
Abstract
The Sabana de Bogotá in the northern Andes is an interesting region to investigate temporal patterns of dietary variation because it counts with a vast archeological and osteological record for the last 10,000 years. In this paper, stable isotope data of human archeological bone collagen and apatite were used to study the evolution of diet and major subsistence transformations taking place during the Holocene (∼9000–600 cal BP). Paleoenvironmental reconstructions and the isotopic ecology of the Sabana de Bogotá were used as an interpretative baseline. Stable isotope measurements (δ13Ccol, δ13Cap, δ15N, and Δ13Ccol-ap) representing hunter-gatherers, horticulturalists, and agriculturalists (N = 134 individuals) were analyzed by using bivariate, regressional, and discriminant statistical techniques. Results show that early Holocene hunter-gatherers (9000–7000 cal BP) consumed mostly C3 vegetal resources locally available. In contrast, animal protein was less important. Middle Holocene hunter-gatherers (6000–4500 cal BP) continued with the food foraging pattern observed in the earlier counterparts and presented a slight increase in C3 animal protein intake. During the initial late Holocene ca. 4000 cal BP, important shifts in subsistence strategies occurred when populations presented a trend toward mixed C3/C4 diets, and by ca. 3500 cal BP, there is a clear signal of C4 crops (i.e., maize) consumption concomitant with the introduction of ceramic technology. During the final late Holocene (last 2000 cal years BP), intensive agriculture was adopted and humans presented relatively diverse diets integrated by C4 and C3 crops, C3-C4 feeding animals, and freshwater resources. Such dietary change coincides with an increase in sociopolitical complexity, population size, and a general decline in health.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Sabana de Bogotá (SB) in the northern Andes is a region with substantial evidence of major subsistence shifts taking place throughout the Holocene. Traditional conceptions indicated that late Pleistocene/early Holocene hunter-gatherers were specialized groups which relied heavily on large-sized and medium-sized mammals and used a homogeneous and specialized lithic industry named Abriense (Correal and van der Hammen 1977; Correal 1986, 1990a; Hurt et al. 1977; van der Hammen et al. 1990). Despite the fact that reliable evidence of extinct mammals like proboscidean gomphotheres, xenarthrans, and American horses has been recovered (e.g., Tibitó), more recent interpretations suggest that generalist foragers with broad spectrum economies inhabited the region during the late Pleistocene, developing throughout the early Holocene diverse subsistence strategies with a strong focus in plants and to a lesser extent in small/medium-sized mammals (e.g., deer, guinea pig among others) (Aceituno et al. 2013; Aceituno and Loaiza 2015; Borrero 2015; Cárdenas 2002; Delgado et al. 2015a,b; Delgado 2015). Reassessments of the radiocarbon chronology along with recent technofunctional and bioarchaeological analyses suggest that several groups relying on C3 resources and with distinct lithic traditions persisted and coexisted over this time span (Aceituno and Rojas 2015; Delgado 2015; Delgado et al. 2015a,b; Nieuwenhuis 2002).
During the middle Holocene non-local forager populations with different settlement patterns, mortuary practices and skeletal morphology arrived in the region (Ardila 1984; Correal and Pinto 1983; Correal 1990b; Delgado 2012a, 2015). Additionally, the appearance of tools for vegetal processing (anvils, scrapers, hammerstones, among others) ca. 5800 cal BP highlights the increasing importance of C3 plant consumption (Ardila 1984; Correal 1990b). Certain cultigens, such as squash (Curcubita pepo), oca (Oxalis tuberosa), and Dioscorea dated around 4500 cal BP are thought to have been introduced during this period (Ardila 1984; Correal 1990b; Cárdenas 2002). Skeletal and isotopic evidence indicates the presence of abrasive diets and oral pathologies related to higher consumption of non-processed foods, as well as continuity in C3 plant consumption and changes in C3 protein sources (Cárdenas 2002; Delgado 2015; Gómez 2012). The increase in temperature/dryness and decrease in rainfall between ca. 7000 and 4500 cal BP caused important ecological changes (i.e., reduction of water tables, habitat fragmentation, etc.), which promoted multiple changes in hunter-gatherer populations such as demographic contraction, population extinction, and dispersals (Delgado 2012a, 2015).
The initial late Holocene period is central for our understanding of the adoption of agriculture given that important changes in subsistence, demography, and food processing technology occurred along with strong phenotypic diversification (Ardila 1984; Botiva 1989; Correal and van der Hammen 1977; Correal and Pinto 1983; Correal 1990b; Delgado 2012a,b, 2015; Rodríguez and Colantonio 2015). The isotopic evidence reveals diverse dietary patterns (C3, C3/C4, and C4), which likely suggest that a change toward a food production system likely began during that time (Cárdenas 2002; Delgado 2015; van der Hammen et al. 1990). This is supported by the extensive archaeobotanical evidence recovered from Aguazuque and Zipacón (ca. 4000/3500 cal BP), which reveals that local populations were already consuming several C3 and C4 domesticated plants by that time period (Correal and Pinto 1983; Correal 1990b). Analyses of multiple bioarchaeological indicators suggest a reduction in food consistence, an increase in carbohydrates and silica-rich plant consumption, a higher biological stress, population contacts/conflicts, and an increase in population size (Delgado 2012a, 2015; Gómez 2012; Rodríguez 2006). The zooarchaeological and isotopic record suggests a slight increase in animal protein consumption and a diversification of animal resources (Correal and Pinto 1983; Correal 1990b; Cárdenas 2002; Delgado 2015).
During the final late Holocene, the archeological evidence suggests that the most remarkable dietary change occurred after 2000 cal BP, when people consumed considerably more tropical plants (primarily, but not exclusively, maize) and animal protein. For the last 1100 cal BP, during a period named Muisca, divided into early (1000–700 cal BP) and late (700–400 cal BP), the archaeobotanical and isotopic record, along with ethnohistorical accounts (restricted to the late period), stresses the importance of several cultigens (e.g., highland tubercles, tropical grains, fruits, and palms) and diverse C3-C4 animal protein sources, including those from freshwater ecosystems (Delgado et al. 2014; Enciso 1996; García 2012; Langebaek 1987; Morcote 1996; Parra 2001; Simón 1981 [1625]). The skeletal evidence further supports the dietary change as revealed by the increase in both occlusal dental wear and the presence of certain pathologies indicating a very high consumption of highly processed carbohydrates, iron deficiency, and physiological stress (Delgado 2015; Martínez 2012; Rodríguez 2006).
As noted above, the major subsistence and dietary transformations in the study region have been addressed by using distinct archeological indicators. While stable isotopes, which provide quantitative dietary information at the individual level, have been used in previous studies of the region, an analysis by using a broad chronological and comparative approach has never been undertaken. Therefore, the aim of the present study is to investigate the main dietary/subsistence changes which occurred in Sabana de Bogotá throughout the Holocene (ca. 9000 to 600 cal BP) by using stable isotopes of the bone collagen and apatite derived from human archeological samples.
Samples and techniques
Currently, stable isotope analysis is a well-established technique widely used for reconstructing diet in distinct spatial and temporal contexts. While not shown to be specific for the area of this study, a common range in δ13C for C3/C4 plants consumed by humans is from −24 to −10 ‰ (−14 ‰), with each per mil more positive than −24 ‰ representing about 7 % maize (or other C4 plant) in the overall diet (Katzenberg 2008; Schoeninger 2009; Tykot 2004). Bone collagen and bone apatite have resorption rates estimated at 7–10 years or more, so that isotopic analyses of adult bones provide the average diet for many years prior to death (Tykot 2006, 2014). Bone collagen δ13C is metabolically enriched about +5 ‰ relative to diet. It is known that while bone collagen is disproportionately produced from the protein portion of the diet, bone apatite is produced from a mixture of dietary protein, carbohydrates, and fats (lipids) (Price and Burton 2011; Tykot 2004). The diet-apatite spacing differs according to the model used and varies according to the C3 and C4 components of the diet. Under a model of +12 ‰ for C3/C4 dietary components, the value for mono-isotopic diets is 7.0 ‰, for C3 protein/C4 total diet is >7.0 ‰, and for C4 total diet/C3 protein is <7.0 ‰ (Tykot et al. 2009). Stable carbon isotope analysis of both bone collagen and apatite thus permits quantitative estimates of several dietary components. Estimating the percentage of C4 plants in human diet is straightforward for herbivores and human agriculturalists, if seafood was not available and animals were not consuming wild C4 grasses, by simple interpolation between the endpoints of bone apatite for a pure C3 -based diet vs. a pure C4 -based diet (Kellner and Schoeninger 2007). Nitrogen isotopes generally reflect trophic-level differences between plants and herbivores, and especially in multilevel aquatic systems, so that the combination of N and C (both collagen and apatite) isotope values allows discrimination between these resource groups.
The available regional dataset includes 134 stable isotope measurements (δ13Ccol, δ13Cap, δ15N, and Δ13Ccol-ap), which were compiled from published sources (Aufderheide 1990; Correal 1990b; Cárdenas 1993, 1996, 2002; Delgado et al. 2014; Rodríguez and Cifuentes 2005; Rodríguez 2011; van der Hammen et al. 1990). Such data represent adult/adolescent (male and female) individuals recovered from several archeological sites distributed in the SB region (Table 1, Fig. 1). Supplementary Table 1 details the isotope data and sample composition investigated here. Five main chronological/economic groups were studied (see Table 1) taking into account contextual archeological and chronological information. This includes early (∼9000–7000 cal BP) and mid-Holocene (∼6000–4500 cal BP) hunter-gatherers, initial late Holocene horticulturalists (∼3800–2200 cal BP), and final late Holocene agriculturalists from the early (∼1015–800 cal BP) and late Muisca (∼700–690 cal BP) periods. Despite the presence of some chronological overlapping between the early and late Muisca samples investigated, especially between the upper and lower temporal ranges, such division may shed interesting information regarding dietary diversity during the Muisca period. Radiocarbon dates were calibrated (2σ) by using the IntCal13 curve on the CALIB program v. 7.0.4 (Stuiver and Reimer 1993). General trends were investigated through bivariate comparisons, plots of temporal dispersion, and mean differences (p < 0.05). The regression model proposed by Kellner and Schoeninger (2007) was used to obtain information on diet, dietary energy, and protein sources. Plots of δ13Ccol against δ13Cap provide a model of three regression lines (C3, C4, and marine diet protein) where the position on each line indicates the energy source (C3, C4, or mixed). In addition, the multivariate statistical approach proposed by Froehle et al. (2012), which includes not only carbon isotopes from bone collagen (of δ13Ccol) and apatite (of δ13Cap) but also from nitrogen (δ15Ncol), was used to distinguish dietary groups more clearly. In order to determine the optimal number of clusters in the dataset, a k-means cluster analysis was performed. Once the most realistic number of groups was determined, a discriminant function analysis (DFA) was carried out in order to obtain significant linear functions and to identify variables that explain the multivariate distribution of individual points and centroids. The analyzed dataset meets the assumptions of multivariate normality within groups and equality of covariance matrices among groups. The statistical and graphical analyses were performed in Systat 13 for windows, Statistica 7.0 Statsoft Inc., and the free R software (R Core Team 2015).

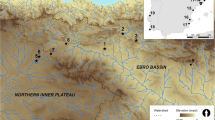
The Sabana de Bogotá region showing the location of archeological sites with human and faunal skeletal remains investigated. Checua (1), Tequendama (2), Potreroalto (3), Galindo (4), Aguazuque (5), Madrid (6), final late Holocene Muisca settlements (Portalegre, Las Delicias, Candelaria, Tibanica) (7)
Paleoenvironmental reconstruction and the regional isotopic ecology
Holocene environmental evolution
Based on palynological, glaciomorphological, isotopic, and diatom evidence, detailed reconstructions of the predominant environmental conditions have been performed for the SB region for the last 13,000 years (Fig. 2). At the El Abra stadial (ca. 12,900–11,800/10,900 cal BP), the regional equivalent to the Younger Dryas Chronozone, the climate was very cold, with inferred temperatures between 4 to 6 °C lower than today according to pollen data (van der Hammen and Hooghiemstra 1995) or 3 °C lower than today in accordance to δ13C data derived from soil organic matter (SOM) samples (Boom et al. 2001; Mora and Pratt 2002). Biome reconstructions revealed a dominance of the cool grassland-shrub biome (Marchant et al. 2002). Around 11,500 cal BP, the climate ameliorated again, with a sudden rise in the average annual temperature (van der Hammen 1992). The biome reconstructions for the 10,800–9500 cal BP interval revealed that there was a remarkable expansion of mesic biomes, which indicates a warm and wet period (Marchant et al. 2002). Between ca. 8300 cal BP and ca. 6800/5800 cal BP, the record shows vegetation changes, fluctuations in precipitations and temperature with a tendency to dry environments, and the subsequent decrease of water tables (Fig. 2) (Gómez et al. 2007; Vélez et al. 2006). Biome reconstructions suggest a marked increase of the cool mixed forest biome (Marchant et al. 2002). Between 5800 and 3800 cal BP, the paleoenvironmental record suggests an increase in temperature (±2 °C) and a decrease in precipitation, which caused a particularly dry period. After the 2500 cal BP, climatic conditions became similar to what we know today (Fig. 2). Pollen and sediment records suggest a high degree of precipitation (reflected by an increase of marshy herbaceous vegetation and open water) as well as intensive human occupation shown by deforestation (Quercus in particular). In addition, pollen data suggest lowering of water tables and increasing disturbance probably caused by erosion (presence of Amaranthaceae/Chenopodiaceae, Borreria, and Dodonaea), crop cultivation (presence of Zea mays), and frequent fires (as shown by the high amounts of charcoal in the sediments) (Gómez et al. 2007; Vélez et al. 2006). Biome reconstructions revealed an increase in the cool grassland-shrub biome (Marchant et al. 2002).

Climatic-vegetation evolution and biome reconstructions for the Sabana de Bogotá region for the last 19,000 14C years on the basis of palynological, glaciomorphological, and diatom evidence (adapted from Delgado 2012a). Pantano de Vargas pollen zone (Zone VG). The gray bars shows the chronological position of the Guantiva interestadial, El Abra stadial, and the mid-Holocene climatic optimum
Establishing the menu: the regional isotopic ecology
Stable isotope data provide reliable quantitative information on diet at the individual level but requires an interpretative context with respect to the potential resources that may have been consumed. Such context is termed isotopic ecology, and it is useful to define the regional dietary spectrum. Regarding vegetal resources, we can obtain some information from isotope-based paleoenvironmental research on the Holocenic altitudinal expansion of wild C4 vegetation (Boom et al. 2001; Guillet et al. 1988; Mora and Pratt 2002) and from previous human stable isotope studies (Cárdenas 1993, 1996, 2002; Delgado et al. 2014). According to paleoenvironmental studies, elevated δ13C values ranging from −9 to −15 ‰ within the Andean/sub-Andean forests and moorland vegetation belts are indicative of the presence of C4 and CAM plants during different moments of the Holocene (supplementary Table 2). For the early/mid-Holocene, some authors hypothesized that the presence of mixed C3/C4 vegetation can be explained by the expansion of lowland C4 grasses caused by lower partial atmospheric CO2 pressure (pCO2), reduced rainfall, and temperature fluctuations related to rapid climatic changes, whereas human-induced deforestation and/or cultivation of C4 crops (maize mainly) and environmental rearrangements could explain the presence of C4 vegetation in the late Holocene (Boom et al. 2001; Guillet et al. 1988; Mora and Pratt 2002). With regard to maize consumption, previous studies on Muisca human skeletal remains (Cárdenas 1993, 1996, 2002; Delgado et al. 2014) suggest that carbon values ranging from −12.8 to −9.7 ‰ likely correspond with a maize-based diet. Isotopic data derived from Native American skeletons from the colonial period suggest that values averaging −9.7 ‰ or higher are indicative of diets based mostly on maize consumption (Cárdenas 2002; Delgado 2007). Although the isotopic research on faunal material is scarce, it has allowed us to make some inferences although restricted to the late Holocene (Table 2; Delgado et al. 2014). As expected, carbon isotopes of two deer samples (Odocoileus virginianus) reflect a C3 diet, which likely reveals intra-specific differences. However, when compared to fossil and experimental evidence (Emery et al. 2000; Kohn et al. 2005), they seem to have been enriched. Carbon enrichment is more evident in the guinea pig and rabbit samples (Table 2), whose values resemble a C3/C4 diet. Despite the fact that human influence may have been a factor, especially for cavia given its probable domestication date established at ca.3500–2600 cal BP (Correal and Pinto 1983; Ijzereef 1978), similar values in rabbit and the detection of C4 and CAM plants in the study region during the late Holocene likely indicate the consumption of C4 wild grasses by these animals. The high nitrogen value found in the rabbit sample is not in line with their feeding habits and local ecology as they can only get nitrogen from the plants they consume. Nitrogen data from carnivorous species such as dog (Canis familiaris) and margay (Leopardus wiedii) revealed their higher trophic position within the local terrestrial food web. Carbon values resembling a C4 diet can be explained by the omnivorous dietary profile of dog and/or human influence and a wide geographic range and feeding behavior for margay, which includes small/medium-sized mammals and birds that inhabit both the tropical lowlands and Andean highlands. Finally, data for a duck species (Dendrocygna auntumnalis) reveal a C3 diet complemented with freshwater resources (e.g., mollusks), which indicates highland open water habitats (Moreano 2011).
Results
Univariate and bivariate comparisons
In Tables 3, 4 , and 5 and Figs. 3 and 4, the main chronological/economic trends for the groups under study are presented. By the early Holocene, the low δ13Ccol values indicate a high dependence on C3 plants. In addition, one individual presented a mixed C3/C4 diet (see Figs. 3a and 4a). Nitrogen isotope ratios (Figs. 3a, b and 4b) revealed two subgroups with low and mean values, respectively, which suggests the presence of distinct protein sources (C3 plants and several C3-feeding animals) at that time. During the middle Holocene, people continued consuming basically C3 plants and there is also a certain increase in nitrogen, which reflects higher consumption of C3-consuming animals and likely freshwater resources. Early and mid-Holocene samples did not present significant differences in their diets (Tables 4 and 5). The carbon isotope data for the initial late Holocene show some changes regarding previous periods, since while some individuals maintained a C3 dietary profile, others presented either mixed C3/C4 or C4 diets. During this time, the slight increase in nitrogen reveals changes in protein sources and/or increase in protein consumption. This trend is reinforced by the t test revealing further differences in carbon (collagen) and nitrogen (Tables 4 and 5) with other time periods. For the final late Holocene, there was a clear C4 signal indicating more reliance on tropical crops, primarily maize along with other C3 and C4 plants. There is also a considerable increase in nitrogen and apatite values (Figs. 3 and 4b, c), which indicates a period of higher consumption of C3-C4 animals and freshwater resources. Interestingly, some differences emerged when the early and late Muisca samples were compared such as the higher protein consumption during the late period (Figs. 3 and 4 and Tables 3, 4, and 5). Furthermore, the diet-apatite spacing values (Fig. 4d) suggest diets integrated by C3 carbohydrates and C3 protein sources during the early and middle Holocene, C4 carbohydrates and C3 protein during the initial late Holocene, and C4 carbohydrates and C3-C4 protein by the final late Holocene.

δ13Ccol plotted against δ15N from Sabana de Bogotá human samples for four chronological samples: early Holocene hunter-gatherers (triangles), middle Holocene hunter-gatherers (circles), initial late Holocene hunter-gatherers/horticulturalists (diamonds), and final late Holocene agriculturalists (squares). a Individual data points and b mean values ± 1 standard deviation. Ninety-five percent confidence ellipses for each chronological group are shown

Scatterplots of four stable isotopes against chronology raging between 9000 and 500 cal BP for human samples from the Sabana de Bogotá region. a δ13Ccol, b δ15Ncol, c δ13Cap, and d ∆13Ccol-ap. Color codes as in Fig. 3
Regression analysis
Figure 5 illustrates the results of Kellner and Schoeninger’s (2007) model. Individuals varied substantially in their source of dietary energy and dietary protein over time, with opposite patterns during the final late Holocene and the early/mid-Holocene. As indicated by the model, while most final late Holocene individuals show a strong emphasis on C4 dietary energy sources, some late individuals fall near the C4 line, albeit their position is closer to mixed C3/C4 dietary energy. At least one individual presented exclusively a C3 protein source. The regression results are consistent with the bivariate comparisons and the t test since they revealed slight differences between the early and late Muisca periods in carbon isotopes. Overall, the data corresponding to the final late Holocene showed that 50–70 % of dietary protein and dietary energy came from C4 sources. Early and mid-Holocene hunter-gatherers fall along the C3 protein line with no or few C4 signal, except for one individual showing mixed C3/C4 protein sources. With the exception of the aforementioned outlier, about 80–100 % of their dietary energy came from C3 sources. In addition, isotope data for the first 5000 years of the Holocene show homogeneous dietary sources and relatively diverse energy sources. Although the initial late Holocene sample is underrepresented, the two samples analyzed here have similar values to those from the early and mid-Holocene, which suggest similar C3 dietary and energy sources. In general, the temporal spread of the data reveals variability in dietary protein and dietary energy composition over the Holocene in the study region, with an emphasis on C3 protein/energy sources during the early and middle Holocene and C4 protein/energy sources during the initial and final late Holocene.

δ13Ccol plotted against δ13Cap for five chronological samples from the Sabana de Bogotá region according to the regression model of Kellner and Schoeninger (2007). Early Holocene hunter-gatherers (triangles), middle Holocene hunter-gatherers (circles), initial late Holocene hunter-gatherers/horticulturalists (diamonds), and final late Holocene agriculturalists (squares). Color codes as in Fig. 3
Multivariate analysis
The results of the DFA are presented in Tables 6 and 7 and Fig. 6. The k-means cluster analysis revealed the existence of four main clusters with distinct multivariate dietary profiles. Only linear functions with higher variance and canonical correlations were interpreted (i.e., functions 1 and 2 = 99.8 % of the total variance). The structure analysis showed that carbon isotope variables have the heaviest influence on function 1, while nitrogen exerted its strongest effect on function 2 (Table 6). The classification analysis based on posterior probabilities and inter-individual Mahalanobis distances (D 2) revealed a very good correspondence between the observed and predicted isotope-based classification (only 0.07 % incorrect classifications). The four clusters are plotted in Fig. 6, both as individual data points (a) and multivariate centroids (b). Function 1 clearly differentiates between clusters with low (negative scores) and high (positive scores) carbon values. Clusters 1 (circles) and 2 (squares) are markedly different from clusters 3 (diamonds) and 4 (triangles). This distinction has economic sense, since clusters 1 and 2 correspond to hunter-gatherers from the early, middle, and initial late Holocene and clusters 3 and 4 correspond to final late Holocene agriculturalists. Function 2 allows us to distinguish between clusters with low (negative scores) and high (positive scores) nitrogen values. This differentiation can be explained in temporal terms since cluster 1 corresponds to early Holocene hunter-gatherers with low nitrogen values and cluster 2 comprises middle and initial late Holocene hunter-gatherers with high values. The distinction between clusters 3 and 4 based on function 2 (nitrogen) supports the bivariate results (Fig. 3; Tables 4 and 5) suggesting that people from the late Muisca period consumed higher amounts of protein than their early counterparts. Overall, the multivariate model suggests that early and middle Holocene foragers presented ca. 100 % of C3 diets and ≥65 % of C3 protein sources, while final late Holocene agriculturalists presented 70 % of C4 diets and ∼50 % of C4 protein sources. The high positive scores of some individuals along function 2 suggest either the presence of marine protein (see Somerville and Froehle 2011) or a strong focus on local freshwater fauna. Alternatively, the elevated nitrogen values could be at least partly attributed to the use of animal fertilizers, burning of vegetation, and/or land management by the final late Holocene societies (see Szpak 2014). Finally, Table 7 presents the matrix of D 2 distances between the four dietary clusters, which revealed significant differences between clusters 1–2 and 3–4 but not between 1 and 2 or between 3 and 4. This suggests that, in dietary terms, cluster 1 (low carbon/low nitrogen), representing early Holocene hunter-gatherers, is similar to cluster 2 (low carbon/high nitrogen), which includes mid and initial late Holocene hunter-gatherers/horticulturalists. Additionally, clusters 1 and 2 are markedly different from clusters 3 (high carbon/low nitrogen) and 4 (high carbon/high nitrogen), which correspond to final late Holocene agriculturalists from the early and late Muisca period, respectively.

Discriminant function scores plotted by cluster representing four multivariate dietary profiles as established by the discriminant function analysis. Cluster 1 (circles), cluster 2 (squares), cluster 3 (diamonds), and cluster 4 (triangles). a Individual data points and b multivariate centroids. Error bands extend to 2σ of the centroids. Ninety-five percent confidence ellipses for each cluster are shown
Discussion: subsistence and dietary trends at the Sabana de Bogotá
The isotopic evidence presented above revealed an important transformation in diet and subsistence at the SB region over time, which can only be assessed on wide archeological timescales (i.e., millennia). Here, the dietary/subsistence trends are addressed by temporal blocks roughly corresponding to periods of remarkable cultural change in the study region.
Early Holocene (ca. 9000–7000 cal BP)
Past interpretations on SB early hunter-gatherer subsistence indicated dietary specializations (Correal and van der Hammen 1977; Correal 1986; Hurt et al. 1977; van der Hammen et al. 1990), while recent studies tend to stress more generalized diets and economies (Aceituno and Loaiza 2015; Borrero 2015; Cárdenas 2002; Delgado 2015; Piperno and Pearsall 1998). Much of the subsistence specialization (i.e., hunting) was inferred from the lithic assemblages; however, the most important and ubiquitous late Pleistocene/early Holocene regional lithic industry (i.e., Abriense) is not a bifacial technology but a very simple, versatile, and multifunctional unifacial technology that reflects environmental constraints rather than specializations (Delgado 2012a). Recent technofunctional analysis revealed the existence of broad spectrum economies with a strong focus on vegetal resources (Nieuwenhuis 1998, 2002). The isotopic results presented here are in agreement with the recent interpretations of the lithic repertoire indicating that early Holocene forager diets were mostly integrated by C3 plants. Although animal protein was less important, it should be noted that this protein came from diverse C3 sources, which, in turn, indicates flexibility in resource acquisition. The multivariate and regression analyses (Figs. 5 and 6; Table 6) strongly support this interpretation as about 80–100 % of the dietary energy of early hunter-gatherers came from C3 vegetal sources. In addition, the temporal dispersion of nitrogen and diet-apatite spacing (Fig. 4b, d) stress that C3 animal protein was also a relatively important dietary component for some early foragers. This pattern is supported by distinct lines of evidence. For example, the archaeobotanical and lithic evidence highlights the importance of roots, tubercles, and seeds (Cárdenas 2002; Correal and van der Hammen 1977; Nieuwenhuis 2002), and the zooarchaeological record points to deer, guinea pigs, and other small mammals as the main animal protein sources (Ijzereef 1978; Peña and Pinto 1996). Bioarchaeological evidence indicates abrasive diets and oral pathologies related to the high consumption of minimally processed foods (Delgado 2015; Gómez 2012). On the one hand, all this evidence indicates that gathering was more important than hunting. On the other, however, there was more dietary diversity than previously assumed, which leads to the hypothesis of non-specialized economies for early Holocene foragers.
The early Holocene was a period without marked environmental changes in the SB region (Fig. 2). Likewise, aside from a slight increase in carbon (collagen) and nitrogen during the middle Holocene, the isotopic record shows overall relative dietary stability between 9000 and 4500 cal BP, which means that few significant changes occurred over millennia in connection with subsistence (Fig. 4; Tables 4 and 5). However, individual data also suggest some interesting issues, especially regarding mobility and the arrival of non-local populations. The most conspicuous finding is the different dietary profile of an early Holocene individual from Galindo site (Pinto 2003) dated to 7730 ± 60 BP (GrN-16345) (8508 cal BP range 2σ 8406–8604), which presented a mixed C3/C4 diet (see Figs. 3, 4, and 5). Although some authors have pointed out that because this skeleton has not been directly dated, its unusual carbon value (−15.3 ‰) could represent a late agricultural individual unrelated to the dated stratum (Cárdenas 2002). A distinct interpretation suggests that such individual could have come from a lowland region and/or that it may have belonged to a highly mobile group exploiting tropical resources frequently. Recent analysis of dental and craniofacial morphology suggests that during the early and middle Holocene, populations with distinct phenotypic patterns could have arrived, favoring the hypothesis of a non-local origin for such individual (Delgado 2015).
Some authors have suggested that the regional isotopic record reflects the importance of logistic mobility (Cárdenas 2002; van der Hammen et al. 1990) during the early Holocene, given the allegedly clear signal of tropical C4 plants and large inter-individual isotopic variation (Cárdenas 2002). This, along with the paucity of the archaeobotanical record for this period, has led to interpret SB as a region for hunting rather than gathering, where much of the vegetal resources come from tropical lowlands (Cárdenas 2002). The results presented here, excluding the Galindo individual, are not consistent with the aforementioned hypothesis. On the contrary, they suggest an entire homogeneous C3 diet indicative of an intensive use of locally available vegetal resources (Figs. 3, 4, 5, and 6; Tables 3, 4, 5, and 6). Additionally, the low preservation of the paleobotanic remains may, in part, explain their alleged absence in early contexts. Although the available isotopic evidence is not very useful to discuss human mobility, it suggests that early Holocene foragers exploited mostly local C3 ecosystems and had therefore a restricted geographic range. This is partially supported by the paleoecological evidence, which suggests a relatively homogeneous distribution of C3 resources over the early/middle Holocene for the SB (Boom et al. 2001; Mora and Pratt 2002). Nevertheless, the finding of both foreign fauna (e.g., Felidae and Atelidae families) and non-local lithic tools (e.g., fluting technology) by using allochthonous raw materials suggests that the widely accepted territorial mobility hypothesis (e.g., Cárdenas 2002; Correal and van der Hammen 1977) must be monitored by using other lines of evidence (Gnecco 2000).
Middle Holocene (ca. 6000–4500 cal BP)
The isotopic data for this temporal block shows few differences regarding the early Holocene, especially in terms of carbon isotopes, which suggest a continuous exploitation of local C3 vegetal resources and that there was virtually no change in gathering. Both the univariate/bivariate and regression/multivariate analyses indicate strong dietary similarity between early and mid-Holocene foragers (Figs. 3, 4, 5, and 6; Tables 3,4, 5,6 and 7). The only change regarding the previous period is a slight increase in nitrogen and diet-apatite spacing, which indicates higher and more diverse animal protein consumption during the middle Holocene (Figs. 3 and 4b; Table 5). The zooarchaeological record indicates that small mammals, such as guinea pigs, became more important than deer during this time period. The increased importance of Cavia porcellus over other animal resources (e.g., deer) for middle Holocene hunter-gatherer economic strategies remains unclear. From the perspective of evolutionary geography (sensu Lahr and Foley 1998), the author (Delgado 2012a, 2015) suggested that mid-Holocene climatic changes (temperature increase and aridity) may have caused significant ecological modifications resulting from climate-dependent chorological changes affecting vegetal and animal communities. Thus, the retraction and/or fragmentation of forested areas likely produced the decline of deer populations, which, in turn, allowed the more intensive use of guinea pigs and other small-sized animals adapted to open habitats (i.e., grasslands) and freshwater resources. The relatively wide range of nitrogen (Figs. 3 and 4b; Table 3) seems to support this interpretation since it suggests the consumption of relatively diverse animal protein sources during this period.
An important aspect that must be taken into account for the interpretation of the emergence of agriculture in the study region from the isotopic record is that despite the fact that the most important changes related to food production are more noticeable during the initial and final late Holocene (e.g., the introduction of tropical crops, the intensive cultivation of C3 plants, etc.), this does not necessarily imply that that an early horticulture of C3 plants was not developed, especially during the middle Holocene. The distinct analyses performed here, both univariate/bivariate and multivariate revealing a strong emphasis on C3 plants, seem to support a mid-Holocene horticulture system (Figs. 3, 4, 5, and 6; Tables 3, 4, 5, and 6). This is in agreement with the rich paleobotanical record highlighting the importance of squash, Dioscorea, oca, and other highland tubercles (Cárdenas 2002; Correal 1990b; Correal and van der Hammen 1977). In addition, the initial appearance of lithic tools for vegetal processing in the local archeological record dated to ca. 5800 cal BP (GrN-14477) (range 2σ 5560–5798) (Correal 1990b) also highlights the intensification of plant use from the middle Holocene onwards. This is interesting, as similar kinds of lithic tools have been found in archeological sites with clear and reliable evidence of plant management, since the Pleistocene/Holocene transition (Aceituno and Loaiza 2015; Gnecco and Mora 1997; Gnecco 2003).
Initial late Holocene (ca. 3800–2200 cal BP)
The initial late Holocene is a crucial period to address plant domestication and the origins of agriculture in the study region because it is during this period that some of the most important changes in food production occurred. The results of this study show the existence of C3, mixed C3/C4, and C4 diets (Figs. 3 and 4; Tables 3, 4, and 5). Although it is possible that during this period groups and/or individuals with distinct diets coexisted, this pattern also can be the result of poor chronological resolution and small sample size. Thus, the transitional character of this period may change in the near future with more and well-dated human samples. The extensive archaeobotanical evidence recovered from Aguazuque (ca. 4300 cal BP) and Zipacón (3500 cal BP (GrN-11125) (range 2σ 3445–3571), which includes sweet potato (Ipomoea batata), calabash tree (Cresentia cujete), avocado (Persea americana), cherry tree (Prunos serotina), and maize (grains and cobs of Z. mays) (Correal 1990b; Correal and Pinto 1983), points out that by the initial late Holocene, SB populations were already consuming several C3 and C4 domesticated plants. Regardless of some fluctuations, isotopic information suggests that the main dietary change occurred ca. 3000–2800 cal BP, when people consumed significantly more C4 plants and animal protein (Fig. 4). The chronological discrepancy between the paleobotanic and the isotopic record regarding the initial time of dietary change could be attributed to the paucity of the isotopic evidence. Yet, the finding of two individuals from the upper levels of Tequendama dated around 3900–3500 cal BP and with carbon values indicative of mixed C3/C4 diets (see Fig. 4a) shows that actually a date of ca. 4000 cal BP may more reliably represent the initial change toward a C4 diet. Bioarchaeological analysis supports the dietary change inferred from the isotopic and archaeobotanical record, since it indicates, on the one hand, an important increase in oral pathologies such as caries, dental calculus, and enamel hypoplasias and, on the other, the reduction of occlusal dental wear rates (Delgado 2015). The results also showed a slight increase in nitrogen (Fig. 4b), which coincides with the preponderance of guinea pig remains, highlighting the economic importance of this species for late Holocene horticultural societies (Correal and van der Hammen 1977; Correal and Pinto 1983; Correal 1990b). This is interesting because on the basis of the analysis of skeletal morphology, Correal and Pinto (1983, p. 183) suggested that the probable date for the initial domestication of this species is around 3500 cal BP, which supports previous proposals (Ijzereef 1978, p. 170) that state that by ca. 2600 cal BP, these animals had been fully domesticated.
One of the main goals of this research was to provide interpretations about the biocultural processes that might have allowed the cultivation of plants. I also attempted to understand whether these biocultural processes were promoted by external factors such as climatic changes and/or population dispersals or whether they were the product of an in situ development. After evaluating the temporal dispersion of the isotopic record (especially carbon isotopes) throughout the Holocene (Figs. 4 and 5; Tables 3 and 4) by using distinct statistical and graphical approximations, the most conspicuous patterns are (1) homogeneity of the C3 signal between 9000 and ca. 4500 cal BP; (2) the presence of C3, C3/C4, and C4 diets between ca. 4000 and 3500 cal BP; and (3) the preponderance of the C4 signal after 2000 cal BP (Figs. 3, 4, 5, and 6; Tables 3, 4, and 5). Despite the fact that in situ development of agriculture is likely, especially given the importance of vegetal resources for the hunter-gatherer economy during the early and middle Holocene, the occurrence of strong, rapid, and multiple biocultural changes suggests that other factors were likely involved, with the most relevant being the entry of non-local populations with an agricultural subsistence. Climate change and environmental rearrangements do not seem to have been an important factor prompting to subsistence changes as the paleoecological record shows only a slight increase in precipitation and humidity during this time span (Marchant et al. 2002). In this context, it is worth mentioning the findings concerning the earliest evidence of maize and ceramic technology in the study region dated to around 3500 cal BP (Correal and Pinto 1983). Such evidence suggests, on the one hand, that despite the fact that maize could have been introduced well before 3500 cal BP given the early evidence of maize in other regions of northwestern South America (Bray et al. 1987; Cavelier et al. 1995; Monsalve 1985), its role as the mainly consumed plant took place later. On the other hand, it is striking that the earliest evidence of maize appeared along with the oldest evidence of pottery related to non-local lowland populations (Botiva 1989), which points to the fact that some of the most important cultivated crops may have been introduced by population expansions. This is in line with the long-held view of local archeologists, who have favored the hypothesis that incipient agriculture was introduced by populations from the Magdalena valley at least since ca. 3500 cal BP (Ardila 1984; Botiva 1989; Correal and Pinto 1983). Even though the isotopic record is fairly incomplete, it suggests similar dates for the initial dietary change around ca.4000 cal BP (Figs. 3 and 4). This is relevant since studies of craniofacial and dental morphology found remarkable evolutionary diversification during the initial late Holocene indicating population discontinuity (Delgado 2012a,b, 2015; Rodríguez and Colantonio 2015). Together, this data indicate that the change from C3 diets related to hunter-gatherers/early horticulturalist to C4 diets characteristic of agricultural societies is concomitant with the appearance and dispersals of food processing technologies (pottery and gridding tools) and the entry of human populations with distinct phenotypic patterns. Accordingly, these lines of evidence strongly support previous hypothesis that non-local populations, likely from the Magdalena valley, with distinct behavioral and biological traits introduced the tropical crop agriculture to the SB region during the initial late Holocene (Correal and van der Hammen 1977; Correal 1990b).
Final late Holocene (ca. 1300–600 cal BP)
The archeological record along with ethnohistorical accounts show that the Muisca society introduced intensive agriculture, developed advanced agricultural strategies, and established wide and complex commercial networks, all of which implied the availability of different foodstuffs from several regions (Cárdenas 1996; García 2012; Kruschek, 2003; Langebaek 1987). The isotopic record coincides with this diverse picture as it shows the existence of relatively heterogeneous diets and a well-established agriculture during the Muisca period, with some differences between the early and late period. The biochemical evidence stresses the importance of C4 plants (primarily maize) complemented with other C3 and C4 crops, as well as C4-feeding animals and freshwater resources (Figs. 3, 4, 5, and 6; Table 2). Although the strong C4 signal suggests that maize was the most commonly used tropical plant, other C4 crops (e.g., manioc), fruits, and palms were likely incorporated into the Muisca diet (García 2012). This means that the overwhelming C4 signal, which until now has been interpreted as maize, in fact reflects more than one tropical resource consumed by these societies corroborating the ethnohistoric evidence (Langebaek 1987).
Previous studies (Cárdenas 1993, 1996) emphasized the absence of Andean foodstuffs in the isotopic record such as highland tubers like potato (Solanum tuberosum), sweet potato (I. batata), arracacha (Arracacia xanthorrhiza), oca (O. tuberosa), mashua (Tropaeolum tuberosum), and ullucu (Ullucus tuberosus), which according to etnohistoric evidence were of crucial importance within the Muisca diet (Langebaek 1987). However, the bivariate results contradict such interpretation, showing instead wide ranges for δ13Ccol (Fig. 3; Tables 3 and 4), where a large part of the Muisca sample averaging ca. −15 ‰ indicates that C3 resources such as highland tubers were also commonly consumed. Similarly, the regression and DFA results (Figs. 5 and 6) point out a C3 trend for some individuals from the early and late Muisca period. What is clear from the isotopic data is that C3 plants were not as important as C4 crops during this time span (Figs. 4a and 5). Interestingly, the local isotopic ecology (Table 2) revealed that some animal species (e.g., guinea pig and rabbit) possessed relatively enriched carbon isotopes, which is explained by the presence of wild C4 plants in the SB region during the final late Holocene (Mora and Pratt 2002; supplementary Table 2). This implies that humans feeding on those animals may have displayed the same phenomenon. These data suggest that the alleged carbon enrichment along with the use of several C4 products might overshadow the isotopic signal of C3 plants.
Some researchers documented a sociopolitical differentiation between the early and the late Muisca periods (Boada 2006; Henderson and Ostler 2005; Kruschek 2003), which coincides with other behavioral aspects, including socially constituted food consumption practices. Here, carbon and especially nitrogen isotopes revealed some differences between both periods. The bivariate results (Figs. 3a, b and 4b–d) show that the sample from the late period present higher nitrogen values, while the sample from the early period present slightly higher carbon ones; some of these differences are statistically significant (Tables 4 and 5). In addition, the DFA results support such chronological differentiation (Fig. 6; Tables 6 and 7). For carbon isotopes, cluster 3 (mostly representative of the early period) shows slightly more negative values indicating the relative importance of C3 resources. For nitrogen, cluster 4 (mostly representative of the late period) shows higher values, which suggests more protein intake, primarily from animal sources (see also Fig. 4b). Cárdenas (1993) interpreted the relatively high nitrogen values in Muisca samples as evidence of marine resource consumption. In line with this, there have also been findings of marine animal remains (mollusks, gastropods, and shells) and the circulation of exotic marine products such as salt, fish flour, and salted and smoked fish within the Muisca territory. However, Cárdenas (1993, 1996) erroneously interpreted the high nitrogen values in infants of 0–3 years old (which are related to breastfeeding actually) as indicative of marine resources. His suggestion regarding the circulation and consumption of exotic marine products is problematic because these resources are not well-represented in the isotopic record. Despite the absence of a highly developed isotopic ecology, especially for local aquatic habitats, the results presented here suggest that the relatively high nitrogen values can be explained by the consumption of freshwater resources (e.g., fish), especially given the vast zooarchaeological evidence for the late period indicating the intensive exploitation of such resources (Enciso 1996).
Taking into account the entire temporal range investigated (ca. 9000–600 cal BP), it is clear that some of the most important changes regarding diet, according to the stable isotope data analyzed (Figs. 3, 4, 5, and 6; Tables 3, 4, 5, 6, and 7), occurred during the late Holocene and more specifically, during the Muisca period. Such dietary change and its associated biocultural transformations had profound implications for the health and behavior of the Muisca societies. Complementary bioarchaeological evidence revealed that during the final late Holocene, there was an increase in the rate and plane of occlusal dental wear potentially related to the intensive use of grinding stones for processing plants. This infused additional environmental grit into the meal in the form of stone chips from the grinding implements producing more pronounced dental wear (Delgado 2015). Increased focus on maize, indicated for the strong C4 signal (Figs. 3, 4, 5, and 6), could have negative consequences on the health of Muisca populations given its association with pathologies that result in iron deficiency, such as porotic hyperosthosis and criba orbitalia. Despite the fact that recent studies suggest more complex etiologies for such diseases (Walker et al. 2009), it is interesting that some authors reported higher frequencies among Muisca populations compared to other periods (Martínez 2012; Rodríguez 2006). Coinciding with the increased maize consumption, there is an increase in the prevalence of carious lesions over the course of the Muisca period (Delgado 2015). Finally, the analysis of dental enamel hypoplasias and non-specific infections (periosteal reactions) provides further evidence that Muisca populations may have experienced a dramatic increase in physiological stress (Delgado 2015; Rodríguez 2006). In general, the introduction of intensive agriculture produced a decline in health for the Muisca populations that reflects a change in the quality of diet and lifestyle.
Concluding remarks
The isotopic record revealed a slow subsistence transformation between the early and middle Holocene (∼9000–4500 cal BP), which suggests dietary stability. Contrary to the expectations of a well-known model on Paleoindian subsistence, early Holocene hunter-gatherers presented a strong focus on plants with some fluctuation in protein sources (i.e., flexibility). The noticeable biocultural and climatic changes that occurred during the middle Holocene (ca. 6000–4500 cal BP) contrast with the stable isotope data only pointing to a slight increase in protein consumption. The appearance of lithic tools for intensive vegetal processing and the finding of some cultigens suggest that C3 plant horticulture emerged during the middle Holocene. However, it is ca. 4000 cal BP when a change toward mixed C3/C4 diets occurred. Around ca. 3500 cal BP, human populations presented a strong C4 signal and a slight increase in protein consumption. During the Muisca period (last 1100 cal years BP), the agricultural societies exhibited a strong reliance on C4 plants such as maize complemented with other C3 and C4 crops, C3-C4 feeding animals, and freshwater resources. In addition, there were differences in animal protein and C3 plant consumption between early and late Muisca samples indicating slight changes in subsistence in the SB region in the previous European contact. Lastly, although this research revealed interesting issues on the evolution of diet over the Holocene in this region, further research is still required. New stable isotope measurements for important periods (e.g., initial late Holocene) are necessary, which include additional data on mobility and dietary mobility along with an enhancement of the chronological resolution and a more complete regional isotopic ecology reconstruction.
References
Aceituno FJ, Loaiza N (2015) The role of plants in the early human settlement of Northwest South America. Quat Inter 363:20–27
Aceituno FJ, Rojas S (2015) Lithic studies in Colombia during the late Pleistocene and the early Holocene. Chungará 47:13–23
Aceituno FJ, Loaiza N, Delgado ME, Barrientos G (2013) The initial human settlement of Northwest South America during the Pleistocene/Holocene transition: synthesis and perspectives. Quat. Inter. 301:23–33
Ardila GI (1984) Chía. Un sitio precerámico en la Sabana de Bogotá. Fundación de Investigaciones Arqueológicas Nacionales. Banco de la República, Bogotá
Aufderheide AC (1990) Results of tests performed on the bones on Aguazuque I. In: Correal G (ed) Aguazuque. Evidencias de cazadores, recolectores y plantadores en la altiplanicie de la Cordillera Oriental. Fundación de Investigaciones Arqueológicas Nacionales, Banco de la República, pp. 305–307
Boada AM (2006) Patrones de asentamiento regional y sistemas de agricultura intensiva en Cota y Suba, Sabana de Bogotá (Colombia). Fundación de Investigaciones Arqueológicas Nacionales, Banco de la República
Boom A, Mora G, Cleef A, Hooghiemstra H (2001) High altitude C4 grasslands in the northern Andes: relicts from glacial conditions? Rev Palaeobot Palynol 115:147–160
Borrero LA (2015) Moving: hunter-gatherers and the cultural geography of South America. Quat Int 363:126–133
Botiva A (1989) La altiplanicie Cundiboyacense. In: Botiva A, Groot AM, Herrera L, Mora S (eds) Colombia prehispánica: regiones arqueológicas. Instituto Colombiano de Antropología e Historia, Bogotá, pp. 77–115
Bray W, Herrera LF, Cardale de Schrimpff M, Botero P, Monsalve JG (1987) The ancient agricultural landscape of Calima, Colombia. In: Denevan WM, Mathewson K, Knaap G (eds) Pre-Hispanic agricultural fields in the Andean region, vol 359. BAR: International Series, pp. 443–481
Cárdenas F (1993) Paleodieta y paleodemografía en poblaciones arqueológicas muiscas (sitios Las Delicias y Candelaria). Revista Colombiana de Antropología. 30:129–148
Cárdenas F (1996) La dieta prehispánica entre poblaciones arqueológicas muiscas. In: Therrien M, Enciso B (eds) Bioantropología de la Sabana de Bogotá. Instituto Colombiano de Antropología e Historia, Bogotá, pp. 85–109
Cárdenas F (2002) Datos sobre la alimentación prehispánica en la sabana de Bogotá, Colombia. Informes Arqueológicos (3). Instituto Colombiano de Antropología e Historia.
Cavelier I, Rodríguez C, Mora S, Herrera LF, Morcote G (1995) No solo de caza vive el hombre: ocupación del bosque amazónico, holoceno temprano. In: Cavelier I, Mora S (eds) . Ámbito y ocupaciones tempranas de la América tropical, ICANH-Fudación Erigai, pp. 27–44
Correal G (1986) Apuntes sobre el medio ambiente pleistocénico y el hombre prehistórico en Colombia. In: Bryan AL (ed) New evidence for the Pleistocene peopling of the Americas. University of Main, Orono, Center for the Study of the Early Man, pp. 115–131
Correal G (1990a) Evidencias culturales durante el Pleistoceno y Holoceno de Colombia. Rev Arqueol Am 1:69–89
Correal G (1990b) Aguazuque. Evidencia de cazadores, recolectores y plantadores en la altiplanicie de la Cordillera Oriental. Fundación de Investigaciones Arqueológicas Nacionales. Banco de la República, Bogotá
Correal G, Pinto M (1983) Investigaciones arqueológicas en el Municipio de Zipacón, Cundinamarca. Fundación de Investigaciones Arqueológicas Nacionales. Banco de la República, Bogotá
Correal G, van der Hammen T (1977) Investigaciones arqueológicas en los abrigos rocosos del Tequendama. 11,000 años de prehistoria en la Sabana de Bogotá. Banco Popular, Bogotá
Delgado ME (2007) El impacto de la colonización Española: Evidencias paleopatológicas e isotópicas de cambios en la salud oral y reducción en la diversidad de la dieta en sociedades Coloniales Nativas del Sur Occidente de Colombia. Revista de Estomatología 15:17–37
Delgado ME (2012a) Mid and late Holocene population changes at the Sabana de Bogotá (northern South America) inferred from skeletal morphology and radiocarbon chronology. Quat Int 256:2–11
Delgado ME (2012b) Dental and craniofacial diversity in the northern Andes, and the early peopling of South America. In: Miotti L, Saleme M, Flegenheimer N, Goebel T (eds) Southbound: late Pleistocene peopling of Latin America. Center for the Study of the First Americans, Department of Anthropology, University of Texas A&M University, Texas, pp. 33–38
Delgado ME (2015) Variación Dental y Craneofacial en el Norte de los Andes durante el Pleistoceno y el Holoceno. Su relevancia para la Discusión de la Colonización Temprana de Sudamérica, Ph.D. Dissertation, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Buenos Aires
Delgado ME, Langebaek C, Aristizabal L, Tykot RH, Johnson L (2014) Indicadores bioquímicos de dieta en Tibanica, un poblado muisca tardío en la Sabana de Bogotá (Colombia). Isótopos estables (δ 13Ccol, δ 13Cap, δ 15N) y elementos traza (Ba, Sr). In: Luna L, Aranda C, Suby J (eds) . Avances Recientes de la Bioarqueología Latinoamericana, Grupo de Investigación en Bioarqueología, pp. 103–127
Delgado ME, Aceituno FJ, Loaiza N (2015a) Multidisciplinary studies on the human-environment interaction during the initial peopling of the Americas. Quat Int 363:1–3
Delgado M, Aceituno FJ, Barrientos G (2015b) 14C data and the early colonization of Northwest South America: a critical assessment. Quat Int 363:55–64
Emery K, Wright L, Schwarcz H (2000) Isotopic analysis of ancient deer bone: biotic stability in collapse period Maya land-use. J Archaeol Sci 27:537–550
Enciso B (1996) Fauna asociada a tres asentamientos muiscas del sur de la Sabana de Bogotá. Siglos VIII-XIV D.C. In: Therrien M, Enciso B (eds) Bioantropología de la Sabana de Bogotá. Instituto Colombiano de Antropología e Historia
Froehle AW, Kellner CM, Schoeninger M (2012) Multivariate carbon and nitrogen stable isotope model for the reconstruction of prehistoric human diet. Am J Phys Anthropol 147:352–369
García JL (2012) The foods and the crops of the muisca: a dietary reconstruction of of the intermediate chiefdoms of Bogotá and Tunja. Master Dissertation, Department of Anthropology, University of Central Florida, Orlando, Florida, Colombia
Gnecco C (2000) Ocupación temprana de bosques tropicales de montaña. Editorial Universidad del Cauca, Popayán
Gnecco C (2003) Against ecological reductionism: late Pleistocene hunter-gatherers in the tropical forest of northern South America. Quat Int 109-110:13–21
Gnecco C, Mora S (1997) Late Pleistocene/early Holocene tropical forest occupations at San Isidro and Peña Roja. Colombia Antiquity 71:683–690
Gómez J (2012) Análisis de marcadores óseos de estrés en poblaciones del Holoceno medio y tardío inicial de la Sabana de Bogotá. Revista Colombiana de Antropología 48:143–168
Gómez A, Berrío J, Hooghiemstra H, Becerra M, Marchant R (2007) A Holocene pollen record of vegetation change and impact from Pantano de Vargas, and intra-Andean basin of Duitama. Colombia Rev Palaeobot Palynol 145:143–157
Guillet B, Faivre A, Mariotti JK (1988) The 14C and 13/12C ratios of soil organic matter as a means of studying the past vegetation in intertropical regions: examples from Colombia (South America). Palaeogeogr Palaeoclimatol Palaeoecol 65:51–58
Henderson H, Ostler N (2005) Muisca settlement organization and chiefly authority at Suta, Valle de Leyva, Colombia: a critical appraisal of native concepts of house for studies of complex societies. J Anthropol Archaeol 24:148–178
Hurt WR, van der Hammen T, Correal G (1977) The El Abra rock shelters, Sabana de Bogotá, Colombia, South America. Occasional papers and monographs, vol 2. Bloomignton Indiana University Museum.
Ijzereef G (1978) Faunal remains from the el Abra Rockshelters (Colombia). Palaeogeogr Palaeoclimatol Palaeoecol 25:163–177
Katzenberg MA (2008) Stable isotope analysis: a tool for studying past diet, demography and life history. In: Katzenberg M, Saunders SR (eds) Biological anthropology of the human skeleton. John Willey & Sons, Inc, pp. 305–327
Kellner C, Schoeninger M (2007) A simple carbon isotope model for reconstructing prehistoric human diet. Am J Phys Anthropol 133:1112–1127
Kohn M, Mckay M, Knight K (2005) Dining in the Pleistocene—who’s the menu? Geology 33:649–652
Kruschek MH (2003) The evolution of the Bogotá chiefdom: a household view. Ph.D dissertation, Faculty of Arts and Sciences. Department of Anthropology, University of Pittsburgh, Pittsburgh
Lahr M, Foley R (1998) Towards a theory of modern human origins: geography, demography, and diversity in recent human evolution. Yearb Phys Anthropol 41:137–176
Langebaek CH (1987) Mercados, poblamiento e integración étnica entre los muiscas, siglo XVI. Colección Bibliográfica Banco de La República, Bogotá
Marchant R, Behling H, Berrío J, Cleef A, Duivenvoorden J, Hooghiemstra H, Kuhry P, Melief R, Schreve-Brinkman E, van Geel B, van der Hammen T, van Reenen G, Wille M (2002) Pollen-based biome reconstructions for Colombia at 3000, 6000, 9000, 12000, 15000 and 18000 14C yr ago: late quaternary tropical vegetation dynamics. J Quat Sci 17:113–129
Martínez I (2012) La salud en poblaciones Muiscas durante la transición del periodo temprano al tardío, estudio comparativo. Departamento de Antropología, Facultad de Ciencias Humanas, Universidad Nacional de Colombia, Master Dissertation
Monsalve JG (1985) A pollen core from the hacienda Lusitania. Pro Calima. 4, 40–44. Basilea: Periodische Publikation der Vereinigung Pro Calima.
Mora G, Pratt LM (2002) Carbon isotopic evidence from paleosols for mixed C3/C4 vegetation in the Sabana de Bogotá basin. Colombia Quat Sci Rev 21:985–995
Morcote G (1996) Evidencia arqueobotánica de cultígenos presentes en grupos muiscas de la Sabana de Bogotá en los siglos VIII y IX. In: Enciso B (ed) Therrien M. Bioantropología de la Sabana de Bogotá, Instituto Colombiano de Antropología e Historia, pp. 59–83
Moreano AC (2011) Análisis arqueozoológico de elementos óseos de fauna recuperados en el complejo funerario muisca Tibanica (Soacha). Universidad de los Andes, Bogotá, Informe Departamento de Antropología
Nieuwenhuis CJ (1998) Unattractive but effective: unretouched pointed flakes as projectile points? A closer look at the Abriense and Tequendamiense artefacts. In: Plew MG (ed) Explorations in American archaeology. Essays in Honor of Wesley R. Hurt. University Press of America, Lanham, pp. 133–163
Nieuwenhuis CJ (2002) Traces on tropical tools: a functional study of Chert artifacts from Preceramic sites in Colombia. Ph.D. dissertation. Leiden University, Archaeological Studies Leiden University. No. 9. Faculty of Archaeology, University of Leiden, Leiden
Parra R (2001) Identificación de fitolitos en el cálculo dental de individuos prehispánicos de Tunja (Boyacá) y Soacha (Cundinamarca). In: Rodríguez JV (ed) Los Chibchas. Adaptación y diversidad en los Andes Orientales de Colombia. Universidad Nacional de Colombia, Bogotá, pp. 237–249
Peña G, Pinto M (1996) Mamíferos más comunes en sitios precerámicos de La Sabana de Bogotá: Guía ilustrada para arqueólogos. Editorial Guadalupe, Bogotá
Pinto M (2003) Galindo, un sitio a cielo abierto de cazadores-recolectores en la Sabana de Bogotá. Fundación de Investigaciones Arqueológicas Nacionales. Banco de la República, Bogotá
Piperno D, Pearsall D (1998) The origins of agriculture in the lowland Neotropics. Academic Press, San Diego
Price TD, Burton JH (2011) An introduction to archaeological chemistry. Springer, New York
R Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3–900051–07-0. Available online at http://www.R-project.org/.
Rodríguez JV (2006) Las enfermedades en las condiciones de vida prehispánicas de Colombia. Universidad Nacional de Colombia, Departamento de Antropología. Facultad de Ciencias Humanas
Rodríguez JV (2011) Los Chibchas: hijos del sol, la luna y los Andes. Universidad Nacional de Colombia, Instituto de Desarrollo Urbano
Rodríguez JV, Cifuentes A (2005) Un yacimiento formativo ritual en el entorno de la antigua laguna de La Herrera, Madrid. Cundinamarca Maguaré 19:103–131
Rodríguez D, Colantonio S (2015) Biological affinities and regional microevolution among pre-Hispanic communities of Colombia's northern Andes. Anthropol Anz 72:141–168
Schoeninger MJ (2009) Stable isotope evidence for the adoption of maize agriculture. Curr Anthropol 50:633–640
Simón P (1981) [1625] Noticias historiales de la conquista de tierra firme en las Indias Occidentales. Bogotá, Banco popular
Somerville AD, Froehle A (2011) New approaches to investigating ancient Maya diet: a three-isotope model for the reconstruction of dietary protein sources. Am J Phys Anthropol 144(Suppl 52):280
Stuiver M, Reimer PJ (1993) Extended 14C database and revised CALIB radiocarbon calibration program. Radiocarbon 35:215–230
Szpak P (2014) Complexities of nitrogen isotopes biogeochemistry in plaint-soil systems: implications for the study of ancient agricultural and animal management practices. Front Plant Sci 5:1–19
Tykot RH (2004) Stable isotopes and diet: you are what you eat. In: Course C, Martini M, Milazzo M, Piacentini M (eds) Proceedings of the International School of Physics “Enrico Fermi”. IOS press, Amsterdam, pp. 433–444
Tykot RH (2006) Isotope analyses and the histories of maize. In: Staller JE, Tykot RH, Benz BF (eds) Histories of maize: multidisciplinary approaches to the prehistory, linguistics, biogeography, domestication, and evolution of maize. Academic Press (Elsevier), Cambridge, pp. 131–142
Tykot RH (2014) Bone chemistry and ancient diet. In: Smith C (ed) Encyclopedia of global archaeology. Archaeological science section. Springer, Berlin, pp. 931–941
Tykot RH, Falabella F, Planella MT, Aspillaga E, Sanhueza L, Becker C (2009) Stable isotopes and archaeology in Central Chile: methodological insights and interpretive problems for dietary reconstruction. Int J Osteoarch 19:156–170
van der Hammen T (1992) Historia, ecología y Vegetación. Banco Popular, Bogotá
van der Hammen T, Hooghiemstra H (1995) The el Abra stadial, a younger Dryas equivalent in Colombia. Quat Sci Rev 14:841–851
van der Hammen T, Correal G, van Klinken GJ (1990) Isótopos estables y dieta del hombre prehistórico en la Sabana de Bogotá. Boletín de Arqueología, Fundación de Investigaciones Arqueológicas Nacionales
Vélez M, Hooghiemstra H, Metcalfe S, Willie M, Berrío J (2006) Late glacial and Holocene environmental and climatic changes from a limnological transects through Colombia, northern South America. Palaeogeogr Palaeoclimatol Palaeoecolo 234:81–96
Walker PL, Bathurst R, Richman R, Gjerdrum T, Andrusko V (2009) The causes of porotic hyperosthosis and criba orbitalia: a reappraisal of the iron-dieficiency-anemia hypothesis. Am J Phys Anthropol 139:109–125
Acknowledgments
I would like to thank German Mora and Arnoud Boom for sharing their data on carbon isotopes from paleoenvironmental researches. Thanks to Javier Giraldo (Universidad del Cauca, Colombia) and Alejandra Ortíz (Arizona State University, USA) for their useful comments on early drafts of this paper. Thanks to Alexandra Delgado for help with the English version of the manuscript. This research was partially supported by the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, Argentina) and Fundación de Investigaciones Arqueológicas Nacionales (FIAN, Colombia) (project no. 409-2010).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Supplementary Table 1
(XLS 52 kb)
Supplementary Table 2
(XLS 48 kb)
Rights and permissions
About this article

Cite this article
Delgado, M. Stable isotope evidence for dietary and cultural change over the Holocene at the Sabana de Bogotá region, Northern South America. Archaeol Anthropol Sci 10, 817–832 (2018). https://doi.org/10.1007/s12520-016-0403-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-016-0403-3