
Abstract
This study provides a taphonomic and paleoenvironmental analysis of the micromammal (˂1 kg) accumulations from the archaeological site Álvarez 4 (AZ4), a rock shelter located at the middle basin of the Limay River, Río Negro Province, northwestern Patagonia, Argentina. Based on taphonomic and taxonomic features, several skeletal remains accumulated in Period I (963 cal. BP), Period II (187 cal. BP), and square C3 (963 cal. BP) are assigned to the action of Strigiformes, while pellets found in the surface layer (SL) (187 cal. BP) would have been deposited by Accipitriformes. Pellet bones were more protected from trampling than bones recovered from the sieved sediments of the SL. In addition, some Caviidae remains from Period II and square C3 were likely incorporated in human consumption. This low-ranked resource might have complemented the otherwise high-ranked animals found in Álvarez 4, which would suggest a similar diet breadth to that frequently seen at several archaeological sites in the area. The paleoenvironmental analysis based on the small mammals recovered from AZ4 shows the development of open areas of shrubby-grassy Patagonian steppe and large rocky outcrops with overall stability of habitats, with a dominance of shrublands and steppes in the last ~1000 years. Past environments were more heterogeneous than today, as evidenced by the absence of some species and the expansion of opportunistic species in the modern pellet samples near to the rock shelter.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Most of the Quaternary small mammal remains from the Limay River, Río Negro Province, northwestern Patagonia come from archaeological sites (e.g., Andrade and Boschín 2015; Ceballos 1982; Crivelli Montero et al. 1996; Fernández et al. 2016, 2018; Guillermo et al. 2020a, b; Hajduk and Albornoz 1999; Massoia 1982, 1987; Pardiñas 1999a, b; Pardiñas and Teta 2013; Pearson 1987; Pearson and Pearson 1993; Tammone et al. 2014, 2020; Teta et al. 2005). The oldest records of micromammals from this area were found at basal levels (sterile from the archaeological point of view) dated toward the end of the Late Pleistocene, during the Last Glacial Maximum (Tammone et al. 2014). However, most findings of micromammals are associated with the Holocene, especially toward its end (Crivelli Montero et al. 1996; Fernández et al. 2016; Pardiñas 1999a; Pardiñas and Teta 2013; Pearson 1987; Pearson and Pearson 1993; Teta et al. 2005). This temporal distribution of the micromammal findings is coincident with the peopling of the area. The oldest dating associated with humans corresponds to the Pleistocene-Holocene transition. The human populations increase up to ca. 6200 14C BP; then, they decrease sharply between ca. 5000 and 3000 14C BP, and finally they grow steadily since 3000 14C BP (e.g., Boschín 2009; Crivelli Montero 2010; Crivelli Montero et al. 1993, 1996; Hajduk et al. 2006, 2007).
The taphonomic studies on fossil assemblages of micromammals in this area stand out for the predatory activity of two types of accumulating agents: Strigiformes birds (Guillermo et al. 2020b; Tammone et al. 2014) and human hunter-gatherers (Andrade and Boschín 2015; Bond et al. 1981; Ceballos 1982, 1987; Fernández et al. 2017a; Fernández 1988-1990; Hajduk and Albornoz 1999; Lezcano et al. 2010; Pardiñas and Teta 2013; Tammone 2017), or a combination of both (Andrade 2015; Crivelli Montero et al. 2017; Fernández et al. 2016; Guillermo et al. 2020a; Pardiñas 1999a, b; Pardiñas and Teta 2013; Silveira and Cordero 2014; Tammone et al. 2020; Teta et al. 2005). However, the present study on the micromammal assemblage from the archaeological site Álvarez 4 (AZ4) reveals a different type of small-mammal accumulation that had not been reported before for northwestern Patagonia. This kind of deposit is characterized by various sequential and temporal deposition episodes yielded by humans and two different non-human agents and has been defined as a palimpsest by different authors in other sites (Bailey 1981, 2007; Binford 1981; Malinsky-Buller et al. 2011).
Micromammal assemblages of the upper and middle basin of the Limay River have shown taxonomic stability, or minor variations, during the Holocene (Crivelli Montero et al. 1996; Fernández et al. 2016, 2018; Pardiñas 1999a; Pardiñas and Teta 2013; Pearson 1987; Pearson and Pearson 1993; Tammone et al. 2020; Teta et al. 2005). However, since the Post-Hispanic period, a gradual entry of domestic exotic fauna (horse, cow, and sheep) to northwestern Patagonia took place (see more details on entry of each species in Menéndez 1896: 218; Rosales 1877; Viedma 1938: 1-26; Vignati 1964: 497), giving rise to a series of environmental modifications, mainly in the soil and vegetation cover, by the action of trampling and overgrazing (e.g., Bertiller and Bisigato 1998). This also led to a restructuring of the small-mammal communities in the area, with the prevalence of generalist or opportunistic species, such as Abrothrix olivacea, Calomys musculinus, Eligmodontia sp., and Oligoryzomys longicaudatus (Crivelli Montero et al. 1996; Fernández et al. 2016; Pardiñas 1999a; Pardiñas and Teta 2013; Pearson 1987; Pearson and Pearson 1993; Tammone et al. 2020; Teta et al. 2005).
The archaeological site Álvarez 4, located in the middle basin of the Limay River, offers an archaeofaunal assemblage from the latest Holocene. It stands out by the presence of mollusks, birds, and mammal remains of different body masses, including micromammals, in addition to pellets preserved in stratigraphy. This study is focused on the taphonomic and taxonomic assessment of the micromammal assemblage collected from AZ4, in order to identify the main agents that account for the bone accumulations, both natural and anthropic. In addition, this study contributes to the knowledge of the paleoenvironmental conditions of the area at the time of the deposition of the remains, as well as to the information on the use of faunal resources by the human groups that inhabited the middle basin of the Limay River during the end of the Late Holocene.
Study area and archaeological context
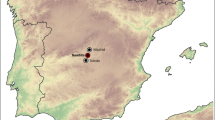
The archaeological site AZ4 is a rock shelter located in the Department of Pilcaniyeu, Province of Río Negro, northwestern Patagonia (40°48′38.8″ S, 70°27′12.9″ W, 917 m asl; Fig. 1a). It is about 300 m west of the Coquelén creek, in a tuff outcrop at 910 m above sea level (Crivelli Montero and Palacios 2010). The Coquelén creek is an affluent of the left bank of the Comallo creek, tributary of the Limay River (Fig. 1b). The valley was carved in granitic rocks of the Mamil Choique Formation that are widely exposed in the area, and in the younger tuffs of the Collón Curá Formation, assigned to the Middle Miocene (Rodríguez et al. 2007). About 200 m from AZ4, there is a small weed/turf habitat (mallín) generally dry, but through which sometimes water flows, as suggested by a narrow runoff channel (Crivelli Montero and Palacios 2010).

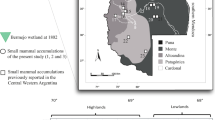
a Map of the study area, northwest Patagonia, Argentina, superimposed on the main phytogeographic districts of the study area (cf. Oyarzabal et al. 2018), including the location of the archaeological site Álvarez 4 (white triangle), other archaeological sites (black triangles) mentioned in this work (LL: Epullán Grande; CLC1: Cañadón Las Coloradas 1; Mar 1: La Marcelina 1; CPL: Cueva y Paredón Loncomán; CPO: Casa de Piedra de Ortega; Sarita I, II, IV caves), and the pellet samples (circles) analyzed in this study: (1) Pampa de Nestares; (2) Barda Esteban; (3) Cañadón Las Coloradas; (4) Cerro Castillo; (5) Pampa de los Guanacos; (6) Cañadón Fuquelén; (7) Corralito; (8) Canteras Comallo; (9) Pipilcura; (10) Paso de los Molles; (11) Cooperativa Escuela; (12) ONO Comallo; (13) Estancia Picañeu; (14) Paraje Leleque. b Detail of the Álvarez 4, Álvarez 3, and Sauce Guacho location based on a satellite image. White arrows indicate the tuffs of the Collón Curá Formation. c Inside rock shelter before excavation (picture taken by Eduardo Crivelli Montero). d Excavation plan of Álvarez 4 (modified from Crivelli Montero and Palacios 2010: Fig. 4). e Schematic profile of Álvarez 4
The average annual rainfall in the area is 220 mm, reaching its maximum in autumn and winter, and a maximum water deficit in summer. The mean annual temperature is 7°C and the annual temperature is 16°C (Paruelo et al. 1998). Concerning phytogeography, AZ4 is located within the Occidental District of the Patagonian Province, close to the Central District, characterized by a shrub-gramineous steppe (Oyarzabal et al. 2018; Fig. 1a). Grasses such as Pappostipa speciosa, P. humilis, Poa ligularis, and P. lanuginosa and the shrubs Adesmia volckmannii, Berberis microphylla, Mulinum spinosum, Senecio filaginoides, and Ephedra frustillata are abundant (Oyarzabal et al. 2018).
The surface of the rock shelter AZ4 is 3.5 × 3.5 m (Fig. 1c, d). Its mouth, oriented in a NW direction, offers little shelter to the prevailing winds from the west or westerlies. The excavation was carried out in 2007, under the direction of Dr. Eduardo Crivelli Montero. Three squares of 1 m2 and two of 0.50 m2 were excavated (Fig. 1d) following visible “natural” strata, when possible. It was also excavated in artificial levels 5 to 10 cm thick; the stratigraphic thickness reached 1.25 m at its contact with the bed-rock (Crivelli Montero and Palacios 2010; Fig. 1e).
The surface of AZ4 was covered by a layer of sheep guano, variable in thickness depending on the square (Fig. 1e), together with few archaeological and modern material. This section was called surface layer (SL), which extends from conventional level 0 to 35–40 cm in depth in squares B3–B4, C2, and C4, while in C3 it reaches 50 cm. In almost the entire square C3 and part of the SL of square C2, there is a disturbed sedimentary fill and evidence of recent looting activity. Other sectors show no clear evidences of alteration (Fig. 1d and e). An accumulation of pellets was found at the base of the SL in square B3, between 40 and 45 cm deep, and dated older than 210 ± 90 14C BP (probabilistic \(\overline{x}\) 187 cal. BP) (Fig. 1e). Below SL, the sediment is homogeneous, with a gravitational matrix, and no occupation events are recognized. The sequence was divided into two temporal sections according to depth and radiocarbon dating: Period I (PI) dated to 1100 ± 70 14C BP (probabilistic \(\overline{x}\) 963 cal. BP) and Period II (PII) dated to 210 ± 90 14C BP (probabilistic \(\overline{x}\) 187 cal. BP). Square C3, with maximum chronologies (963 cal. BP), was studied separately due to evidence of recent human disturbance.
In AZ4, various types of lithic projectile points made of dacite and silex were found, as well as two plate fragments engraved with carved signs and ceramic fragments (Crivelli Montero and Palacios 2010). Additionally, the zooarchaeological and taphonomic studies of the mollusks, birds, and medium to large mammals (more than 900 specimens) have revealed that the subsistence of the human groups that inhabited AZ4 was based on the exploitation of the guanaco (Lama guanicoe) and was complemented with smaller mammals such as Molina’s Hog-nosed Skunk (Conepatus chinga), the large hairy armadillo (Chaetophractus villosus), the lesser rhea (Rhea pennata), the Pampas gray fox (Lycalopex gymnocercus), the pichi (Zaedyus pichiy), and the southern viscacha (Lagidium viscacia) (Guillermo and Fernández 2021). Two exotic species were also identified—the sheep (Ovis orientalis aries) and the European hare (Lepus europaeus)—although no taphonomic evidence suggesting their incorporation into human subsistence for historical context was found (Guillermo and Fernández 2021). The scarcity of archaeological finds and the low anthropic alteration of the sediments preliminarily indicated that the site had brief and not very intensive human occupations (Crivelli Montero and Palacios 2010). However, in the vicinity of AZ4, there are two archaeological sites (Fig. 1b), one located at the base of the cliff (Álvarez 3) and a large open-air site (Sauce Guacho), where residential activities were carried out (Crivelli Montero and Palacios 2010).
Material and methods
All the recovered small mammal remains were observed with a 10× hand lens and under a Leica A60 binocular magnifying glass, with a zoom of up to 40×, when necessary. Pellets were photographed and measured with a digital caliper before their disaggregation and cleaning. Afterwards, they were immersed in a container with warm water for 5 s, and then, the bone remains were separated with surgical instruments. In this work, 400 specimens were analyzed, 263 were scattered in the stratigraphy, and 137 were contained in pellets recovered from SL.
Anatomical identification and quantification were based on reference collections from the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (MACN, Ciudad Autónoma de Buenos Aires) and osteological atlas (Pearson 1995; Udrizar Sauthier et al. 2020). Body mass classification of the mammals herein considered includes small mammals below 1 kg (Didelphidae, Ctenomyidae, Caviidae, and Cricetidae) and mesomammals ranging from 1 to 15 kg (Mephitidae and unidentified medium mammals recovered from pellet samples). The average body mass of the species represented was obtained from the Macroecological Database of Mammalian Body Mass (MOM), version 4.1 (Smith et al. 2003). The percentage of biomass contributed by each prey species for each archaeological and recent pellet sample was estimated as (Bi) = [(Spi Ni) / Σ(Spi Ni)] × 100, where Spi is the weight of species i and Ni is the number of consumed individuals of species i (Marti et al. 2007).
The taphonomic analysis followed the methodology proposed by Andrews (1990), Fernández-Jalvo and Andrews (1992), and Fernández et al. (2017b). The minimum number of specimens (NISP), minimum number of skeletal elements (MNE), and minimum number of individuals (MNI) were calculated following Badgley (1986).
Assessment of the relative abundance of skeletal elements considers the representation of each element in the context of the MNI in the sample, MNEi / Ei × MNI) × 100, where MNEi is the minimum number of a given skeletal element in the sample, and Ei is the expected number of this skeletal element in one individual (Andrews 1990). Two indexes of proportion of elements were evaluated using MNE: relation between cranial and postcranial elements (femora + humeri) / (mandibles + maxillae) × 100, and relation between proximal and distal limb elements (tibiae + ulnae) / (femora + humeri) × 100.
Breakage of skull and mandibles was evaluated considering the methodology proposed by Montalvo et al. (2020), and breakage of postcranial elements following Andrews (1990).
The categories of digestive corrosion on teeth were also evaluated according to Fernández et al. (2017b), who distinguished digestive corrosion patterns on different tooth morphologies of South American rodents and marsupials. Digestive corrosion on the postcranial elements was evaluated using proximal portions of femora and distal portions of humeri (Andrews 1990; Marín-Monfort et al. 2019).
Anthropic exploitation of micromammals was evaluated according to the taphonomic attributes of cutting marks and thermoalterations, and taxonomical concepts that include abundance of large species and/or of diurnal and social habits (Pardiñas 1999b; Fernández et al. 2017a, and references therein). Thermoalteration was evaluated distinguishing between partial (mainly on the distal ends of premaxillaries and incisors, and zeugopodial bones) and complete (e.g., Medina et al. 2012; Pardiñas 1999b), and through a chromatic scale: white-cream = unburnt bones, yellow-reddish-brown-black = burnt bones, and blue-grayish = calcined bones (Shipman et al. 1984). Both attributes help to differentiate between natural/human-indirect fires (generally burnt or calcined bones on the whole surface, respectively) and human cooking from exposing small mammals directly to charcoal fire (López and Chiavazza 2020, and references therein).
Paleoenvironmental analyses are usually based on presence/absence of some species and relative changes of their frequencies of MNI. Most small mammals have relatively narrow environmental requirements, being frequently associated with particular habitats. With adequate taphonomic control, temporal changes in the abundance and distribution of small-mammal taxa can reveal the environmental conditions at the time of deposition of the assemblages (e.g., Andrews 1990; Pardiñas 1999a; Stahl 1996).
Fourteen samples of micromammals from pellets of avian raptors (Tyto furcata, Bubo magellanicus (Strigiformes), and Geranoaetus melanoleucus (Accipitriformes)) were used as actualistic parameters of the zooarchaeological materials. These samples were taken within a 40 km radius from the archaeological site and in the main environmental units of the study area (Fig. 1a; Table S1).
Taking into account the variation in sample size of the different archaeological and recent pellet samples, rarefaction curves were calculated for each sample in order to assess reliability of richness (S) and relative abundance of species in terms of MNI. A box plot was used to graph the median and interquartile range of the measurements of each pellet recovered from SL and pellet samples of different avian raptors reported by Trejo and Ojeda (2002: Table 1) for the study area. The Shannon diversity index (H) was estimated for each sample based on MNI. A correspondence analysis was conducted in order to explore species and sample ordination in a multivariate space. This exploration was made on a data matrix of relative abundance (MNI) log-transformed (ln). Statistical analyses were performed using the program PAST (PAleontological STatistics), version 4.03 (Hammer 1999-2020).
Finally, a taxonomic habitat index (THI) was calculated for archaeological and selected recent pellet samples. The THI was employed following the proposal of Evans et al. (1981), slightly modified by Pardiñas (1999a), as follows: THI = Σ ([pi. MNIi] / MNI) / S, where pi = proportion of the species i in a landscape unit; MNIi = MNI for the i species; MNI = MNI for the total sample; and S = total number of species in the sample. This index was applied to show the overall representation of each type of habitat (i.e., forest, shrubland, steppe, grassland, weed/turf, rocky, and bare). The probability of occurrence of a species in each habitat, expressed as a proportion, derives from the trapping data provided by Pearson (1995: Table 2) for northwestern Patagonia, although, due to the lack of data for some species, other more general bibliographic sources were used (e.g., Patton et al. 2015; SAREM 2019). This index is a good complement to those aforementioned methodologies to infer environmental evolution on a large scale (e.g., Pardiñas 1999a).
Results
Pellet description
Seven pellets in good state of preservation (Fig. 2a–g) and two incomplete ones (Fig. 2h, i) were recovered from the base of SL in B3–B4 squares. Measurements of each specimen are detailed in Table 1. Mean and standard deviation of each variable of complete pellets are as follows: length \(\overline{x}\) = 4.7 (σ = 1.8); width \(\overline{x}\) = 2.6 (σ = 0.4); thickness \(\overline{x}\) = 2.4 (σ = 0.2).

Pellets recovered from the surface layer of the archaeological site Álvarez 4. a–g Complete pellets. h, i Incomplete pellets. Scales = 1 cm
Anatomical representation
The total MNE is 368, of which 134 were recovered from pellets, so the highest MNE comes from the SL, and 128 correspond to cranial elements (Table 2). It should be noted that, in addition, 26 bone remains (not included in Table 2) were recovered with evidence of thermoalteration.
Calculated indexes are shown in Table 3. In all archaeological units, crania (including teeth) are the most frequent skeletal elements. For limb bones, tibiae and humeri are the most frequent in PI and SL, and femora and tibiae predominate in PII and C3 (Table 3).
The curves derived from the analysis of relative abundance of skeletal elements are shown in Fig. 3a. The average of relative abundance is low in the PI (16.9%), PII (12.7%), C3 (12.8%), and SL (20.2%) samples. In PI and, to a lesser degree, in PII, the maxillae, mandibles, and isolated incisors reveal similar percentages, followed by tibiae with slightly lower values. C3 results show very low percentages for postcranial elements and only slightly higher for the cranial elements. SL is the level with the most complete representation, with high frequency values for cranial elements, tibiae, and, to a lesser extent, humeri.

a Relative abundance of skeletal elements (based on the MNI of each unit) from archaeological samples of Álvarez 4. b Relative abundance of skeletal elements from Periods I (PI) and II (PII), and square C3, compared with small-mammal assemblages generated by average of Athene cunicularia (Montalvo et al. 2020), Bubo magellanicus (Montalvo et al. 2016), and Tyto alba (Andrews 1990). c Relative abundance of skeletal elements from the surface layer (SL), compared with small-mammal assemblages produced by average of Caracara plancus, Geranoaetus melanoleucus (two samples described by López et al. 2017), and G. polyosoma (Montalvo and Tallade 2009)
Breakage degree
Categories of breakage of skulls, mandibles, and long bones are detailed in Table 4 and Fig. 4.

Taphonomic examples from the archaeological site Álvarez 4. a Femur of a mesomammal, with extreme digestion on the distal epiphysis (surface layer, pellet H). b Mandibular fragment of Caviinae with heavy digestion (surface layer). c Skull fragment of Ctenomys cf. C. haigi with light digestion on incisors (surface layer). d Femur of Sigmodontinae, with extreme digestion on the proximal epiphysis (surface layer). e Tibia of Mammalia indet., with heavy corrosion on proximal epiphysis (surface layer). f Mandible of Phyllotis xanthopygus, with heavy digestion on teeth (surface layer). g Tibia of Caviinae with burned distal end (Period II). Scales = 1 cm
Regarding the completeness of skulls, category 4 is represented in all levels in high frequencies (Table 4). Complete skulls were recovered only in SL (particularly from pellets), but also very broken skulls (categories 7 and 8) were found in the sediment without pellet remains (pellets would protect the skulls). In PI and PII, skulls of categories 5 and 7 are also recorded; in PII and C3, remains are less modified (categories 2 and 3). Evaluation of mandible breakage reveals that the most frequent specimens belong to categories 2 and 3 in all levels and in pellets (Table 4). Unbroken mandibles are present in C3 and SL. The greatest modifications are found in PI and C3 (category 5) and in PII (category 4). Concerning postcranial elements, the most frequent in all levels is the presence of complete remains, with femora being the best preserved. In SL there are differences in breakage between the remains recovered from sieved sediments and from pellets, although the postcranial elements were mainly complete.
Digestion degree
Evidences of digestive action are indicated for all levels in Table 5. Figure 4a–f illustrate different examples of modifications by digestive corrosion.
In all samples (NISP = 279), remains without evidence of digestion predominate (NISP = 194); only one remain from PI presents slight evidence of digestion. In PII, remains with light evidence of digestion increase compared to PI, and there are also one incisor with light digestion and one with heavy digestion. In C3, there are three remains with light and four with moderate evidences of digestion. In SL, remains with light digestion predominate, but there are remains (NISP=12) with moderate, heavy, and extreme evidences. Extreme digestion is mainly found among remains still included in the pellets
Processes affecting bone color
In PI and PII, impregnation with manganese oxides is observed in several remains (NISP%=0.7), including changes of the original surface color to small dark brown stains.
Evidence of thermoalteration (NISP = 26) is recorded in low proportions in cranial and postcranial remains recovered from PI (NISP% = 8.0), PII (NISP% = 10.4), and C3 (NISP% = 13.4) (Fig. 4), whereas thermoaltered remains were not found in SL. In PI, some isolated molars (NISP% = 6.0) and maxillae (NISP% = 2.0) are completely burnt (reddish-brown). In PII, some mandibles (NISP% = 0.9), maxillae (NISP% = 1.9), humeri (NISP% = 0.9), tibiae (NISP% = 5.7), and one humerus are also completely burnt (reddish-brown). In C3, isolated incisors (NISP% = 1.2) and remains of mandibles (NISP% = 2.4), maxillae (NISP% = 1.2), and tibiae (NISP% = 4.9) are completely burnt (reddish-brown), and one mandible from this unit is completely calcined. Finally, two tibiae of Caviinae from PII and C3 (NISP% = 0.9 and NISP% = 1.2, respectively) present burning pattern on their distal extremities (Fig. 4g).
Taxonomical representation and paleoenvironmental analysis
Rarefaction curves reflect the strong relationship between specific richness (S) and sample sizes (MNI). The archaeological units and recent pellet samples never reached an asymptote, unlike other pellet samples (Fig. S1), a situation that, coupled with the small MNI studied, hinders the scope of our interpretation. This circumstance, however, does not prevent us from observing general trends on a coarse scale.
The taxonomic identification of mammals in AZ4, their abundances (expressed as NISP and MNI), and the percentage of biomass they represent are detailed in Table 6. Recorded taxa at genus or species level include one marsupial, one mephitid carnivore, three caviomorph rodents, and eight sigmodontine rodents. PI is the unit with the lowest taxonomic richness (S = 4) and diversity (H′ = 1.3). On the contrary, PII registers the highest taxonomic richness (S = 9) and diversity (H′ = 1.9). Intermediate levels of richness and diversity are found in C3 (S = 7; H′ = 1.8) and SL (S = 6; H′ = 1.6).
All bones assigned to Ctenomys sp. are referred to C. haigi, the smallest morphotype recorded in the Limay River basin for the Holocene (e.g., Fernández et al. 2016; Pardiñas 1999a; Pardiñas and Teta 2013; Tammone et al. 2020). The caviomorphs Ctenomys cf. haigi and Microcavia australis and the sigmodontine Phyllotis xanthopygus are recorded in all units; the caviid Galea leucoblephara is present in PII and C3, and the sigmodontine Reithrodon auritus is found in PII, C3, and SL. However, most small mammals are only observed in one or two units: the marsupial Lestodelphys halli in PII and C3; the sigmodontines Eligmodontia sp., Euneomys chinchilloides, and Loxodontomys micropus in PII; the sigmodontines Abrothrix hirta, Akodon iniscatus, and Calomys musculinus in PI, C3, and SL, respectively; and the mephitid Conepatus chinga only in the pellet sample of SL.
In term of biomass, the caviomorphs are the most important prey items in AZ4 with values ranging from 30 to 70% (Table 6), whereas the sigmodontine rodents hardly contribute to the biomass (up to 30%) in all units of AZ4 (Table 6). Recent samples from Strigiformes (B. magellanicus and T. furcata) show overall similar tendencies (samples 1–6, 9, 10, and 13; Table S1). It is also noted that in the pellets included in SL, the contribution of mesomammals to the biomass is higher than that of the small mammals, as within the modern sample #7 produced by G. melanoleucus (Table S1).
These micromammal assemblages broadly suggest the development of open areas of shrubby-grassy Patagonian steppe and large rocky outcrops (e.g., Pardiñas et al. 2003). In addition, the aforementioned taxa are also found in recent pellets yielded by different raptor species (B. magellanicus, G. melanoleucus, and T. furcata) occurring within a ~40 km radius from the rock shelter, suggesting fairly stable ecological conditions for the last ~1000 years. A minor change is suggested by the presence of G. leucoblephara, which is known to inhabit the Monte desert, and is found in PII and C3 units of AZ4, but not within the recent samples recovered from close to the rock shelter. However, in some modern pellet samples (1, 4, 5, 8, 10, 12–14), less diversity is detected (Table S1).
The values of the taxonomic habitat index (THI) show an overall fairly stable habitat from the latest Late Holocene to recent times (Fig. 5a). A major proportion of shrublands and steppes (ranging from ~15 to 25%) is observed during the end of the Late Holocene, and a visible increment of weed/turf (~25 to 30%) and bare habitat (~5 to 18%) in recent samples (Fig. 5a). In fact, small mammal assemblages from AZ4 and recent pellet samples show remarkable similarity, suggesting only minor environmental changes during the last ~1000 years (Table 6). In overall terms, past and present samples are characterized by the continuous co-occurrence of P. xanthopygus and R. auritus along with other taxa (e.g., Eligmodontia sp. and E. chinchilloides) which guarantees the permanency of the aforementioned habitats (Fig. 5b).

a Taxonomical habitat index calculated for the archaeological site Álvarez 4 and selected owl pellet samples from the main phytogeographic units of the area close to the archaeological site. b Temporal variation of the relative abundance (MNI) of selected rodent taxa for the same samples in the previous graph
Correspondence analysis results (Fig. 6) of the archaeological sample ordination reveal positive values on axis 1 (29% of the total variance) and negative values on axis 2 (19.4% of the variance), mainly affected by the caviid species (G. leucoblephara and M. australis). Contrary to the archaeological samples, some generalist and opportunistic modern species (Abrothrix olivacea, Eligmodontia sp., and Oligoryzomys longicaudatus) are ordered toward negative values on axis 1 and positive values on axis 2. Positive values of both axes group a mixture of Patagonian (L. halli, R. auritus) and Monte taxa (Akodon dolores, A. iniscatus, Calomys musculinus, and Thylamys pallidior), whereas negative values group taxa typically allied to the forest and humid environments (A. hirta, Geoxus valdivianus, L. micropus, and Paynomys macronyx). The species most strongly associated with the forest (G. valdivianus) is not present in AZ4, suggesting that this biome did not reach this area during the last ~1000 BP.

Correspondence analysis of the relationships among species abundance between archaeological and modern samples at regional scale
Discussion
Taphonomical interpretation
The finding of pellets in SL of AZ4 is not striking in the context of Patagonia, where the preservation of these regurgitations is quite common in archaeological sites (e.g., Crivelli Montero et al. 1996; Fernández et al. 2012, 2016; Pardiñas 1999a, b). This conservation is due to features of the site (cave and rock shelters), high-speed burial in soft soils, low biological activity (mainly lepidopteran larvae that feed on hairs and feathers), low temperatures, and low humidity (Andrews 1990; Pardiñas 1999b; Terry 2004). The box plot in Figure S2 shows that the length of the pellets from SL is above the mean of pellets from avian raptors that inhabit the area, being similar to the value of pellets assigned to Geranoaetus polyosoma (Trejo and Ojeda 2002). The width calculated for pellets from SL is slightly above that observed for 50% of the samples, matching with the width of pellets of Parabuteo unicinctus (Trejo and Ojeda 2002). Modern samples assigned to P. unicinctus were not evaluated in this work, but the distribution of this species is within the area of AZ4. However, Trejo and Ojeda (2002) found an overlapping in pellet measurements among the large raptors, suggesting that the aforementioned identifications regarding AZ4 pellet origin may be questionable.
In PII and C3, the proportion between biomass values obtained for Ctenomys cf. haigi and M. australis is similar to those indicated for B. magellanicus (Zapata et al. 2015). However, the values from PI, PII, and C3 also coincide with the possible range of contribution of caviomorph rodents to the diet of Athene cunicularia (Montalvo et al. 2020, and references therein). In this sense, several studies stated the great variability of the contribution of caviomorphs to the diet of A. cunicularia, pointing out the opportunistic feeding habits of this owl (Montalvo et al. 2020, and references therein). In SL, the proportion of biomass values for caviomorph and cricetid rodents, together with the identification of small to medium mammals, may suggest accumulations produced by diurnal birds (e.g., Bó et al. 2007). Accordingly, some diurnal birds inhabiting the area, such as G. melanoleucus and Caracara plancus, besides consuming microvertebrates, may feed on dead animals of different sizes (Montalvo and Fernández 2019).
Cranial elements predominate at all levels, as well as, to a lesser extent, isolated teeth and limb bone elements (with slight differences between levels). Indexes obtained for all units are similar to those for South American prey of diurnal birds and mammals (Montalvo and Fernández 2019). Even so, it must be considered that factors unrelated to predatory activity (trampling, dispersal) may have modified the proportion of each type of remain. When only the SL level containing the pellets is evaluated, the indexes are close to those obtained for prey of G. melanoleucus (López et al. 2017).
The average of relative abundance is low in all cases. It is worth noting that Montalvo and Fernández (2019) reported an average relative abundance of 42.9% for Strigiformes samples and an average of 20% for diurnal raptor pellets. Values for levels PI, PII, and C3 are lower than those of modern raptors, and only the average of SL displays a similar value to that obtained for different diurnal birds (Montalvo and Fernández 2019), but it should be noted that the small size of the archaeological samples may be contributing to this variance. In Fig. 3c, the curves of relative abundance of units PI, PII, and C3 are compared with the averages obtained from samples generated by A. cunicularia (Montalvo et al. 2020), B. magellanicus (Montalvo et al. 2016), and T. alba (Andrews 1990) (Fig. 3b), whereas those of level SL (that includes the pellets) are compared with samples generated by C. plancus (Montalvo and Tallade 2009), G. melanoleucus, and G. polyosoma (López et al. 2017).
In PI, PII, and C3, percentages of preserved remains are in all cases lower than those of nocturnal raptors, except for the isolated incisors recovered from PI. There are strong differences in the representation of limb bones, as well as in the skull elements, which are very low in the stratigraphic units. These results may be biased by the breakage and loss of skeletal elements, related to trampling in spaces with reduced mobility, such as the rock shelter AZ4 (Pardiñas 1999a). Trampling by predators themselves may cause dispersion and destruction of consumed bones. The abundance of isolated incisors may be related to skull and mandible breakage; the scarcity of isolated molars, as well as other fragile skeletal elements such as scapulae and vertebrae, may also be attributed to this process. The comparison between the degree of breakage of bones from pellets and those recovered from the sieved sediments of the SL shows that the proportion of broken elements is higher in the latter (pellets = 31.2%; SL without pellets = 73.7%). This difference is statistically significant (chi-square = 32.337; df = 6; p = 0.0001; Monte Carlo p = 0.0002). In particular, the standardized residuals observed for broken postcranial elements from the SL sieved sediments and for complete postcranial elements from pellets have higher values than expected (>1.96). This result supports that the pellet bones are better protected from trampling than elements on SL sediments (see Andrews 1990; Pardiñas 1999b). In SL and mainly among remains from pellets, which are the most frequent in this level, there is a high representativeness of cranial elements and tibiae, and the generated curve is similar (although with low proportions) to pellets of the two species of Geranoaetus (Fig. 3c). Two skulls and five mandibles were found complete in the archaeological pellets, as well as complete mandibles in C3. In all units, the representativeness of the different breakage categories of skulls and mandibles varies. Regarding the postcranial elements, femora, radii, and ulnae are preserved complete in all units in which they were recorded. Humeri display more variability; although complete humeri are abundant in all units of the profile, distal portions are also preserved in a lower proportion. Based on these features, the breakage degree is similar to the average observed for Strigiformes (Montalvo and Fernández 2019), and the differences may be attributed to trampling processes. The completeness of the remains found in pellets is compared with prey bones accumulated by species of Geranoaetus, Caracara plancus, and Milvago chimango, which show a very high degree of skull breakage, but mandibles have a high proportion of complete elements. The breakage degree of limb bones is variable for these species, but some complete elements were recorded (Montalvo and Fernández 2019).
The degree of digestion on bones from units PI, PII, and C3 is similar to that observed in prey of nocturnal raptors (Strigiformes). The values obtained for prey remains of Tyto furcata show that the absence of digestion is the most common evidence (Montalvo and Fernández 2019), whereas prey of A. cunicularia and B. magellanicus, as in AZ4 levels, shows light, moderate, and extreme digestion values in different proportions (Montalvo et al. 2016, 2020). These species nest in caves and rocky shelters, and mainly feed on nocturnal and twilight-active small mammals within a hunting range of 3 km (Andrews 1990). In the particular case of PII and C3, the record of some isolated teeth with moderate and heavy digestive corrosion, respectively, suggests the action of B. magellanicus (samples from A. cunicularia and T. furcata show scarce isolated teeth with these features; Montalvo and Fernández 2019). The averages obtained for AZ4 units are also comparable to those of the species of Geranoaetus (Fig. S3). The results show strong differences in the proportions of each digestion category, mainly concerning remains without digestive evidence. In sum, the presence of remains without evidence of digestion supports the action of a nocturnal raptor for the accumulations of PI, PII, and C3 of AZ4.
Remains with moderate, heavy, and extreme digestion predominate in pellets. Only a few skeletal elements lack evidence of digestion, as was stated by López et al. (2017) and Montalvo and Tallade (2009) for samples of Geranoaetus spp. and C. plancus, respectively. A moderate to high proportion of specimens with light digestive corrosion is also observed, as well as a moderate to high percentage of postcranial remains with moderate digestive corrosion. Heavy digestion is observed on teeth in situ, while the extreme type is recorded in femora. These results are similar to those obtained in samples of diurnal raptors (Montalvo and Fernández 2019). Particularly, the obtained values are similar to those of samples of Geranoaetus spp. from Mendoza Province studied by López et al. (2017), although there are some differences that may be attributed to the sample size. In AZ4, the MNE (134; Fig. S4a) is low with respect to the samples from Villavicencio (MNE = 424; Fig. S4b), Agua de la Mula (MNE = 318; Fig. S4c), and Arroyo Panchino II (MNE = 1177; Fig. S4d).
Small mammals and human subsistence
The finding in PII and C3 of two tibiae of Caviinae rodents distally burned indicates human consumption. This thermoalteration pattern is related to the cooking of the caviomorph rodents directly on charcoal fire, through which the muscles and tissues of distal zeugopodials are retracted, leaving bones exposed to high temperatures (e.g., Fernández et al. 2017a; López and Chiavazza 2020; Medina et al. 2012; Pardiñas 1999b). Two species of Caviinae—Microcavia australis and Galea leucoblephara—are identified in AZ4. These species and Ctenomys have also been part of the food of humans recorded in several archaeological sites located in the Limay River basin, mainly of Late Holocene age (Andrade 2015; Andrade and Boschín 2015; Bond et al. 1981; Ceballos 1982, 1987; Crivelli Montero et al. 1996, 2017; Fernández et al. 2016; Guillermo et al. 2020a; Hajduk and Albornoz 1999; Lezcano et al. 2010; Pardiñas 1999a, b; Pardiñas and Teta 2013; Silveira and Cordero 2014; Tammone 2017; Teta et al. 2005). These low-ranked resources play a complementary role in the overall context of diet breadth, where large- (Lama guanicoe and Rhea pennata) and medium-sized (Chaetophractus villosus, Conepatus chinga, Lagidium viscacia, Lycalopex gymnocercus, and Zaedyus pichiy) animals have a greater relevance to the human economy, such as at AZ4 and other synchronic archaeological sites in the area (e.g., Andrade and Boschín 2015; Fernández et al. 2016, 2017a; Guillermo and Fernández 2019; Guillermo et al. 2020a; Silveira and Cordero 2014).
The broader diet is framed in a regional intensification process that embraces a set of cultural patterns that have certain visibility in the archaeological record. In a general framework of environmental stability with some variability (vide infra) ~3000 years BP (taking into account uncertainties of radiocarbon dates), the human population of the Limay River basin increased. This is inferred from the observation of successive new technologies, such as pottery, bone tools and arrowheads, more use of local lithic sources, increased artifact deposits, the proliferation of rock and mobiliar art, and the occurrence of specialized archaeological sites (e.g., Boschín 2009; Crivelli Montero 2010; Crivelli Montero and Fernández 2004). According to this scenario, the Late Holocene regional hunter-gatherers reduced their residential mobility and moved toward seminomadic conditions (living within well-defined territories), increased their logistical mobility (Bettinger et al. 2015; Binford 1980), and exploited a wide range of resources from high to low energy return. In this way they decreased their foraging efficiency compared to hunter-gatherers from the Early and Middle Holocene (e.g., Ceballos 1982; Cordero 2009, 2011a, b; Crivelli Montero et al. 1993, 1996; Hajduk et al. 2004, 2007).
Paleoenvironments
The taphonomic analysis suggests that the micromammal remains found in AZ4 were incorporated by nocturnal and diurnal raptors, and also humans. These predators have different home ranges and hunting preferences; hence, they yield their own kind of accumulations. For example, South American Strigiformes feed mainly on nocturnal and solitary sigmodontine rodents within a small home range, whereas Accipitriformes and humans direct their hunting toward larger and diurnal rodents, such as caviids and ctenomyids, within a larger home range (e.g., Andrews 1990; Fernández et al. 2017a). These differential taphonomic trajectories, along with the small-sized sample and the disturbed area found in part of the AZ4 sequence, yield limitations for paleoenvironmental interpretations.
The paleoenvironmental analysis based on the small mammals recovered from AZ4 shows a fairly overall stability of type of habitats, where the dominance of shrublands and steppes, intermixed with large rocky outcrops, signed the last ~1000 years. The finding of a higher taxonomic diversity and the presence of Galea leucoblephara in AZ4 (absent in the modern pellet samples nearest to the rock shelter) suggests more heterogeneous environments than today. Some variability is also expressed in other small mammal fossil records close to AZ4. For example, the sequence of small mammals from the archaeological sites Cueva y Paredón Loncomán, La Marcelina 1, and Sarita I, II, and IV caves (Fig. 1a) is fairly similar to that of AZ4, although some elements are allied to more humid conditions (Holochilus vulpinus in La Marcelina 1) for the last 2800 years (Crivelli Montero et al. 2017; Massoia 1982; Teta et al. 2005). In contrast, the archaeological site Casa de Piedra de Ortega, located in the same environment as AZ4 (Fig. 1a), revealed a small mammal assemblage typically allied to Monte desert (Calomys musculinus, Galea leucoblephara, Graomys griseoflavus) and grasslands (Reithrodon auritus) about 300 years BP (Guillermo et al. 2020a). Other small mammal fossil records from the upper and middle Limay River show minor variations during the Holocene (Crivelli Montero et al. 1996; Fernández et al. 2016, 2018; Guillermo et al. 2020b; Pardiñas 1999a; Pardiñas and Teta 2013; Pearson 1987; Pearson and Pearson 1993; Tammone et al. 2014, 2020; Teta et al. 2005).
The pollen records and charcoal data from the upper Limay River basin show a general stability through the Late Holocene, with some minor variations associated with the interannual climate variability of El Niño-Southern Oscillation (e.g., Bianchi 2007; Heusser 1993; Markgraf 1983; Whitlock et al. 2006). In addition, the pollen record from the archaeological site Epullán Grande, located in the middle Limay River basin in the same biogeographic unit as AZ4 (Fig. 1a), displays a mixed scrubby-herbaceous steppe ~2200 years cal. BP (Prieto and Stutz 1996). On the other hand, the progressive advance of forests due to the increasing rainfall during the Early and Late Holocene, detected by these paleoenvironmental proxies, did not reach the AZ4 area and surroundings. This is evidenced by the absence of small mammals typically allied to forests (e.g., Geoxus valdivianus) in the archaeological sequences of AZ4, Cañadón Las Coloradas 1, Casa de Piedra de Ortega, Cueva y Paredón Loncomán, Epullán Grande, La Marcelina 1, and Sarita I, II, and IV caves (Fig 1a), among other archaeological sites in the Patagonian steppe (Crivelli Montero et al. 1996, 2017; Fernández et al. 2016; Guillermo et al. 2020b; Massoia 1982; Pardiñas 1999b; Pardiñas and Teta 2013; Teta et al. 2005).
On a broader scale, the small mammals, parasites, pollen, macrobotanical remains, and packrat midden records recovered from the archaeological site Cueva Huenul 1, located in an ecotone of Monte and Patagonian steppe in the extreme north of Neuquén Province, revealed a fair stability with minor changes since the end of the Last Glacial Maximum to the present (Fernández et al. 2012; Llano et al. 2020). These records show an environment characterized by a mosaic of shrubby steppes, open bare areas, and large rocky outcrops, possibly a little less arid than today by the absence of typical Monte elements (Fernández et al. 2012; Llano et al. 2020). This stability contrasts with other Late Pleistocene/Holocene small mammal records from upper Limay River basin and suggests differential biotic responses that could be associated with distance to under-ice zones (Pearson 1987; Pearson and Pearson 1993; Tammone et al. 2014, 2020).
Finally, modern pellet samples of the area have a lower taxonomic diversity and an increase of some generalist and opportunistic species, such as Abrothrix olivacea, Eligmodontia sp., and Oligoryzomys longicaudatus (Crivelli Montero et al. 1996; Fernández et al. 2016; Pardiñas 1999a; Pardiñas and Teta 2013; Pearson 1987; Pearson and Pearson 1993; Tammone et al. 2014, 2020; Teta et al. 2005). This new configuration of modern small-mammal communities was triggered by the widespread environmental degradation mostly caused by livestock during the Anthropocene (e.g., Tammone et al. 2020), characterized by the increase of shrubby areas with large patches of bare soils and the loss of herbaceous cover (e.g., Bertiller and Bisigato 1998).
Conclusion
In view of their taxonomic and taphonomic characteristics, the accumulations of micromammals from PI, PII, and C3 (963–187 cal. BP) may be attributed to the action of Strigiformes (possibly Athene cunicularia or Bubo magellanicus). In turn, pellets and dispersed bones found in the SL (187 cal. BP) would have been deposited by Accipitriformes (likely Geranoaetus). Bones from pellets were more protected from trampling than the other bones recovered from the sieved sediments of the SL. In addition, some remains of Caviidae (Galea leucoblephara and/or Microcavia australis) from PII and square C3 would have been incorporated by human consumption. This low-ranking resource may have complemented the otherwise high-ranking animals found in AZ4, suggesting a significant diet breadth as it is frequently seen in other archaeological sites in the same area.
In this context, the results of the taphonomic analysis of the micromammal assemblage from AZ4 suggest the action of nocturnal and diurnal raptor birds, and humans. This palimpsest generated by these three types of predators is recorded for the first time in the Limay River basin.
The paleoenvironmental analysis based on the small mammals recovered from AZ4 shows the development of open areas of shrubby-grassy Patagonian steppe and large rocky outcrops with a rather overall stability of habitats, where the dominance of shrublands and steppes signed the last ~1000 years. The past environments were more heterogeneous than today as evidenced by the absence of the species Galea leucoblephara and the expansion of generalist and opportunistic species, such as Abrothrix olivacea, Eligmodontia sp., and Oligoryzomys longicaudatus, in the modern pellet samples nearest to the rock shelter.
References
Andrade A (2015) Distinguishing between cultural and natural depositional agents: micromammal taphonomy from the archaeological site Cueva y Paredón Loncomán (Patagonia, Argentina). J Archaeol Sci Reports 3:122–131. https://doi.org/10.1016/j.jasrep.2015.06.012
Andrade A, Boschín MT (2015) Explotación de roedores por las sociedades cazadoras-recolectoras de Patagonia durante el Holoceno tardío: de la evidencia arqueológica al registro histórico. Zephyrus 75:101–118. https://doi.org/10.14201/zephyrus201575107124
Andrews P (1990) Owls, Caves and Fossils. University of Chicago Press, Chicago
Badgley C (1986) Counting individuals in mammalian fossil assemblages from fluvial environments. Palaios 1:328–338. https://doi.org/10.2307/3514695
Bailey G (1981) Concepts, timescales and explanations in economic prehistory. In: Sheridan A, Bailey G (eds) Economic archaeology. British Archaeological Reports International Series 9, Oxford, pp 97–117
Bailey G (2007) Time perspectives, palimpsests and the archaeology of time. J Anthropol Archaeol 26:198–223
Bertiller M, Bisigato A (1998) Vegetation dynamics under grazing disturbance. The state-and-transition model for the Patagonian steppes. Ecología Austral 8:191–199
Bettinger R, Garvey R, Tushingham S (2015) Hunter-gatherers archaeological and evolutionary theory. Springer, Nueva York
Bianchi MM (2007) El cambio climático durante los últimos 15.000 años en Patagonia Norte: Reconstrucciones de la vegetación en base a polen y carbón vegetal sedimentario. Boletín Geográfico Universidad Nacional de Comahue 30:39–53
Binford L (1980) Willow smoke and dogs’ tails: hunter-gatherer settlement systems and archaeological site formation. Am Antiquity 45:4–20. https://doi.org/10.2307/279653
Binford L (1981) Bones: ancient men and modern myths. Academic Press, New York
Bó M, Baladrón A, Biondi L (2007) Ecología trófica de Falconiformes y Strigiformes: tiempo de síntesis. Hornero 22:97–115
Bond M, Caviglia S, Borrero L (1981) Paleoetnozoología del Alero de los Sauces (Neuquén, Argentina); con especial referencia a la problemática presentada por los roedores en sitios patagónicos. In: de Bórmida S (ed) Prehistoria de la cuenca del río Limay. 1ª parte: Los aleros del Limay norte (Área de El Chocón). Universidad de Buenos Aires-Facultad de Filosofía y Letras, Buenos Aires, pp 93–109
Boschín MT (2009) Tierra de hechiceros: arte indígena de Patagonia septentrional argentina. Universidad de Salamanca, Salamanca
Ceballos R (1982) El sitio Cuyín Manzano. Centro de Investigaciones Científicas de Río Negro. Serie Estudios y Documentos 9:1–66
Ceballos R (1987) Estrategias de subsistencia en el valle del río Pichileufú Depto. Pilcaniyeu (Río Negro). Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano 12:197–202
Cordero JA (2009) Arqueofauna de las primeras ocupaciones de cueva Epullán Grande. Cuadernos de Antropología 5:159–188
Cordero JA (2011a) Arqueofauna de las ocupaciones tempranas de Cueva Traful I, provincia del Neuquén, Argentina. Arqueología 11:161–194. https://doi.org/10.34096/arqueologia.t17.n0.1842
Cordero JA (2011b) Subsistencia y movilidad de los cazadores-recolectores que ocuparon Cueva Traful I durante el Holoceno Medio y Tardío. Comechingonia 5:158–202
Crivelli Montero EA (2010) Arqueología de la cuenca del río Limay. In: Masera R, Casamiquela R, Miotti L, Berón M, Martínez G, Cúneo EM, Crivelli Montero EA, Prates L, Cortes H, Lew J, Membribe A, Cortese V, García R (eds) Los ríos mesetarios norpatagónicos: aguas generosas del Ande al Atlántico. Gobierno de Río Negro - Ministerio de Producción, Viedma, pp 261–338
Crivelli Montero EA, Curzio D, Silveira M (1993) La estratigrafía de la cueva Traful I (provincia del Neuquén). Præhistoria 1:9–160
Crivelli Montero EA, Fernández M (2004) Demografía, movilidad y tecnología bifacial en sitios de la cuenca del río Limay. In: Masera R, Casamiquela R, Miotti L, Berón M, Martínez G, Cúneo E, Crivelli Montero EA, Prates L, Cortes H, Lew J, Membribe A, Cortese V, García R (eds) Los ríos mesetarios norpatagónicos: aguas generosas del Ande al Atlántico. Gobierno de Río Negro - Ministerio de Producción, Viedma, pp 89–103
Crivelli Montero EA, Ramos M, Cordero JA, Fernández FJ, Vitores M, Homar A (2017) Arqueología del cañadón Fta Miche, provincia de Río Negro, noroeste de la Patagonia. Informe preliminar. In: de Haro MT, Rocchiettti A, Runcio A, Fernández M y Hernández de Lara O (comp) Anti. Latinoamérica: una mirada desde el presente hacia el pasado. IX Coloquio Binacional Argentino Peruano. Aspha, Buenos Aires, pp 67-98
Crivelli Montero EA, Palacios O (2010) Dos fragmentos de placas grabadas procedentes del alero Álvarez 4, Coquelén. Pcia. de Río Negro. Rastros. Arqueología e Historia de la cuenca del río Limay 3:597–621
Crivelli Montero EA, Pardiñas UFJ, Fernández M, Bogazzi M, Chauvin A, Fernández V, Lezcano M (1996) La Cueva Epullán Grande (Provincia de Neuquén, Argentina). Informe de Avance. Praehistoria 2:185–265
Evans E, Van Couvering J, Andrews P (1981) Palaeoecology of Miocene sites in Western Kenya. J Human Evol 10:99–116. https://doi.org/10.1016/S0047-2484(81)80027-9
Fernández FJ, del Papa L, Mange E, Teta P, Crivelli Montero EA, Pardiñas UFJ (2016) Human subsistence and environmental stability during the last 2200 years in Epullán Chica cave (northwestern Patagonia, Argentina): a perspective from the zooarchaeological record. Quaternary International 391:38–50. https://doi.org/10.1016/j.quaint.2015.06.013
Fernández FJ, Montalvo CI, Fernández-Jalvo Y, Andrews P, López JM (2017b) A re-evaluation of the taphonomic methodology for the study of small mammal fossil assemblages of South America. Quaternary Sci Rev 155:37–49 https://doi.org/10.1016/j.quascirev.2016.11.005
Fernández FJ, Tammone M, Aguilar JP, Crivelli Montero EA, Pardiñas UFJ (2018) La acción volcánica sobre pequeños mamíferos terrestres en cuatro sitios arqueológicos de la Norpatagonia. In: Gascón M (coord) Historias de volcanes y sociedades. Biblos, Buenos Aires, pp 57–76
Fernández FJ, Teta P, Barberena R, Pardiñas UFJ (2012) Small mammal remains from Cueva Huenul 1, Argentina. Taphonomy and paleoenvironments since the Late Pleistocene. Quaternary Int 278:22–31. https://doi.org/10.1016/j.quaint.2012.01.005
Fernández FJ, Teta P, Pardiñas UFJ (2017a) Evidencias arqueológicas de explotación antrópica de micromamíferos en el extremo austral de América del Sur. Anales de Arqueología y Etnología 72:9–32
Fernández J (1988-1990) Bioarqueología. La Cueva Haichol. Arqueología de los pinares cordilleranos del Neuquén. Anales de Arqueología y Etnología 43-45:583-597
Fernández-Jalvo Y, Andrews P (1992) Small mammal taphonomy of gran Dolina, Atapuerca (Burgos), Spain. J Archaeol Sci 19:407–428. https://doi.org/10.1016/0305-4403(92)90058-B
Guillermo A, Fernández FJ (2019) Análisis zooarqueológico y tafonómico del sitio Vaca Mala (Alicurá, Neuquén). In: Laguens A, Bonnin M, Marconetto B (c) (eds) Libro de Resúmenes XX Congreso Nacional de Arqueología Argentina: 50 años de arqueologías. Universidad Nacional de Córdoba, Córdoba, pp 314–315
Guillermo A, Fernández F (2021) Análisis zooarqueológico del sitio Álvarez 4 (noroeste de Patagonia, Argentina). Un aporte al estudio de la subsistencia de los grupos humanos de la cuenca media del río Limay durante el Holoceno tardío. Arqueología: in press
Guillermo A, Fernández FJ, Cordero JA (2020a) Impacto de la fauna exótica doméstica en la subsistencia humana en la cuenca superior del río Limay: la evidencia de Casa de Piedra de Ortega (Río Negro, Argentina). Arqueología 26:171–195. https://doi.org/10.34096/arqueologia.t26.n1.5821
Guillermo A, Fernández FJ, Crivelli Montero EA (2020b) Zooarqueología y tafonomía del sitio Cañadón Las Coloradas 1 (Holoceno tardío, Río Negro, Argentina). Chungará Revista de Antropología Chilena. https://doi.org/10.4067/S0717-73562020005002301
Hajduk A, Albornoz A (1999) El sitio Valle Encantado I. Su vinculación con otros sitios: un esbozo de la problemática local diversa del Nahuel Huapi. In: Belardi J, Fernández P, Goñi R, Guráieb G, De Nigris M (eds) Soplando en el Viento. Universidad Nacional de Comahue – Instituto Nacional de Antropología y Pensamiento Latinoamericano, Neuquén-Buenos Aires, pp 371–391
Hajduk A, Albornoz A, Lezcano M (2004) El “Mylodon” en el patio de atrás. Informe preliminar sobre los trabajos en el sitio El Trébol, ejido urbano de San Carlos de Bariloche, Provincia de Río Negro. In: Civalero MT, Fernández PM, Guráieb AG (eds) Contra viento y marea. Arqueología de Patagonia. Instituto Nacional de Antropología y Pensamiento Latinoamericano - Sociedad Argentina de Antropología, Buenos Aires, pp 715–731
Hajduk A, Albornoz A, Lezcano M (2006) Levels with extinct fauna in the forest rockshelter El Trébol (Northwest Patagonia, Argentina). Current Research in Pleistocene 23:55–57
Hajduk A, Arias Cabal P, Chauvín A, Albornoz A, Armendariz Gutiérrez A, Cueto Raspado M, Fernández Sanchéz P, Fernández V, Goye S, Lezcano M, Tapia Sagarna J, Teira Mayolini L (2007) Poblamiento temprano y arte rupestre en el área del lago Nahuel Huapi y cuenca del río Limay (Pcias. de Río Negro y Neuquén, Argentina). In: Tras las Huellas de la Materialidad. Resúmenes Ampliados del XVI Congreso Nacional de Arqueología Argentina, Tomo III. Universidad Nacional de Jujuy, San Salvador de Jujuy, pp 393–399
Hammer Ø (1999-2020) PAST PAleontological STatistics Version 4.03. Reference manual. University of Oslo, Oslo
Heusser C (1993) Palinología de la secuencia sedimentaria de la Cueva Traful I (provincia del Neuquén, República Argentina). Præhistoria 1:206–210
Lezcano M, Hajduk A, Albornoz A (2010) El menú a la carta en el bosque ¿entrada o plato principal?: una perspectiva comparada desde la Zooarqueología del sitio El Trébol (Parque Nacional Nahuel Huapi, Pcia. Río Negro). In: De Negris M, Fernández P, Giardina M, Gil A, Gutiérrez M, Izeta A, Neme G, Yacobaccio H (eds) Zooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio. Ediciones del Espinillo, Buenos Aires, pp 243–257
López JM, Chiavazza H (2020) From owl prey to human food: taphonomy of archaeological small mammal remains from Late Holocene wetlands of arid environments in Central Western Argentina. Archaeol Anthropol Sci 12. https://doi.org/10.1007/s12520-020-01213-z
López JM, Fernández FJ, Montalvo CI, Chiavazza H, De Santis L (2017) The role of the Accipitriformes Geranoaetus melanoleucus and Geranoaetus polyosoma as small mammal bones accumulator in modern and archaeological sites from Central Western Argentina. J Taphonomy 15:91–108
Llano C, De Porras M, Barberena R, Timpson A, Beltrame M, Marsh E (2020) Human resilience to Holocene climate changes inferred from rodent middens in drylands of northwestern Patagonia (Argentina). Palaeogeography, Palaeoclimatol, Palaeoecol 557:109894. https://doi.org/10.1016/j.palaeo.2020.109894
Malinsky-Buller A, Hovers E, Marder O (2011) Making time: ‘living floors’, ‘palimpsests’ and site formation processes – a perspective from the open-air Lower Paleolithic site of Revadim Quarry, Israel. J Anthropol Archaeol 30:89–101. https://doi.org/10.1016/j.jaa.2010.11.002
Marín-Monfort MD, García Morato S, Olucha R, Yravedra J, Piñeiro A, Barja I, Andrews P, Fernández-Jalvo Y (2019) Wildcat scats: taphonomy of the predator and its micromammal prey. Quaternary Sci Rev 225:106024. https://doi.org/10.1016/j.quascirev.2019.106024
Markgraf V (1983) Late and postglacial vegetational and paleoclimatic changes in subantarctic, temperate, and arid environments in Argentina. Palynology 7:43–70
Marti C, Bechard M, Jaksic F (2007) Food habits. In: Bird D, Bildstein K (eds) Raptor research and management techniques. Hancock House, Blaine, pp 129–152
Massoia E (1982) Restos de mamíferos recolectados en el paraje Paso de los Molles, Pilcaniyeu, Río Negro. Revista de Investigaciones Agropecuarias 17:39–53
Massoia E (1987) Restos de mamíferos recolectados en el sitio arqueológico de Chenque Haichol, Picunche, Provincia del Neuquén. Estudios de Antropología Pampeana 2:15–21
Medina M, Teta P, Rivero D (2012) Burning damage and small-mammal human consumption in Quebrada del Real 1 (Cordoba, Argentina): an experimental approach. J Archaeol Sci 39:737–743. https://doi.org/10.1016/j.jas.2011.11.006
Menéndez F [1791-1794] (1896) Diarios del P. Fr. Francisco Menéndez, Predicador General Apostólico del Colegio de Santa Rosa de Ocopa. Niemeyer, Valparaíso
Montalvo CI, Fernández FJ (2019) Review of the actualistic taphonomy of small mammals ingested by South American predators. Its importance in the interpretation of the fossil record. Publicación Electrónica de la Asociación Paleontológica Argentina 19:18–46. https://doi.org/10.5710/PEAPA.11.03.2019.275
Montalvo CI, Fernández FJ, Tallade P (2016) The role of Bubo virginianus magellanicus as rodent bone accumulator in archaeological sites. A case study for the Atuel River (Mendoza, Argentina). Int J Osteoarchaeol 26:974–986. https://doi.org/10.1002/oa.2509
Montalvo CI, Fernández FJ, Tomassini RL, Mignino J, Kin MS, Santillán MA (2020) Spatial and temporal taphonomic study of bone accumulations of the burrowing owl (Athene cunicularia) in central Argentina. J Archaeol Sci Reports 30:1–11. https://doi.org/10.1016/j.jasrep.2020.102197
Montalvo C, Tallade P (2009) Taphonomy of the accumulations produced by Caracara plancus (Falconidae). Analysis of prey remains and pellets. J Taphonomy 7:235–248
Oyarzabal M, Clavijo J, Oakley L, Biganzoli F, Tognetti P, Barberis I, Maturo H, Aragón R, Campanello P, Prado D, Oesterheld M, Rolando L (2018) Unidades de vegetación de la Argentina. Ecología Austral 28:40–63. https://doi.org/10.25260/EA.18.28.1.0.399
Pardiñas UFJ (1999a) Los Roedores Muroideos del Pleistoceno Tardío-Holoceno en la Región Pampeana [Sector Este] y Patagonia [República Argentina]. Aspectos Taxonómicos, Importancia Bioestratigráfica y Significación Paleoambiental (Ph.D thesis). Universidad Nacional de La Plata, La Plata
Pardiñas UFJ (1999b) Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia (Argentina). Arqueología 9:265–340
Pardiñas UFJ, Teta P (2013) Holocene stability and recent dramatic changes in micromammalian communities of northwestern Patagonia. Quaternary Intl 305:127–140. https://doi.org/10.1016/j.quaint.2012.08.001
Pardiñas UFJ, Teta P, Cirignoli S, Podestá D (2003) Micromamíferos (Didelphimorphia y Rodentia) de Norpatagonia Extra Andina, Argentina: taxonomía alfa y biogeografía. Mastozoología Neotropical 10:69–113
Paruelo JM, Beltrán A, Jobbágy E, Sala OE, Golluscio RA (1998) The climate of Patagonia: general patterns and controls on biotic processes. Ecología Austral 8:85–101
Patton JL, Pardiñas UFJ, D’Elia G (2015) Mammals of South America 2 rodents. University of Chicago Press, Chicago and London
Pearson OP (1987) Mice and the postglacial history of the Traful valley of Argentina. J Mammal 68:469–478. https://doi.org/10.2307/1381583
Pearson OP (1995) Annotated keys for identifying small mammals living in or near Nahuel Huapi National Park or Lanin National Park, southern Argentina. Mastozoología Neotropical 2:99–148
Pearson AK, Pearson OP (1993) La fauna de mamíferos pequeños de Cueva Traful I, Argentina: pasado y presente. Praehistoria 1:211–224
Prieto A, Stutz S (1996) Vegetación del Holoceno en el norte de la estepa Patagónica: palinología de la cueva Epullán Grande (Neuquén). Prehistoria 2:267–277
Rosales D (1877) Historia general del Reyno de Chile. Flandes indiano. Imprenta del Mercurio, Valparaíso
Rodríguez MF, Leanza HF, Salvarredy Aranguren M (2007) Neuquén, provincias del Neuquén, Río Negro y La Pampa 1:250,000. Instituto de Geología y Recursos Minerales. Boletín 370:1–165
SAREM (2019) Categorización de los Mamíferos de Argentina. http://cma.sarem.org.ar/index.php/es. Accessed 11 January 2021
Shipman P, Foster G, Schoeninger M (1984) Burnt bones and teeth: an experimental study of color, morphology, crystal structure and shrinkage. J Archaeol Sci 11:301–325
Silveira M, Cordero JA (2014) Zooarqueología del sitio La Marcelina 1 (provincia de Río Negro, Argentina). AtekNa 4:67–141
Smith FA, Lyons S, Morgan Ernest SK, Jones KE, Kaufman DM, Dayan T, Marquet PA, Brown JH, Haskel JP (2003) Body mass of Late Quaternary mammals. Ecology 84:3403
Stahl PW (1996) The recovery and interpretation of microvertebrate bone assemblages from Archaeological contexts. J Archaeol Method Theory 3:31–75
Tammone MN (2017) Pérdida de diversidad genética: implicaciones para la evolución y conservación de dos especies de Ctenomys (Rodentia, Ctenomiydae) en Patagonia norte (Ph. D thesis). Universidad Nacional de Comahue, San Carlos de Bariloche
Tammone MN, Hajduk A, Arias P, Teta P, Lacey EA, Pardiñas UFJ (2014) Last Glacial Maximum environments in northwestern Patagonia revealed by fossil small mammals. Quaternary Res 82:198–208. https://doi.org/10.1016/j.yqres.2014.04.015
Tammone MN, Lacey EA, Pardiñas UFJ (2020) Dramatic recent changes in small mammal assemblages from Northern Patagonia: a caution for paleoenvironmental reconstruction. SAGE 11(30):1579–1590. https://doi.org/10.1177/0959683620941096
Terry RC (2004) Owl Pellet Taphonomy: a preliminary study of the post-regurgitation taphonomic history of pellets in a temperate forest. Palaios 19:497–506
Teta P, Andrade A, Pardiñas UFJ (2005) Micromamíferos (Didelphimorphia y Rodentia) y paleoambientes del Holoceno tardío en la Patagonia noroccidental extra-andina (Argentina). Int J Archaezool 14:183–197
Trejo A, Ojeda V (2002) Identificación de egagrópilas de aves rapaces en ambientes boscosas y ecotonales del noroeste de la Patagonia argentina. Ornitología Neotropical 13:313–317
Udrizar Sauthier DE, Formoso AE, Andrade A, Podestá D, Teta P (2020) Key to cranial and mandibular remains of non-flying small mammals from southern South America. J Archaeol Sci: Reports. https://doi.org/10.1016/j.jasrep.2020.102310
Viedma F [1779](1938) Carta de Francisco de Viedma al Virrey Vértiz del 4 de junio de 1779. Revista de La Biblioteca Nacional 7:504
Vignati M (1964) Antecedentes para la protoetnografía del norte de la Patagonia. Boletín de La Academia Nacional de La Historia 34:493–525
Whitlock C, Bianchi MM, Bartlein PJ, Markgraf V, Marlon J, Walsh M, McCoy N (2006) Postglacial vegetation, climate, and fire history along the east side of the Andes (late 41-42.5S), Argentina. Quaternary Res 66:187–201. https://doi.org/10.1016/j.yqres.2006.04.004
Zapata SC, Procopio D, Travaini A (2015) Caviomorphs as prey: general patterns for mammalian carnivores and a local study for raptors in Patagonia. In: Vassallo A, Antenucci D (eds) Biology of caviomorph rodents: diversity and evolution. SAREM series, Buenos Aires, pp 295–322
Acknowledgements
We thank the archaeological team formed by Eduardo Crivelli Montero, Oscar Palacios, and Agustín Cordero; and to Cecilia M. Deschamps and Esperanza Cerdeño for the English revision.
Availability of data and material
All novel data sets are presented directly in the manuscript; other data used in the analyses are referenced within the text.
Code availability
Not applicable.
Funding
This research was funded by the Consejo Superior de Investigaciones Científicas de Cooperación Internacional (CSIC, Spain; project i-COOPB-20287), the Universidad Nacional de La Plata (Project 11/N769), and the Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa (Project 05-G), Argentina.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Microvertebrate Studies in Archaeological Contexts: Middle Paleolithic to early Holocene past environments




Rights and permissions
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Guillermo, A.A., Montalvo, C.I. & Fernández, F.J. Palimpsest of micromammal deposits in an archaeological rock shelter (Álvarez 4, Late Holocene) from northwestern Patagonia, Argentina. Archaeol Anthropol Sci 13, 178 (2021). https://doi.org/10.1007/s12520-021-01437-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-021-01437-7