
Abstract
During 2015–2016, a strong El Niño event, nicknamed the “Godzilla El Niño,” similar to events in 1982–1983 and 1996–1997 occurred in the Pacific Ocean. Based on in situ and satellite observations, this paper aims to assess the water masses and chlorophyll-a surface distribution in the Bay of La Paz, Gulf of California, Mexico, after this event. A conductivity, temperature, and depth (CTD) rosette system equipped with a fluorescence sensor was used to obtain high-resolution measurements and surface water samples for spectrophotometric determinations of chlorophyll-a. Satellite images of sea surface temperature and chlorophyll-a were obtained for the dates when the cruise took place. The results showed the presence of three water masses: Gulf of California Water, Subtropical Subsurface Water, and Pacific Intermediate Water, the last present in Boca Grande. The highest surface chlorophyll-a concentrations observed were 4.51 mg m-3, 4.63 mg m-3, and 1.00 mg m-3 from the fluorescence sensor, spectrophotometric determination, and satellite observations, respectively. The horizontal distribution patterns for the three methods employed matched well. Despite the fact that Godzilla El Niño had several adverse effects in different regions, at the Bay of La Paz, they seem to have had no dramatic effect on the chlorophyll-a at the time of our observations, even though the values observed were higher compared to previous reports. This apparent lack of effect on the surface waters due to this climate disruption could be associated to the seasonal variability of the phytoplankton productivity, which presents a peak during autumn and winter and to the local fertilization mechanisms reported in the study area; however, further observations are required to confirm this.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
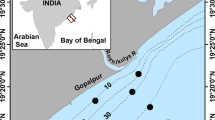
The Bay of La Paz (BoP), located on the southeastern margin of the Baja California peninsula between 24.1° and 24.9° N and 110.3° and 110.8° W, is the largest basin within the Gulf of California (GC), Mexico (Fig. 1). The region represents an environment of great biodiversity and productivity that supports an abundant variety of megafauna (Silverberg et al. 2014). The bay communicates with the GC by two openings: Boca Grande (wide and deep) to the north and San Lorenzo Channel (narrow and shallow) to the south. The regional climate is semiarid and the cloud cover is one of the lowest in Mexico, occurring only when a tropical storm or an atmospheric front approaches the area. Evaporation (300 mm/year) exceeds precipitation (180 mm/year) and river discharge is nonexistent (Monreal-Gómez et al. 2001).

Study area: left panel, Gulf of California, and right panel, Bay of La Paz. Bathymetry is in m; bold dots represents hydrographic and sampling stations
The BoP exchanges water masses with the GC predominantly through Boca Grande, involving Tropical Surface Water (TSW), Gulf of California Water (GCW), and Subsurface Subtropical Water (StSsW) (Lavín et al. 2009). The circulation pattern is dominated by a quasipermanent mesoscale cyclonic gyre (Monreal-Gómez et al. 2001) which induces nutrient pumping (Coria-Monter et al. 2017), differential distribution of phytoplankton (Coria-Monter et al. 2014), and differential aggregation of zooplankton (Duran-Campos et al. 2015) by different mechanisms that have been associated with the rotational movement of the eddy, transport of nutrients to the euphotic zone, and morphological structures, as well as the feeding habitats and ecological characteristics of each plankton group.
Because of its location, near the connection of the GC to the open Pacific Ocean, the site represents a sensitive recorder of regional variations in the gulf and the larger scale climate circulation of the subtropical Eastern Pacific Ocean (Pérez-Cruz 2006). The major source of interannual and monthly variability in the southern GC, and consequently the BoP, has been related to the El Niño/Southern Oscillation (ENSO) (Pérez-Cruz 2013; Guevara-Guillén et al. 2018), which classified them into two different ENSO: one has its most prominent equatorial Pacific sea surface temperature anomalies extending westward from the South American Coast and the other has its most prominent sea surface temperature anomalies extending toward eastern Pacific (Kug et al. 2009). For these reasons, they are referred to as the eastern Pacific ENSO and central Pacific ENSO (Paek et al. 2017).
In 2015, one of the strongest ENSO events on record was forecast by the National Oceanic and Atmospheric Administration (NOAA) to intensify in the winter of 2015–2016 (Klein 2015; Whitney 2015); it was named the “Godzilla El Niño” (Schiermeier 2015), with an intensity similar to the two previous very strong ENSO years documented, 1982–1983 and 1997–1998, the latter of which was also referred to as “the climate event of the twentieth century” (Changnon 2000) and both having a negative effect on the hydrographic properties of the water column as well as on the biological production. During the 1997–1998 ENSO, persistent warm anomalies throughout the GC were documented (Pavia et al. 2006). In the BoP, during the 1982–1983 ENSO, a major input of TSW that modified the plankton assemblages was documented (Pérez-Cruz and Molina-Cruz 1988), and an increase in the temperature resulting in the sinking of the thermocline and isotherms was reported (Obeso-Nieblas et al. 2004). Also, this climate disruption has been associated with changes that affect the habitat where some organisms perform biological functions, producing behavioral changes in the BoP populations (Shirasago-Germán et al. 2015). To date, the effects of the Godzilla El Niño event in the southern GC have been evaluated by Sánchez-Velasco et al. (2017) who, contrary to expectations, did not report dramatic changes in the total fish larvae abundance during this warm event, suggesting that the unexpected relatively high larval abundance indicated a possible adaptation of tropical species to prolonged periods of warming in the oceans. Similar observations where documented by Coria-Monter et al. (2018) who reported that, due to the dynamics and the mechanism of productivity enhancement in the area, the Godzilla El Niño event did not have as large of an impact on the phytoplankton biomass, expressed as chlorophyll-a, in the southern GC compared to other areas and previous strong El Niño events.
Although the Godzilla El Niño apparently did not have a negative impact on the biological production of the southern region of GC (Sánchez-Velasco et al. 2017; Coria-Monter et al. 2018), there is still controversy about the role of this event, particularly on the planktonic ecosystem, perhaps due to the lack of in situ observations, both hydrographic and phytoplankton samplings, from ships. This paper aims to assess the effects after the Godzilla El Niño on the hydrographic structure and surface phytoplankton biomass, expressed as chlorophyll-a concentrations, in the BoP, from in situ observations collected on an oceanographic research cruise during November of 2016 and by satellite observations. We hypothesized an incursion of warm water masses into the bay with a negative effect on the phytoplankton biomass.
Materials and methods
High-resolution hydrographic data and water samples for chemical analyses were obtained on board the R/V El Puma from the Universidad Nacional Autónoma de México during the multidisciplinary research cruise “Paleomar-II” to the BoP, November 15–21, 2016.
Sampling
A CTD Seabird 19 plus equipped with a calibrated WET Labs ECO series fluorometer sensor was used to record conductivity, temperature, pressure, and chlorophyll-a fluorescence at 43 stations within the bay (Fig. 1, right panel). A General Oceanics rosette equipped with 10-L Niskin bottles was used to take surface water samples (at 2 m depth) for spectrophotometric determination of chlorophyll-a. Immediately after collection, sub-samples of ~ 3 L were vacuum-filtered at less than 10 psi through 47-mm-diameter nitrocellulose membrane filters with a pore size of 0.45 μm (Millipore Corp., Bedford, MA, USA) and then stored in dark conditions at − 20 °C in plastic centrifuge tubes prior to processing.
Data reduction
The CTD data were initially processed using the manufacturer’s standard package (SBE Data Processing V.7.26.7), averaged to 1 dbar. Then, the conservative temperature (Θ, °C), absolute salinity (SA, g/kg), and density anomaly (σΘ, kg m-3) were calculated from the in situ temperature and practical salinity using the Thermodynamic Equation of Seawater 2010 (IOC et al. 2010). The chlorophyll-a concentration (mg m-3) was estimated from the in situ fluorescence using the nominal factory calibration of the fluorometer.
Satellite images of sea surface temperature (SST) and chlorophyll-a were obtained for the dates with concurrent in situ SST and chlorophyll-a measurements from the Moderate Resolution Imaging Spectroradiometer (MODIS-AQUA), launched in 2002 and still in operation. Available images were obtained from NASA Data Browser (https://oceancolor.gsfc.nasa.gov/cgi/browse.pl) on November 15, 17, 18, and 20, 2016. The images, with a spatial resolution of 1 km/pixel, were processed as follows: first, using level 2 (before mapping), values of SST and chlorophyll-a were extracted with SeaDAS version 7.4. In order to screen bad or low-quality data when generating the images, LAND, CLDICE, HILT, and STRAYLIGHT flags/masks were applied. LAND and CLDICE are raised then a mask pixel is applied to MODIS L2 products. HILT is flagged if any of the bands or detectors reach the physical saturation, whereas STRAYLIGHT indicates the influence of brightness of adjacent pixels on pixels reflectance value (Pieri et al. 2015). Then, maps of both variables were generated with Matlab (Mathworks, Inc. 2011).
Laboratory analyses
In the laboratory, chlorophyll was extracted for > 12 h in dark conditions at − 20 °C using 90% acetone as the solvent (Strickland and Parsons 1972; Parsons et al. 1984). The absorbance in three replicates was measured at 750, 664, 647, and 630 nm in a Genesys 10S UV-VIS (Thermo Scientific) spectrophotometer, and then the chlorophyll-a concentration (mg m-3) was calculated according to Strickland and Parsons (1972).
Results
According to the classification proposed by Lavín et al. (2009), the water mass distribution determined by the T-S diagram showed the presence of three water masses: GCW, StSsW, and Pacific Intermediate Water (PIW), the last in Boca Grande region (Fig. 2).

Conservative temperature (Θ, °C)—absolute salinity (g/kg) diagram: Tropical Surface Water (TSW), Gulf of California Water (GCW), Subtropical Subsurface Water (StSsW), and Pacific Intermediate Water (PIW).
The surface conservative temperature was in the range of 27.4 to 27.8 °C (Fig. 3a). Its horizontal distribution showed the presence of areas, one warm, located in the southwestern bay off San Juan de la Costa and reaching values of 27.7 °C, and another cold area, located at the central part of the bay with a value of 27.5 °C. A third warm area was observed with a value of 27.8 °C located close to Espíritu Santo Island.

Surface horizontal distributions (at 2 m depth): (a) in situ conservative temperature (°C), (b) chlorophyll-a (mg m-3) derived from the fluorescence sensor, and (c) chlorophyll-a (mg m-3) from spectrophotometric determination. The black dot on the west coast is the location of San Juan de la Costa.
The concentrations obtained by the fluorescence sensor rose in a range from 0.01 to 4.51 mg m-3 (Fig. 3b). An area of high concentration (> 3.8 mg m-3) was observed in the southwestern bay, close to San Juan de la Costa. Another area of relatively high concentration (> 1.2 mg m-3) was observed close to Espíritu Santo Island. The spectrophotometric determination of chlorophyll-a showed values from 0.13 to 4.63 mg m-3 (Fig. 3c). A consistent pattern of distribution was observed with an area of high concentration off San Juan de la Costa reaching values > 3.6 mg m-3. A secondary area of enhancement was observed close to the coast at 24.5° N latitude reaching values > 2.0 mg m-3.
The satellite images obtained for the dates of the cruise showed a pattern that matched well with the in situ measurements obtained at the surface. In November 15, the SST (Fig. 4a) and the chlorophyll-a images (Fig. 4e) strongly support the results derived from in situ measurements of both SST and chlorophyll-a levels, ranging from 27.8 to 28.5 °C and from 0.5 to 2.2 mg m-3, respectively. The images of November 17 showed SST values ranging from 28 to 29.5 °C, the highest anomalies observed at the southern bay and in a region close to Espíritu Santo Island (Fig. 4b), whereas the chlorophyll-a values (Fig. 4f) showed changes inside the bay with values similar as observed in Fig. 3b and c, in the range 0.5 to 2.2 mg m-3 with the highest concentration in the southern bay near the coast. The SST values, as well as the phytoplankton pigment concentrations on November 18 (Fig. 4c, g) and 20 (Fig. 4d, h) are also reasonably similar to those shown in the in situ measurements, showing values in the same order of magnitude. From Fig. 4, it is also evident the presence of filaments inside the bay, related to mesoscale processes reported as recurrent in the area.

Satellite images from MODIS-AQUA, for the period of study (November 15–20, 2016). Top panel shows the SST images in November (a) 15, (b) 17, (c) 18, and (d) 20. In the bottom panel are the chlorophyll-a images in November (e) 15, (f) 17, (g) 18, and (h) 20
While in situ chlorophyll-a fluorescence is a proxy for the chlorophyll-a concentration, and hence phytoplankton biomass, there is a large natural variation in the relationship between in situ fluorescence and extracted chlorophyll-a concentration due to variability in the specific absorption of the phytoplankton and in the fluorescence quantum yield (Roesler et al. 2017). This is the reason for the slight variation in the chlorophyll-a concentration observed between the methods used in this study.
Discussion
The water masses observed in this study are partially in agreement with previous studies in the BoP when a strong El Niño event took place. Monreal-Gómez et al. (2001) evaluated the hydrographic structure at the end of spring 1998 and observed the presence of Equatorial Surface Water (now renamed as TSW), GCW, and StSsW inside the bay and also at Boca Grande the presence of PIW. Similar observations were also made during the 1997–1998 El Niño when TSW filled the GC but was blocked on the Pacific side of the peninsula (Durazo and Baumgartner 2002). Obeso-Nieblas et al. (2014) pointed out that in periods not affected by El Niño, TSW and StSsW are not usually present in the southern GC, but during this episode, their presence varies in space showing a strong signal in the interior of the gulf and in the BoP. The presence of StSsW was also documented by Álvarez-Borrego and Schwartzlose (1979) who reported important incursions of this water mass during 1957 (an El Niño year) and by Castro et al. (2000) who reported a strong signal in the mouth of the GC during the 1997–1998 El Niño. Although the presence of the water masses observed in this study are partially in agreement with those described for the previous El Niño events, their presence seems to have no effect on the trophic state of the study area, even when considering that the high temperature of the GCW could confer an oligotrophic regime in the area. Monreal-Gómez et al. (2001) documented values of chlorophyll-a ranging from 0.04 to 0.50 mg m-3 at the end of spring 1998, under a predominance of GCW inside the BoP; however, our results indicate higher concentrations of chlorophyll.
To date, although there are reports on the negative effects of the Godzilla El Niño such as a strong convection of warm air masses along the Equator (Kintisch 2016), low nutrient values in the upper 350-m layer in the Pacific Ocean (Stramma et al. 2016), and a decrease in nutrient transport resulting in substantial reductions in phytoplankton biomass in the eastern North Pacific (Whitney 2015), the Godzilla El Niño seems to have had no apparent effect on the chlorophyll-a concentration observed inside the BoP. Similarly, Sánchez-Velasco et al. (2017) described no dramatic changes in the pelagic ecosystem and documented a relatively high abundance of fish larvae during the 2015 event in the southern GC as a result of the high tolerance of species with tropical affinity characterized by their domain in the Eastern Tropical Pacific to sustained warming. More recently, Coria-Monter et al. (2018) documented the absence of an effect of the Godzilla El Niño on the phytoplankton biomass in the southern portion of the GC, associated with the dynamics of the gulf which are characterized by the presence of intense tides that are produced mainly by co-oscillation with those of the Pacific Ocean, which in turn generate hydrodynamic processes such as mesoscale cyclonic eddies. Particularly at Bay of La Paz, Shirasago-Germán et al. (2015) also documented that the sea lion population inside the bay is not directly affected by the changes caused by El Niño.
The chlorophyll-a concentrations observed here seem to be more closely related to a typical seasonal variation reported. In the BoP, the chlorophyll-a mean annual cycle showed two concentration periods, a higher one from December to March and lower one from April to November (Guevara-Guillén et al. 2018). High values of chlorophyll-a (> 6.8 mg m-3) were observed during the winter season (Martínez-López et al. 2001). At the end of spring 2004, chlorophyll-a values of ~ 2.1 mg m-3 were associated with the pycnocline and nutricline (Coria-Monter et al. 2017), whereas in summer of 2009, the surface chlorophyll-a concentrations within the BoP were ~ 0.10 mg m-3 (Coria-Monter et al. 2014). The relatively high concentrations observed in this study suggest a strong relation with environmental conditions close to the winter season, due to the seasonal changes resulting from periods of heating and cooling of the surface layers; during summer, a strong stratification in the water column prevents flux of nutrients to the euphotic zone, and in contrast, winter winds and convection induce mixing in the water column which in turn promotes an increase in the phytoplankton biomass. This is in agreement with the paradigm for the seasonal variation in phytoplankton biomass in subtropical zones, where the limiting factor is the nutrient supply (Reynolds 2006).
Several mechanisms associated with the dynamics of the BoP could be involved in this apparent lack of effect due to the Godzilla El Niño. The presence of a quasipermanent mesoscale cyclonic eddy, which induces nutrient pumping, has been suggested as the main mechanism of fertilization within the bay (Coria-Monter et al. 2014; Coria-Monter et al. 2017). Coastal upwelling along the GC and the advection of high-nutrient waters to the BoP is another mechanism involved in the high productivity of the region (Martínez-López et al. 2001). Another mechanism of fertilization could be related to the presence of a bathymetric sill along Boca Grande, close to Espíritu Santo Island (Molina-Cruz et al. 2002), where mixing induced by the interaction between the currents and the bottom could promote a high concentration of nutrients. Additionally, this particular Godzilla El Niño occurred after a negative (cold) phase of the multivariate ENSO index, whereas the two previous mega ENSOs (1982–1983 and 1997–1998) occurred after a prolonged positive phase, which would set the stage for different ecosystem responses, especially in the higher trophic level that has a longer response time, as discussed by Coria-Monter et al. (2018). However, why were the “Godzilla El Niño” and previous extreme El Niños different? Paek et al. (2017) showed that the 1997–1998 event evolved in a way that suggests that it was dominated by the eastern Pacific El Niño dynamics, while the evolution of the Godzilla event suggests that a mixture of both the eastern Pacific and the central Pacific El Niño dynamics was at work, and the two events began to differ from each other in their decaying phases, during which SST anomalies retracted to the South American Coast beginning in January 1998 for the 1997–1998 event but stayed in the equatorial central Pacific from late winter to spring of 2016 for the 2015–2016 event. This framework shows that the two last strongest extreme El Niño events on record are very different in terms of their underlying dynamics and climate impacts. This would also set the stage for different ecosystem responses in the southern GC.
The consistent high chlorophyll-a concentration area observed off San Juan de La Costa by the three methods employed could be strongly related to the high phosphorite content of the lithology in the zone where mining takes place (Roca Fosfórica Mexicana II, Rofomex), which represents an additional fertilization process in the water column.
Satellite remote sensing observations of ocean color are crucial for understanding marine biology from a synoptic view with an adequate spatial and temporal resolution which in turn represent a powerful tool to evaluate the impacts of extreme events (Pieri et al. 2015), as ENSO events, bringing a unique opportunity to observe daily changing characteristic of the phytoplankton structure, a fundamental component of the ecosystem due to their capability to control the carbon dioxide uptake in the oceans and, therefore, in the global carbon cycle (Siegel et al. 2014). The results from in situ and satellite data presented here agrees on the important opportunity to combine both methods and highlight the values of effort to understand the impacts of ENSO events on high-productive ecosystems, and also show that combining both in situ observations from ships and satellite-derived observations are needed.
Based on the previous studies and our results, the principal remarks may be during winter, the northwesterly wind is strong, about 12 m s-1, inducing mixing and the sinking of the thermocline, and also a southward current along the Peninsula coast induces an output of GCW and inhibits the input of TSW. The mixing promotes the nutrient resuspension into the euphotic layer, then stimulated the phytoplankton production and high chlorophyll-a values. The results at the time of this research cruise have shown the absence of the TSW, which could be evidence that the Godzilla had no effect in the bay, or may be that the phytoplankton community has been in recovery after six months of the end of Godzilla
Conclusion
Based on the results, the surface chlorophyll-a concentration values apparently seem to have not been dramatically affected by the Godzilla El Niño, although the water masses detected in the T-S diagram showed an incursion of warm water as GCW and the concentrations observed are higher with respect to previous reports. This could be strongly related to the seasonal variability of the phytoplankton productivity, which presents a peak during autumn and winter and to the different fertilization mechanisms reported for the area, which promote nutrient availability for the phytoplankton. There are many other aspects such as differences in hydrographic properties and food-web dynamics that could be modified by the advection of warm oligotrophic waters, therefore many more detailed studies of these relationships in the BoP are required.
References
Álvarez-Borrego S, Schwartzlose RA (1979) Masas de agua del Golfo de California. Cienc Mar 6:43–63
Castro R, Mascarenhas AS, Durazo R, Collins CA (2000) Seasonal variation of the temperature and salinity at the entrance to the Gulf of California, Mexico. Cienc Mar 26(4):561–583
Changnon SA (2000) El Niño 1997-1998: the climate event of the century. Oxford University Press, New York 215 p
Coria-Monter E, Monreal-Gómez MA, Salas de León DA, Aldeco-Ramírez J, Merino-Ibarra M (2014) Differential distribution of diatoms and dinoflagellates in a cyclonic eddy confined in the Bay of La Paz, Gulf of California. J Geophys Res Oceans 119:6258–6268. https://doi.org/10.1002/2014JC009916
Coria-Monter E, Monreal-Gómez MA, Salas de León DA, Durán-Campos E (2017) Wind driven nutrient and chlorophyll-a enhancement at a cyclonic circulation in Bay of La Paz, Gulf of California, Mexico. Estuar Coast Shelf Sci 196:2990–2300
Coria-Monter E, Monreal-Gómez MA, Salas de León DA, Durán-Campos E (2018) Impact of the “Godzilla El Niño” event of 2015-2016 on sea surface temperature and chlorophyll-a in the southern Gulf of California, Mexico, as evidenced by satellite and in situ data. Pac Sci 72(4):411–422
Duran-Campos E, Salas de León DA, Monreal-Gómez MA, Aldeco-Ramírez J, Coria-Monter E (2015) Differential zooplankton aggregation due to relative vorticity in a semi-enclosed bay. Estuar Coast Shelf Sci 164:10–18
Durazo R, Baumgartner TR (2002) Evolution of oceanographic conditions off Baja California: 1997-1998. Progr Oceanogr 54(1-4):7–31
Guevara-Guillén C, Shirasago-Germán B, Pérez-Lezama EL, Obeso-Nieblas MJ (2018) Variability of chlorophyll-a concentration and halocline depth in La Paz Bay (Gulf of California, Mexico) and the relationship to Pacific Large-Scale climatic phenomena. J Coast Res 34(4):920–927
IOC, SCOR, IAPSO (2010) The international thermodynamic equation of seawater 2010. Calculation and use of thermodynamic properties. Intergovernmental Oceanographic Commission, Manuals and Guides No. 56, UNESCO (English), 196 pp
Kintisch E (2016) How a ‘Godzilla’ El Niño shook up weather forecasts. Science 352(6293):1501–1502
Klein K (2015) NOAA predicts strong El Niño. Eos, Transactions American Geophysical Union 96. https://doi.org/10.1029/2015EO035535
Kug J-S, Jin F-F, An S-I (2009) Two types of El Niño events: cold tongue El Niño and warm pool El Niño. J Clim 22:1499–1515
Lavín MF, Castro R, Beier E, Godínez VM, Amador A, Guest P (2009) SST, thermohaline structure, and circulation in the southern Gulf of California in June 2004 during the North American Monsoon Experiment. J Geophys Res Oceans 114:C02025. https://doi.org/10.1029/2008JC004896
Martínez-López A, Cervantes-Duarte R, Reyes-Salinas A, Valdez-Holguín JE (2001) Cambio estacional de la clorofila a en la Bahía de La Paz, B.C.S., México. Hidrobiologica 11(1):45–52
Mathworks, Inc (2011) MATLAB. Global Optimization Toolbox: User’s Guide (r2011b). www.mathworks.com/help/pdf_doc/gads/gads_tb.pdf
Molina-Cruz A, Perez Cruz L, Monreal-Gómez MA (2002) Laminated sediments in the Bay of La Paz Gulf of California: a depositional cycle regulated by pluvial flux. Sedimentology 49:1401–1410
Monreal-Gómez MA, Molina-Cruz A, Salas de León DA (2001) Water masses and cyclonic circulation in Bay of La Paz, during June 1998. J Mar Syst 30:305–315
Obeso-Nieblas M, Shirasago B, Sánchez-Velasco L, Gaviño-Rodriguez J (2004) Hydrographic variability in Bahia de La Paz, B.C.S, Mexico during the 1997-1998 El Niño. Deep-Sea Res II 51:689–710
Obeso-Nieblas M, Gaviño-Rodríguez JH, Obeso-Huerta H, Muñoz-Casillas SI (2014) Spatial termohaline variability, water masses and geostrophic circulation in Bahía de La Paz, Gulf of California. Rev Biol Mar Oceanogr 49(3):413–426
Paek H, Yu J-Y, Qian C (2017) Why were the 2015/2016 and 1997/1998 extreme El Niños different? Geophys Res Lett 44. https://doi.org/10.1002/2016GL071515
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis. Pergamon Press, Oxford, 173 p
Pavia EG, Graef F, Reyes J (2006) PDO-ENSO effects in the climate of Mexico. J Clim 19:6433–6438
Pérez-Cruz L, Molina-Cruz A (1988) El Niño 1983: effect on the distribution of silicoflagellates in the Gulf of California. Cienc Mar 14(3):9–38
Pérez-Cruz L (2006) Climate and ocean variability during the middle and late Holocene recorder in laminated sediments from Alfonso Basin, Gulf of California, Mexico. Quat Res 65:401–410
Pérez-Cruz L (2013) Hydrological changes and paleoproductivity in the Gulf of California during middle and late Holocene and their relationship with ITCZ and North American Monsoon variability. Quat Res 79:138–151
Pieri M, Massi L, Lazzara L, Nuccio C, Lapucci C, Maselli F (2015) Assessment of three algorithms for the operational estimation of [CHLA] from MODIS data in the Western Mediterranean Sea. Eur J Remote Sens 48:383–401
Reynolds C (2006) Ecology of Phytoplankton. Cambridge University Press, Cambridge 535 p
Roesler C, Uitz J, Claustre H, Boss E, Xing X, Organelli E, Briggs N, Bricaud A, Schmechtig C, Poteaaus A, D’Otenzio F, Ras J, Drapeau S, Haëntjens N, Barbieux M (2017) Recommendations for obtaining unbiased chlorophyll estimates from in situ chlorophyll fluorometers: A global analysis of WET Labs ECO sensors. L&O Methods 15:572–585
Sánchez-Velasco L, Beier E, Godínez VM, Barton ED, Santamaría-del-Angel E, Jiménez-Rosemberg SPA, Marinone SG (2017) Hydrographic and fish larvae distribution during the “Godzilla El Niño 2015-2016” in the northern end of the shallow oxygen minimum zone of the Eastern Tropical Pacific Ocean. J Geophys Res Oceans 122:2156–2170. https://doi.org/10.1002/2016JC012622
Schiermeier Q (2015) Hunting the Godzilla El Niño. Nature 526:490–491
Shirasago-Germán B, Pérez-Lezama EL, Chávez EA, García-Morales R (2015) Influence of el Niño-Southern oscillation on the population structure of sea lion breeding colony in the Gulf of California. Estuar Coast Shelf Sci 154:69–76
Siegel DA, Buesseler KO, Doney SC, Sailley, SF, Behrenfeld MJ, Boyd PW (2014) Global assessment of ocean carbon export by combining satellite observations and food-web models. Global Biogeochem Cycles 28:181–196. https://doi.org/10.1002/2013GB004743
Silverberg N, Aguirre-Bahena F, Mucci A (2014) Time-series measurements of settling particulate matter in Alfonso Basin, La Paz Bay, southwestern Gulf of California. Cont Shelf Res 84:169–187
Stramma L, Fischer T, Grundle DS, Krahmann G, Bange HW, Marandino CA (2016) Observed El Niño conditions in the eastern tropical Pacific in October 2015. Ocean Sci 12:861–873
Strickland JDH, Parsons TR (1972) A practical handbook of seawater analysis. 2nd edn. J Fish Res Board Can. No. 067 (167), 311 pp
Whitney FA (2015) Anomalous winter winds decrease 2014 transition zone productivity in the NE Pacific. Geophys Res Lett 42:428–431
Acknowledgments
The ship time for the Paleomar-II research cruise onboard the R/V El Puma was funded by the Universidad Nacional Autónoma de México (UNAM). We would like to express our gratitude to the NASA Ocean Biology Processing Group (OBPG) for the data products used in this study. We thank the participants of the research cruise, including the captain and crew. We acknowledge Jorge Castro by improving the figures, F. Sergio Castillo-Sandoval for technical support during the chemical analyses and Marco Belacicco for suggestions and comments. We appreciate the helpful comments by two anonymous reviewers.
Funding
Consejo Nacional de Ciencia y Tecnología (CONACYT) sponsored E. Coria-Monter during this study. This study was supported by the Instituto de Ciencias del Mar y Limnología (UNAM) (grants number 144, 145, and 342).
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: Haroun Chenchouni
Rights and permissions
About this article

Cite this article
Coria-Monter, E., Monreal-Gómez, M.A., Salas de León, D.A. et al. Water masses and chlorophyll-a distribution in a semi-enclosed bay of the southern Gulf of California, Mexico, after the “Godzilla El Niño”. Arab J Geosci 12, 473 (2019). https://doi.org/10.1007/s12517-019-4636-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-019-4636-1