
Abstract
Recent archaeobotanical research on 16 archaeological sites in the Sierras de Córdoba, central Argentina, provides new insights into the livelihoods and subsistence practices of the peoples who inhabited this mountainous region from c. 3,000–250 bp. Significantly, the plant macro- and microbotanical remains, identified as primarily fruit from wild trees, crops and weeds, provide evidence for a continuation in the consumption and manipulation of plant resources. During the late pre-Hispanic period (1,500–350 bp) people used domesticated plants such as maize, as well as new types of plant processing techniques that permitted the consumption of otherwise inedible wild seeds such as chenopods. The introduction of cultivated plants through contact with agricultural societies at around 1,000 bp was slow and did not substantially change the existing foraging way of life. Instead, crop plants were added to the existing, highly diversified subsistence systems in use in the Sierras de Córdoba, rather than replacing wild plant gathering.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Traditional archaeological accounts of the people who inhabited the Sierras de Córdoba from the early Late Holocene (3,000 bp) to colonial times (250 bp) have presented a simplistic view of plant consumption practices of both hunter-gatherer and farming societies. Archaeologists proposed that forager subsistence included only fruits of wild trees; in this scenario, the emergence of agriculture at around 1,000 bp resulted in an immediate change to a reliance on domesticated plants, and it was within this agricultural landscape that sedentary village life began (Gonzalez 1943; Serrano 1945; Berberián 1984; Laguens 1999; Berberián and Roldan 2001, 2003, among others). This view was based on indirect evidence including grinding tools, cultivating implements such as hoes, pottery and historical documents. Thereby, ancient human life in the Sierras de Córdoba was interpreted as: (1) human forager groups with high mobility patterns and consumption of items such as large game, Lama guanicoide Müller (guanaco), and secondarily eggs of Rhea sp. Brisson (rhea) and plants such as Prosopis sp. L. (algarrobo) and Geoffroea decorticans (Gill. ex Hook. & Arn.) Burkart (chañar); (2) farming societies with a sedentary way of life in pit house villages which were occupied all year round, and produced resources, mainly Zea mays L. (maize), near where they lived. Farming was thought to have been immediately adopted and with high levels of production, in tandem with cultural changes that were necessary for adopting the new mode of economic organization.
Recent archaeobotanical studies of 16 mainly single component archaeological sites in the Sierras de Córdoba provide the basis for new interpretations of prehistoric subsistence practices that show the importance of plants in the ancient economies of central Argentina. New data show that human occupation here during the Pleistocene-Holocene transition (c. 12,000–9,000 bp) was not intensive, with a low visibility archaeological record. Only three types of evidence are known from this period: stone artefacts (fish-tail projectile points), extinct megafaunal remains and human bones (Rivero et al. 2015). Botanical remains recovered from the El Alto 3 site were identified as the wood of Polylepis australis Bitter and Maytenus boaria Molina (López 2015). Although economic use of plants was evident, food remains were not identified from this early time. After c. 8,000 bp when modern environmental conditions began to be established, people could colonize the Sierras de Córdoba, leading to settlement of this mountainous region. They inhabited open air and cave sites and much material has been recovered, such as Ayampitin lanceolate or foliate projectile points, bone artefacts and grinding tools. While faunal remains were retrieved and identified as Lama sp. (camelids), Mazama sp. Rafinesque and Ozotoceros beozoarticus Linnaeus (cervids, deer) and small vertebrates including Cricetidae, Caviinae, etc. (Rivero and Berberián 2011; Rivero and Medina 2013), edible plants have only been recovered from sites dating from 3,000 bp and later, when important changes were observed in the valleys. From up to 50 large open air sites, triangular projectile points, grinding tools and numerous burials show the intensity of occupation in the Sierras de Córdoba; the introduction of agriculture appears to have been a slow process, evident only in late contexts (c. 1,100 bp). Small-scale food production would have been involved, complementing a sustained process of intensification of hunting and gathering. The incorporation of innovations such as pottery, new types of projectile points and the growing of crops, involved gradual changes to traditional ways of life (Berberian et al. 2008). “The social and political systems in the Sierras de Córdoba were built on flexible subsistence and mobility patterns, where strategies were changed according to the availability of resources and the social environment” (Medina et al. 2016, p 102).
The aim of this paper is to present the results of the archaeobotanical study of open air and cave sites. Macro and microbotanical remains from the early Late Holocene have been studied in order to investigate continuities and change in the consumption and manipulation of plant resources from 3,000 bp through to the first years of Spanish colonization. During the last 15 years, archaeobotanical analyses have focused on new questions related to changes in the systems of sociopolitics, economics and landscape use.
The following methodologies for the recovery of both macro- and microbotanical plant remains were used. For seeds and fruits, a systematic sampling, followed by fine sieving with 2–1 mm meshes was carried out to identify human activities, especially food production and consumption. Microscopic remains such as phytoliths and starch grains were recovered from stone tools, pottery and dental tartar as well as from sediment samples. Standard laboratory methods were applied to remove microremains from artefacts without the use of heavy liquid, using the point of a fine needle; residues from the artefacts were then mounted on glass slides using immersion oil (Pearsall 2000; Piperno 2006).
Archaeological sites throughout the Sierras de Córdoba, central Argentina, have similar environmental conditions. This low altitude mountain range consists of peaks, valleys and plateaus, ranging from 500 to 2,800 m a.s.l. Known geologically as the Sierras Pampeanas Orientales, they can be divided into a northern area that covers the Sierra de Norte and a central-western area that includes the Sierras Chicas, Sierras Grandes and Sierra de Pocho-Guasayan-Serrezuela. The Sierra de Norte is considered a small part of the Sierras Grandes, which make up the central body of the Sierras Pampeanas Orientales (Cioccale 1999). The valleys in Sierra Chaco are comprised of a semi-arid woodland dominated by trees and shrubs with edible wild fruits such as Geoffroea decorticans (chañar), Prosopis alba Griseb. and P. nigra Griseb. Hieron. (algarrobo), Sarcomphalus mistol (Griseb.) Hauenschild (mistol), Lithraea molloides (Vell.) Engl. (molle de beber) and Condalia buxifolia Reissek, C. microphyla Cav. and C. montana A. Cast. (piquillín), among others. These were prime areas for farming, with slopes around them. The modern fauna is dominated by low-diversity neotropical small mammals such as the Mazama gouazoubira Fisher (brown-brocket deer), Pecari tajacu Linnaeus (collared peccary), Lagostomus maximus Desmarest (vizcacha), Dasypodidae (armadillos) and Caviinae and Ctenomyinae (small caviomorph rodents including cavies and guinea pigs). Above 1,500 m a.s.l., the vegetation is a mosaic of Polylepis australis woods and upper mountain grasslands with mainly Festuca hieronymi Hack., Deyeuxia hieronymi (Hack.) Türpe, Sorghastrum pellitum (Hack.) Parodi and Poa stuckertii (Hack.) Parodi. The wild fauna there is characterized by Puma concolor Linnaeus (puma) and Vultur gryphus Linnaeus (condor). In the past, it supported wild ungulate herds such as Lama guanicoe (guanaco), Ozotoceros bezoarticus (pampas deer) and Rhea americana Linnaeus (rhea) (Giorgis et al. 2011; Poca et al. 2014; Grau et al. 2015).
The earliest archaeobotanical data from the Sierras de Córdoba, central Argentina
The earliest evidence of plant processing and consumption has been found at two sites, the Quebrada del Real 1, a cave in the Pampa de Achala, and Cruz Chiquita 3, an open air settlement in Valle de Traslasierra (Fig. 1; Table 1). Quebrada del Real 1 had a lengthy occupation period, from c. 7,400 bp to the final Late Holocene (c. 1,000–500 bp). Only the middle layers, Component 2 (C2), are discussed here because of their rich archaeobotanical record. The C2 archaeological assemblage, dated at 2,950 ± 90 bp (LP-2042; charcoal), was characterized by triangular projectile points with slightly to markedly concave bases and no pottery, resembling other Middle Holocene and early Late Holocene deposits (Rivero 2009). Seven manos (hand mills) were recovered from these layers, which appear to have been used for processing seeds and other plant parts. The phytolith and starch evidence shows that people processed chenopod seeds and maize grains at this site (Pastor et al. 2012; López et al. 2015).

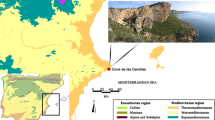
Locations of the Sierras de Córdoba (central Argentina), the archaeological areas in it and the archaeological sites mentioned in this paper
The identification of starch grains resembling Chenopodium spp. from grinding tools in QR1 suggests that chenopods have been used as food since c. 3,000 bp in central Argentina (Fig. 2). The most probable taxa would be wild ones such as Chenopodium hircinum Schrad. or C. (=Dysphania) ambrosioide (L.) Mosyakin & Clemants, as Component 2 represents early Late Holocene hunter-gatherer occupations (López et al. 2015). The presence of these annual camp-following plants, which are high in edible biomass but which require extensive processing to consume them, suggest intensification of plant use (Aldenderfer 1998; Kuznar 2001). Similar data from the early Late Holocene in central Chile (c. 3,300 bp) have been interpreted as evidence of greater use of plants that led to the domestication of Chenopodium in that Andean region (Planella and Tagle 2004; Planella et al. 2005, 2011). The compounded characteristics of the starch grains suggest that the hand stones were not only used to grind Chenopodium spp. seeds. The starch aggregation suggests seeds which had been rather gently pounded with hand-stones, mainly to remove the pericarps and their steroid content, which gives them a bitter taste and a toxic effect from the saponin which they contain (Fonturbel 2003; López 2012). Thus, these data not only suggest an early knowledge of plants, but also that people had the technology to remove the inedible portions of the seeds (López et al. 2011, 2015).

Identified microbotanical remains. a, b cf. Chenopodium spp. starch grains from QR1; c, d, cf. Solanum cf. tuberosum starch grain from Pz1; e, f cf. Oxalis sp. starch grain from BP2; g, cf. Zea mays cob phytolith from PE1; h cf. Cucurbita sp. fruit phytolith from ATC1; i cf. Zea mays leaf phytoliths from ATC1; j cf. Phaseolus sp. leaf phytolith from ATC1; a, c, e optical microscope with a direct light source; b, d, f with polarized light. Scale bar 20 µm
Excavations at the open air Cruz Chiquita 3 site revealed a simple burial without a well-defined border, which was covered with a layer of stones. Inside were the remains of an adult male individual in a flexed position without any adornments or accompanying objects. This was AMS radiocarbon dated to 2,466 ± 51 bp (AA68146, from bone collagen). Phytolith analysis of microfossils in this individual’s dental tartar indicated consumption possibly of Zea mays (Pastor et al. 2012). Such early evidence of processed and consumed maize is important for understanding the subsistence system in central Argentina and the Andean region. It contributes to a wider discussion of the process of adoption of cultivated plants through interactions between hunter-gatherer and farming populations. Moreover, it suggests that the expansion of farming was accompanied by flexible and variable patterns of development and a slow and gradual transformation of the diverse local societies. While current information is insufficient to determine the importance of maize in the diet at this time and place, there is presently no indication that it was great. However, information available at the macro-regional and sub-continental level (Gil 1997–1998; Staller 2007; Nuñez et al. 2009; Babot 2011) suggests consideration of other factors, for example, the ritual significance of this exotic resource in numerous contexts (Pastor et al. 2012).
Plant evidence from pre-Hispanic contexts (1,500–350 bp)
Abundant evidence of cultivation, gathering and harvesting, processing and cooking of plants in central Argentina appears in late pre-Hispanic contexts, c. 1,500–350 bp. This period is characterized by innovations such as containers and the growing of crops, as well as flexible patterns of subsistence, where seasonal mobility played a key role. This mobility allowed people to take advantage of different environments around the year. In this context, agriculture that was developed on a small scale and without larger works, such as irrigation canals or cultivation terraces, was not essential to the life of the pre-Hispanic communities (Pastor and López 2011; Medina et al. 2016).
Several of the 16 sites discussed here span the period from 1,500 to 350 bp (Table 1; Fig. 1). Carbonized macrobotanical remains were identified, on the one hand, fruits of wild trees, Sarcomphalus mistol (mistol), Lithraea molloides (molle de beber), Condalia sp. (piquillín), Prosopis sp. (algarrobo), Geoffroea decorticans (chañar) and Schinus sp. cf. areira L. (aguaribay); on the other hand, there were also seeds of crops and weeds such as Zea mays (maize), Phaseolus vulgaris var. vulgaris L. and P. lunatus L. (beans), Phaseolus vulgaris var. aborigineus (Burkart) Baudet (wild bean), Chenopodium quinoa var. quinoa (quinoa) and C. quinoa Willd. cf. var. melanospermum Hunz. (quinoa negra) (Fig. 3). The microfossil analysis supports the macrofossil evidence for crops and further indicates the presence of wild underground storage organs (USOs) of plants in late contexts. Starch grains similar to those of Solanum sp. cf. tuberosum L. (potato), phytoliths of maize (grain) and Cucurbita L. sp. (cucurbit, fruit) were recovered from pottery and grinding tools. Starch grains similar to Oxalis L. sp. (oca, yam) were recovered from a bone tool with a notched working edge. This genus is an endemic plant in the Andean region and includes wild taxa with edible storage organs in Argentina such as O. conorrhiza Jacq. and O. lasiopetala Zucc. (Giorgis et al. 2011). Finally, phytoliths from maize and bean leaves were recovered from sediments in ancient cultivation plots (Fig. 2; Pastor and López 2011; López 2015; López and Recalde 2016).

Identified macrobotanical remains. a Sarcomphalus mistol fruits from QN7; b Lithraea molloides fruits and endocarps from QN7; c Condalia sp. endocarps from QN7; d Prosopis sp. seeds and endocarps from QN7; e Geoffroea decorticans endocarp from QN7; f Schinus cf. areira seeds from QN7; g Zea mays grain from C.Pun.39; h Phaseolus lunatus cotyledons from ATC1; i Phaseolus vulgaris cotyledons from ATC1; j Chenopodium quinoa var. quinoa seed from QN7. Scale bar 10 mm
Taken together, archaeobotanical and archaeological evidence of agricultural practices, such as stone artefacts for digging, indicates that plant cultivation was adopted around c. 1,500 bp. Nevertheless, no dramatic changes in the settlement and subsistence patterns are evident, but rather, subsistence generally followed the pre-existing early Late Holocene foraging patterns. Thus, the transition to growing crops was a slow process in which foragers increasingly took up horticulture to support their already diversified economies. Cultivation fitted into the pre-established foraging pattern, and its adoption was an opportunistic effort to minimize seasonally occurring shortfalls of food, rather than a means to a new subsistence system.
The fruits of wild trees were important staple resources and the diversity of taxa shows that people manipulated and consumed all plants that the environment had to offer. Mistol fruits were used in the whole Sierras de Córdoba and their remains offer the first evidence of food storage. Current mistol consumption now only includes arrope (syrup) and bolanchao (balls of ground fruits) but it was likely that a multipurpose food resource such as the dried fruit could possibly also have been used to make flour. The same may be the case for the fruit of molle de beber, which is not consumed nowadays (Arias Toledo 2009).
The low frequencies of Prosopis (algarrobo) fruits at these sites remain a question, as various archaeobotanical and historical records point to the importance of this tree fruit in the past (see above). Algarrobo has been recovered from numerous archaeological sites in north-west Argentina (Oliszewski 1999; Giovannetti et al. 2008; Capparelli and Lema 2011). The fact that it is also abundant now in the Sierra Chaco woods in the Sierras de Córdoba in central Argentina has led to the view that Prosopis was also an important food resource in this region. These assumptions have been strengthened by recent ethnobotanical studies in northwest, northeast and central-west Argentina which show the importance of algarrobo as a traditional food and in which periods (Capparelli 2007; Sciammaro 2015). Spanish documents from the early colonial period in the 16th-17th centuries ad also have numerous references to the consumption and storage of algarrobo and its importance to the indigenous people (Berberián 1987; Castro Olañeta 2004).
However, the role of Prosopis may have been different in central Argentina in the past. Although a highly useful resource that could be processed into a range of edible products (Capparelli 2007), archaeological remains of algarrobo fruit are not only scarce, but several other factors also need to be considered, beginning with the ecology of Prosopis. The abundance of algarrobo in central Argentina can probably be explained by its colonization of the Sierra Chaco after the loss of Aspidosperma quebracho blanco Schltdl. (quebracho blanco), which was formerly the main native non-edible species in the Sierra Chaco. Quebracho blanco was severely reduced by fire and livestock (mainly European) permitting the invasion of Prosopis (Morello and Saravia 1959). Algarrobo trees are thus more abundant in this region today than they were in the past. Also, environmental conditions such as moisture and temperature influence the fruiting of algarrobo from year to year and from tree to tree (Villagra et al. 2004), and therefore the availability of fruit. Ethnobotanical studies show that algarrobo fruits are used as fodder by modern communities as well as having had a significant role in folklore (Arias Toledo 2009). On the other hand, colonial records contain numerous inaccuracies, because Spaniards erroneously viewed the indigenous people in the Sierras de Córdoba as being similar to those in northwest Argentina. Although the colonial documents can provide important evidence about the food consumption practices of pre-Hispanic people, the archaeobotanical evidence does not support many of their assumptions. Finally, the Sierras de Córdoba continued to be treated as a marginal area into the 20th century and archaeologists likewise treated the area as being similar to north-west Argentina, with the same features such as an agricultural economy and the preponderance of algarrobo among the wild fruits.
The late pre-Hispanic people moved around the landscape to take advantage of seasonal availabilities of various wild resources as well as crops adapted to different micro-environments. Cultivation and foraging were both important elements of the ancient subsistence economy, so that the remains of settlements and small sites are scattered across the mountainous landscape (Medina et al. 2016). The proposed model of foraging together with small-scale farming recognizes a varied mosaic of foraging and farming patterns that did not involve all the people in the region in the same way, during the long-lasting late pre-Hispanic period (Medina et al. 2016). Archaeobotanical analyses have revealed differences and similarities between two well defined areas: the northern and the central-western parts of Provincia de Córdoba (Fig. 1; Table 2). Although different numbers of sites were analyzed in each area, the variations in the finds of remains of wild and cultivated plants indicate corresponding variations in the dynamics of using different parts of the landscape, as well as different patterns of mobility around it. Although subsistence strategies were similar in both regions, consisting mainly of agricultural practices, these led to the development of different landscape types as a result of the different ways of handling both wild and domesticated plants.
In the northern area, the richness of macrobotanical remains according to the number of taxa was higher, but mistol and molle were the most abundant, while in the central-western area, taxon richness was lower and mistol predominated. On the other hand, maize, a few beans and quinoa were the primary crops in the north, while a little maize and abundant Phaseolus vulgaris and P. lunatus beans dominated the assemblage from the central-west. Weed seeds were present in both regions and could be part of the subsistence systems, but quinoa negra was only found in the north and wild bean only in the central-west, and the bean could also grow outside of cultivated plots. Microbotanical remains of crops showed differences too. Even if USOs were processed in both areas, potato was recorded in the northern area where it was processed and cooked. Cucurbita (squash) was only processed and cooked in the central-western area.
Archaeological evidence and archaeobotanical data in particular, together with depositional patterns, if taphonomic biases are similar, allow us to assume that societies at around 1,500 years bp had settlement systems with different aspects of mobility related to, for example, frequent moves, by some people or all of them, and a degree of re-use of sites from year to year. The low density of macrobotanical remains from the central-western area probably shows short occupations with a low discard of waste as the result of maximizing the consumption of vegetables as scarce resources. On the contrary, in the northern area, the high density of macrobotanical remains presumably shows site occupation for a longer period of time and/or most people living at the site, at which tree fruits were stored and there was a different relationship between the people and the land, where crops with different growth cycles were being grown.
The first evidence of plant use during early colonial times (350–250 bp)
The single archaeological site in the Sierras de Córdoba from colonial times is Alero Tala Huasi (Fig. 1; Table 1). This medium-size 4 ha open-air site is composed of a grinding tool area and a cave, which are near a small stream known as Arroyo Seco. Microbotanical analyses of residues from pottery produced phytoliths of cf. Zea mays (maize, grains) and no evidence of European plants. Although European livestock was important here, the consumption of maize persisted. The pottery types of storage vessels and pucos (serving dishes) showed the same pre-Hispanic patterns in their design elements. Also, no evidence of glass, metal or faience was recovered (Pastor and Medina 2013).
These authors proposed that at this site, people intentionally continued their ancient customs as a way of maintaining their own traditions and to resist Spanish culture. Resistance against the colonial regime was also expressed by the theft of Spanish property, particularly livestock. Questions remain about the significance of maize, such as whether this native food was part of the indigenous resistance to Spanish culture, or was it in fact part of a cross-cultural exchange (sensu Dietler 2007) in which European livestock was adopted through an “indigenization” of exotic food? Or, did both maize and European livestock take on different roles in the new context?
Maize, along with other pre-Hispanic resources, shows the continuity of indigenous food consumption practices during the early years of colonial times. The incorporation of European livestock could represent an indigenous effort to demonstrate their power to the Spanish authorities, by appropriating resources that were otherwise denied to them. However, in the long term, these resistance tactics were unsuccessful (Pastor and Medina 2013). It may otherwise be that the “indigenization” of exotic resources happened by incorporating them into “a routinized set of practices without altering the perception of continuity” (Dietler 2007, p 224); in this way, European resources were adopted if they could be used to reproduce traditional food products (beyond particular ingredients) and therefore assume a cultural significance (López 2015). More analyses are necessary to find out more about the role of plants in colonial times, but the evidence presented here provides issues to be discussed in the future.
Conclusions
Recent archaeobotanical studies of several sites in central Argentina have offered a new view about the processing and consumption of various plant products including wild fruit trees, weed seeds, tubers and crops. Despite recent progress in the archaeobotany of this otherwise understudied region, the results have raised new questions that go beyond the presence or absence of plant taxa, among them: did post-harvest processing practices change over time? Did post-harvest processing differ between the northern and central-western areas? Were there cultural differences in plant selection and consumption in these two areas? Excavations of new sites, focusing on archaeobotanical sampling and recovery would help to answer these questions and improve our interpretation of the subsistence systems of indigenous people in central Argentina.
References
Aldenderfer M (1998) Montane foragers. Asana and the South-Central Andean Archaic. University of Iowa Press, Iowa City
Arias Toledo B (2009) Uso de plantas alimenticias y medicinales en Córdoba. Fitogeografía y cultura. Tesis para optar al grado de doctor en ciencias biológicas. Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de Córdoba
Babot MP (2011) Cazadores-recolectores de los andes centrosur y procesamiento vegetal. Una discusion desde la puna meridional argentina (c. 7,000–3,200 anos A.P.). Chungara 43(Numero Especial 1):413–432
Berberian E, Pastor S, Rivero D, Medina M, Recalde A, Lopez L, Roldán F (2008) Últimos avances de la investigación arqueológica en las Sierras de Córdoba. Comechingonia 11:143–174
Berberián E (1984) Potrero de Garay: Una entidad sociocultural tardía de la región serrana de la Provincia de Córdoba (Rep. Argentina). Comechingonia 4:71–138
Berberián E (1987) Crónicas del Tucumán. Siglo XVI. Comechingonia, Córdoba
Berberián E, Roldán F (2001) Arqueología de las Sierras Centrales. In: Berberián E, Nielsen A (eds) Historia Argentina Prehispánica. Editorial Brujas, Córdoba, pp 635–691
Berberián E, Roldán F (2003) Limitaciones a la producción agrícola, estrategias de manejo de terrenos cultivables y ampliación de la dieta en comunidades formativas de la región serrana de la provincia de Córdoba. Relaciones de la Sociedad Argentina de Antropología 28:117–131
Capparelli A (2007) Los productos alimenticios derivados de Prosopis chilensis (Mol.) Stuntz y P. flexuosa DC., Fabaceae, en la vida cotidiana de los habitantes del NOA y su paralelismo con el algarrobo europeo. Kurtziana 33(1) (Volumen especial de Etnobotánica):1–19
Capparelli A, Lema V (2011) Recognition of post-harvest processing of algarrobo (Prosopis spp.) as food from two sites of Northwestern Argentina: an ethnobotanical and experimental approach for desiccated macroremains. J Archaeol Anthropol Sci 3:71–92
Castro Olañeta I (2004) Transformaciones y continuidades de sociedades indígenas en el sistema colonial. El pueblo de indio de Quilino a principios del s. XVII. Alción Editora, Córdoba
Cioccale M (1999) Investigación geomorfológica de cuencas serranas. Estudio geomorfológico integral: morfodinámica, morfometría y morfogpenesis del flanco oriental de las Sierras Chicas de Córdoba. Doctoral Thesis. Facultad de Ciencias Exactas, Físicas y Naturales. Universidad Nacional de Córdoba
Dietler M (2007) Culinary encounters: food, identity, and colonialism. In: Twiss K (ed) The archaeology of food and identity (Center for Archaeological Investigations, Occasional Paper 34). Board of Trustees. South Illinois University, Illinois, pp 218–242
Fonturbel F (2003) Problematica de la produccion y comercialización de Chenopodium quinoa W. (Chenopodiaceae), debida a la presencia de saponinas. Revista Digital Ciencia Abierta 21:1–10
Gil A (1997–1998) Cultígenos prehispánicos en el sur de Mendoza. Discusión en torno al límite meridional de la agricultura andina. Relaciones de la Sociedad Argentina de Antropología 22–23:295–318
Giorgis M, Cingolani A, Chiarini F et al. (2011) Composición florística del Bosque Chaqueño Serrano de la provincia de Córdoba, Argentina. Kurtziana 36:9–43
Giovannetti M, Lema V, Bartoli C, Capparelli A (2008) Starch grain characterization of Prosopis chilensis (Mol.) Stuntz and P. flexuosa DC, and the analysis of their archaeological remains in Andean South America. J Archaeol Sci 35:2,973–2,985
González A (1943) Arqueología del yacimiento indígena de Villa Rumipal (Pcia. de Córdoba). (Publicaciones del Instituto de Arqueología, Lingüística y Folklore 4) Universidad Nacional de Córdoba, Córdoba
Grau R, Torres R, Gasparri I, Blendinger P, Marinaro S, Macchi L (2015) Natural grasslands in the Chaco. A neglected ecosystem under threat by agricultural expansion and forest-oriented conservation. J Arid Environ 123:40–46
Kuznar L (2001) Ecological mutualism in Navajo corrals: Implications for Navajo environmental perceptions and human/plant coevolution. J Anthropol Res 57:17–39
Laguens A (1999) Arqueología del contacto Hispano-Indígena: Un estudio de cambios y continuidades en las Sierras Centrales de Argentina (BAR Int Ser 801). Archaeo, Oxford
López ML (2012) Estudio de macro y micro restos de quinoa de contextos arqueológicos del último milenio en dos regiones circumpuneñas. Doctoral thesis, Universidad Nacional de Córdoba
López ML (2015) La cocina como medio para la reproducción social de los grupos prehispánicos de las sierras de Córdoba. In: Salazar J (ed) Condiciones de posibilidad de la reproducción social en sociedades prehispánicas y coloniales tempranas en las Sierras Pampeana (República Argentina). CEH Prof. Carlos S.A. Segreti, Córdoba, pp 177–212
López ML, Recalde MA (2016) First quinoa (Chenopodium quinoa Willd) botanical macro-remains at Sierras del Norte (Central Argentina) and its implications in pre-Hispanic subsistence practices. J Archaeol Sci Rep 8:426–433
López ML, Capparelli A, Nielsen A (2011) Traditional postharvest processing to make quinoa grains (Chenopodium quinoa var. quinoa) apt for consumption in Nortern Lipez (Potosi, Bolivia): Ethnoarchaeological and archaeobotanical analyses. J Anthropol Archaeol Sci 3:49–70
López ML, Medina M, Rivero D (2015) First records of Chenopodium spp./Amaranthus spp. starch grains and their relevance to the study of the Late Holocene human subsistence in Central Argentina. Holocene 25:288–295
Medina M, Pastor S, Recalde A (2016) The archaeological landscape of Late Prehispanic mixed foraging and cultivation economy (Sierras of Cordoba, Argentina). J Anthropol Archaeol 42:88–104
Morello J, Saravia C (1959) El bosque chaqueño. I: Paisaje primitivo, paisaje natural y paisaje cultural en el oriente de Salta. Revista agronómica del noroeste argentino 3:5–79
Nuñez L, MacRostie V, Cartajena I (2009) Consideraciones sobre la recolección vegetal y la horticultura durante el Formativo Temprano en el sureste de la Cuenca de Atacama. Darwiniana 47:56–75
Oliszewski N (1999) La importancia del algarrobo en el Campo del Pucará (Andalgalá, Catamarca) durante el Período Formativo. In: Aschero C, Korstanje A, Vuoto P (eds) En los tres reinos: prácticas de recolección en el cono sur de América. Ediciones Magna Publicaciones, Tucumán, pp 171–186
Pastor S, López ML (2011) Consideraciones sobre la agricultura prehispánica en el sector central de las sierras de Córdoba (Argentina). In: Korstanje A, Quesada M (eds) En arqueología de la agricultura: casos de estudio en la región Andina Argentina. Ediciones Magna, Buenos Aires, pp 208–233
Pastor S, Medina M (2013) Prácticas resistentes, elusión y reproducción social en un contexto histórico adverso. Una mirada a los indígenas de Córdoba (Argentina) en tiempos coloniales tempranos. Memoria Americana 21:65–92
Pastor S, López ML, Rivero D (2012) Access to maize (Zea mays) and its manipulation in hunter-gatherer context in central Argentina (ca. 3,000–2,500 bp). Before Farm Archaeol Anthropol Hunt Gather 2012/4:1–10
Pearsall D (2000) Paleoethnobotany. A handbook of procedures. Academic, San Diego
Piperno D (2006) Phytoliths. A comprehensive guide for archaeologists and paleoecologists. Altamira, Oxford
Planella MT, Tagle B (2004) Inicios de presencia de cultígenos en la zona central de Chile, periodos arcaico y alfarero temprano. Chungara 36(Volumen Especial 1):387–399
Planella MT, Cornejo L, Tagle B (2005) Alero Las Morrenas 1: evidencias de cultígenos entre cazadores recolectores de finales del período Arcaico en Chile Central. Chungará 37:59–74
Planella MT, Scherson R, McRostie V (2011) Sitio El Plomo y nuevos registros de cultígenos iniciales en cazadores del Arcaico IV en Alto Maipo, Chile Central. Chungará 43:189–202
Poca M, Pérez-Harguindeguy N, Vaieretti M, Cingolani A (2014) Descomposición y calidad físico-química foliar de 24 especies dominantes de los pastizales de altura de las sierras de Córdoba, Argentina. Ecol Austral 24:249–257
Rivero D (2009) Ecología de cazadores-recolectores del sector central de las Sierras de Córdoba (Rep. Argentina). (BAR International Series 2007). Archaeo, Oxford
Rivero D, Berberián E (2011) Paleoindian occupation in the central mountains of Argentina: was it a failed colonization? Archaeol Latin Am CRP 28:118–120
Rivero D, Medina M (2013) Human Holocene colonization, diet breadth and niche construction in Sierras of Cordoba (Argentina). In: Cardillo M, Muscio M (eds) Darwin’s Legacy: the status of evolutionary archaeology in Argentina. IMHICIHU-CONICET. Archaeopress, Publisher of British Archaeological Reports, Oxford, pp 76–88
Rivero D, Heider G, Pastor S (2015) Identificación de una punta cola de pescado en las Sierras de Córdoba. Implicancias para el poblamiento del Centro de Argentina. Cuadernos INAPL 24:151–155
Sciammaro L (2015) Caracterización fisicoquímica de vainas y harinas de algarrobo (Prosopis alba y Prosopis nigra). Aplicaciones en productos horneados y fermentados. Doctoral thesis. Universidad Nacional de La Plata
Serrano A (1945) Los Comechingones. (Serie Aborígenes Argentinos 1) Instituto de Arqueología, Lingüística y Folklore de la Universidad Nacional de Córdoba, Córdoba
Staller J (2007) Un aproximación interdisciplinaria para nuestra comprensión de la introducción y el rol temprano del maíz (Zea mays L.) en los Andes Occidentales. In: Oliva F, De Grandis N, Rodríguez J (comp) Arqueología Argentina en los inicios de un nuevo siglo, Tomo I. Publicación del XIV Congreso Nacional de Arqueología Argentina. Universidad Nacional de Rosario, Rosario, pp 23–38
Villagra P, Cony M, Mantován N et al. (2004) Ecología y manejo de los algarrobales de la Provincia Fitogeográfica del Monte. In: Arturi M, Frangi J, Goya J (eds) Ecología y manejo de bosques nativos de Argentina. Editorial Universidad Nacional de La Plata, La Plata, pp 1–32
Acknowledgements
I would like to acknowledge Michele Wollstonecroft and Aylen Capparelli for their valuable comments, allowing me to improve the first version of the text, to the anonymous referees for their invaluable suggestions and to the editors and J. Greig for helping me to improve the English writing. This research received institutional support from Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) PIP 2014–2016 GI and Programa Incentivos-Universidad Nacional de La Plata (Ref. 11/N734), funding to Aylen Capparelli.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Tengberg.
Rights and permissions
About this article

Cite this article
López, M.L. Archaeobotany in central Argentina: macro- and microscopic remains at several archaeological sites from early Late Holocene to early colonial times (3,000–250 bp). Veget Hist Archaeobot 27, 219–228 (2018). https://doi.org/10.1007/s00334-017-0627-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-017-0627-x