
Abstract
Macro- and microscopic plant remains recovered from excavations at Teotihuacan over several decades contribute to an understanding of the subsistence resources available to the city’s inhabitants. However, associated contextual evidence may be inconclusive regarding the specific uses of the plants in question and, particularly, whether their presence or relative abundance indicates processing and consumption or the consequence of depositional processes. While the analysis of archeological plant remains is necessarily an interdisciplinary endeavor, involving aspects of botany, ecology, ethnography and history among other disciplines, contextual archeological evidence provides the matrix for interpretation. In this paper, we explore direct archeological evidence for plant use, considering the significance of appropriate contextual evidence (e.g., activity areas, associated ceramics, lithics, etc.) and the potential complementary role of analytical techniques such as residue analyses, and carbon isotope signatures in faunal and human osteological remains as well as stratigraphic sediments.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Research into foodways and nutrition from an anthropological perspective seeks to develop an interdisciplinary understanding of the relationship between natural resources, their uses as foods, and the nutritional consequences of consumption for human groups. The availability of resources varies on an annual cycle and requires an understanding of the environment and related biomass. Aspects to consider include resource exploitation and production, food preparation, consumption, and associated norms that are culture-specific. Their study calls for a holistic approach that considers technology, food preferences, consumption behavior, and habits, all within their appropriate timeframes.
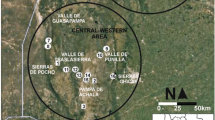
In this paper, we summarize carbonized macrobotanical remains recovered from several excavations carried out at the prehispanic urban center of Teotihuacan, Mexico, in some cases over four decades ago. Corresponding to the so-called Classic period in central Mexico, Teotihuacan (ca. AD 1-650) situated in the NE sector of the Basin of Mexico (Fig. 7) was the site of the first major city in the Americas, the capital of a complex state society that grew to dominate the Basin and adjacent valleys of central Mexico, with significant contacts in southern Mesoamerica (Millon 1988). Teotihuacan’s urban inhabitants occupied characteristic domestic structures generally referred to as apartment compounds, in which several related nuclear families were believed to reside. Various domestic activities were carried out in these structures, ranging from daily subsistence activities including food preparation and consumption, different types of craft production, and ritual practices (Cowgill 2015). Less is known about rural domestic activities on the outskirts of the Classic period city as few excavations in this part of the valley have been undertaken (Cabrera Cortés 2011).
We argue that the study of macrobotanical remains as well as microbotanical materials is fundamental to a more detailed understanding of subsistence practices among the inhabitants of the prehispanic city, and complements other approaches which offer a more general view of plant use. However, in this paper, we focus on macrobotanical evidence.
Archeological studies of plant remains at Teotihuacan
Macroscopic plant remains have been recovered from excavations at Teotihuacan over several decades (Ford and Elias 1972, McClung de Tapia 1977, 1978, 1979, 1987, 1980). For the most part, macroscopic plant materials are preserved as carbonized remains (Minnis 1981), although in some exceptional cases, uncarbonized materials have been recovered in what are clearly unaltered archeological contexts. Furthermore, in some instances, the recovery of carbonized plant materials has been entirely incidental to routine excavation procedures. For example, carbonized plant specimens from the compound known as Zacuala Patios by Laurette Sejourné (1959) were summarily mentioned in the description of burial 11, although additional specimens not mentioned in that publication found their way to the Laboratorio de Paleoetnobotánica y Paleoambiente (LPP), Instituto de Investigaciones Antropológicas (IIA), Universidad Nacional Autónoma de México (UNAM) decades later. Recently, however, the recovery of plant remains is more often included as a basic component of archeological technique, beginning with several of the excavations carried out during the Teotihuacan Mapping Project (TMP, Millon 1973), and continuing with Tlajinga 33 (Storey 1992), Oztoyahualco (Manzanilla 1993), La Ventilla (Cabrera-Castro 1996; Casales-Gómez and Tavera-Sánchez 1996; Montufar 1996), Teopancazco (Martínez and Manzanilla 2005, Paper presented at the Society for Economic Botany; Manzanilla 2012), to mention some examples.
Analysis of the flotation remains recovered from the Teotihuacan Mapping Project (TMP) excavations was first reported in a paper presented by Richard Ford and Joel Elias at the 1972 Society for American Archeology meeting. McClung de Tapia (1979, 1987) provided an initial list of the range of plants that could be expected from archeological contexts in the prehispanic urban zone, as well as a basis for the development of hypotheses concerning economic and ritual uses attributed to these plants. Because the TMP excavations consisted mainly of relatively small stratigraphic test pits in order to support the establishment of a reliable ceramic chronology as well as to define the extension and limits of certain spaces that were unclear following the interpretation of aerial photographs to produce the map of the urban center (Millon 1973), the activity areas discovered could not be defined in detail. Although a considerable range of carbonized plant specimens was recovered from these test pits, particularly underlying the remains of later structures, it was impossible to interpret the contexts in detail or associate them with specific activities beyond evidence for burning. As a result, it was feasible to construct a scheme of plants “available” to the urban residents through time, based on the chronological sequence represented by ceramics associated with the test pits, although interpretations of the uses of these plants remained hypothetical. An attempt was undertaken to analyze the differential availability of both plant and faunal resources at two of the Mapping Project excavations, Tetitla and Yayahuala (McClung de Tapia 1979), suggesting a higher socioeconomic level among residents of the former apartment compound. With respect to animal bones as indicators of food resources, these are considerably more resistant and are better preserved in the archeological record at Teotihuacan. Furthermore, they are often visible in situ, facilitating their recovery, or picked out of the screened sediment during the course of excavation (O’Connor 2000; Reitz and Wing 2001). To date, no evidence of human coprolites has been detected at Teotihuacan.
In more recent excavations in which a concentrated effort was made to recover evidence from activity areas (e.g., Storey 1992, Manzanilla 1993, see also, Montufar 1996, Casales-Gómez and Tavera-Sánchez 1996), macrobotanical plant remains were not always well preserved, or not clearly associated with the activities carried out in particular spaces, but rather associated with different fill materials or more recent intrusion via bioturbation or other processes. In some cases, the minimal depth of the excavations together with historic and modern agricultural practices in their vicinity may have rendered them liable to such disturbance. Another factor, often jokingly cited as a cause for archeologists’ frustration, was the tendency for the urban residents of Teotihuacan to maintain their living surfaces exceptionally clean (Millon 1973). Consequently, it has been difficult to substantiate earlier hypotheses regarding plant use.
Based on descriptions of plant use contained in sixteenth century documents, such as Sahagún (Dibble and Anderson 1963), Martín de la Cruz (Emmart 1940) and Hernández (1942), together with ethnographic descriptions of plant preparation and consumption, working hypotheses may be developed concerning the potential uses of these same taxa recovered from excavations. However, the difficulty in subjecting such hypotheses to verification remains with us due to the vicissitudes associated with the preservation of archeological materials, particularly organic remains. Also, the introduction of European components in the diet has altered modern food preparation techniques with respect to their prehispanic counterparts and ethnographic descriptions of plant use must be used carefully.
Thus, in spite of the relatively substantial database from Teotihuacan now in existence, including a broad range of plants, unquestionable archeological evidence for their uses is limited insofar as the details of preparation, storage and consumption of vegetal materials are concerned. Although larger concentrations of plant remains are occasionally recovered, frequently in association with ceremonial/ritual contexts, they are most often present in low densities.
Selected food plant remains recovered from excavations in Teotihuacan
In addition to domesticates such as maize (Zea mays), or beans (Phaseolus vulgaris), and squash (Cucurbita spp.), many of the plant taxa identified in flotation samples from TMP and other excavations at Teotihuacan pertain to genera and species that could have been intentionally cultivated or exploited as wild plants. It is evident from a glance at the map of Teotihuacan’s urban area (Millon 1973) that at the height of its development very little space was available within the city for either agricultural fields or home gardens. However, the nature of Teotihuacan’s urban configuration during the earliest phases of its development (Patlachique and Tzacualli /Miccaotli phases, ca. AD 0-150) is not well understood as a result of the overlying construction associated mainly to the Tlalmimilolpa and Xolalpan phases. The presence of sediments pertaining to agricultural fields in the fill of monumental structures (Rivera et al. 2007; McClung de Tapia 2012) suggests that the area originally used for crop production was reduced as the city expanded. We do not know the extent or distribution of this area during earlier phases, although it seems reasonable to assume that sediments destined for construction fill were obtained from as nearby as possible.
Table 1 provides a list of selected plant taxa representing food or potential food resources recovered from diverse excavations carried out in Teotihuacan over the past four decades, including Tlajinga 33(33:S3W1; Widmer 1987; Widmer and Storey 1992; Storey 1992), Tlajinga PATT (17:S3W1, 18:S3W1; Carballo and Barba 2014), the Teotihucan Mapping project excavations at Tetitla (I:N2W2) and Yayahuala (1:N3W3; McClung de Tapia 1978, 1979, 1987) as well as Oztoyahualco (15B:N6W3; Manzanilla 1993). Codes following the site names refer to locations on Millon’s 1973 cuadrant maps. It is not a complete list of recovered materials, but rather emphasizes potential food plants and their sources (agricultural or gathered products). Table 2 provides counts and ubiquities for carbonized macrobotanical specimens recovered from the domestic structures previously mentioned. In Table 2, N refers to the number of flotation samples considered from each excavation.
Maize (Zea mays L., Fig. 1) is a domesticated plant unable to reproduce without human intervention and thus cannot be confused with non-domesticates that may be either cultivated or wild. Amaranth (Amaranthus spp.) may be an important agricultural resource as well; in fact, Sanders et al. (1979) considered it a significant contender for this role. However, archeological evidence for domesticated amaranth at Teotihuacan is inconclusive. In many cases, amaranth cannot be differentiated clearly from the fills with which it is usually associated. Seeds are not present in significant quantities in deposits and both carbonized and uncarbonized specimens often occur together in the same samples. Furthermore, although amaranth unquestionably had ritual significance during the Late Postclassic period in the Basin of Mexico (Dibble and Anderson 1963, Hernández 1942), there is no clear archeological evidence as yet to suggest a similar role during the Classic period at Teotihuacan. In addition, amaranth may also have been valued as a quelite, a broad category of tender edible greens from immature plants, in which case its achenes would not be present at all in the archeological record. Chile (Capsicum annuum, Fig. 2), beans (Phaseolus spp., Fig. 3) and squash (Cucurbita spp.), for which little archeological evidence is available at Teotihuacan, were likely important. However, the seeds of both beans and squash were consumed, often leaving few traces. Seasonally available greens (quelites) include Amaranthus spp., Chenopodium spp. and Portulaca oleracea (Fig. 4), all of which are present in archeological deposits at Teotihuacan. Fruits, such as capulín (Prunus serotina ssp. capuli), tejocote (Crataegus mexicana) together with various cactus products including tunas (prickly pears, Fig. 5, and pencas (leaves of Opuntia spp.) and fruits of garambullo (Myrtillocactus geometrizans) are also present. Seeds of cactus fruits other than Opuntia spp. are indicated as Cactaceae in Table 2 as many of these specimens require revision. Chía (Salvia spp., Fig. 6) while not frequent is also present at different excavated sites in the city. Common names for the taxa and their sources are provided in Table 1.

Maize. Zea mays L

Chile. Capsicum annuum L

Common beans. Phaseolus vulgaris L

Purslane (Verdolaga). Portulaca oleracea

Nopal (prickly pear). Opuntia sp.

Chía. Salvia sp.
In Table 3, selected Mexican food plants are described in terms of their potential nutritional value (Pérez-Lizaur and Marván-Laborde 2001). The variability in net weight evident for different foods in the table reflects modern data organized by nutritionists focused on portions that can be understood by users on a daily basis. These data are organized by specialists to facilitate construction of suitable diets based on the comparison between different foods. Although, this kind of information allows us to conclude that prehispanic Mesoamerican populations and Teotihuacan residents in particular had access to the makings of a well-balanced diet, it does not tell us the extent to which these elements were prepared, combined with other components, such as faunal resources, and consumed.
Macrobotanical remains recovered from excavations in two apartment compounds in the Tlajinga area at the southern extreme of the Teotihuacan Valley exemplify what is frequently found in other domestic contexts. Storey (1992) reported maize, beans and amaranth as well as nopal (tuna) at Tlajinga 33. Unpublished specimens recovered from recent excavations by the authors in other structures at Tlajinga undertaken by David Carballo of Boston University, include maize, beans, amaranth, goosefoot, purslane, nopal, and chía (sage), all of which have been reported from TMP excavations and other contexts at Teotihuacan. These data are consistent with earlier summaries of plant remains conserved in the prehispanic city (McClung de Tapia 1979, 1987). TMP excavations are represented by Tetitla and Yayahuala in the central sector of the urban zone, and Manzanilla’s excavation at Oztoyohualco (Manzanilla 1993) in the northwest are also considered. All of these plant materials are catalogued and stored in the Reference Collection of the Laboratorio de Paleoetnobotánica y Paleoambiente (LPP-IIA-UNAM). The volume of sediment recovered from different excavations varied considerably, particular in earlier excavations when standardized methodologies had not yet been developed for research in Central Mexico.
Recent advances in the study of Teotihuacan diet
References to the consumption of different plants and their possible preparation at Teotihuacan have traditionally fallen back on the detailed descriptions of plant use in contexts associated with ritual, medicinal, and food consumption provided by sixteenth century sources, particularly Sahagún (Dibble and Anderson 1963) and Francisco Hernández (1942). These references are of considerable importance for the study of prehispanic plant use in the Basin of Mexico (McClung de Tapia et al. 2014). However, although there is a notable continuity between these sixteenth century descriptions and ethnographic observations by the authors in parts of central México, Teotihuacan flourished and collapsed over a millennium before the Aztec society referred to in these accounts. While it is reasonable to assume that many traditions established by the sixteenth century have their roots in the Teotihuacan period or even earlier, this is difficult to demonstrate given the state of preservation of plants and activity areas in the archeological record for Teotihuacan. In light of these limitations, it is important to apply complementary techniques for the assessment of diet and foodways at Teotihuacan. With respect to botanical evidence for plant use, additional available techniques include the analysis of pollen, phytoliths, and starch grains, as well as the identification of wood and charcoal when present. The recovery of microbotanical remains from stone implements, particularly manos, metates, tecolotes, and molcajetes, has been applied only sparingly and has not been published. However, in spite of the limited specificity of some microbotanical remains with respect to macrobotanical materials, this area of research has enormous potential for the future.
Macro and microbotanical remains associated with storage facilities in archeological contexts are potential sources of evidence—both direct and indirect—for food use, although at Teotihuacan significant densities of plant remains have not been recovered from such contexts. One exception is the recovery of phytoliths and pollen from sediment samples associated with a possible storage area in the apartment compound of Oztoyahualco, indicating the presence of squash (Cucurbita sp.) (González-Vázquez et al. 1993).
Established qualitative techniques for the recovery of pH, phosphates, carbonates, fatty acids, and protein residues from stucco floors have been widely employed in recent years at Teotihuacan. The distributions of these concentrations point to a broad range of different activities involving both domestic and ritual practices, including food storage, preparation, and consumption as well as ritual procedures (Barba-Pingarrón 2007). Similarly, their use in detecting chemical residues preserved in ceramic vessels (Barba-Pingarrón 2009; Barba et al. 2014) is useful. Notable recent applications in this area include determining the presence of cacao in Maya ceramics (Powis et al. 2002; Hurst 2006). More recently, evidence for pulque was determined in Teotihuacan ceramics (Correo-Ascencio et al. 2014; Robertson et al. this volume). In this particular case, it is of interest to note that while several sherds from the urban barrio of La Ventilla showed evidence for pulque, sherds from site 520, a rural community on the city’s margin but situated within a potential zone for agave cultivation and pulque production apparently did not. The determination of specific chemical signatures conserved in vessels is of enormous importance, not only to detect specific food uses and preparation techniques but also to better understand the social role of these plants (and animals).
Pathologies related to nutritional factors in human osteological remains
Unfortunately, evidence for paleopathological conditions related to diet and nutrition in human osteological remains recovered at Teotihuacan is limited as well. Serrano and Lagunas (1999) analyzed burials from La Ventilla B in the southwestern sector of the urban zone, in which dental wear appeared insignificant and degenerative conditions such as arthritis was also sparse. In addition, over 300 burials from additional excavations at La Ventilla were described by Gómez-Chávez and Nuñez-Hernández (1999), although pathologies and traumas present were not discussed in detail. Of 42 individuals in 18 burials from the apartment compound of Oztoyahualco, only three showed evidence of pathologies other than moderate to strong dental wear (Manzanilla et al. 1999a, b). One hundred and ten individuals were recovered from 68 burials in the apartment compound designated as Tlajinga 33 (Storey 1992) in which iron deficiencies manifest in porotic hyperostosis were detected in only four. Manzanilla (2012) reports approximately 116 burials at Teopancazco in the southern sector of the urban zone. Of the 51 sexed individuals, a relatively small number showed evidence for nutritional deficiencies indicated by such pathologies as porotic hyperostosis (11) and cribra orbitalia (4) as well evidence for scurvy (6) and intensity of tooth decay (3) ranging from normal to severe (Manzanilla 2015). In 7 individuals, 2 or 3 of these conditions were simultaneously present, further reducing their overall presence in the sample.
In general terms, chronic infections such as digestive ailments are believed to have affected biological stress levels in Teotihuacan populations (González-Sobrino 2003; Ochoa-Ocaña 2003), possibly associated with inadequate hygiene related to urban living conditions. The quantitative evidence for pathologies associated with inadequate nutrition, therefore, is not significant in statistical terms, but rather insofar as it contributes to a picture of adaptation to pre-industrial urban life. While nutritional differences would be expected among members of distinct social or ethnic groups, the analysis of material archeological evidence for such distinctions is still in its infancy. The number of burials recovered from excavations over the past eight decades does not account for the estimated population of Teotihuacan at its height—ca. 100,000 inhabitants. Obviously, insufficient preservation of human osteological remains as well as a lack of detailed analysis of their characteristics in some cases contributes to this situation. Also, cremation was probably an important mortuary practice, although its intensity is not clear (Arnaud 2014; Meza-Peñaloza 2008; Rattray 1997). However, several authors have noted the apparent absence of evidence for nutritional deficiencies (Rojas-Lago and Serrano-Sánchez 2011), suggesting that much is yet to be learned about the ways in which Teotihuacan’s residents were able to obtain sufficient food to meet their biological needs (see Storey and Widmer this volume).
Stable carbon isotope evidence for human food consumption: issues and limitations
Other significant techniques that have been undertaken more frequently include the recovery of stable isotopes from human remains. At Teopancazco (Manzanilla 2012) and La Ventilla (Arnaud 2014), δ13C values obtained from collagen and tooth enamel indicated tendencies and individual differences relative to the consumption of C3, C4, and CAM plants. In very simplistic terms, these designations refer to the number of atoms associated with photosynthesis; in the case of C3 plants, the first organic compound produced during photosynthesis has three carbon atoms while four are produced in the so-called C4 plants. CAM plants are mainly succulents and members of the Cactaceae family. More detailed discussion of the significance of these categories with reference to the phytosynthetic pathways followed by different types of plants is available from several sources, in the context of Teotihuacan (Arnaud 2014, Morales et al. 2012) and the Maya region (Somerville et al. 2013).
As discussed in McClung de Tapia and Adriano-Morán (2012), differences in the photosynthetic pathways of plant taxa are reflected in different proportions of 13C with respect to 12C. δ13C ratios of C3 plants range from approximately −32 to −20 ‰ with a mean of −27 ‰. The range for C4 plants varies between −17 and −9 ‰, with a mean of approximately −13 ‰. The ratios for C3 and C4 plant taxa therefore differ from one another by roughly −14 ‰. The CAM have δ13C values similar to C4 plants while C3 plants range from −28 to −10 ‰. The CAM taxa analyzed in the Teotihuacan Valley have a mean δ13C ratio of −13.30 ‰ (Lounejeva-Baturina et al. 2006). δ13C ratios can be used to quantify aspects of vegetation dynamics in ecosystems, such as the changes in relative proportions of plants and, by association, the environmental conditions (e.g., temperature and humidity) within which they developed. In Mesoamerican archeology, they have also been employed in soils to support the hypothesis of maize cultivation (Webb et al. 2004, Seinfeld et al. 2009). Nevertheless, it is important to point out that stable carbon isotope ratios obtained from off-site stratigraphic sediments do not confirm the existence of particular environmental conditions through time, nor do they indicate the cultivation of specific plants at any one period. However, they do provide a baseline for the development and support of hypotheses related to these issues, in conjunction with macro- and microbotanical remains recovered from stratigraphic deposits as well as archeological contexts. Recent investigations at Río San Pablo in the Teotihuacan Valley (Fig. 7) detected maize pollen and phytoliths together with δ13C ratios indicating a predominance of C4 vegetation in strata from a buried soil, referred to in the literature as the “black soil,” dated to the Middle-Late Formative period (Solleiro-Rebolledo et al. 2011; Sánchez-Pérez et al. 2013). These data likely indicate maize cultivation rather than semi-arid environmental conditions as δ13C ratios alone might suggest (Solleiro-Rebolledo et al. 2006).

The Teotihuacan Valley, central Mexico
Interestingly, the black soil previously mentioned and described elsewhere in the Teotihuacan Valley was preserved as part of the fill in the earliest structure underlying the Moon Pyramid (ca. AD 100, Sugiyama and Cabrera 2007). Samples also yielded δ13C values (between −15.33 and −13.79 0/00), consistent with maize cultivation (Rivera et al. 2007) even though neither maize pollen nor phytoliths were recovered from this context. Micromorphological evidence indicating human-induced alteration of the soil includes abundant microscopic charcoal and agrocutans in the clay fraction, supporting the hypothesis that nearby agricultural fields were affected as soil was removed to provide fill for monumental structures (McClung de Tapia 2012).
Furthermore, insofar as human consumption of C3, C4, and CAM plants detected from collagen is concerned, direct as well as indirect routes are discernible. That is, plants may be consumed directly or meats from herbivores or omnivores may be consumed alone or together with other plants, thus confounding the specificity of the isotope signatures in human remains. In the Teotihuacan region, an important aspect of stable carbon isotope values is the apparent overlap between C4 and CAM plants. For example, macrobotanical remains recovered from domestic contexts attest to maize and amaranth (C4), as well as nopal fruits (CAM) being significant components of the Teotihuacan diet (McClung de Tapia 1979, 1987; González-Vázquez 1986, González-Vázquez et al. 1993; see also Martínez and Manzanilla 2005, Paper presented at the Society for Economic Botany). Table 4 indicates that the δ13C values of these and other plants fall within a comparable range, pointing to the difficulty in differentiating their consumption based on stable isotope ratios alone.
Discussion
The presence of macrobotanical and microbotanical evidence for specific plants in archeological deposits, particularly activity areas, is significant as a means to validate and complement the information obtained from stable carbon isotopes.
While it is clear that stable carbon isotope values highlight the categories of foods consumed by individual residents of Teotihuacan, as well as contributing to our understanding of the geographical origins and histories of individuals found buried in the city, they do not resolve specific questions about what foods were consumed, how they were prepared, and the ways in which they were combined to structure diet. Although recent efforts to discern traces of traditional prehispanic foodways in central Mexico rely partially on sixteenth century descriptions and ethnographic accounts to develop hypotheses concerning these aspects (McClung de Tapia et al. 2014), only botanical remains from well-preserved archeological contexts can provide the information necessary to demonstrate their validity. The rapid advances in archaeometric techniques available to archeologists is significant. Additional techniques such as the recovery of food residues from artifacts, in the form of chemical signatures and microbotanical remains (for example, phytoliths and starch grains) from tools associated with food preparation and chemical residues from ceramic vessels offer potentially specific results beyond the commonly used qualitative techniques. No single technique offers a complete picture and traditional archeological field practices combined with in-depth sampling for diverse remains of plant materials referred to above is fundamental to advance questions concerning the foodways of ancient Teotihuacan. However, the recovery of macro- and microbotanical remains continues to provide a fundamental source of information concerning prehispanic plant use, as it is the only direct evidence available to archeologists at Teotihuacan in the absence of coprolites.
Unfortunately, the low quantities of identifiable plant remains recovered so far from excavations at Teotihuacan have restricted the applicability of paleoethnobotanical information beyond general hypotheses that are difficult to test. It is clearly necessary to increase the amount of evidence for plant use in order to explore the relative importance of different plants. Some attempts have focused on differentiating domesticated, cultivated and gathered plants (Martínez-Yrízar and McClung de Tapia 2009). Others, such as Garine and Vargas (2006) propose a more detailed scheme. These authors outlined a perspective for the anthropological study of foodways and nutrition in modern communities, which provides a broad framework within which to consider the kinds of questions raised by archeological evidence for food use. They provide a cultural classification of foods based on their uses, consisting of basic, primary, secondary, and marginal components. Based on the range and diversity of preparation techniques, the association of plants to ritual practices and the degree to which members of a society are familiar with specific plants.
Classifying plant remains from archeological contexts in categories such as these may help to interpret their relative importance as foods, for example, facilitating comparison among sites within regional frameworks, within periods or through time, among other possibilities. Traditions reported in the ethnographic literature may be useful as the study of food consumption in modern rural communities provides elements to organize data and construct hypotheses for the analysis of prehispanic behavior. However, these kinds of questions require substantial quantities of plant remains, both macrobotanical and microbotanical, from carefully excavated contexts. More appropriate methodologies for the recovery of botanical remains from Teotihuacan are necessary. Undoubtedly, it will be necessary to increase the volume of sediment samples recovered for flotation, although this will tax laboratory infrastructure in some instances. Although extensive excavations are vital to provide an overall understanding of activities the associated use of space, our experience suggests that stratigraphic test excavations reaching undisturbed depths may have better preservation of organic materials. Needless to say, at a complex site such as Teotihuacan, this strategy may not always be feasible. Furthermore, the difficulty of interpretation in the absence of spatial contextual information is an important limitation.
We reiterate the importance of coordinated interdisciplinary efforts to address these kinds of problems with the goal of enhancing paleoethnobotanical research at Teotihuacan and other Mesoamerican archeological sites.
References
Arnaud S M. (2014) Procedencia y Dieta de una muestra ósea de la Ventilla 92–04, Teotihuacan. Master’s thesis in Anthropology. Facultad de Filosofía y Letras/Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México
Ayerza R Jr, Coates W (2005) Chia. Rediscovering a forgotten crop of the Aztecs. University of Arizona Press, Tucson
Barba L, Ortiz A, Pecci A (2014) Los residuos químicos. Indicadores arqueológicos para entender la producción, preparación, consumo y almacenamiento de alimentos en Mesoamerica. Anales de Antropología 48(1):201–239
Barba-Pingarrón L (2007) Chemical residues in lime-plastered archaeological floors. Geoarchaeology 22(4):439–452
Barba-Pingarrón L (2009) Los residuos químicos en cerámica. Indicadores arqueológicos para entender el procesamiento de alimentos y el uso de recipientes. Memorias del XXX Congreso Internacional de americanistica, Centro Studi Americanistici Circolo Amerindiano, Perugia, pp. 721–728
Cabrera Cortés M O (2011) Craft production and socio-economic marginality: living on the periphery of urban Teotihuacan. PhD dissertation Arizona State University
Cabrera-Castro R (1996) Las excavaciones en La Ventilla. Un barrio Teotihuacano. Revista Mexicana de estudios antropológicos. Sociedad Mexicana de Antropología, Tomo XLII, Teotihuacan, pp. 5–30
Carballo D, Barba L (2014) Proyecto Arqueologico Tlajinga Teotihuacan. Segunda Temporada 2013. Informe Técnico al Instituto Nacional de Antropología e Historia, México
Casales-Gómez M, Tavera-Sánchez J (1996) Estudio arqueobotánico en pozos del frente 2 en La Ventilla, Teotihuacan. Revista Mexicana de estudios antropológicos. Sociedad Mexicana de Antropología, Tomo XLII, Teotihuacan, pp. 77–86
Correo-Ascencio M, Robertson IG, Cabrera-Cortes O, Cabrera-Castro R, Evershed PR (2014) Pulque production from fermented agave sap as a dietary supplement in prehispanic Mesoamerica. Papers of the National Academy of Science 111(39):14223–14228
Cowgill G L (2015) Ancient Teotihucan. Early urbanism in Central Mexico. Cambridge University Press
Dibble C E, Anderson A J O (1963) Florentine Codex. General history of the things of New Spain Book 11—Earthly things. Dibble, C E, Anderson, A J O (ed.) Monographs of the School of American Research and the Museum of New Mexico. Santa Fe. Number 14 Part XII
Emmart EW (1940) The Badianus manuscript (codex Barberini, Latin 241. Vatican library): an Aztec herbal of 1552. Johns Hopkins Press, Baltimore
Ford RI, Elias JE (1972) Teotihuacan paleoethnobotany. Paper presented at the 37th annual meeting of the Society for American Archaeology, Miami Florida
Garine I, Vargas LA (2006) Introducción a las investigaciones antropológicas sobre alimentación y nutrición. In: Alonzo-Padilla AL, Peña-Saint-Martin F (eds) Cambio Social, Antropología y Salud. Escuela Nacional de Antropología/Programa de Mejoramiento del Profesorado (PROMEP)/Subsecretaría de Educación Pública, México, pp 103–120
Gómez-Chávez S, Nuñez-Hernández J (1999) Análisis preliminar del patrón y la distribución espacial de entierros en el Barrio de la Ventilla. In: Manzanilla, Serrano (eds) Practicas funerarias en la ciudad de los dioses. Instituto de Investigaciones Antropológicas/Dirección General de Asuntos del Personal Académico, Universidad Nacional Autónoma de México, pp 81–147
González-Sobrino, B Z (2003) Víctimas sacrificados en el Templo de Quetzalcoatl, Teotihuacan. In: Serrano-Sánchez C (coord) Una perspectiva biocultural. Contextos arqueológicos y osteología del Barrio de La Ventilla, Teotihuacan (1992-1994). Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México, México, pp 139–148
González-Vázquez J (1986) La Identificación de Semillas Carbonizadas de Cactáceas Procedentes del Sitio Arqueológico de Tetitla, Teotihuacan, México. Unpublished thesis in Biology, Facultad de Ciencias, Universidad Nacional Autónoma de México
González-Vázquez J, Ibarra-Morales E, Zurita-Noguera J, McClung de Tapia E, Tapia-Recillas H (1993) Macrofósiles botánicos, fitolitos y polen. In: Manzanilla L (ed) Anatomía de un conjunto residencial teotihuacano en Oztoyahualoco. II Los Estudios Específicos, Instituto de Investigaciones Antropológicas, UNAM, pp. 661–728
Guralnick LJ, Cline A, Smith M, Sage RF (2008) Evolutionary physiology: the extent of C4 and CAM photosynthesis in the genera Anacampseros and Grahamia of the Portulacaceae. J Exp Bot 58(7):1735–1742
Hernández F (1942) Historia de las plantas de Nueva España. Instituto de Biología, Universidad Nacional Autónoma de México, tomo I, II, III. Imprenta Universitaria, UNAM
Hurst WJ (2006) The determination of cacao in samples of archaeological interest. In: McNeil CL (ed) Chocolate in Mesoamerica, a cultural history of cacao. University of Florida Press, Gainesville, pp. 105–113
Lounejeva-Baturina E, Morales-Puente P, Cabadas-Báez HV, Cienfuegos-Alvarado E, Sedov S, Vallejo-Gómez E, Solleiro-Rebolledo E (2006) Late Pleistocene to Holocene environmental changes from δ13C determinations in soils at Teotihuacan, México. Geofísica Internacional 45(2):85–98
Manzanilla L (1993) Anatomía de un conjunto residencial teotihuacano en Oztoyahualoco I, Las excavaciones. Instituto de Investigaciones Antropológicas, UNAM
Manzanilla L (2012) Introducción. In: Estudios Arqueométricos del centro de barrio de Teopancazco en Teotihuacan, Manzanilla (ed) Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México. pp. 17–68
Manzanilla L (2015) Cooperation and tensions in multiethnic corporate societies using Teotihuacan, Central Mexico as a case study. Papers of the National Academy of Science. www.pnas.org/cgi/doi/10.1073/pnas.1419881112
Manzanilla L, Millones M, Civera M (1999a) Los entierros de Oztoyahualco. In: Manzanilla, Serrano (ed) Practicas funerarias en la ciudad de los dioses. IIA/DGAPA, UNAM, pp. 247–283
Manzanilla L, Tejeda S, Martínez JC (1999b) Implicaciones del análisis de calcio, estroncio y Zinc en el conocimiento de la dieta y la migración en Teotihuacan, México. Anales de Antropología 33:13–28
Martínez-Yrízar D, McClung de Tapia E (2009) Las plantas como recursos en Santa Cruz Atizapan, EN Y. Sugiura (Coord.). La gente de la ciénaga en tiempos antiguos: la historia de Santa Cruz Atizapan. IIA-UNAM/Colegio Mexiquense, pp 175–193
McClung de Tapia E (1977) Recientes estudios paleo-etnobotánicos en Teotihuacan, México. Anales de Antropología, XIV, IIA, UNAM. pp. 49–61
McClung de Tapia E (1978) Aspectos ecológicos del desarrollo y la decadencia de Teotihuacan. Anales de Antropología, XV, IIA, UNAM, pp. 53–65
McClung de Tapia E (1979) Plants and subsistence in the Teotihuacan Valley A.D. 100–700, Ph. D. Dissertation, Faculty of the Graduate School of Arts and Sciences Brandeis University, Department of Anthropology
McClung de Tapia E (1980) Interpretación de restos botánicos procedentes de sitios arqueológicos, Anales de Antropología, XVIII(I). IIA, UNAM, pp. 149–166
McClung de Tapia E (1987) Patrones de subsistencia urbana en Teotihuacan. In: McClung de Tapia E, Rattray E C (eds) Teotihucan: Nuevos Datos, Nuevas Síntesis, Nuevos Problemas. Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México, pp. 57–74
McClung de Tapia E (2012) Silent Hazards – Invisible Risks: Prehispanic Erosion in the Teotihuacan Valley, Central Mexico. In: Cooper J, Sheets P (eds) Living with the Dangers of Sudden Environmental Change: Understanding Hazards, Mitigating Impacts, Avoiding Disasters. University Press of Colorado, pp. 139–161
McClung de Tapia E, Adriano-Morán CC (2012) Stable carbon isotopes applied to landscape reconstruction in the Teotihuacan Valley, Mexico. Bol Soc Geol Mex 64(2):161–169
McClung de Tapia E, Martínez-Yrízar D, Ibarra-Morales E, Adriano-Morán CC (2014) Los orígenes prehispánicos de una tradición alimentaria en la Cuenca de México. Anales de Antropología 48(1):97–121
Meza-Peñaloza A (2008) Estudio de morfometría geométrica de sinus frontales de los antiguos teotihuacanos: afinidades biológicas y contextos culturales. PhD thesis. Posgrado de Estudios Mesoamericanos, Universidad Nacional Autónoma de México, México, D. F.
Millon R (1973) Urbanization at Teotihuacan: the Teotihuacan map. Part one: text, part two: maps. Texas University Press, Austin
Millon R (1988) The last years of Teotihuacan dominance. In: Yoffee N, Cowgill GL (eds) The collapse of ancient states and civilizations. University of Arizona Press, Tucson, pp. 102–164
Minnis P (1981) Seeds in archaeological sites: sources and some interpretive problems. Am Antiq 46:143–152
Montufar A (1996) Estudio arqueobotánico en vasijas de La Ventilla, Teotihuacan, México, Revista Mexicana de estudios antropológicos. Sociedad Mexicana de Antropología, Tomo XLII, Teotihuacan, pp. 71–76
Morales P, Cienfuegos E, Manzanilla L, Otero F (2012) Estudio de la paleodieta empleando isótopos estables de los elementos carbono, oxígeno y nitrógeno en restos humanos y de fauna encontrados en el barrio Teotihuacano de Teopancazco. In: Manzanilla L (ed) Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México, pp. 347–424
O’Connor T (2000) The Archaeology of Animal Bones. Sutton Publishing
Ochoa-Ocaña M A (2003) Dieta y estatus. Estudio comparativo de paleonutrición en Teotihuacan. In: Serrano-Sánchez C (coord.) Contextos arqueológicos y osteología del Barrio de la Ventilla, Teotihuacan (1992–1994). Instituto de Investigaciones antropológicas, Universidad Nacional Autónoma de México. México pp 123–128.
Pérez-Lizaur AB, Marván-Laborde L (2001) Sistema Mexicana de Alimentos Equivalentes. Fomento de Nutrición y Salud, A. C, México
Powis TG, Valdez F Jr, Hester TR, Hurst JW, Tarka SM (2002) Spouted vessels and cacao use among the preclassic Maya. Lat Am Antiq 13(1):85–106
Rattray E C (1997) Entierros y Ofrendas en Teotihuacan. Excavaciones, Inventario, patrones mortuorios. Instituto de Investigaciones antropológicas, Universidad Nacional Autónoma de México. México
Reitz E J, Wing E S (2001) Zooarchaeology. Cambridge Manuals in Archaeology. Cambridge University Press
Rivera M, Sedov S, Solleiro-Rebolledo E, Pérez-Pérez J, McClung E, González A, Gama-Castro J (2007) Degradación ambiental en el Valle Teotihuacan: evidencias geológicas y paleopedológicas. Boletin de la Sociedad Geológica Mexicana 59(2):203–217
Rojas-Lago R, Serrano-Sánchez C (2011) Paleopatología de Teotihuacan. Teotihuacan, cosmogonía y medicina. Vida + Salud. Biblioteca Mexiquense del Bicentenario, Gobierno del Estado de México, Toluca, pp 100–135
Sánchez-Pérez S, Solleiro-Rebolledo E, Sedov S, McClung de Tapia E, Golyeva A, Prado B, Ibarra-Morales E (2013) The Black San Pablo paleosol of the Teotihuacan Valley, Mexico: pedogenesis, aptitude, utilization in ancient agricultural and urban systems. Geoarchaeology 28(3):249–267
Sanders WT, Parsons JR, Santley RS (1979) The basin of Mexico. Ecological processes in the evolution of a civilization. Academic Press, Orlando
Seinfeld DM, von Nagy C, Pohl MD (2009) Determining Olmec maize through bulk stable carbon isotope analysis. J Archaeol Sci 36:2560–2565
Sejourné L (1959) Un palacio en la ciudad de los dioses. Instituto Nacional de Antropología e Historia, México
Serrano C, Lagunas Z (1999) Prácticas mortuorias prehispánicas en un barrio de artesanos La Ventilla B Teotihuacan. In: Manzanilla L, Serrano-Sánchez (eds) Practicas funerarias en la ciudad de los dioses. IIA/DGAPA, UNAM, pp. 35–80
Solleiro-Rebolledo E, Sedov S, McClung de Tapia E, Cabadas H, Gama-Castro J, Vallejo-Gómez E (2006) Spatial variability of environment change in the Teotihuacan Valley during late quaternary: paloepedological inferences. Quaternary Internacional 156-157:13–31
Solleiro-Rebolledo E, Sycheva S, Sedov S, McClung de Tapia E, Rivera Y, Salcedo-Berkovich C (2011) Fluvial responses and paleopoedgenesis in the Teotihuacan Valley, Mexico. Responses to the late quaternary landscape dynamics. Quat Int 233:40–52
Somerville A, Fauvelle M, Froehle A (2013) Applying new approaches to modeling diet and status: isotopic evidence for commoner resiliency and elite variability in the classic maya lowlands. J Archaeol Sci 40:1539–1553
Storey R (1992) Life and death in the ancient city of Teotihuacan, a modern paleodemographic synthesis. The University of Alabama Press
Sugiyama S, Cabrera R (2007) The moon pyramid project and the Teotihuacan state polity. Anc Mesoam 18(1):109–125
Webb EA, Schwarcz HP, Healy PF (2004) Detection of ancient maize in lowland Maya soils using stable carbon isotopes: evidence from Caracol, Belize. J Archaeol Sci 31:1039–1052
Widmer R (1987) The evolution of form and function in a Teotihuacan apartment compound. In: McClung de Tapia E, Rattray E (eds) Teotihuacan. Nuevos datos, Nuevas síntesis, Nuevos problemas. Instituto de Investigaciones Antropológicas, Universidad Nacional Autónoma de México, pp. 317–368
Widmer R J, Storey R (1992) Social Organization And Household Structure Of A Teotihuacan Apartment Compound: S3 W1:33 Of The Tlajinga Barrio. In: Robert S S, Hirth K G (eds) Prehispánica Doméstic Units In Western Mesoamerica, Saudíes Of The Household, Compound And Residence. CRC Press, pp. 87–104
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
McClung de Tapia, E., Martínez-Yrizar, D. The potential of paleoethnobotanical evidence for the study of Teotihuacan foodways. Archaeol Anthropol Sci 9, 39–50 (2017). https://doi.org/10.1007/s12520-016-0414-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-016-0414-0