
Abstract
This paper discusses isotopic analyses carried out on human bone samples corresponding to complex hunter-gatherers from the Paraná River basin in northeastern Argentina. Based on the δ15N and δ13C values obtained, the dietary patterns of 23 individuals recovered from pre-Hispanic archaeological sites of the Late Holocene are characterized. These sites are associated with the archaeological unit generically called Goya-Malabrigo, which is identified in South America over the entire middle basin of the Paraná River and on a part of the lower river. The values obtained show diets based on depleted δ13C proteins linked to the C3 photosynthetic pathway, which is in turn consistent with the isotopic values detected in the main food sources of these human groups: freshwater fishes and continental mammals. The δ15N values and the results of a multivariate model also indicate a low consumption of plant foods. Although archaeobotanical information from the area has reported maize and other cultigens, the diet of the individuals studied was based on animal proteins, especially on freshwater fish. Moreover, the intake of maize was not isotopically detectable in the human bones analyzed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The middle and lower Paraná River basin was occupied at least within the time span of 2400–500 years BP by human groups whose economic base was centered on hunting, fishing, and gathering. These activities were framed within a context of substantial and increasing social and technological complexity, with numerous adaptations driven by human population density. These societies developed certain features such as central residential camps with some stability, cemeteries used for generations, storage systems, territoriality, ownership of territories and productive patches, intensification in the exploitation of the environment, and development of a complex trade network (Ceruti 2003; Loponte and Acosta 2006; Loponte 2008). Among the intensification-oriented behaviors, we must emphasize the extensive use of pottery vessels to process food and enhance the ability to extract nutrients by boiling (Acosta 2005; Loponte 2008). The use of pottery for this purpose was sufficiently intense that almost all of the pottery recorded shows evidence of fatty acids associated with vegetable and/or animal foods (Naranjo et al. 2010; Ottalagano 2013; Pérez et al. 2013). These populations also developed a complex kit of bone and stone artifacts, including a sophisticated weapon system that included the bow and arrow, hand-held spears, spear-throwers, bola stones, detachable harpoons, and fishing nets as well as traps (Buc 2010; Ceruti 2003; Loponte and Acosta 2006; Loponte 2008).
Freshwater fishes were an important part of the diet, especially towards the end of the Late Holocene. The subsistence systems also included the exploitation of deer (Blastocerus dichotomus; Mazama gouazoubira; Ozotoceros bezoarticus), freshwater clams, and small-to-medium rodents (Myocastor coypus, Cavia aperea) (Acosta 2005; Acosta et al. 2010, 2013a, b; Arrizurieta et al. 2010; Barboza and Píccoli 2013; Bonomo et al. 2011a; Ceruti and González 2007; Cione and Tonni 1978; Loponte and Acosta 2004; Loponte 2008; Loponte et al. 2012; Musali 2010; Nóbile 1993; Ottalagano et al. 2015; Pérez Jimeno 2007; Salemme et al. 1987; Salemme and Tonni 1983; Santiago 2004; Sartori and Colasurdo 2012; Tonni et al. 1985, among others). Also, one large rodent (Hydrochaeris hydrocaheris) was exploited, although seemingly only marginally since the archaeological representation is small, and hunting of this animal largely appears to have had a symbolic meaning, especially in the Lower Paraná River basin (Acosta 2005; Loponte 2008; Salemme 1987). Agroforestry of local palms and the development of sporadic and small gardens have also been postulated (Acosta and Ríos Román 2013; Loponte 2008). In relation to these activities, the archaeological presence of phytolith and starch grains assignable to species such as Cucurbita sp. (squash), Prosopis sp. (algarrobo), Phaseolus sp. (beans), Ipomea sp. (sweet potato), Zea mays (maize), and palms (Syagrus romanzoffiana) has been reported (Acosta et al. 2013b; Bonomo et al. 2011a, b, c; Colobig and Ottalagano 2015; Cornero and Rangone 2015; Sánchez et al. 2013; Zucol and Loponte 2008).
One of the archaeological units corresponding to complex hunter-gatherers in the region is generally known as “Goya-Malabrigo” (Ceruti 2003; Rodríguez 2001; Serrano 1946, 1972), and although this unit is still somewhat ill-defined, its main diagnostic feature is the presence of pottery with zoomorphic appendages, usually representing different types of birds (especially parrots). These figures may have constituted symbols of group identity within a context of growing social complexity (Ottalagano 2013). On numerous occasions, archaeologists studying the region have argued for the possibility that these societies had developed small-scale horticultural practices, often based on ethnohistorical sources but also as suggested by certain archeological features (Ceruti 2003; Loponte 2008; Rodríguez 2001; Serrano 1972). Archaeobotanical identification of micro-remains from Zea mays and other cultigens such as Cucurbitaceae, Cannaceae, and Phaselous on ceramic and stone artifacts corresponding to this archaeological unit (e.g., Bonomo et al. 2011a, b, c; Colobig and Ottalagano 2015; Cornero and Rangone 2015; Sánchez et al. 2013) has recently led some authors to suggest the existence of a mixed economy, which would imply a significant degree of dependence on plant products (Bonomo et al. 2011a, b). However, the current data are insufficient to allow assessment of the existence of dependency on cultivated resources in “Goya-Malabrigo” contexts.
Over three decades ago, stable isotope analysis of carbon and nitrogen began to be used to infer human dietary patterns (e.g., Ambrose and DeNiro 1986; DeNiro and Schoeninger 1983; Van der Merwe and Vogel 1978; Vogel and Van der Merwe 1977). Unlike archaeofaunal, archaeobotanical, or fatty acid analysis, isotopic studies provide a direct source of data on human diets. This is because the isotopic composition of human tissues is derived from the daily food intake of the individual and then remains constant after death. Therefore, measurement of isotopic values facilitates the identification of different resources present in the diet and the ratios of their consumption. In the lower Paraná River basin, isotopic studies began over a decade ago in relation to other archaeological units (Acosta and Loponte 2001; Loponte and Acosta 2007; Loponte 2008; Loponte et al. 2015), with only one study related to Goya-Malabrigo contexts (Loponte and Kozameh 2009). In that analysis, no evidence of maize consumption was found, at least as isotopically detectable.
The aim of the present paper is therefore to introduce and discuss new δ15N and δ13C isotopic data obtained from human remains recovered from archaeological sites associated with the Goya-Malabrigo archaeological unit, located in the middle and lower Paraná areas, in order to characterize the dietary patterns existing there and to assess any eventual dependence upon plant foods. To improve the analytical strength of this analysis, we have also incorporated the values obtained from human remains recovered from Guaraní sites located in the lower Paraná River basin. These last contexts belong to Amazonian horticulturist groups, who are known to have had a mixed diet with a significant intake of maize and other cultivated plants as well as consumption of animal food products derived from hunting and fishing activities (Loponte and Acosta 2013; Loponte et al. 2015). These data thus provide local isotopic values for an economy that had a significant horticultural component.
Environment
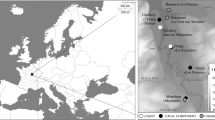
The study area comprises a section of the middle Paraná River basin as well as its lower section, covering a total length of 700 km along the watercourse (Fig. 1). The Paraná is an extensive, high-volume South American river that originates from rainfall in the tropics that is mostly produced by the South American summer monsoon (SASM), which is associated with the intertropical convergence zone (ITCZ) (Gan et al. 2004; Vuille et al. 2003; Zhou and Lau 1998). The tropical environment of the headwaters extends as a vector of moisture and temperature along the river basin, allowing for development of the southernmost subtropical wetland in South America, with the presence of a multi-layered riparian forest with a continuous 8–15-m canopy, associated with riverine prairies highly fragmented by streams (Cabrera 1976; Kalesnik et al. 2008; Roesler and Agostini 2012). In the middle Paraná, this wetland environment is narrow and circumscribed to the river’s banks and the small islands within it. In the lower Paraná, however, for a distance of 320 km, the wetlands become 60-km wide on average, cutting across the temperate environment of the Pampean steppe. This facilitates the dispersal of flora and fauna and extends the humid conditions from more northern areas, creating a giant azonal and hierarchical patch of resources (Burkart et al. 1999; Loponte 2008). Most of the vegetation cover and the main dietary energy sources in the Paraná fluvial system have a C3 pattern (Madanes et al. 2013; Marchese et al. 2014), and therefore virtually all of the region’s fishes and semi-aquatic and terrestrial mammals also possess a C3 pattern. In Table 1, we present the isotopic data available for the main preys considered in this paper. The data come mostly from archaeological samples except those corresponding to capybara, which are modern. Due to the Suess effect, which is defined as a global decrease in the 13C content of atmospheric carbon dioxide, primarily because of fossil fuel burning over the last 150 years that started with the industrialization process (Francey et al. 1999; Indermühle et al. 1999), we applied a time-dependent correction of −0.005 ‰ per year between 1860 and 1960 and −0.022 ‰ per year since 1960 (Chamberlain et al. 2005) until 2013, when the modern samples were collected. This correction provides an approximate value of δ13C for comparing ancient and modern ecosystems.

Location of the archaeological sites included in this study: 1, Arroyo Las Mulas I; 2, Arroyo Arenal I; 3, La Palmera II; 4, Los Tres Cerros I; 5, Los Marinos; 6, Escuela 31
Stable isotopes
As mentioned above, one of the key aspects for understanding the success of isotopic studies is that this type of data provides information about an individual’s diet for the last 7–10 years of life, especially when these analyses are carried out on bone tissue (Ambrose 1993). Numerous studies have shown that δ13C values measured in an individual’s collagen (δ13Cco from here forward) are enriched by about 5 ‰ compared with the diet. Thus, herbivorous diets based on resources with a C3 photosynthetic pathway show values of approximately −21 ‰, while monoisotopic C4 diets present values of around −7 ‰ (DeNiro and Epstein 1978a, b; Koch 2007; Krueger and Sullivan 1984; Lee-Thorp et al. 1989; Vogel and Van der Merwe 1977). In the inorganic fraction (δ13Cap from here forward), monoisotopic diets present values of −13.5 and −3 ‰ and below, respectively (Kellner and Schoeninger 2007; Lee-Thorp and Van der Merwe 1987; Sullivan and Krueger 1981).
Various studies have also shown that carbon values from collagen more closely reflect the content of the protein intake in the diet, while those from apatite have a closer relationship with the total diet (protein + fat + carbs) (Ambrose and Norr 1993; Katzenberg 2008; Krueger and Sullivan 1984; Tykot 2006). This leads to the conclusion that differences between the two carbon sources (δ13Cco-ap, or Δ13C from here forward) largely reflect the plant component of the diet. This model developed by Krueger and Sullivan (1984) is used as an indicator of the trophic level of diets and/or to identify marine components in them (Ambrose and Norr 1993; Ambrose et al. 1997, 2003; Ambrose and Krigbaum 2003; Harrison and Katzenberg 2003; Lee-Thorp et al. 1989; Loftus and Sealy 2012; Norr 1995; Pate 1995; Rand et al. 2013; Roksandic et al. 1988; Waterman et al. 2015; White et al. 2006). Larger differences are typical of herbivores, with mean values of 6.8 ± 1.4 ‰, while carnivorous diets are below 4.0 ± 1.0‰ (Ambrose and Norr 1993; Ambrose and Krigbaum 2003; Clementz et al. 2009; Krueger and Sullivan 1984; Lee-Thorp et al. 1989; Loftus and Sealy 2012). However, there are no absolute values for demarcating thresholds, and instead these should be defined empirically according to the particular context analyzed (Ambrose and Norr 1993; Kellner and Schoeninger 2007). These thresholds also depend upon the combination of photosynthetic proteins and carbohydrates ingested (Ambrose and Norr 1993; Ambrose and Krigbaum 2003; Ambrose et al. 2003; Cormie and Schwarcz 1996; Kellner and Schoeninger 2007; Metges et al. 1990; Stevens et al. 2008; Van Klinken et al. 1994). However, the spacing of values related with trophic levels has been supported empirically in many studies, and these analyses are reinforced when used in conjunction with nitrogen values. In fact, the δ15N increases at a rate of 3–4 ‰ as the body incorporates foods of higher trophic levels (Bocherens and Drucker 2003; Hedges and Reynard 2007; Schoeninger and DeNiro 1984). Thus, the combination of both carbon and nitrogen isotopes is used to estimate the trophic position of individuals (Ambrose et al. 1997, 2003; Ambrose and Norr 1993; Froehle et al. 2010, 2012; Loftus and Sealy 2012; Tykot 2006; Warinner and Tuross 2009).
Based on laboratory experiments, Kellner and Schoeninger (2007) have argued that isotopic analysis is more reliable if regression lines are used on both values of δ13Cco and δ13Cap. Some authors have suggested that this model has methodological limitations in relation to metabolic aspects and the intake of other nutrients that are not included in the analysis of controlled diets, as well as a lack of consideration of nitrogen values (Froehle et al. 2012; Loftus and Sealy 2012; Warinner and Tuross 2009). To overcome some of these problems, Froehle et al. (2012) developed a multivariate model incorporating the δ15N values from calculation of discriminant functions, which are then plotted in clusters of known diets. This allows more accurate evaluation of the archaeological diets, especially those that incorporate maize (a C4 plant), which has played an especially role in isotopic studies (Ambrose et al. 2003; Balasse et al. 1999; Boyd et al. 2008; Calo and Cortes 2009; Gheggi and Williams 2013; Gil et al. 2010; Hart et al. 2011; Froehle et al. 2012; Killian et al. 2012; Laguens et al. 2009; Metcalfe et al. 2009; Rand et al. 2013; Scherer et al. 2007; Slovak and Paytan 2009; Smalley and Blake 2003; Staller et al. 2006; Stronge 2012; Tykot 2002, 2004, 2006; White et al. 2006).
Materials and methods
In this paper, we included isotopic values for 23 individuals recovered from six Goya-Malabrigo archaeological contexts (Fig. 1). These sites are located along the middle and lower Paraná River, within a chronological range between 460 ± 50 and 1807 ± 47 years 14C BP (Table 2). Human remains from the Arroyo Las Mulas I and Arroyo Arenal I sites were recovered by Ceruti (2003), while those from La Palmera II and Escuela 31 sites were excavated by Ottalagano et al. (2015) and by Loponte (unpublished work), respectively. For the isotopic analysis, fragments of long bones, ribs, and skull were selected, which were seen as preferable as elements that offer a higher proportion of cortical tissue and since they are more suitable for collagen extraction (Ambrose 1993; Falabella et al. 2007). We also preferably considered burials with a high level of integrity, although in a few cases we also had to resort to isolated human remains recovered from the archaeological layers. Most of the analyses were performed in the USA, mainly at the SIRFER laboratory at the University of Utah, although others were carried out at the University of Georgia (CAIS) and at the University of Arizona (Environmental Isotope Laboratory). A few samples were analyzed at the Institute of Isotope Geology and Geochronology in Buenos Aires (INGEIS). We have incorporated into our analysis four isotopic values of δ13Ccol, δ13Cap, and δ15N previously published by Loponte and Kozameh (2009) and two δ13Cco values obtained as part of the AMS dating performed at the NSF Arizona AMS Lab (Table 2). There are six direct AMS radiocarbon dates available from different individuals. The rest of the samples were associated chronologically with their dated contexts (Table 2).
Results and discussion
Carbon values for individuals associated with the Goya-Malabrigo archaeological unit show a C3 monoisotopic trend, with depleted δ13Cco values with a mean of −19.4 ± 1.6 ‰ with low variability (CV = 8.2 %) (Table 2). Similarly, the average δ13Cap of −13 ± 1.2 ‰ (CV = 9 %), although we currently have a small sample for apatite analysis, reflects an almost pure monoisotopic diet (Kellner and Schoeninger 2007; Sullivan and Krueger 1981; Lee-Thorp and Van der Merwe 1987), while the high levels of nitrogen (11.8 ± 1.1 ‰; CV = 9.2 %) indicate a substantially carnivorous diet (Ambrose 1993) (Table 2). Thus, the δ13C values found in the individuals analyzed correspond more to C3 protein intake rather than consumption of C3 plants.
Regarding spatial variations between the two analyzed sectors of the basin, there is little difference in the confidence intervals. The values from collagen are slightly enriched in the samples from the middle Paraná (CI = −19.9 ‰, −18.2 ‰) compared to those from the lower Paraná (CI = −21.0 ‰, −18.9 ‰). In turn, nitrogen values are similar (CI = 11.6–12.3 ‰ in middle Parana, CI = 11.0–13.4 ‰ in lower Paraná, excluding the anomalous value of individual EIL-1021; including this value the LL is 9.5 ‰, see Table 2).
The samples can also be grouped into three chronological blocks to assess the impact of plant resources in the diet, including maize: an early period (1807 ± 47 to 1732 ± 50 14C BP); an intermediate period (1056 ± 47 to 950 ± 120 14C BP); and a late period (775 ± 85 to 460 ± 50 14C BP). The first has a value of \( \overline{\mathrm{x}} \) 3 = δ13Cco −20.1 ± 1.10 ‰, the second \( \overline{\mathrm{x}} \) 12 = −19.4 ± 1.3 ‰ (CI = −20.3 ‰, −18.6 ‰), and the last \( \overline{\mathrm{x}} \) 8 = −19.1 ± 0.7 ‰ (CI = −20.7 ‰, −17.5 ‰). There are therefore no significant differences seen among these values (Fig. 2). The nitrogen values, in turn, show an increase in the intake of animal-derived protein over time (Fig. 3), which could indicate an intensification of fishing-based resource exploitation, a trend that has been verified in other archaeological units in the area based on faunal analysis (Loponte et al. 2012).

Temporal trends for the carbon values from collagen

Temporal trends for the nitrogen values
The mean carbon values from both collagen and apatite show hardly any fractionation with respect to the average values for prey. However, a more detailed analysis is required here. The large rodent known as capybara (Hydrochaeris hydrocaheris), which shows the most enriched diet of all of the prey considered, has a very low representation in the faunal assemblages: At the Arroyo Las Mulas I site, it is practically absent (Serrano 1946); at the La Palmera II site, it makes up only 0.5 % of the NISP (Ottalagano et al. 2015); and at the Escuela 31 site, it is observed in an equally low proportion (detailed quantification does not exist for the rest of the sites considered).Footnote 1 Brocket deer (Mazama sp.) must also be considered to be an occasional resource, as these remains are either scarce or not recorded at sites in the area (see Acosta 2005; Arrizurieta et al. 2010; Bonomo et al. 2011c; Loponte 2008; Nóbile 1993; Ottalagano et al. 2015).
Excluding these two taxa mentioned, the average δ13Cco for the most common preys is −20.0 ± 1.5 ‰, a magnitude that also shows hardly any fractionation compared to the human remains analyzed (−19.4 ± 1.6 ‰), where the expected difference between the isotopic values of prey and predator collagen ranges from 0 to 2 ‰ (Bocherens and Drucker 2003). In turn, for this discussion, what is really crucial are the isotopic values for the most abundant fishes from the Paraná River system. While Table 1 shows the average for two omnivorous Siluriformes taxa (Pterodoras granulosus and Pimelodus sp.) recovered from two archaeological sites in the lower Paraná, the faunal evidence indicates that the most heavily exploited species was Prochilodus lineatus, a bottom-feeding species that represents more than 50 % of the fish biomass of the Paraná River and which is considered to be a key species for the fluvial system (Loponte 2008). This characiform has extremely thin and fragile bones and is therefore notably under-represented in the archaeological record because of problems with preservation and identification (Acosta 2005; Loponte 2008; Musali 2010). A study performed on this species in northern areas of the Paraná basin yielded values for muscle tissue ranging between δ13Cco −24.8 ± 3 and −31.5 ± 2.6 ‰ (Lopes et al. 2007), with an average of −27.5 ‰ (Suess-corrected δ13C −26.1 ‰).Footnote 2 Another analysis conducted by Marchese et al. (2014) on individual specimens captured in the middle Paraná yielded a similar average (−27.3 ± 3.1 ‰, Suess-corrected δ13C −25.7 ‰).Footnote 3 Given that all of these values obtained from muscle tissue are depleted by 1.1–1.6 ‰ with respect to bone collagen in fishes (Sholto-Douglas et al. 1991), an approximate mean of δ13C = −24.5 ± 3 ‰ can be considered as an accepted average value for bone collagen.Footnote 4 Other fishes that are also common in the archaeological sites of the region, such as Leporinus obtusidens, Pimelodus maculatus, Hoplias malabaricus, Salminus brasiliensis, Ageneiosus brevifilis, and Pseudoplatystoma reticulatum (Acosta 2005; Arrizurieta et al. 2010; Loponte 2008; Loponte et al. 2012; Musali 2010), show a depleted δ13C average of −26.7 ± 1 ‰ in muscle tissue (calculated based on Saigo et al. 2015: Table 2), and thus a value of −25.3 ‰ can be considered for bone collagen (Suess-corrected −23.7 ‰). Bulk value of collagen of fish (−24.1 ‰)Footnote 5 therefore provides monoisotopic C3 signals as do the other preys (−20.1 ‰), but substantially depleted, and this may help to understand the carbon levels found in the human samples. Some authors (i.e., Bocherens and Drucker 2003) have suggested that to understand the values of collagen of the predators, it is necessary to consider all the significant preys in the archaeological record. Although we do not know exactly the proportion between terrestrial mammals and fish resources in the diet of individuals analyzed, considering an average between the two types of resources, we can explain the values of collagen registered in humans within the ranges that are reasonable to understand the observed isotopic fractionation between preys and humans (−22.1 vs. −19.4 ‰, respectively), including here probably also some minor intake of C3 plants.
A similar situation occurs in the case of nitrogen. The human samples analyzed show enriched δ15N values (\( \overline{\mathrm{x}} \) = 11.8 ± 1.1 ‰) that cannot be explained by consumption of the preys listed in Table 1 (\( \overline{\mathrm{x}} \) = 6 ± 1.5 ‰). Although is also likely to other species with depleted δ13Cco values contributing in a significant manner to human subsistence, these human nitrogen levels could be explained in part by high consumption of fish. Indeed, P. lineatus shows an average of 9.13 ± 0.82 ‰ (in muscle) (Marchese et al. 2014) for individual fish captured in the middle Paraná and between 10.7 ± 2.2 and 11.9 ± 0.7 ‰ for specimens obtained in the upper Paraná (Lopes et al. 2009), for an average of 10.6 ± 1.2 ‰ for this species. Thus, the nitrogen value for P. lineatus would be around 8.3 ± 1.2 ‰ in bone collagen since it is depleted by 2.3–2.4 ‰ compared to muscle tissue (Sholto-Douglas et al. 1991). The remaining fish species mentioned above, which are present in regional archaeofaunal assemblages, show an average of 10.4 ± 1.1 ‰ (calculated on the values obtained from the dorsal muscle, Saigo et al. 2015: Table 2), averaging ∼8 ‰ in bone collagen (see also Bocherens and Drucker 2003; Hedges and Reynard 2007; Schoeninger and DeNiro 1984). Considering the nitrogen level in preys listed in Table 1 (6 ‰) and other fishes (∼8 ‰), an average of 7 ‰ is an approximate value for nitrogen signal intake for humans. The resulting isotopic fractionation (∼4.8 ‰) is within the known range between predators and preys (Bocherens and Drucker 2003).
Therefore, the enriched nitrogen values and depleted carbon levels observed among the hunter-gatherer individuals analyzed in the present study can be explained within the framework of a diet with a strong emphasis on fish consumption and, certainly, limited direct consumption of plant resources.
It is estimated that human individuals with bone collagen values ranging between −17.0 and −14.0 ‰ correspond to a mixed diet, which could have been made up of 25–50 % C4 plants and/or C4 plant-based animal protein (Coltrain and Leavitt 2002). The only individual who falls within these parameters comes from Arroyo Arenal I site (δ13Cco = −15.9 ‰, SIRFER 13-8008 in Table 2). However, this individual has a high nitrogen value, similar to the rest of the population, also suggesting in this case a limited intake of plant products.
The general trend towards a C3 diet based on animal resources is clearly observed when the values from the human individuals analyzed from Goya-Malabrigo sites are plotted together with those for local herbivorous and omnivorous preys and human individuals recovered from Guaraní sites of the area (Amazonian horticulturalists), whose diets actually included a substantial element of cultigens, including maize (Figs. 4 and 5). To further understand the incidence of maize within these two populations, and for comparative purposes, we have also plotted the values published by Sommerville et al. (2013) for individuals from the southern Maya lowlands throughout the Classic Period, where there was a substantial dependence upon cornFootnote 6 (Fig. 4).

Values for δ13Cco versus δ15N for individuals from societies with different economic systems, complex hunter-gatherers (Goya-Malabrigo), horticulturalist economy (Guaraní), and agriculture-based societies (Maya), along with the main preys exploited along the middle and lower Paraná River by pre-Columbian human populations

Values for δ13Cco and δ13Cap following the model proposed by Kellner and Schoeninger (2007) for individuals with a horticulturalist economy (including maize) (Guaraní) and complex hunter-gatherers (Goya-Malabrigo). It should be noted that the graph is based on the six apatite values currently available
If the discriminant functions developed by Froehle et al. (2012) are used to consider the nitrogen values, the Goya-Malabrigo individuals analyzed fall into the box of diets composed of 100 % protein and energy intake related with a C3 photosynthetic pathway. In contrast, the Guaraní samples fall into an intermediate position between the box for C3/C4 energy and protein intake and the box representing the highest consumption of C4 energy (Fig. 6). There is also an isotopic gap seen when these two populations are plotted by their means plus/minus one standard deviation (Fig. 7). Finally, the carnivorous diet of the human individuals recovered at Goya-Malabrigo sites is also quite clear when a typical carnivore from the area (the jaguar, Pantera onca) is added to the plot (Fig. 7).

F1 and F2 discriminant function values from individuals recovered at Goya-Malabrigo and Guaraní sites, plotted against previously generated dietary clusters (see Froehle et al. 2012)

The isotopic signals observed in the analyzed individuals corresponding to the Goya-Malbrigo archaeological unit, as well as data provided by archaeobotanical studies in relation to these groups (see Bonomo et al. 2011a, b, c; Colobig and Ottalagano 2015; Cornero and Rangone 2015; Sánchez et al. 2013), are consistent with the observations of sixteenth-century travellers in reference to the post-Hispanic societies then occupying the Paraná River areas where the highest concentrations of Goya-Malabrigo sites have now been recorded. That is, a low incidence of vegetable foods, use of small quantities of maize, and the importance of animal resources, especially fishes. For example, one historical source remarks: “…They have certain lagoons that have large fisheries, and they take out fish and store them for later” (Fernández de Oviedo y Valdés 1944:195, own translation). However, no such clear references seem to exist in relation to the storage of maize or other crops. In some cases, historical references indicate that the local people only ate “meat and fish” (Schmidl [ca. 1531] 1948:71), whereas in other references it is mentioned that in addition to these resources, the local population also engaged in small-scale consumption and/or cultivation of maize, squash, sweet potatoes, and beans (García de Moguer [1528] 1908:245; Ramírez [1528] in Madero 1939:384). In this sense, they could be seen as complex foragers with low-level food production (cf. Keeley 1988; Kelly 1995; Smith 2001; Zeder 2012) as has been suggested on several occasions in regional archaeological studies from throughout the twentieth century (see a summary in Loponte 2008).
An economic base focused on animal resources also clearly has a correlation with other aspects of the bioarchaeological record. For example, it is known that consumption of cultivated plants, especially maize, has a multiplier effect on the level of dental caries seen in individuals (Larsen 1984; Larson 1995, 1997; Rose et al. 1984; Turner 1979). In the mortuary context of the Los Marinos site, where three dates confirm an antiquity close to the time of European contact, the individuals studied have very low levels of dental caries, typical of non-agricultural populations (Kozameh and Barbosa 1996; Mazza and Barrientos 2012). The same applies to the Escuela 31 site, where none of the three individuals recovered have even a single instance of dental caries. This is especially remarkable considering that these individuals also show the lowest nitrogen levels of the samples analyzed in the present study. Studies currently in progress regarding the bioarchaeological record from other sites referred to in this article should allow for a more detailed understanding of these aspects.
Final remarks
The results presented and discussed in this article indicate that human individuals recovered from sites corresponding to the archaeological unit called Goya-Malabrigo had a monoisotopic C3 diet. In virtually all of the samples examined, maize was not isotopically detectable, indicating that this crop had a marginal to null impact on the local diets. The carbon and nitrogen levels show a preponderantly carnivorous diet, most likely based on freshwater fishes, and secondly on terrestrial species with depleted δ13Cco values, such as coypu and some ungulates. The nitrogen levels detected are even higher than those observed for complex hunter-gatherers who are associated with other archaeological units from the southernmost part of the Paraná River basin (Loponte 2008). Furthermore, the isotope values recorded in the individuals studied also clearly differ from those obtained from local horticulturist groups corresponding to the Guaraní archaeological unit, which had a mixed diet that undoubtedly included maize consumption. These data lead to questioning the hypothesis that crop plants played a significant role for the groups labeled as Goya-Malabrigo, as has been proposed by some authors based on archaeobotanical record (Bonomo et al. 2011a, b). The first exploration of this issue by isotopic data, presented in this paper, suggests on the contrary that they could not be classified as societies with a mixed economy. Rather, isotopic information obtained on individuals analyzed in this study seems to indicate an economic base focused on animal resources.
This interpretation of low or possibly null importance of cultivated plants and vegetable foods, as established by the isotopic analysis, mainly through δ15N and δ13C values, is not contrary to the detection of phytoliths and starch grains as reported by archaeobotanical studies, since this evidence could be associated with only a low level of processing and/or consumption of cultigens. Temporal analysis of the samples also suggests that there was no existing trend towards the incorporation of maize or other cultigens over time. To the contrary, our analysis essentially points to an isotopically stable diet for the chronological period of 1800 ± 47 to 460 ± 50 14C year BP, with a slight trend towards increased consumption of animal protein during the later portion of this range. New samples currently under analysis should soon allow isotopic characterization of the diets of these groups to be expanded and should lead to a better understanding of their spatio-temporal variability.
Notes
An NISP of n = 7 for Hydrochaeris hydrocaheris is recorded for the Arroyo Arenal I site (Nóbile 1993); however, the percentage this represents with respect to the total sample is not specified.
Samples were collected between November 2009 and December 2010 (Marchese et al. 2014); therefore, we consider the Suess effect until 2010.
Taken an average of δ13C −25.9 ‰ (Suess correction) plus 1.35 ‰ (average 1.1–1.6) for bone collagen.
Averaging corrected values for shad (−24.5 ‰) and corrected values for other fishes (−23.7 ‰).
We considered the average reported by Somerville (2013:1544) from a total of 102 samples, which grouped together different time periods and which includes both commoners and elite individuals, ensuring greater variability in the comparative sample.
References
Acosta A (2005) Zooarqueología de cazadores-recolectores del extremo nororiental de la provincia de Buenos Aires (humedal del Paraná inferior, Región Pampeana, Argentina). Tesis doctoral, Universidad Nacional de La Plata
Acosta A, Loponte D (2001) Tendencias paleodietarias a través del análisis de isótopos estables de poblaciones prehispánicas del Norte de la provincia de Buenos Aires. XIV Congreso Nacional de Arqueología Argentina, Rosario
Acosta A, Loponte D, Mucciolo L (2010) Comparando estrategias de explotación faunística en el humedal del Paraná inferior: cazadores-recolectores vs. horticultores amazónicos. In: Gutiérrez M, De Nigris M, Fernández P, Giardina M, Gil A, Izeta A, Neme G, Yacobaccio H (eds) Zooarqueología a principios del siglo XXI. Aportes teóricos, metodológicos y casos de estudio. Ediciones del Espinillo, Buenos Aires, pp 177–188
Acosta A, Loponte D, Tchilinguirián P (2013a) Nuevos aportes para la arqueología del humedal del Paraná inferior: el sitio Médanos de Escobar. Relaciones de la Sociedad Argentina de Antropología 38:19–35
Acosta A, Ríos Román V (2013) Explotación prehispánica de palmeras por grupos cazadores-recolectores y horticultores del extremo sur de Sudamérica: el caso del humedal del Paraná inferior (Argentina). Pesquisas Antropologia 70:197–216
Acosta I, Leiva G, Malec L (2013b) Estudio de herramientas líticas del humedal del Paraná inferior. Análisis de los gránulos de almidón. Cuadernos del INAPL (series especiales) 1(2):174–184
Ambrose SH (1993) Isotopic analysis of paleodiets: methodological and interpretive considerations. In: Sandford MK (ed) Investigations of ancient human tissue. Chemical analysis in anthropology. Gordon and Breach Scientific, New York, pp 59–130
Ambrose S, DeNiro M (1986) Reconstruction of African human diet using bone collagen carbon and nitrogen isotope ratios. Nature 319:321–324
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert JB, Grupe J (eds) Prehistoric human bone. Archaeology at the molecular level. Springer, Berlin, pp 1–37
Ambrose SH, Butler BM, Hanson DB, Hunter-Anderson RL, Krueger HW (1997) Stable isotopic analysis of human diet in the Marianas archipelago, western Pacific. Am J Phys Anthropol 104:343–361
Ambrose HS, Buikstra J, Krueger HW (2003) Status and gender differences in diet at Mound 72, Cahokia, revealed by isotopic analysis of bone. J Anthropol Archaeol 22:217–226
Ambrose SH, Krigbaum J (2003) Bone chemistry and bioarchaeology. J Anthropol Archaeol 22:193–199
Arrizurieta MP, Mucciolo L, Musali J (2010) Análisis faunístico preliminar del sitio Cerro Lutz. In: Luna L, Bonomo M, Montalvo C, Aranda C, Carrera Aizpitarte M (eds) Mamül Mapu: pasado y presente desde la arqueología pampeana, volumen I. Editorial Libros del Espinillo, Ayacucho, pp 261–276
Balasse M, Bocherens H, Mariotti A (1999) Intra-bone variability of collagen and apatite isotopic composition used as evidence of a change of diet. J Archaeol Sci 26:593–598
Barboza MC, Píccoli C (2013) Ocupaciones humanas en la llanura aluvial del Paraná medio durante el Holoceno tardío. El registro arqueológico del sitio Los Bananos (Goya, Corrientes, Argentina). Anuario de Arqueología 5:117–132
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bonomo M, Aceituno Bocanegra FJ, Politis G, Pochettino ML (2011a) Pre-Hispanic horticulture in the Paraná Delta (Argentina): archaeological and historical evidence. World Archaeol 43(4):557–579
Bonomo M, Colobig MM, Passeggi E, Zucol A, Brea M (2011b) Multidisciplinary studies at Cerro Tapera Vázquez site, Pre-Delta National Park, Argentina: the archaeological, sedimentological and paleobotanical evidence. Quat Int 245:48–61
Bonomo M, Politis G, Gianotti C (2011c) Montículos, jerarquía social y horticultura en las sociedades indígenas del delta del río Paraná (Argentina). Lat Am Antiq 22(3):297–333
Boyd M, Varney T, Surette C, Surette J (2008) Reassessing the northern limit of maize consumption in North America: stable isotope, plant microfossil, and trace element content of carbonized food residue. J Archaeol Sci 35:2545–2556
Buc N (2010) Tecnología ósea de cazadores-recolectores del humedal del Paraná inferior (bajíos ribereños meridionales). In: Loponte DM, Acosta A (eds) Arqueología de la Cuenca del Plata serie monográfica. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires
Burkart R, Barbaro N, Sanchez R, Gómez D (1999) Eco-regiones de la Argentina. Secretaría de Recursos Naturales y Desarrollo Sustentable, Buenos Aires
Cabrera AL (1976) Regiones fitogeográficas argentinas. ACME, Buenos Aires
Calo MC, Cortes L (2009) A contribution to the study of diet of formative societies in northwestern Argentina: isotopic and archaeological evidence. Int J Osteoarchaeol 19:192–203
Ceruti C (2003) Entidades culturales presentes en la cuenca del Paraná Medio (margen entrerriana). Mundo de Antes 3:111–135
Ceruti C, González MI (2007) Modos de vida vinculados con ambientes acuáticos del Nordeste y Pampa bonaerense de Argentina. Relaciones de la Sociedad Argentina de Antropología 32:101–140
Chamberlain CP, Waldbauer JR, Fox-Dobbs K et al (2005) Pleistocene to recent dietary shifts in California condors. Proc Natl Acad Sci 12(46):16707–16711
Cione A, Tonni E (1978) Paleoethnozoological context of site Las Lechiguanas islands, Paraná Delta, Argentina. El Dorado. Newsl Bull South Am Anthropol 3(1):76–86
Clementz M, Fox-Dobbs K, Wheatley P, Koch P, Doak D (2009) Revisiting old bones: coupled carbon isotope analysis of bioapatite and collagen as an ecological and palaeoecological tool. Geol J 44:605–620
Colobig MM, Ottalagano FV (2015) Estudio arqueobotánico de los residuos orgánicos adheridos en alfarerías prehispánicas de la cuenca del Paraná medio. Revista Arqueología, in press
Colonese A, Collins M, Lucquin A et al (2014) Long-term resilience of Late Holocene coastal subsistence system in southeastern South America. PLoS One. doi:10.1371/journal.pone.0093854
Coltrain J, Leavitt S (2002) Climate and diet in Fremont prehistory: economic variability and abandonment of maize agriculture in the Great Salt Lake basin. Am Antiq 67:453–485
Cormie AB, Schwarcz HP (1996) Effects of climate on deer bone δ15N and δ13C: lack of precipitation effects on δ15N for animals consuming low amounts of C4 plants. Geochim Cosmochim Acta 60:4161–4166
Cornero S, Rangone L (2015) Análisis arqueobotánicos en sitios de la entidad arqueológica Goya-Malabrigo ubicados en el centro-norte de santa fe. Anuario de Arqueología 7:85–94
Corriale MJ, Loponte D (2015) Use of stable carbon isotope ratio for foraging behavior analysis of capybara (Hydrochoerus hydrochaeris) from Esteros del Iberá, Argentina. Mammalian Biology, in press
DeNiro MJ, Epstein S (1978a) Influence of diet on the distribution of carbon isotopes in animals. Geochimica et Cosmochimca Acta 42:495–506
DeNiro MJ, Epstein S (1978b) Carbon isotopic evidence for different feeding patterns in two hyrax species occupying the same habitat. Science 210:906–908
DeNiro MJ, Schoeninger MJ (1983) Stable carbon and nitrogen isotope ratios of bone collagen: variations within individuals, between sexes, and within populations raised on monotonous diets. J Archaeol Sci 10:199–203
Falabella F, Planella MT, Aspillaga E, Sanhueza L, Tykot RH (2007) Dieta en sociedades alfareras de chile central: aporte de análisis de isótopos estables Diet in pottery-making societies from central chile: the contribution of stable isotope analyses. Chungará 39:5–27
Fernández de Oviedo y Valdés G (1944) Historia General y Natural de las indias. Editorial Guarania, Asuncion del Paraguay
Francey RJ, Allison CE, Etheridge DM et al (1999) A 1000 year record of d13C in atmospheric CO2. Tellus 51:170–193
Froehle AW, Kellner CM, Schoeninger MJ (2010) FOCUS: effect of diet and protein source on carbon stable isotope ratios in collagen: follow up to Warinner and Tuross (2009). J Archeol Sci 37:2662–2670
Froehle AW, Kellner CM, Schoeninger MJ (2012) Multivariate carbon and nitrogen stable isotope model for the reconstruction of prehistoric human diet. Am J Phys Anthropol 147:352–369
Gan MA, Kousky VE, Ropelewski C (2004) The South American monsoon circulation and its relationship to rainfall over West-Central Brazil. J Clim 17:47–66
García de Moguer D (1908) Relación y derrotero de Diego García. In: Medina J (ed) Los viajes de Diego García de Moguer al Río de la Plata. Imprenta Elzeviriana, California, pp 232–246
Gheggi MS, Williams VI (2013) New data on food consumption in Pre-Hispanic populations from northwest Argentina (ca. 1000–1550 A.D.): the contribution of carbon and nitrogen isotopic composition of human bones. J Anthropol. doi:10.1155/2013/258190
Gil AF, Neme G, Tykot R (2010) Isótopos estables y consumo de maíz en el centro occidente argentino: tendencias temporales y espaciales. Chungará 42(2):497–513
Harrison RG, Katzenberg MA (2003) Paleodiet studies using stable carbon isotopes from bone apatite and collagen: examples from Southern Ontario and San Nicolas Island, California. J Archaeol Sci 22:227–244
Hart P, Anderson LM, Feranec RS (2011) Additional evidence for cal. seventh-century A. D. maize consumption at the kipp island site, New York. In: Rieth C, Hart J (eds) Current research in New York archaeology: A.D. 700–1300. University of New York, New York, pp 27–40
Hedges R, Reynard L (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251
Indermühle A, Stocker TF, Joos F et al (1999) Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor Dome, Antarctica. Nature 398:121–126
Kalesnik F, Vallés L, Quintana R, Aceñolaza P (2008) Parches relictuales de selva en galería (Monte Blanco) en la región del bajo Delta del río Paraná. Serie Miscelánea INSUGEO-CONICET 17:169–193
Katzenberg M (2008) Stable isotope analysis: a tool for studying past diet, demography, and life history. In: Katzenberg M, Saunders S (eds) Biological anthropology of the human skeleton. Wiley-Liss, New York, pp 413–442
Keeley LH (1988) Hunter-gatherer economic complexity and ‘population pressure’: a cross cultural analysis. J Anthropol Archaeol 7:373–411
Kellner C, Schoeninger M (2007) A simple carbon isotope model for reconstructing prehistoric human diet. Am. J Phys Anthropol 133:1112–1127
Kelly RL (1995) The foraging spectrum. Diversity of hunter-gatherer lifeways. Smithsonian Institution Press, Washington
Killian V, Galván A, Olivera D, Gallegos E (2012) Una aproximación isotópica al consumo de maíz en la localidad arqueológica río Doncellas (Dpto. de Cochinoca, prov. de Jujuy). In: Babot MP, Marschoff M, Pazzarelli F (eds) Las manos en la masa. Arqueologías, antropologías e historias de la alimentación en Suramérica. Museo de Antropología, Córdoba, pp 319–338
Koch PL (2007) Isotopic study of the biology of modern and fossil vertebrates. In: Michener R, Lajtha K (eds) Stable isotopes in ecology and environmental science. Blackwell Publishing, Boston, pp 99–154
Kozameh L, Barbosa JE (1996) Influencia de la dieta sobre el desgaste y patologías dentarias en una población agrícola temprana del Noreste Argentino. Revista Española de Antropología Biológica 17:5–26
Kozameh L, Brunás O (2013) Enfermedad de paget en un individuo prehispánico del Delta del Paraná, confirmado por examen histológico y datación radiocarbónica. Cuadernos del INAPL (series especiales) 1(1):114–120
Krueger HW, Sullivan CH (1984) Models for carbon isotope fractionation between diet and bone. In Stable Isotopes in Nutrition. Am Chem Soc Symp Ser 258:205–222
Laguens AG, Fabra M, Santos GM, Demarchi A (2009) Palaeodietary inferences based on isotopic data for pre-Hispanic populations of the central mountains of Argentina. Int J Osteoarchaeol 19:237–249
Larsen CS (1984) Health and disease in prehistoric Georgia: the transition to agriculture. In: Cohen MN, Armelagos GJ (eds) Paleopathology at the origins of agriculture. Academic, Orlando, pp 367–392
Larson CS (1995) Biological changes in human populations with agriculture. Annu Rev Anthropol 24:185–213
Larson CS (1997) Bioarchaeology: interpreting behavior from the human skeleton. Cambridge University Press, Cambrige
Lee-Thorp J, Van der Merwe NJ (1987) Carbon isotope analysis of fossil bone apatite. S Afr J Sci 83:712–715
Lee-Thorp JA, Sealy JC, Van der Merwe NJ (1989) Stable isotope carbon ratio differences between bone collagen and bone apatite, and their relationship to diet. J Archaeol Sci 16:585–599
Loftus E, Sealy J (2012) Technical note: interpreting stable carbon isotopes in human tooth enamel: an examination of tissue spacing from South Africa. Am J Phys Anthropol 147:499–507
Lopes CA, Benedito-Cecilio E, Martinelli L (2007) Variability in the carbon isotope signature of Prochilodus lineatus (Prochilodontidae, Characiformes) a bottom-feeding fish of the Neotropical region. J Fish Biol 70:1649–1659
Lopes CA, Benedito E, Martinelli L (2009) Trophic position of bottom-feeding fish in the Upper Paraná River floodplain. Braz J Biol 69(2):573–581
Loponte DM (2008) Arqueología del humedal del Paraná inferior. Bajíos Ribereños meridionales. In: Loponte DM, Acosta A (eds) Arqueología de la Cuenca del Plata serie monográfica. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires
Loponte D, Acosta A (2004) Late Holocene hunter-gatherers from the Pampean wetlands, Argentina. In: Mengoni Goñalons G (ed) Zooarchaeology of South America. BAR International Series, Oxford, pp 39–57
Loponte D, Acosta A (2006) Integrating zooarchaeology from Pampa and Patagonia. In: Maltby M (ed) Integrating zooarchaeology. Oxbow Books Series, Londres, pp 5–16
Loponte DM, Acosta A (2007) Horticultores amazónicos en humedal del Paraná inferior: los primeros datos isotópicos de la dieta. Arqueología de la Pampas. In: Bayón C, Pupio A, González MI, Flegenheimer N, Frére M (eds) Arqueología en las Pampas. Sociedad Argentina de Antropología, Buenos Aires, pp 75–93
Loponte DM, Kozameh L (2009) Nuevos datos para el conocimiento de las dietas prehispánicas del Delta superior. Comechingonia 12:115–118
Loponte DM, Acosta A, Mucciolo L (2012) Contribución a la arqueología del Delta del Paraná: El nivel acerámico del sitio Isla Lechiguanas 1. Comechingonia 16:229–268
Loponte D, Corriale MJ (2012) Isotopic values of diet of Blastocerus dichotomus (marsh deer) in Paraná Basin, South America. J Archaeol Sci 40:1382–1388
Loponte DM, Acosta A (2013) La construcción de la unidad arqueológica Guaraní en el extremo meridional de su distribución geográfica. Cuadernos del Instituto Nacional de Antropología y Pensamiento latinoamericano (Series Especiales) 1(4):193–235
Loponte DM, Acosta A, Corriale MJ (2015) Complex hunter-gatherers in the wetlands of the lower Paraná River. Trends in isotopic diet. Submitted for publication
Madanes N, Kalesnik F, Vargas D (2013) Análisis de los recursos vegetales comestibles del macromosaico de humedales del Paraná inferior del Holoceno tardío. Cuadernos del INAPL (series especiales) 1(1):196–2010
Madero E (1939) Historia del puerto de Buenos Aires. Ediciones Buenos Aires, Buenos Aires
Marchese MR, Saigo M, Zillia F, Capello S, Devercellia M, Montalto L, Paporello G, Wantzen KM (2014) Food webs of the Paraná River floodplain: assessing basal sourcesusing stable carbon and nitrogen isotopes. Limnologica 46:22–30
Mazza B, Barrientos G (2012) Patologías orales de origen infeccioso en poblaciones cazadoras-recolectoras del Holoceno tardío procedentes del Humedal del Paraná inferior, Argentina. XII Congreso de la Asociación Latinoamericana de Antropología Biológica, San José
Metcalfe JZ, White CD, Longstaffe FJ, Wrobel G, Cook DC, Pyburn KA (2009) Isotopic evidence for diet at Chau Hixx, Belize: testing regional models of hierarchy and heterarchy. Lat Am Antiq 20:15–36
Metges C, Kempe K, Schmidt H (1990) Dependence of the carbon-isotope contents of breath carbon-dioxide, milk, serum and rumen fermentation products on the delta-C-13 value of food in dairy-cows. Br J Nutr 63:187–196
Musali J (2010) El rol de la pesca entre los grupos humanos de la Baja Cuenca del Plata. Ictioarqueología de conjuntos prehispánicos del Holoceno tardío en el humedal del río Paraná inferior. Tesis doctoral, Universidad de Buenos Aires
Naranjo G, Malec L, Pérez M (2010) Análisis de ácidos grasos en alfarería arqueológica del humedal del Paraná inferior. Avances en el conocimiento de su uso. In: Bárcena R, Chiavazza H (eds) Arqueología Argentina en el bicentenario de la Revolución de Mayo, vol IV. Universidad Nacional de Cuyo, Mendoza, pp 1493–1498
Nóbile J (1993) Análisis arqueofaunísticos del sitio Arroyo Arenal I, Paraná medio (provincia de Entre Ríos). Tesina de grado, Universidad Nacional de Rosario
Norr L (1995) Interpreting dietary maize from bone stable isotopes in the American tropics: the state of the art. In: Stahl PW (ed) Archaeology in the Lowland American Tropics: current analytical methods and recent applications. Cambridge University Press, Cambridge, pp 198–223
Ottalagano F (2013) Aves simbólicas, estilo e identidad en la arqueología del gran río sudamericano: Un estudio contextual del arte cerámico de las sociedades prehispánicas de la cuenca del río Paraná. In: Loponte DM, Acosta A (eds) Arqueología de la Cuenca del Plata serie monográfica. Instituto Nacional de Antropología y Pensamiento Latinoamericano, Buenos Aires
Ottalagano F (2014) Explorando la variabilidad petrográfica entre vasijas decoradas y lisas del sitio La Palmera 2 (Entre Ríos, Argentina): hacia un abordaje microscópico del arte cerámico prehispánico de la cuenca del río Paraná. Revista Chilena de Antropología, in press
Ottalagano F, Darigo M, Pereyra B, Brancatelli C, Iannelli L (2015) Investigaciones arqueológicas en el sitio La Palmera 2 (cuenca del Paraná medio, provincia de Entre Ríos, nordeste de Argentina). Revista de Antropología del Museo de Entre Ríos 1(1):55–65
Pate D (1995) Stable carbon isotope assessment of hunter-gatherer mobility in prehistoric South Australia. J Archaeol Sci 22:81–87
Pérez M, Acosta I, Naranjo G, Malec L (2013) Uso de la alfarería y conductas alimenticias en el humedal del Paraná inferior a través del análisis de ácidos grasos. Cuadernos del INAPL (series especiales) 1(1):26–45
Pérez Jimeno L (2007) Investigaciones arqueológicas en el sector septentrional de la llanura aluvial del Paraná, margen santafecina. Tesis doctoral, Universidad Nacional de La Plata
Rand AJ, Healy PF, Awe JJ (2013) Stable isotopic evidence of ancient Maya diet at Caledonia, Cayo District, Belize. Int J Osteoarchaeol. doi:10.1002/oa.2308
Rodríguez J (2001) Nordeste prehispánico. In: Berberián E, Nielsen A (eds) Historia Argentina prehispánica, volumen II. Editorial Brujas, Córdoba, pp 693–736
Roesler I, Agostini MG (2012) Inventario de los vertebrados de la reserva natural Punta Lara, provincia de Buenos Aires, Argentina. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires
Roksandic Z, Minagawa M, Akazawa T (1988) Comparative analysis of dietary between Jamon and Ainu hunter-gatherers from stable carbon isotopes of human bone. Journal of the Anthropological Society of Nippon 96:125–136
Rose JC, Burnett BA, Nassaney MS, Blaeuer MW (1984) paleopathology at the origins of maize agriculture in the lower Mississippi Valley and Caddoan culture areas. In: Cohen MN (ed) Armelagos GJ Paleopathology at the origins of agriculture. Academic, Orlando, pp 139–425
Saigo M, Zilli FL, Marchese MR, Demonte D (2015) Trophic level, food chain length and omnivory in the Paraná’ River: a food web model approach in a floodplain river system. Ecol Res. doi:10.1007/s11284-015-1283-1
Salemme MC (1987) Paleoetnozología del sector bonaerense de la región pampenana. Tesis doctoral, Universidad Nacional de La Plata
Salemme MC, Tonni E (1983) Paleoetnozoología de un sitio arqueológico de la Pampa ondulada. Sitio Río Luján (Partido de Campana, provincia de Buenos Aires). Relaciones de la Sociedad Argentina de Antropología 15:77–90
Salemme M, Tonni E, Ceruti C, Iriondo M, Cione A (1987) Los vertebrados del sitio arqueológico La Palmera V, Departamento Paraná, provincia de Entre Ríos. III Jornadas de Ciencias Naturales del Litoral, Corrientes
Sánchez J, Colobig MM, Zucol A, Politis G, Bonomo M, Castiñeira C (2013) Primeros resultados sobre el uso prehispánico de los vegetales en el sitio arqueológico Los Tres Cerros 1 (Victoria, Entre Ríos, Argentina): análisis del registro biosilíceo. Darwiniana (nueva serie) 1(2):201–219
Santiago F (2004) Los roedores en el menú de los habitantes de Cerro Aguará (provincia de Santa Fe): Su análisis arqueofaunístico. Intersecciones en Antropología 5:3–18
Sartori J, Colasurdo MB (2012) La recurrencia del coipo (myocastor coypus) en los registros arqueofaunísticos de la cuenca inferior del río Paraná (Argentina). Arqueología Iberoamericana 13:23–36
Scherer AK, Wright LE, Yoder CJ (2007) Bioarchaeological evidence for social and temporal differences in diet at Piedras Negras, Guatemala. Lat Am Antiq 18:85–104
Schmidl U (1948) Crónicas del viaje a las regiones del Plata, Paraguay y Brasil. Peuser, Buenos Aires
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639
Serrano A (1946) Arqueología del Arroyo Las Mulas en el noroeste de Entre Ríos. Universidad de Córdoba, Córdoba
Serrano A (1972) Líneas fundamentales de la arqueología del Litoral (una tentativa de periodización). Publicación del Instituto de Antropología de la Universidad de Córdoba 32:1–79
Sholto-Douglas AD, Field JG, James AG, Van der Merwe NJ (1991) 13C/12C and 15N/14N isotope ratios in the Southern Benguela Ecosystem: indicators of food web relationships among different size-classes of plankton and pelagic fish; differences between fish muscle and bone collagen tissues. Mar Ecol Prog Ser 78:23–31
Slovak NM, Paytan A (2009) Fisherfolk and farmers: carbon and nitrogen isotope evidence from middle horizon Ancón, Perú. Int J Osteoarchaeol. doi:10.1002/oa.1128
Smalley J, Blake M (2003) Stalk Sugar and the domestication of maize. Curr Anthropol 44(5):675–703
Smith B (2001) Low-level food production. J Archaeol Res 9(1):1–43
Sommerville A, Fauvelle M, Froehle A (2013) Applying new approaches to modelling diet and status: isotopic evidence for commoner resiliency and elite variability in the Classic Maya lowlands. J Archaeol Sci 40:1538–1553
Staller J, Tykot R, Benz B (2006) Histories of maize. Multidisciplinary approaches to the prehistory, linguistics, biogeography, domestication, and evolution of maize. Academic Press. Elsevier, New York
Stevens R, Jacobi R, Street M, Germonpré M, Conard N, Münzel S, Hedges R (2008) Nitrogen isotope analyses of reindeer (Rangifer tarandus), 45,000BP to 9,000BP: Paleoenvironmental reconstructions. Paleogeogr Paleoclimatology Paleoecology 262(1–2):32–45
Stronge S (2012) Examining Chronological Trends in Ancient Maya diet at Minanha, Belize, Using the Stable Isotopes of Carbon and Nitrogen. Anthropology M.A. Thesis, Trent University, Peterborough
Sullivan CH, Krueger HW (1981) Carbon isotope analysis in separate chemical phases in modern and fossil bone. Nature 292:333–335
Tonni E, Ceruti C, Iriondo M (1985) Los vertebrados del sitio Arroyo Arenal 1, Dpto. La Paz, provincia de Entre Ríos (Argentina). Revista de la Asociación de Ciencias Naturales del Litoral 16(2):157–167
Turner CG (1979) Dental anthropological indicators of agriculture among the Jomon people of central Japan. X. Peopling of the Pacific. Am J Phys Anthropol 51:619–636
Tykot RH (2002) Contribution of stable isotope analysis to understanding dietary variation among the Maya. In: Jakes KA (ed) Archaeological chemistry: materials, methods and meaning. American Chemical Society, Washington, pp 214–230
Tykot RH (2004) Stable isotopes and diet: You are what you eat. In: Martini M, Milazzo M, Piacentini M (eds) Proceedings of the International School of Physics Enrico Ferni Course CLIV. IOS Press, Amsterdam, pp 433–444
Tykot RH (2006) Isotope analyses and the histories of maize. In: Staller JE, Tykot RH, Benz BF (eds) Histories of maize: multidisciplinary approaches to the prehistory, linguistics, biogeography, domestication, and evolution of maize. Academic Press/Elsevier, New York, pp 131–142
Van der Merwe N, Vogel JC (1978) 13C content of human collagen as a measure of prehistoric diet in Woodland North America. Nature 276:815–816
Van Klinken GJ, Van der Plicht J, Hedges RE (1994) Bone 13C/12C ratios reflect (palaeo) climatic variations. Geophys Res Lett 21:445–448
Vogel JC, Van der Merwe NJ (1977) Isotopic evidence for early maize cultivation in New York State. Am Antiq 42:238–242
Vuille M, Bradley RS, Werner M, Healy R, Keimig F (2003) Modeling δ 18O in precipitation over the tropical Americas: 1. Interannual variability and climatic controls. J Geophys Res 108(D6):1–24
Warinner C, Tuross N (2009) Alkaline cooking and stable isotope tissue-diet spacing in swine: archeological implications. J Archeol Sci 36:1690–1697
Waterman AJ, Tykot RH, Silva AM (2015) Stable isotope analysis of diet-based social differentiation at late prehistoric collective burials in south-western Portugal. Archaeometry. doi:10.1111/arcm.12159
White CD, Longstaffe FJ, Schwarcz HP (2006) Social directions in the isotopic anthropology of maize in the Maya region. In: Staller J, Tykot R, Benz B (eds) Histories of maize: multidisciplinary approaches to the prehistory, linguistics, biogeography, domestication, and evolution of maize. Academic, Burlington, pp 143–159
Zeder M (2012) The broad spectrum revolution at 40: resource diversity, intensification, and an alternative to optimal foraging explanations. J Anthropol Archaeol 31:241–264
Zhou J, Lau KM (1998) Does a monsoon climate exist over South America? Climate 11:1020–1040
Zucol A, Loponte D (2008) Análisis comparativo metodológico y estudio de la abundancia fitolítica en tártaro de dientes humanos de sitios arqueológicos de la provincia de Buenos Aires, Argentina. In: Korstanje MA, Babot MP (eds) Matices Interdisciplinarios en estudios fitolíticos y de otros microfósiles/ Interdisciplinary nuances in phytolith and other microfossil studies. BAR Internacional Series, Oxford, pp 39–45
Acknowledgments
This research was supported by Argentina’s National Agency for Scientific and Technological Promotion (AGENCIA) (PICT 2011–344) and National Council for Scientific and Technical Research (CONICET) (PIPs N° 11220110100565). We are indebted to Carlos Ceruti, since he recovered a large part of the burials discussed here, as well as his suggestions and comments to develop the research in the middle basin of Parana River. We must also thank the reviewers who have improved the manuscript. All opinions of this paper are responsibility of the authors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ottalagano, F., Loponte, D. Stable isotopes and diet in complex hunter-gatherers of Paraná River Basin, South America. Archaeol Anthropol Sci 9, 865–877 (2017). https://doi.org/10.1007/s12520-015-0308-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-015-0308-6