutricional de la población medieval de l’Esquerda (Osona, Barcelona): oligoelementos y estriación dentaria. Trab Antropología 21(3):2667–280
Lalueza C, Pérez-Pérez A, Turbón D (1996) Dietary inferences through buccal microwear analysis of middle and upper Pleistocene human fossils. Am J Anthropol 100(3):367–87. https://doi.org/10.1002/(SICI)1096-8644(199607)100:3%3c367::AID-AJPA5%3e3.0.CO;2-R
Louys J, Zaim Y, Rizal Y, Aswan, Puspaningrum M, Trihascaryo A, Price GJ, Petherick A, Scholtz E, DeSantis LRG (2021) Sumatran orangutan diets in the Late Pleistocene as inferred from dental microwear texture analysis. Quat Intern 603:74–81. https://doi.org/10.1016/j.quaint.2020.08.040
Lucas PW, Omar R, Al-Fadhalah K, Almusallam AS, Henry AG, Michael S, Thai LA, Watzke J, Strait DS, Atkins AG (2013) Mechanisms and causes of wear in tooth enamel: implications for hominin diets. J R Soc Interface 10:20120923. https://doi.org/10.1098/rsif.2012.0923
Mahoney P (2006) Dental microwear from natufian hunter-gatherers and early Neolithic farmers: comparisons within and between samples. Am J Phys Anthropol 130:308–319
Mainland I (2003) Dental microwear in grazing and browsing Gotland sheep (Ovis aries) and its implications for dietary reconstruction. J Archaeol Sci 30(11):1513–1527
Malgosa A (2016) La població: característiques físiques i estat de salut. In: Gómez A & Molist M (eds) La prehistòria al pla de Barcelona. Documents per a una nova síntesi. Ajuntament de Barcelona, Institut de Cultura, Museu d’Història de Barcelona, Barcelona, pp 151-156.
Martín A, Guilaine J, Thommeret J, Thommeret Y (1981) Estratigrafía y dataciones C14 del yacimiento de la «Cova del Frare» de St. Llorenç del Munt (Matadepera, Barcelona). ZEPHYRVS 32–33:101–111
Martínez LM, Pérez-Pérez A (2004) Post-mortem wear as indicator of taphonomic processes affecting enamel surfaces of hominin teeth from Laetoli and Olduvai (Tanzania): implications to dietary interpretations. Anthropologie (1962-) 42(1):37–42
Martínez LM, Galbany J, Pérez-Pérez A (2004) Palaeodemography and dental microwear of homo habilis from East Africa. Anthropologie (1962-) 42(1):53–58
Martínez LM, Estebaranz-Sánchez F, Galbany J, Pérez-Pérez A (2016) Testing dietary hypotheses of East African mominines using buccal dental microwear data. PLoS ONE 11(11):e0165447. https://doi.org/10.1371/journal.pone.0165447
Martínez LM, Estebaranz-Sánchez F, Ferràndez-Cañadell C, Romero A, Ribot F, Galbany J, Gibert L, Pérez-Pérez A (2020) Buccal dental-microwear and feeding ecology of Early Pleistocene Theropithecus oswaldi from Cueva Victoria (Spain). J Hum Evol 142:102736. https://doi.org/10.1016/j.jhevol.2019.102736
Martínez LM, Estebaranz-Sánchez F (2018, unpublished). 1. Estudi Antropològic de les restes esquelètiques de la Gardunya. In: Estudi del jaciment arqueològi de la Plaá de la Gardunya (Fase III). Anàlisi de la formació dels dipòsits arqueològics. Coordination: Dr. Miquel Molist. Universitat Autònoma de Barcelona, 3-113
Martínez LM, Estebaranz-Sánchez F, Pérez-Pérez A (2015) Buccal dental microwear as an indicator of diet in modern and ancient human populations. In: Hardy J, Kubiak-Martens L (eds) Wild harvest: plants in the hominin and pre-argarian human worlds. Oxbow Books, United Kingdom, pp 155-170. https://doi.org/10.2307/j.ctvh1dmjj
Maya JL (1997) Reflexiones sobre el Bronce Inicial en Cataluña.SAGVNTVM. Papeles del Laboratorio de Arqueología de Valencia 30:11-27. https://doi.org/10.7203/SAGVNTVM..2140
Maya JL, Petit MÀ (1986) El Grupo del Nordeste. Un nuevo conjunto de cerámicas con boquique en la Península Ibérica. An Prehist Arqueol 2:49–71
Mihlbachler MC, Beatty BL, Caldera-Siu A, Chan D, Lee R (2012) Error rates and observer bias in dental microwear analsis using light microscopy. Palaeontol Electron 15(1):1–22
Nadal E, Castillo RJ (2009) Intervenció arqueològica al C/ d’Espalter, 1b-11; C/ de Sant Pau, 66; C/ de Sant Josep Oriol, 7-11; pl. de Salvador Seguí. https://arxiu.arqueologiabarcelona.bcn.cat/intervencio-arqueologica-als-carrers-espalter-1b-11-carrer-de-sant-pau-66-carrer-sant-josep-oriol-7-11-placa-salvador-segui-de-setembre-de-2007-a-maig-de-2009
Ortner DJ (2003) Identification of pathological conditions in human skeletal remains. Academic Press, San Diego
Pérez-Pérez A (2004) Why buccal microwear? Anthropologie (1962-) 42(1):1–4
Pérez-Pérez A, Alesan A, Roca L (1990) Measurement error: inter-and intraobserver variability. An empiric study. Int J Anthropol 5(2):129–135. https://doi.org/10.1007/BF02442082
Pérez-Pérez A, Lalueza C, Turbón D (1994) Intraindividual and intragroup variability of buccal tooth striation pattern. Am J Phys Anthropol 94(2):175–187. https://doi.org/10.1002/ajpa.1330940203
Pérez-Pérez A, Bermúdez de Castro JM, Arsuaga JL (1999) Nonocclusal dental microwear analysis of 300.00-year-old Homo heilderbergensis teeth from Sima de los Huesos (Sierra de Atapuerca, Spain). Am J Phys Anthropol. 108(4):433–457. https://doi.org/10.1002/(SICI)1096-8644(199904)108:4%3C433::AID-AJPA5%3E3.0.CO;2-5
Pérez-Pérez A, Espurz V, Bermúdez de Castro JM, de Lumley MA, Turbón D (2003) Non-occlusal dental microwear variability in a sample of Middle and Late Pleistocene human populations from Europe and the Near East. J Hum Evol. 44(4):497–513. https://doi.org/10.1016/S0047-2484(03)00030-7
Pérez-Pérez A, Lozano M, Romero A, Martínez LM, Galbany J, Pinilla B, Estebaranz-Sánchez F, Bermúdez de Castro JM, Carbonell E, Arsuaga JL (2017) The diet of the first Europeans from Atapuerca. Sci Rep 7:43319. https://doi.org/10.1038/srep43319
Pérez-Pérez A, Martínez LM, Gómez M, Estebaranz F, Romero A (2018) Correlations among dietary proxies in African fossil hominins: dental buccal microwear, occlusal textures and 13C stable isotope. J Archaeol Sci: Rep 22:384–391. https://doi.org/10.1016/j.jasrep.2018.03.013
Piera J, Moreno I, Bordas A, Suau L (2005) Memòria de la intervenció arqueològica a l’Illa d’en Robador: C/Robador, Sant Rafel, Sant Josep Oriol i Sadurní. Ciutat Vella, Barcelona. http://cartaarqueologica.bcn.cat/Docs/19/2010_12_03_18_16_38_Mem%C3%B2ria%20125-04.pdf
Pinilla Pérez B, Romero A, Pérez-Pérez A (2011) Age-related variability in buccal dental-microwear in Middle and Upper Pleistocene human populations. Anthropol Rev 74:25–37. https://doi.org/10.2478/v10044-010-0005-0
Polo-Cerdá M, Romero A, Casabó J, De Juan J (2007) The Bronze Age burials from Cova Dels Blaus (Vall d′Uixó, Castelló, Spain): an approach to palaeodietary reconstruction through dental pathology, occlusal wear and buccal microwear patterns. HOMO- J Comp Hum Biol 58(4):297–307. https://doi.org/10.1016/j.jchb.2006.10.005
Puech PF, Prone A, Kraatz R (1980) Microscopie de l’usure dentaire chez l’Homme fossile : bol alimentaire et environnement. C R Acad Sc Paris 290:1413–1416
Radini A, Nikita E, Buckley S, Copeland L, Hardy K (2017) Beyond food: the multiple pathways for inclusion of materials into ancient dental calculus. Am J Phys Anthropol 162:71–83. https://doi.org/10.1002/ajpa.23147
Rivals F, Semprebon GM (2010) What can incisor microwear reveal about the diet of ungulates? Mammalia 74:401–406. https://doi.org/10.1515/MAMM.2010.044
Romero A, de Juan J (2007) Intra- and interpopulation human buccal tooth surface microwear analysis: inferences about diet and formation processes. Anthropologie (1962-) 45(1):61–70
Romero A, Martínez-Ruiz N, de Juan J (2004) Non-occlusal dental microwear in a Bronze-Age human sample from East Spain. Anthropologie (1962-) 42(1):65–70
Romero A, Galbany J, Martínez-Ruiz N, de Juan J (2009) In vivo turnover rates in human buccal dental-microwear. Am J Phys Anthropol 48:223–224
Romero A, Galbany J, de Juan J, Pérez-Pérez A (2012) Brief communication: short- and long-term in vivo human buccal-dental microwear turnover. Am J Phys Anthropol 148(3):467–472. https://doi.org/10.1002/ajpa.22054
Romero A, Ramírez-Rozzi FV, de Juan J, Pérez-Pérez A (2013) Diet-related buccal dental microwear patterns in Central African Pygmy foragers and Bantu-speaking farmer and pastoralist populations. PLoS ONE 8(12):e84804. https://doi.org/10.1371/journal.pone.0084804
Rovira MC (2006) El Bronze Inicial a Catalunya des de la perspectiva metal·lúrgica. Cypsela: revista de prehistòria i protohistòria 16:135-145
Salazar-García DC, Romero A, García-Norja P, Subirà ME, Richards MP (2016) A combined dietary approach using isotope and dental buccal-microwear analysis of human remains from the Neolithic, Roman and Medieval periods from the archaeological site of Tossal de les Basses (Alicante, Spain). J Archaeol Sci: Rep 6:610–619. https://doi.org/10.1016/j.jasrep.2016.03.002
Shapland F, Lewis ME (2013) Brief communication: a proposed osteological method for the estimation of pubertal stage in human skeletal remains. Am J Phys Anthropol 151(2):302–310. https://doi.org/10.1002/ajpa.22268
Soriano I (2013) Metalurgia y Sociedad en el Nordeste de la Península Ibérica: (finales del IV – II milenio cal ANE). University of Michigan Press, Ann Arbor. https://doi.org/10.30861/9781407311180
Soriano I (2018) Les pràctiques funeràries durant el calcolític i el bronze antic i mitjà. La fi és el principi. Pràctiques funeràries a la Catalunya prehistòrica. Generalitat de Catalunya, Barcelona, pp 79–102
Soriano I, Chamón J (2012) Estudio arqueometalúrgico del punzón de Cova de la Pesseta (Torrelles de Foix, Barcelona). Datos composicionales, metalográficos, isotópicos y funcionales MARQ. Arqueol Museos 5:73–89
Suau L, Matas Ò (2002) Memòria de la intervenció arqueològica de la zona C del projecte d’infraestructures troncals des del carrer Carders a la central de RSU, en el mercat de Santa Caterina, al districte de Ciutat Vella, de Barcelona. https://arxiu.arqueologiabarcelona.bcn.cat/memoria-de-la-intervencio-arqueologica-del-projecte-dinfrastructures-sic-troncals-des-del-carrer-carders-a-la-central-de-rsu-en-el-mercat-de-santa-caterina-al-districte-de-ciutat-vella-zona-d-desembre-2002-abril-2003;isad?sf_culture=en
Teaford MF (1988) Scanning electron microscope diagnosis of wear patterns versus artifacts on fossil teeth. Scanning Microsc 2(2):1167–1175
Teaford MF, Glander KE (1991) Dental microwear in live, wild-trapped Alouatta palliata from Costa Rica. Am JPhys Anthropol 85(3):313–319. https://doi.org/10.1002/ajpa.1330850310
Teaford MF, Oyen OJ (1989) Live primates and dental replication: new problems and new techniques. Am J Phys Anthropol 80(1):73–81. https://doi.org/10.1002/ajpa.1330800109
Ungar PS, Grine FE, Teaford MF (2008) Dental microwear and diet of the Plio-Pleistocene hominin Paranthropus boisei. PLoS ONE 3(4):e2044. https://doi.org/10.1371/annotation/195120f0-18ee-4730-9bd6-0d6effd68fcf
Ungar PS, Abella EF, Burgman JHE, Lazagabaster IA, Scott JR, Delezene LK, Manthi FK, Plavcan JM, Ward CV (2020) Dental microwear and Pliocene paleocommunity ecology of bovids, primates, rodents and suids at Kanapoi. J Hum Evol 140:102315. https://doi.org/10.1016/j.jhevol.2017.03.005
Velasco A, Terrats N, Gómez A, Molist M (2017) L’assentament de la Plaça de la Gardunya a Inicis del II mil·lenni: noves dades sobre les ocupacions de l’Edat del Bronze Inicial al pla de Barcelona. Quarhis: Quaderns d’Arqueologia i Història de la Ciutat de Barcelona 13:70-89
Weber K, Winkler DE, Kaiser TM, Žigaitė Ž, Tütken T (2021) Dental microwear texture analysis on extant and extinct sharks: ante- or post-mortem tooth wear? Palaeogeogr Palaeoclimatol Palaeoecol 562:110147. https://doi.org/10.1016/j.palaeo.2020.110147
Weber K, Winkler DE, Schulz-Kornas E, Kaiser TM, Tütken T (2021) The good, the bad and the ugly – a visual guide for common post-mortem wear patterns in vertebrate teeth. Palaeogeogr Palaeoclimatol Palaeoecol 578:110577. https://doi.org/10.1016/j.palaeo.2021.110
Acknowledgements
Special thanks are extended to Núria Miró, Emili Revilla and Carme Miró for their assistance and granting access the osteological collections of the Museu d’Història de Barcelona (MUHBA). Gratitude is also owed to Meritxell Vendrell Flotats from the Servei de Microscòpia of the Universitat Autònoma de Barcelona for her help in sample preparation and obtaining SEM images. Additionally, we are grateful to Dr. Alejandro Pérez-Pérez from the University of Barcelona (UB) for facilitating the use of his laboratory facilities to obtain the positive results of the analysed teeth. Moreover, we sincerely appreciate the valuable comments and suggestions provided by the anonymous reviewer, which significantly contributed to the enhancement of this manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This research has been funded by the project Prehistòria al pla de Barcelona, cooperated by the ICUB Servei d’Arqueologia, the Museu d’Història de Barcelona and the Prehistory Department of the Autonomous University of Barcelona. This study has also been supported by the Spanish Ministerio de Ciencia e Innovacion (project numberd PID2019-106399GB-I00) and funded by PID2020-112963GB-I00 by MNCIN/AEI/10.13039/501100011033 and by “ERDF A way of making Europe” by the “European Union”) and finally the GRAMPO research group (2017SGR-1302). Alicia Gluitz has been granted with a FPU research scholarship from the Spanish Ministerio de Universidades (FPU20/03612), a support that has enabled the successful completion of this study.
Author information
Authors and Affiliations
Contributions
Not applicable
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gluitz, A., Martínez, L.M., Molist, M. et al. Diet in the Early Bronze Age: a buccal microwear analysis from the plain of Barcelona (Spain). Archaeol Anthropol Sci 15, 157 (2023). https://doi.org/10.1007/s12520-023-01855-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-023-01855-9