
Abstract
This study investigates the diet of an eleventh century CE parish community located in northwestern Germany. We assessed the isotopic compositions of human (n = 24) and faunal (n = 17) bone collagen (δ 13Ccol, δ 15Ncol) and human structural carbonate (δ 13Csc) using skeletal material recovered from the Dalheim cemetery. Traditional interpretation of the isotopic data indicates that Dalheim residents likely relied on a C3 plant-based diet and consumed some terrestrial animal products without evidence of marine resource input in the diet. Bivariate and multivariate models used as an additional means to assess diet indicate minor consumption of C4 plant foods in this community. The multivariate-isotope model identified regional similarities and differences in C4 plant/marine food consumption and in dietary protein sources by comparing data from Dalheim with those of other medieval sites from the published literature. We did not observe sex differences in this population but differences in δ 15Ncol suggest that juveniles consumed the lowest trophic level protein.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The goal of this research is to provide preliminary insight into the diet of a medieval community in rural Germany through a multi-isotope investigation of human bone. The samples analyzed here are from a small cemetery predating the nearby Dalheim Monastery located in the northwestern city of Lichtenau (Figs. 1a, b). Radiocarbon results link of the cemetery to the eleventh century (Hofmann et al. 2008; Warinner et al. 2014a), but the community may have used the burial ground for long given that the neighboring parish church stood on the same grounds from as early as the ninth century until circa 1429 when Augustinian monks purchased the land to build the monastery (Pieper 2000).

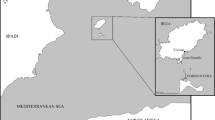
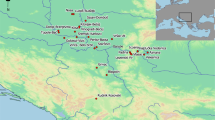
a Regional map of northwest Germany indicating the location of the Dalheim site, nearby waterways and present-day cities. b Site map with excavated areas indicated by diagonal-lined boxes. While the majority of human burials were recovered from area B (now covered by an outbuilding), some human remains were also excavated in area G. Isolated faunal bones were found in grave fill and in a location adjacent to the north side of the church (area W)
Regional variation in diet during the medieval period was impacted by many factors including location and population density, crop type and productivity, and access to traded foodstuffs (e.g., see Woolgar et al. 2006). Food economies became increasingly complex as medieval cities emerged, expanded, and established market relationships with the surrounding rural communities (Pounds 2005). Historic documents suggest that age, sex, social status, and religion also strongly influenced food access and consumption among Northern and Central European populations (Postan 1972; Bynum 1987; Carlin and Rosenthal 1998; Adamson 2004).
For the residents of medieval Dalheim, economic and social factors regulated food options, as they would have in populations across Europe. For example, historical literature suggests that the poor primarily relied on cereal crops (oats, barley, millet), vegetables (cabbage, broad beans, turnips), and to a lesser extent on dairy, eggs, and pork, while wealthy individuals had greater access to expensive foods like wheat, meats (including game), and fish (Adamson 1995, 2004). Humoral ideology was another factor affecting consumption in medieval society. In this belief system, proper health could be maintained by balancing the body’s four humors or fluids (black bile, yellow bile, phlegm, and blood). Age and sex played roles in determining individual humoral constitutions and thus influenced personal food choices (van Winter 2007; Gilchrist 2012). In addition, food was believed to play critical roles in maintaining and restoring human health, and thus, the humoral properties of foodstuffs were exploited in both medical and everyday dietary practices (Scully 1995).
The influence of the Catholic Church was also considerable as Christian populations across Europe were forbidden from eating most meats, dairy products, and eggs for more than one third of the calendar year (Henisch 1976). Fish was an accepted alternative, and consumption of marine fish (e.g., cod and herring) has been documented at urban or trade centers in medieval European sites using isotopic analyses (e.g., Barrett and Richards 2004; Müldner and Richards 2007; Salamon et al. 2008; Barrett et al. 2008, 2011; Salazar-García et al. 2014a; Alexander et al. 2015; Salazar-García et al. 2016). Even though saltwater fish is believed to have been a part of most medieval European diets by the fourteenth century, historical documents suggest that rural Central European populations far removed from the ocean were more likely to consume freshwater fish (e.g., pike and perch) than marine foods (Adamson 2004). Dalheim is one such population, situated more than 200 km inland from the nearest ocean port, and far from the country’s largest rivers (trade routes), which likely prevented easy access to saltwater foods.
Reitsema and Vercellotti (2012) have called for more research on dietary adaptations at the local level given that traditional understanding of dietary practices is generally derived from broad historical accounts of Europe’s religious and sociopolitical elite. These authors stress that isotopic research has provided insight into weaning age, sex- and status-based differences in diet, and group changes in diet over time and continues to be the key to progressively refining our understanding of local and regional variation in medieval European diet. Thus, our research is uniquely poised to address several key questions. First, how do the stable isotope data for Dalheim correspond to historical accounts of diet among Central European peasantry and to medieval European diet more generally? What (if any) age or sex-based differences in diet are visible in smaller, more rural populations like Dalheim, and do these patterns reflect ideological food proscriptions? And finally, what can this dataset tell us about the temporal and spatial variability of medieval diet within the region? Additionally, we use isotopic modeling to provide more detailed insight into the composition of ancient diets. The data for Dalheim are interrogated using a bivariate (simple) (Kellner and Schoeninger 2007; Froehle et al. 2010) and a multivariate carbon-isotope model (Froehle et al. 2012) and compared with previously published isotopic data for medieval diet. This study is the first to our knowledge that explores medieval European diet using the multivariate-isotope approach.
Isotopic background
The stable isotopic compositions of collagen (a major class of bone proteins) and structural carbonate (a minor component of bone mineral [bioapatite]) are used here in order to establish the relative importance of isotopically distinct foods in the diet at Dalheim. Stable isotope ratios are measures of our interactions with the environment and expressed in delta (δ) notation as per mil (‰) differences relative to a standard. In the food chain, the primary levels of isotopic variation are determined by climatic and environmental influences on plant species and by the photosynthetic and nitrogen-fixing pathways employed by those plants (Ambrose 1993). The tissue isotopic compositions of plant consumers reflect those of plants but with some degree of isotopic fractionation between each level of the food chain (Schoeller, 1999). Paleodiet studies measure the carbon-isotope ratios found in collagen and structural carbonate to examine the relative consumption of two isotopically distinct groups of plants, i.e., C3 and C4. Most fruits, vegetables, legumes, and grains are C3 plants, which have lower δ 13C relative to C4 plants (Smith and Epstein 1971; O’Leary 1988). The most common C4 plants available to medieval Europeans were millet and sugarcane, although sugarcane would have been a rare and expensive commodity for most medieval individuals (Adamson 2004). Stable carbon isotopic compositions are also indices of marine food consumption because the source of carbon used by plants differs in oceanic and terrestrial ecosystems, causing the tissue δ 13C of marine mammals and fish to be ~8‰ higher than those of terrestrial animals (Chisholm et al. 1982). Brackish and estuarine fish, by contrast, typically have lower δ 13C than marine fish (Salazar-García et al., 2014b). Krueger and Sullivan (1984) have shown that dietary protein is the primary influence on the carbon-isotope composition of collagen (δ 13Ccol) and thus does not provide clear insights into the isotopic composition of the remaining macronutrients (i.e., lipids and carbohydrates). In contrast, the carbon-isotope composition of structural carbonate (δ 13Csc) reflects that of whole diet (i.e., protein, lipids, and carbohydrates) but cannot identify primary sources of dietary protein (Ambrose and Norr 1993; Tieszen and Fagre 1993). Although the exact degree of 13C enrichment from diet to human bone collagen is debatable, commonly accepted values range from ~1 to 5‰ (DeNiro and Epstein 1978; van der Merwe and Vogel 1978) and predator-to-prey differences in carbon-isotope ratios are ~0 to 2‰ (Bocherens and Drucker 2003).
Bone collagen nitrogen-isotope ratios reflect protein source and trophic level. Isotopic variation begins at the base of the food chain where plant δ 15N normally ranges from about −3 to +6‰ on average in pre-industrial ecosystems (Virginia and Delwiche 1982; Heaton 1987; Ambrose 1993). In general, δ 15Ncol is ~3 to 6‰ higher than diet, and the δ 15Ncol of carnivores tends to be higher than those of omnivores which, in turn, are higher than those of herbivores in terrestrial populations (DeNiro and Epstein 1981; Schoeninger 1985; O’Connell et al. 2012). Higher δ 15Ncol among humans consuming primarily vegetarian diets could reflect the consumption of domesticated crops grown using animal manure, which could increase plant δ 15N by ~3 to 5‰ (Bogaard et al. 2007). Breastfeeding infants tend to sit at a higher trophic level relative to the parent population (Fogel et al. 1989; Fuller et al., 2006) though the reasons for this pattern are still debated (Beaumont et al. 2015). In the larger ecosystem, aquatic animals commonly have the highest δ 15Ncol because aquatic plants typically have higher δ 15N relative to terrestrial plants and because there are more trophic levels in marine and freshwater systems (Schoeninger et al. 1983; Schoeninger and DeNiro 1984). Some freshwater ecosystems, however, exhibit a high degree of isotopic variation (e.g., Häberle et al. 2016). Diseases that affect bone can influence the nitrogen-isotope composition of collagen (Katzenberg and Lovell 1999; Strange 2006) but paleodiet interpretations of nitrogen-isotope data are not confounded when bone lesions are avoided during sampling (Olsen et al. 2014).
Materials and methods
Sample
In 1989–1990, the Westphalian Museum of Archaeology excavated the Dalheim cemetery as part of a salvage operation intended to create space for new building construction. The human sample subset analyzed here (n = 24, Table 1) was obtained from the Dalheim skeletal collection curated at the Institute of Evolutionary Medicine at the University of Zurich, Switzerland. Burials at the cemetery did not include coffins or burial objects; all skeletons faced east, and many were deformed or damaged as a result of shallow burial (Hofmann et al. 2008). Burials were excavated primarily in area B of the site but were also recovered from area G (see Fig. 1b). Only well-preserved skeletons were selected for analysis and include juveniles and both male and female adults. Hofmann et al. (2008) determined the age and sex of each burial (Table 1). Harris lines identified on four adults suggest that nutritional or infectious stressors affected these individuals during their lifetimes. We selected the cortical bone from the ribs and vertebrae for isotopic analysis.
Contemporary, fragmented faunal remains of terrestrial herbivores and one Felis species carnivore (n = 17, Table 2) were also recovered during the cemetery’s excavation. Faunal remains were found in grave fill within areas B and G and from a location adjacent to the north side of the church (area W; see Fig. 1b). All of the faunal bones were found as isolated elements; none were deposited intentionally. The cortical bone, primarily from the long bones, was selected from all of the faunal samples recovered during excavation for isotopic analysis.
Analytical procedures
Human bone collagen
The human samples were analyzed at the Laboratory for Stable Isotope Science (LSIS) at The University of Western Ontario (London, Canada), following a modified Longin method to extract the cortical bone collagen (Longin 1971; DeNiro and Epstein 1978; Chisholm et al. 1983), which was then analyzed for its stable carbon- and nitrogen-isotope compositions. Bone samples (~1.0 g) were ground and placed into 0.5 M HCl solution until the mineral component of the bone dissolved. In addition, a 0.1 M NaOH solution was used to remove humic contaminants (Schoeninger and DeNiro 1984). Collagen extracts were solubilized in 0.001 M HCl in a 90 °C oven for 16 h. The solubilized collagen was transferred to 4-mL glass vials and dried completely over an additional 24–36 h in the 90 °C oven.
All samples were analyzed using a Costech elemental combustion system coupled with a Thermo Scientific Delta Plus XL stable-isotope-ratio mass spectrometer (IRMS). Sample δ 13Ccol was calibrated to VPDB using NBS-22 (−30.03‰) and IAEA-CH-6 (+10.45‰). We used international standards USGS-40 and USGS-41 to verify the calibration curve. The former had a δ 13C of −26.42 ± 0.07‰ (mean ± SD; n = 12), as compared to its accepted value of −26.39‰. The latter had a δ 13C of +37.40 ± 0.21‰ (n = 11), as compared to its accepted value of +37.63‰. The δ 15Ncol was calibrated to AIR using IAEA-N1 (+0.4‰) and IAEA-N2 (+20.3‰) or USGS-40 (−4.5‰) and IAEA-N2 (+20.3‰). The calibration curve was verified using USGS-41 and had a δ 15N of +47.42 ± 0.33‰ (n = 11), as compared to its accepted value of +47.6‰. A keratin standard (#902111, MP Biomedicals) was analyzed every four to five samples throughout all analytical sessions. The mean δ 13C and δ 15N of the keratin standard (n = 28) were −24.08 ± 0.05‰ and 6.23 ± 0.18‰, respectively, for the duration of laboratory analyses, which compare well to the accepted values of −24.04 and 6.36‰. The average reproducibility of duplicate sample analyses for the bone collagen (n = 10) was ±0.04‰ for δ 13Ccol and ±0.11‰ for δ 15Ncol.
Human bone structural carbonate
Structural carbonate was isolated by first grinding and sieving cortical bone samples (~0.2 g) to a grain size smaller than 180 μm. Reagent-grade (2%) bleach was used to remove the organic components of the bone. After 72 h, the samples were rinsed with distilled water, followed by a 4-hour soak in 0.1 M acetic acid to eliminate contaminant carbonates. The samples were rinsed again, frozen, and lyophilized.
The isotopic integrity of structural carbonate was assessed using crystallinity index (CI) and C/P values (the ratio of carbonate to phosphate in bioapatite), obtained by Fourier transform infrared (FTIR) spectroscopy of untreated samples following the methods of Shemesh (1990), Weiner and Bar-Yosef (1990), Wright and Schwarcz (1996), and Surovell and Stiner (2001). In addition, FTIR spectra were screened for the presence of secondary calcite and francolite (diagenetic minerals).
We analyzed stable carbon-isotope compositions using a Micromass Multiprep coupled with a VG Optima dual-inlet IRMS. Sample δ 13Csc was calibrated to VPDB using NBS-19 (+1.95‰) and Suprapur (−35.28‰). We used NBS-18 and an internal laboratory calcite standard (WS-1) to verify the calibration curve. The former had a δ 13C of −4.98 ± 0.02‰ (mean ± SD; n = 6), as compared to its accepted value of −5.00‰. The latter had a δ 13C of +0.82 ± 0.07‰ (n = 8), as compared to its accepted value of +0.76‰. The average reproducibility of duplicate sample analyses for structural carbonate (n = 4) was ±0.04‰ for δ 13Csc.
Faunal bone collagen
Collagen extraction and analysis of faunal remains took place at the Stable Light Isotope facilities of the University of Cape Town (Cape Town, South Africa). Collagen extraction methods followed the process described in Richards and Hedges (1999). Bone fragments of ~300 mg from each sampled bone were demineralized at 5 °C in 0.5 M HCl over the course of a week and were then rinsed with deionized water until pH became neutral. Gelatinization followed this process at 70 °C during 48 h in a pH ranging from 2 to 3. Filtering and ultrafiltering using 50–90-μm EZEE© filters, and previously cleaned 30 kDa Amicon© ultrafilters respectively, was also carried out. The purified solutions were frozen and lyophilized before being weighed into tin capsules and loaded onto the mass spectrometer.
The carbon and nitrogen-isotope ratios in collagen were measured in duplicate, using a Delta XP continuous-flow IRMS interfaced with an elemental analyzer, Flash EA 2112 (Thermo Scientific©). Stable carbon-isotope ratios were expressed relative to the VPDB scale (Vienna PeeDee Belemnite) and stable nitrogen-isotope ratios were measured relative to the AIR scale (atmospheric N2), using the delta notation (δ) in parts per thousand (‰). Repeated analysis of internal and international standards indicated an analytical error better than 0.1‰ (1σ) for δ 13C and δ 15N.
Sample integrity
Tables 1 and 2 provide the results for individual tests of sample integrity. One faunal sample (ID 19961) was excluded from interpretation due to a low collagen yield. Otherwise, excellent bone collagen preservation was indicated by collagen yields (human mean = 9.9 ± 4.7% SD, n = 24; faunal mean = 3.1 ± 1.3% SD, n = 16) that exceeded the minimum standard (1%; van Klinken 1999) and carbon/nitrogen ratios (human mean = 3.2 ± 0.1 SD, n = 24; faunal mean = 3.3 ± 0.05 SD, n = 16) that fell within the acceptable range (2.9 to 3.6; DeNiro 1985).
Excellent structural carbonate preservation was indicated by crystallinity index (CI) values (mean = 2.7 ± 0.1 SD, n = 24) similar to literature values for well-preserved untreated archeological bone (e.g., Nielsen-Marsh and Hedges 2000) and by carbonate/phosphate ratios (mean = 0.42 ± 0.05 SD, n = 24) which fell within the range established for well-preserved samples (0.3–0.5; Nielsen-Marsh and Hedges 2000). We found no discrete peaks in the FTIR spectra that would indicate the presence of secondary calcite or francolite contaminants in the samples. Preservation of post-mortem isotopic integrity is further indicated by a lack in significant correlations between CI values and carbonate yields (Pearson’s r = 0.44, df = 22), C/P values (Pearson’s r = 0.07, df = 22), or δ 13Csc (Pearson’s r = −0.05, df = 22).
Results
Tables 1 and 2 summarize the isotopic data for the Dalheim human and faunal samples. For the human dataset (n = 24), the mean isotopic compositions are −20.0 ± 0.2‰ SD (δ 13Ccol), +9.9 ± 1.0‰ SD (δ 15Ncol), and −13.2 ± 0.6‰ SD (δ 13Csc). The average collagen isotopic compositions match well with data published by Warinner et al. (2014a), who analyzed collagen from four Dalheim individuals. The average spacing between δ 13Ccol and δ 13Csc (∆ 13Csc-col) is 6.8 ± 0.6‰ SD. The isotopic compositions for the single terrestrial carnivore recovered from the site are −20.3‰ (δ 13Ccol) and +9.1‰ (δ 15Ncol), while the mean isotopic compositions for terrestrial herbivores (n = 15) are −21.4 ± 0.6‰ SD (δ 13Ccol) and +5.70 ± 1.2‰ SD (δ 15Ncol).
We identified no systematic differences in either the carbon- or nitrogen-isotope data that were associated with the sex of the sampled individuals. No differences were identified between individuals recovered in area B of the cemetery excavation and those recovered in area G (i.e., individuals G03, G07, G11). Skeletons exhibiting Harris lines did not stand out as having different isotopic compositions from the rest of the sample. One-way analysis of variance (ANOVA) tests found one significant difference between age groups for nitrogen-isotope compositions [F(3,18) = 3.83, p = 0.03]. Tukey post hoc comparisons found that δ 15Ncol for juveniles (mean = +9.1 ± 0.6‰ SD) was significantly lower than those of younger adults (mean = +11.0 ± 1.6‰ SD) at the p < 0.02 level (Fig. 2).

Human and faunal collagen carbon- and nitrogen-isotope compositions for Dalheim. a Individual values. b Average compositions ± SD
Discussion
Interpreting diet
The low δ 13Ccol (−20.0‰) at Dalheim suggests that C4 resources (plants or plant consumers) and marine foods were not substantial components of the diet. In other medieval European populations, higher δ 13Ccol has indicated either millet (Reitsema et al. 2010; Reitsema and Vercellotti 2012; Salazar-García et al., 2014a; Alexander et al., 2015) or marine food (Müldner and Richards 2007; Salazar-García et al., 2016) consumption. The Dalheim population likely depended on C3 plant foods given that the degree of 13C enrichment from diet to human bone collagen is generally thought to be less than 5‰ (DeNiro and Epstein 1978; van der Merwe and Vogel 1978), but small amounts of animal protein consumption cannot be ruled out given that predator-to-prey differences in carbon-isotope ratios range from 0 to 2‰ (Bocherens and Drucker 2003).
The mean δ 15Ncol of the sampled individuals is +9.9‰, while the mean δ 15Ncol for terrestrial herbivores is +5.7‰. The herbivore mean is consistent with other medieval German samples that have δ 15Ncol ranging from +5.2 to +7.4‰ (Strott et al. 2007; Doppler et al. 2010; von Heyking 2012; Knipper et al. 2013; Olsen 2013). If the Dalheim population had subsisted primarily on C3 plants, the average nitrogen-isotope composition would resemble that of terrestrial herbivores (e.g., cows or sheep). Because nitrogen-isotope compositions increase by ~+3 to +6‰ between trophic levels (DeNiro and Epstein 1981; Schoeninger 1985; O’Connell et al. 2012), the human nitrogen-isotope composition at Dalheim is consistent with the regular consumption of terrestrial mammal protein. The low δ 15Ncol among juveniles (children 7 to 12 years old) at Dalheim suggests that they consumed protein from lower trophic levels relative to younger adults (20 to 40 years old). The data compare well with Richards et al. (2002) who suggested that a more plant-based diet was responsible for significantly lower δ 15Ncol among young children (4 to 8 years) relative to older children and adults at a tenth–sixteenth century English site. Reitsema et al. (2016) have also identified lower relative protein consumption among some children at the medieval Italian site of Trino Vercellese (eighth to thirteenth centuries) and suggest that the lack of protein directly impacted the survival of these individuals.
Although ∆ 13Csc-col spacings can provide a secondary index of trophic level for ancient food webs independently of nitrogen-isotope compositions, they do not show any obvious differences among age groups in our sample. The individuals with the highest δ 15Ncol may have had access to higher trophic level protein like pork, domesticated fowl, or freshwater resources. The paucity of regional isotopic data for freshwater foods, however, limits our understanding of their dietary contribution. In this temporal/regional context, the higher δ 15Ncol may also reflect the consumption of domesticated crops grown using animal manure (see Bogaard et al. 2007). We also cannot differentiate meat consumption from egg and dairy consumption using carbon- and nitrogen-isotope data alone (O’Connell and Hedges 1999). Thus, the Dalheim population could have relied more on these secondary animal products if meat products were less accessible. While we consider the possibility of meat consumption in this community with caution, recent analyses of genetic material extracted from dental calculus indicate that meat protein (pig/boar, and sheep) was consumed by some Dalheim individuals (Warinner et al. 2014a). Additionally, host ancient DNA investigation of 18 individuals in the Dalheim cemetery, on one hand, revealed a high level of genetic lactase persistence, indicative of milk consumption and genetic selection for the trait at some point in the population’s past (Krüttli et al., 2014). On the other hand, protein analysis of the dental calculus of nine Dalheim individuals did not identify any milk proteins (Warinner et al. 2014b).
Isotopic modeling: simple carbon model
Taking advantage of the different isotopic signals preserved by tissue δ 13Ccol versus δ 13Csc described earlier, Kellner and Schoeninger (2007) and Froehle et al. (2010) developed the “simple carbon model” in order to estimate the ratio of C3 to C4 foods (plants and plant consumers combined) in the diet and to determine whether consumed protein was either “C3-derived” or “C4/marine-derived”. C4-derived and marine-derived proteins are paired here because they share similar carbon isotopic compositions and the model cannot differentiate between them. Kellner and Schoeninger (2007) and Froehle et al. (2010) combined isotopic data for animals from controlled diet experiments to produce parallel regression lines that characterize known diet types. They consider the model appropriate for mapping human datasets because the diet-bone relationships found in the isotopic compositions of experimental animals (rats, mice, pigs) did not differ significantly from those of humans and because archeological populations with well-identified diets fit the model.
Figure 3 illustrates the δ 13Ccol and δ 13Csc for Dalheim in relation to the regression lines of the simple carbon model. Primary sources of dietary protein are indicated by the proximity of the values to the regression lines, while proximity to the poles of the regression lines denotes the ratio of C3 to C4 foods in the diet (Froehle et al. 2010). The carbon isotopic compositions for Dalheim cluster tightly on the C3 protein line, indicating that dietary protein primarily came from C3 foods (i.e., C3 plants or C3 plant consumers). The δ 13Csc values are also low on the regression line, suggesting that the proportion of total C3 foods in the diet was much higher than that of C4 foods, thus confirming expectations. Because the δ 13C values are not at the 100% C3 endpoint, however, it is possible that a small proportion of carbon (i.e., less than ~25%) came from non-C3 sources. Meat from farm animals foddered on non-C3 sources are less likely to constitute the non-C3 portion of the diet because these protein sources would shift individual isotopic compositions away from the C3 protein line and towards the C4/marine protein line.

Isotopic modeling: multivariate-isotope model
Because the simple carbon model does not differentiate C4 food and marine resource consumption, Froehle et al. (2012) introduced the multivariate-isotope model, which resolves this issue by incorporating δ 15Ncol results and using cluster and discriminant function analyses. These analyses generated two functions that describe how the test samples varied in terms of the isotopic data:
-
Function 1
(Carbon) = (0.322 × δ 13Csc) + (0.727 × δ 13Ccol) + (0.219 × δ 15Ncol) + 9.354
-
Function 2
(Nitrogen) = (−0.393 × δ 13Csc) + (0.113 × δ 13Ccol) + (0.622 × δ 15Ncol) − 8.703
The function scores enable the plotting of data into five clusters of dietary types (Froehle et al. 2012; see also Sommerville et al. 2013). The data for Dalheim are modeled this way in Fig. 4 and compared with modeled published isotopic dietary data from Strott et al. (2007) and Grupe et al. (2009). A general lack of structural carbonate (δ 13Csc) data in the isotopic literature on medieval European diet barred additional relevant comparisons.

Discriminant function values for Dalheim and two additional medieval German populations plotted on the Froehle et al. (2012) multivariate-isotope model. a Individual values. b Group means ± SD. Asterisk indicates Upper Bavarian sites including Etting, Unterigling, Kelheim, Enkering, and Grossmehring (see Strott et al. 2007)
Some of the Dalheim individuals plot within cluster 1, reflecting a diet based entirely on C3 foods, but most do not fit within any of the currently established fields. Although the Dalheim individuals have the greatest proximity to clusters 1 and 4, the diet of Dalheim residents likely differed from those of the populations used to create the dietary clusters. Froehle et al. (2012) generated cluster 1 using data primarily from a non-farming population in Ontario (Canada) that consumed C3 plants, wild game, waterfowl, and freshwater fish, while cluster 4 was mainly derived from a similar Ontarian population that also relied on maize (C4) agriculture. Lower nitrogen (Function 2) scores at Dalheim may reflect a more vegetarian diet or a lesser dependence on freshwater foods. The placement of the Dalheim data on the x-axis (below the lower limits of the cluster 4 box) suggests that less than 30% of the diet was derived from C4 resources. This finding matches well with the simple carbon model estimate of <~25% C4 foods in the Dalheim diet. Small amounts of millet in the diet could explain this pattern. Millet was generally avoided by the wealthy, while poor medieval Central Europeans consumed millet more regularly in soups and porridges, and especially in times of famine (Adamson 2004). The four adults in the Dalheim population with tibial Harris lines could indicate an underlying nutritional stress in this sub-population.
For all sites, the placement of individuals near clusters 1 and 4 suggests a high plant dependence and low consumption of freshwater animal species. The separation of the Central European populations along the y-axis (function 2, nitrogen) of Fig. 4b, however, likely reflects dietary differences among the three medieval regions compared here. For example, elite individuals from sixth–tenth century cemeteries in southeastern Germany (Strott et al. 2007) have a function mean score similar to that of Dalheim, and data from both sites lie close to clusters 1 and 4. This consistency in diet is remarkable given that the cemeteries represent socially privileged individuals from an earlier period in the medieval timeframe and are located more than 500 km to the southeast of Dalheim. This close comparison highlights broad regional and temporal similarities in diet. By contrast, the function mean score for Schleswig, a site found ~400 km to the north of Dalheim, reflects a clear regional difference in food access. Grupe et al. (2009) suggest that inhabitants of this eleventh–thirteenth century site located along the Schlei Fjord consumed domestic mammals, wild game, and freshwater fish but few or no marine resources. Although a lack of seafood consumption is common to both Schleswig and Dalheim, the presence of foods like freshwater fish and waterfowl in the diet could explain the relatively higher function 2 (nitrogen) values for the Schleswig site.
Medieval diet
If the rural community in Dalheim followed the dietary guidelines issued by the Catholic Church, it did not rely heavily on marine fish as a replacement for terrestrial animal products. The isotopic data suggest that the population derived its protein from non-marine resources (e.g., terrestrial mammals), even though dried and salted fish like herring and cod were widespread dietary staples traded throughout medieval Europe. Although German populations located far from the ocean more commonly consumed freshwater fish (Adamson 2002), low δ 15Ncol indicate that Dalheim’s residents may not have depended much on this resource unless the fish were relatively low trophic level species. Considering the high variation in δ 15N of freshwater fish, more data are needed on European freshwater food webs to fully interpret the medieval Dalheim tissue isotopic compositions (e.g., Fuller et al. 2012; Häberle et al. 2016).
It is not clear how pervasive humoral ideology would have been among the less literate segments of Central European society, but families and communities still had to make decisions about the kinds of foods they acquired, how they chose to prepare those foods, and how they were distributed within the family and community. Thus, social understanding in some form must have influenced these decisions. At Dalheim, juveniles (7 to 12 years) have the lowest δ 15Ncol in the population, and the results are significantly different from those for younger adults (20 to 40 years). This difference suggests that juveniles consumed protein from a lower trophic level (i.e., fewer terrestrial and/or freshwater meats). From the humoral point of view, children and adults require different diets because a person’s temperament changes with age, and appropriate food options would change accordingly (van Winter 2007). Poor economic circumstances, however, could also explain a differential access to protein and could have been a contributing factor to the early deaths of these children.
Differences between the sexes might also have been expected if humoral rules governing food choices at Dalheim were the primary determinant of diet. Previous studies of coastal (Richards et al. 2006) and urban medieval centers (Kjellström et al. 2009; Reitsema et al. 2010; Reitsema and Vercellotti 2012) identified sex-based variation in diet. Various interpretations of these differences include differing status, occupation, and responsibilities within a household, but humoral belief systems may have also been a factor as women and men were considered to have contrasting humoral constitutions (Gilchrist 2012). No dietary differences were observed between males and females in our sample. Perhaps rural living in the more isolated community of Dalheim precluded food options that would allow for sex-based variation in diet. In addition, isotopic analysis would not be able to differentiate foods from similar trophic levels with different humoral characteristics (e.g., terrestrial meats, eggs, and dairy).
Conclusions
The objective of this research was to provide preliminary insight into the lives of the rural medieval community living at Dalheim through a multi-isotope investigation of the human and animal bone. A traditional approach to interpretation of the carbon isotopic data suggests that C3 plants and/or plant consumers were the main dietary resources, while the nitrogen isotopic data indicate that the population likely consumed animal products, possibly including dairy, eggs, and meat. This isotopic study suggests that part of their dietary protein derived from terrestrial and/or freshwater animal sources but not marine animal sources. We also investigated age- and sex-based isotopic variation and found differences in δ 15Ncol that suggest juveniles in the community consumed protein from a lower trophic level than adults, which reflects differential access to resources and could be consistent with socially proscribed differences in consumption. Additional research on medieval populations like Dalheim is essential to furthering a nuanced understanding of religious and cultural influences on diet.
Two isotopic modeling techniques used here as an additional means to assess diet identified possible minor consumption of C4 resources at Dalheim. In the absence of evidence for marine protein consumption, millet is the most likely C4 resource consumed. Its presence in the diet could reflect famine conditions that regularly affected medieval populations during this period. A comparison of the Dalheim isotopic data to previously reported results for other Central European medieval sites underscores the presence of temporal and spatial variability of medieval diet in the region. Even though the isotopic data primarily fall outside the defined modeling clusters, this exercise still provides a useful comparison. Further refinement of the multivariate-isotope model may be helpful in expanding its applicability to a wider range of archeological populations.
References
Adamson MW (1995) “Unus theutonicus plus bibit quam duo latini” food and drink in late medieval Germany. Medium Aevum Quotidianum 33:8–20
Adamson MW (2002) Medieval Germany. In: Adamson MW (ed) Regional cuisines of Europe: a book of essays. Routledge, New York, pp. 153–196
Adamson MW (2004) Food in medieval times. Greenwood Press, Westport
Alexander MM, Gerrard CM, Gutiérrez A, Millard AR (2015) Diet, society and economy in late medieval Spain: stable isotope evidence from Muslims and Christians from Gandía, Valencia. Amer J of Phys Anthropol 156:263–273
Ambrose SH (1993) Isotopic analysis of paleodiets: methodological and interpretive considerations. In: Sandford MK (ed) Investigations of ancient human tissue: chemical analyses in anthropology. Gordon and Breach Science Publishers, Langhorne, pp. 59–130
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert J, Grupe G (eds) Prehistoric human bone: archaeology at the molecular level. Springer-Verlag, Berlin, pp. 1–37
Barrett JH, Richards MP (2004) Identity, gender, religion and economy: new isotope and radiocarbon evidence for marine resource intensification the early historic Orkney, Scotland, UK. Eur J Archaeol 7:249–271
Barrett JH, Johnstone C, Harland J, Van Neer W, Ervynck A, Makowiecki D, Heinrich D, Hufthammer AK, Enghoff IB, Amundsen C, Christiansen JS, Jones AKG, Locker A, Hamilton-Dyer S, Jonsson L, Lõugas L, Roberts C, Richards MP (2008) Detecting the medieval cod trade: a new method and first results. J Archaeol Sci 35:850–861
Barrett JH, Orton D, Johnstone C, Harland J, Van Neer W, Ervynck A, Roberts C, Locker A, Amundsen C, Enghoff IB, Hamilton-Dyer S, Heinrich D, Hufthammer K, Jones AKG, Jonsson L, Makowiecki D, Pope P, O’Connell TC, de Roo T, Richards MP (2011) Interpreting the expansion of sea fishing in medieval Europe using stable isotope analysis of archaeological cod bones. J Archaeol Sci 38:1516–1524
Beaumont J, Montgomery J, Buckberry J, Jay M (2015) Infant mortality and isotopic complexity: new approaches to stress, maternal health, and weaning. Am J Phys Anthropol 157:441–457
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343
Bynum CW (1987) Holy feast and holy fast: the religious significance of food to medieval women. University of California Press, Berkeley
Carlin M, Rosenthal JT (eds) (1998) Food and eating in medieval Europe. Hambledon Press, London
Chisholm BS, Nelson DE, Schwarcz HP (1982) Stable-carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science 216:1131–1132
Chisholm BS, Nelson DE, Hobson KA, Schwarcz HP, Knyf M (1983) Carbon isotope measurement techniques for bone collagen: notes for the archaeologist. J Archaeol Sci 10:355–360
DeNiro MJ (1985) Post-mortem preservation and alteration of in vivo bone collagen isotope ratios in relation to paleodietary reconstruction. Nature 317:806–809
DeNiro MJ, Epstein S (1978) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Ac 42:495–506
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Ac 45:341–351
Doppler S, Vohberger M, von Carnap-Bornheim C, Heinrich D, Peters J, Grupe G (2010) Biodiversity of archaeological fauna in the estuarine palaeoecosytem of the Schlei fjord, Northern Germany: isotopic evidence. In: Grupe G, Peters J (eds) Documenta archaeobiologiae, vol 8, archaeobiodiversity—a European perspective. Verlag Marie Leidorf GmbH, Rahden/Westf, pp. 21–70
Fogel ML, Tuross N, Owsley O (1989) Nitrogen isotope tracers of human lactation in modern and archaeological populations. Carnegie I Wash 1989:111–117
Froehle AW, Kellner CM, Schoeninger MJ (2010) FOCUS: effect of diet and protein source on carbon stable isotope ratios in collagen: follow up to Warinner and Tuross (2009). J Archaeol Sci 37:2662–2670
Froehle AW, Kellner CM, Schoeninger MJ (2012) Multivariate carbon and nitrogen stable isotope model for the reconstruction of prehistoric human diet. Am J Phys Anthropol 147:352–369
Fuller BT, Fuller JL, Harris DA, Hedges REM (2006) Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am J Phys Anthropol 129:279–293
Fuller BT, Müldner G, Van Neer W, Ervyncke A, Richards MP (2012) Carbon and nitrogen stable isotope ratio analysis of freshwater, brackish and marine fish from Belgian archaeological sites (1st and 2nd millennium AD). J Anal At Spectrom 27:807–820
Gilchrist R (2012) Medieval life: archaeology and the life course. Boydell Press, Woodbridge
Grupe G, Heinrich D, Peters J (2009) A brackish water aquatic foodweb: trophic levels and salinity gradients in the Schlei fjord, Northern Germany, in Viking and medieval times. J Archaeol Sci 36:2125–2144
Häberle S, Fuller BT, Nehlich O, Van Neer W, Schibler J, Hüster Plogmann H (2016) Inter- and intraspecies variability in stable isotope ratio values of archaeological freshwater fish remains from Switzerland (11th–19th centuries AD). Enviro Archaeol: J Human Palaeoecol 21:119–132. doi:10.1179/1749631414Y.0000000042
Heaton THE (1987) The 15N/14N ratios of plants in South Africa and Namibia: relationship to climate and coastal/saline environments. Oecologia 74:236–246
Richards MP, Hedges REM (1999) Stable isotope evidence for similarities in the types of marine foods used by late Mesolithic humans at sites along the Atlantic Coast of Europe. J Archaeol Sci 26:717–722
Henisch BA (1976) Fast and feast: food in medieval society. Pennsylvania State University Press, University Park
Hofmann MI, Böni T, Alt KW, Woitek U, Rühli FJ (2008) Paleopathologies of the vertebral column in medieval skeletons. Anthropol Anz 66:1–17
Katzenberg MA, Lovell NC (1999) Stable isotope variation in pathological bone. Int J Osteoarchaeol 9:316–324
Kellner CM, Schoeninger MJ (2007) A simple carbon isotope model for reconstruting prehistoric human diet. Am J Phys Anthropol 133:1112–1127
Kjellström A, Storå J, Possnert G, Linderholm A (2009) Dietary patterns and social structures in medieval Sigtuna, Sweden, as reflected in stable isotope values in human skeletal remains. J Archaeol Sci 36:2689–2699
Knipper C, Peters D, Meyer C, Maurer AF, Muhl A, Schöne BR, Alt KW (2013) Dietary reconstruction in migration period central Germany: a carbon and nitrogen isotope study. Archeol Anthropol Sci 5:17–35
Krueger HW, Sullivan CH (1984) Models for carbon isotope fractionation between diet and bone. In: Turnland JE, Johnson PE (eds) Stable isotopes in nutrition. American Chemical Society, Washington, pp. 205–220
Krüttli A, Bouwman A, Akgül G, Della Casa P, Rühli F, Warinner C (2014) Ancient DNA analysis reveals high frequency of European lactase persistence allele (T-13910) in medieval central Europe. PLoS One 9(1):e86251. doi:10.1371/journal.pone.0086251
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230:241–242
Müldner G, Richards MP (2007) Stable isotope evidence for 1500 years of human diet at the city of York, UK. Am J Phys Anthropol 133:682–697
Nielsen-Marsh CM, Hedges REM (2000) Patterns of diagenesis in bone II: effects of acetic acid treatment and the removal of diagenetic CO3 2. J Archaeol Sci 27:1151–1159
O’Connell TC, Hedges REM (1999) Investigations into the effect of diet on modern human hair isotopic values. Am J Phys Anthropol 108:409–425
O’Connell TC, Kneale CJ, Tasevska N, Kuhnle GGC (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149:426–434
O’Leary MH (1988) Carbon isotopes in photosynthesis. Bioscience 38:328–336
Olsen KC (2013) A multi-isotope investigation of two medieval German populations: insight into the relationship among diet, disease, and tissue isotopic compositions, dissertation, University of Western Ontario
Olsen KC, White CD, Longstaffe FJ, von Heyking K, McGlynn G, Grupe G, Rühli FJ (2014) Intraskeletal isotopic compositions (δ 13C, δ 15N) of bone collagen: Nonpathological and pathological variation. Am J Phys Anthropol 153:598–604
Pieper R (2000) Dalheim: Pfarrort - Kloster - Staatsdomäne. Ardey-Verlag, Münster
Postan MM (1972) The medieval economy and society; an economic history of Britain, 1100–1500. University of California Press, Berkley
Pounds N (2005) The medieval city. Greenwood Press, Westport
Reitsema LJ, Vercellotti G (2012) Stable isotope evidence for sex- and status-based variations in diet and life history at medieval Trino Vercellese, Italy. Am J Phys Anthropol 148:589–600
Reitsema LJ, Crews DE, Polcyn M (2010) Preliminary evidence for medieval Polish diet from carbon and nitrogen stable isotopes. J Archaeol Sci 37:1413–1423
Reitsema LJ, Vercellotti G, Boano R (2016) Subadult dietary variation at Trino Vercellese, Italy, and its relationship to adult diet and mortality. Am J Phys Anthropol 160:653–664
Richards MP, Mays S, Fuller BT (2002) Stable carbon and nitrogen isotope values of bone and teeth reflect weaning age at the medieval Wharram Percy site, Yorkshire, UK. Amer J of Phys Anthropol 119:205–210
Richards MP, Fuller BT, Molleson TI (2006) Stable isotope paleodietary study of humans and fauna from the multi-period (iron age, Viking, and late medieval) site of Newark Bay, Orkney. J Archaeol Sci 33:122–131
Salamon M, Coppa A, McCormick M, Rubini M, Vargiu R, Tuross N (2008) The consilience of historical and isotopic approaches in reconstructing the medieval Mediterranean diet. J Archaeol Sci 35:1667–1672
Salazar-García DC, Richards MP, Nehlich O, Henry AG (2014a) Dental calculus is not equivalent to bone collagen for isotope analysis: a comparison between carbon and nitrogen stable isotope analysis of bulk dental calculus, bone and dentine collagen from same individuals from the medieval site of El Raval (Alicante, Spain). J Archaeol Sci 47:70–77
Salazar-García DC, Aura E, Olària C, Talamo S, Morales JV, Richards MP (2014b) Isotope evidence for the use of marine resources in the Eastern Iberian Mesolithic. J Archaeol Sci 42:231–240
Salazar-García DC, Romero A, García-Borja P, Subirà E, Richards MP (2016) A combined dietary approach using isotope and dental buccal-microwear analysis of humans from the Neolithic, Roman and medieval periods from archaeological site of Tossal de les Basses (Alicante, Spain). J Archaeol Sci Reports 6:610–619
Schoeller DA (1999) Isotope fractionation: why aren’t we what we eat? J Archaeol Sci 26:667–673
Schoeninger MJ (1985) Trophic level effects on15N/15N and 13C/12C ratios in bone collagen and strontium levels in bone mineral. J Hum Evol 14:515–525
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotope composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Ac 48:625–639
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios reflect marine and terrestrial components of prehistoric human diet. Science 220:1381–1383
Scully T (1995) The art of cookery in the middle ages. Boydell Press, Woodbridge
Shemesh A (1990) Crystallinity and diagenesis of sedimentary apatites. Geochim Cosmochim Ac 54:2433–2438
Smith BN, Epstein S (1971) Two categories of 13C/12C ratios for higher plants. Plant Physiol 47:380–384
Sommerville AD, Fauvelle M, Froehle AW (2013) Applying new approaches to modeling diet and status: isotopic evidence for commoner resiliency and elite variability in the Classic Maya lowlands. J Archaeol Sci 40:1539–1553
Strange MR (2006) The effect of pathology on the stable isotopes of carbon and nitrogen: implications for dietary reconstruction, Master’s Thesis, Binghamton University
Strott N, Czermak A, Grupe G (2007) Are biological correlates to social stratification depicted in skeletal finds? Investigation of early medieval separate burial grounds in Bavaria. In: Grupe G, Peters J (eds) Documenta archaeobiologiae, vol 7, skeletal series and their socio-economic context. Verlag Marie Leidorf GmbH, Rahden/Westf, pp. 67–86
Surovell TA, Stiner MC (2001) Standardizing infra-red measures of bone mineral crystallinity: an experimental approach. J Archaeol Sci 28:633–642
Tieszen LL, Fagre T (1993) Effect of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. In: Lambert JB, Grupe G (eds) Prehistoric human bone: archaeology at the molecular level. Springer-Verlag, Berlin, pp. 121–155
van der Merwe NJ, Vogel JC (1978) 13C content of human collagen as a measure of prehistoric diet in woodland North America. Nature 276:815–816
van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695
van Winter JM (2007) Medieval food culture. In: Spices and comfits: collected papers on medieval food. Prospect Books, Totnes, pp. 51–66
Virginia RA, Delwiche CC (1982) Natural 15N abundance of presumed N2-fixing and non-N2-fixing plants from selected ecosystems. Oecologia 54:317–325
von Heyking K (2012) Anthropologie einer frühstädtischen Randgruppe morphologische und archäometrische Untersuchung eines hoch- bis spätmittelalterlichen Armenhausgräberfeldes in Regensburg, Dissertation, Ludwig-Maximilians-Universität München
Warinner C, Rodrigues JFM, Vyas R, Trachsel C, Shved N, Grossmann J, Radini A, Hancock Y, Tito RY, Fiddyment S, Speller C, Hendy J, Charlton S, Luder HU, Salazar-García DC, Eppler E, Seiler R, Hansen LH, Castruita JAS, Barkow-Oesterreicher S, Teoh KY, Kelstrup CD, Olsen JV, Nanni P, Kawai T, Willerslev E, von Mering C, Lewis CM Jr, Collin MJ, Gilbert MTP, Rühli F, Cappellini E (2014a) Pathogens and host immunity in the ancient human oral cavity. Nat Gen 46:336–346
Warinner C, Hendy J, Speller C, Cappellini E, Fischer R, Trachsel C, Arneborg J, Lynnerup N, Craig OE, Swallow DM, Fotakis A, Christensen RJ, Olsen J, Liebert A, Montalva N, Fiddyment S, Mackie M, Canci A, Bouwman A, Rühli F, Gilbert MTP, Collins MJ (2014b) Direct evidence of milk consumption from ancient human dental calculus. Scientific Reports 4:7104
Weiner S, Bar-Yosef O (1990) States of preservation of bones from prehistoric sites in the near east: a survey. J Archaeol Sci 17:187–196
Woolgar CM, Serjeantson D, Waldron T (2006) Food in medieval England: diet and nutrition. Oxford University Press, Oxford
Wright LE, Schwarcz HP (1996) Infrared and isotopic evidence for diagenesis of bone apatite at Dos Pilas, Guatemala: paleodietary implications. J Archaeol Sci 23:933–944
Acknowledgements
The study was supported by the Social Sciences and Humanities Research Council doctoral scholarship, and grants from the Canada Research Chairs Program, the Natural Sciences and Engineering Research Council of Canada, the Canada Foundation for Innovation, the Ontario Research Fund, the University of Cape Town, the Generalitat Valenciana (VALi+d APOSTD 2014/123 and GV2015/060), and the Mäxi Foundation. The authors thank Kimberley Law, and Li Huang from Western’s Laboratory for Stable Isotope Science (LSIS) and John Lanham at the facilities of the University of Cape Town for their assistance. We wish to acknowledge K. W. Alt (formerly of the Institute of Anthropology, Johannes Gutenberg University, Germany) for providing the bone material. We also acknowledge Paul Szpak, Katherine Bishop, Winona Bailey, and Ian Newton for their contributions to sample preparation and analysis, and Zoe Morris for the site maps. This is LSIS contribution #317.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Olsen, K.C., White, C.D., Longstaffe, F.J. et al. Isotopic anthropology of rural German medieval diet: intra- and inter-population variability. Archaeol Anthropol Sci 10, 1053–1065 (2018). https://doi.org/10.1007/s12520-016-0432-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-016-0432-y