
Abstract
Agriculture sector is facing a lot of constraints such as climate change, increasing population and the use of chemicals, and fertilizers which have significant influence on sustainability. The excessive usage of chemical fertilizers and pesticides has created a significant risk to humans, animals, plants, and the environment. To reduce the dependency on chemical fertilizers and pesticides a biological-based alternative is required. Seaweeds are essential marine resources that contain bioactive compounds and they have several uses in agriculture. The use of seaweed extracts in agriculture can mitigate stress, enhance nutrient efficiency, and boost plant growth. The use of seaweed extracts and their components activate several signaling pathways and defense-related genes/enzymes. In this review, an attempt has been made to explain how seaweed extracts and their bioactive components induce tolerance and promote growth under stress conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The agricultural industry is confronted with several challenges including increasing the production to feed world’s rising population and increasing resource use efficiency, all while limiting the environmental impact on ecosystems and human health. The agro-ecosystem is consequently disturbed by extreme climatic events and such changes might have impact on both the quality and quantity of crops. The increasing level of pollutants, as well as the negative impact of climate change on soil microbiome, are additional constraints to agriculture. Increase in atmospheric carbon dioxide (CO2), temperature, and humidity, in particularly, are predicted to exacerbate the frequency of major plant diseases, insect infestation, and the migration of major pathogens and insect pests into new geographic areas. To increase agricultural production and fulfil the food requirement of increasing population inorganic fertilizer and pesticides are used liberally and it has serious implications on both agro-ecosystem and human health. In developing countries, farmers rely on the use of inorganic pesticides, insecticides, chemical fertilizers, and herbicides and their use has become an unavoidable risk in agriculture. Moreover, agriculture World is the backbone of our economy, and extended and excessive use of these inorganic fertilizers results in massive economic losses. Though their use has led to increased crop production their improper employment has caused agro contamination which is a troublesome issue. The use of chemical fertilizer could impact the agricultural environment by changing its parameter such as eutrophication, reducing the crop yield, salinization of soil, and increasing in pH of the soil (Prakash et al. 2018). To enhance agricultural production several technological advancements have been proposed which also include the identification of new plant biostimulants and efficient methods for their application. The use of plant bio-stimulants is promising and environmental friendly (Colla and Rouphael 2015) and the alternative to chemical fertilizer would be the use of organic fertilizer such as seaweed liquid fertilizer, humic acid, vermicompost, and essential microorganisms and these are biodegradable, non-toxic, and non-polluting. Kauffman et al.(2007) reported that bio-stimulants are not fertilizers, but when applied in low quantities improve plant growth (Kauffman et al. 2007). The biostimulant sector is expected to develop at an annual pace of 11.24% and reach USD 4.9 billion by 2025 (Caradonia et al. 2018). There is a growing interest in the farming sector for novel biostimulant compounds and much research is being conducted in this emerging sector of the business. Seaweeds serve as a valuable source of several elicitors that may be used for inducing physiological development and multitude of defensive responses. This review discusses the utilisation of various seaweed extracts (SEs) as biostimulants to mitigate biotic and abiotic stress. The mechanism of action, composition and function of several seaweed are discussed elaborately.
Seaweeds
Seaweeds or marine algae are marine aquatic plants that inhabit the coasts of oceans and seas. These are macroscopic algae that are found attached to the solid bottom of rocks, shells, and other plant material. It constitutes about 6000 species with a great diversity of forms and size and only 5% of it is being used. Seaweeds are a rich source of bioactive compounds and produce secondary metabolites with a varying range of biological functions such as antibacterial (Singh and Chaudhary 2010), antifungal (De Corato et al. 2017), antiviral, anti-inflammatory, nematocidal, and anticoagulant (Caijiao et al. 2021). They are divided into three categories based on their pigmentation, such as red algae (Rhodophyta), green algae (Chlorophyta), and brown algae (Phaeophyta) (Kim 2011). They are rich in minerals (Mg, Ca, P, K, and I), proteins, vitamins, undigestable carbohydrates, and fibers. The composition of minerals varies from species to species and the source of dietary fibers varies chemically and physiochemically from those of the plants found in the land (Jiménez-Escrig and Cambrodón 1999). Seaweed active compounds are used in wide range of sector such as food, wastewater treatments, agriculture, pharmaceutical, and cosmetics (Pereira and Correia 2015). Beyond these applications, the global demand for seaweed usage has increased. Seaweeds are beneficial alternative feeds for domesticated animals, because they contain valuable source of nutrients, particularly chelated microminerals, the availability of which is greater than that of inorganic minerals; complex carbohydrates with prebiotic properties; and pigments and polyunsaturated fatty acids that are beneficial to consumer health.
Seaweed is extremely important in agriculture and human lives and the huge seaweed reserve along the world's coastal areas should be used more effectively and strategically to reduce the waste of these vital resources (Parmar et al. 2017). The seaweeds have been utilized as organic fertilizer for centuries and in coastal regions seaweeds have been used either directly or in the composted form to improve crop productivity (Craigie 2010). Seaweeds are employed as biofertilizers to compensate for soil nutrient deficiencies. Seaweed extract (SEs) includes regulators, plant growth hormones, carbohydrates, auxins, gibberellins, and vitamins which can help to improve crop productivity and to maintain soil fertility. The SEs target several pathways to improve tolerance under stress and the certain plundering caused by bacteria, fungi, insects, and parasites can be reduced in plants. SEs biochemical composition is complex and as a result, understanding seaweed mode of action is exceedingly complex. Because of the various interactions between a large number of bioactive compounds within the same extract, a multidisciplinary approach is required.
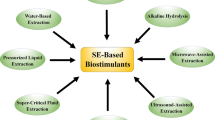
The extraction of seaweed is still the most important phase in the production of agronomically efficient products. The seaweed extract is obtained through various processes but the widely used method is alkali extraction with or without heating (Khan et al. 2009). The seaweed-based biostimulant is produced in either liquid or soluble powder from a different range of seaweeds (Michalak et al. 2016). The use of seaweed extract’s intrinsic chelating characteristics, which prevent trace metal ions from precipitating, the extract is commonly fortified with plant fertilizer and micronutrients. The raw materials used and the extraction procedure have a significant impact on the various mechanism of action of seaweed-derived biostimulants (Shukla et al. 2019).
There are many seaweeds extracts available in the market and a considerable amount of seaweed is used as nutrient supplements and as biostimulants or biofertilizers. The most commonly manufactured seaweed species include Ascophyllum nodosum, Macrocystis pyrifera, Ecklonia msaxima, Sargassum species, Laminaria species, and Fucus serratus (Sharma et al. 2013). Numerous studies have found that seaweed extract applications have a wide range of favourable impact on crops (Fig. 1) which include early seed germination and establishment, improved crop performance and yield, increased resilience to biotic and abiotic stress and increased postharvest shelf-life of perishable products. For example A. nodosum one of the most studied seaweed when used as foliar spray improved the flowering phenomenon in tomatoes (Dookie et al. 2021).

The schematic illustration of different application of seaweed extracts in plants
The biostimulant prepared from different seaweeds elicits plant defense through a distinct mode of action. The effect of specified applications of seaweed extract (rates and timings are crucial for specific responses to be evoked) is also linked to the activation of many molecular and biochemical changes in the treated plants, demonstrating the possible impact of SEs on genetic pathways.
Effect of seaweed extract on plant growth and development
The seaweed extract has been demonstrated to positively affect seed germination and plant development at all phases including harvest and post-harvest (Table 1). Seaweed products have been demonstrated to improve germination rate and a considerable increase in seedling vigor by enhancing root length and density. Recently it was reported that Padina gymnospora, Gracilaria edulis, and Ulva fasciata aqueous extracts are used as seaweed biofertilizers for the seed germination of Capsicum annuum (Shamya Arokia rajan et al. 2020). Rengasamy et al.(2015) reported that the use of eckol and phloroglucinol isolated from the Ecklonia maxima increased the germination rate in maize seeds (Rengasamy et al. 2015). This has been linked to the activity of the enzyme α-amylase, which facilitates the conversion of starch to simple sugars in the roots of maize seedlings treated with eckol and phloroglucinol-treated. The sugar produced was transported to the embryo in order to provide the necessary energy for metabolism. The use of Kappaphycus alvarezii seaweed extract on maize during the grain-filling stage boosted yield attributes such as cob length, the number of grains per cob, and length of grain fill (Trivedi et al. 2018). Different concentration of K. alvarezii was applied together with 100% of the required amount of fertilizer on maize (Singh et al. 2015) and potato tuber (Pramanick et al. 2017) and increased growth, yield, and quality was observed in both studies. Treatment with A. nodosum has been shown to promote growth and nutrient uptake from the soil in Brassica napus (winter rapeseed) and strawberry (Jannin et al. 2012). Application of A.nodosum to wheat increased the nutrient uptake and the grain nutrient accumulation was linearly associated with grain production showing enhanced soil nutrient absorption and remobilization to the reproductive organs (Stamatiadis et al. 2018).
Seasol a commercially available extract prepared from the seaweeds Duvillaea potatorum and A. nodosum increased the root length density and the yield (Mattner et al. 2018). Soybean treated with A. nodosum extract showed increased photosynthetic efficiency and chlorophyll content while also promoting the growth of the root system (do Rosário Rosa et al. 2021). Seaweed extract-treated tomato and pepper showed enhanced chlorophyll content (Ali et al. 2019). The extract obtained from Macrocystis pyrifera promoted the growth of tomato plants and increased the adventitious root formation in the mung bean (Briceño-Domínguez et al. 2014).
Finnie and Staden (1985) reported that Ecklonia maxmia seaweed promoted the in-vitro growth of tomato root. Both root elongation and root extension were significantly enhanced when applied (Finnie and van Staden 1985). Liquid seaweed extracts (LSEs) derived from seaweed such as Ulva lactuca, Caulerpaa sertularioides, P. gymnospora and Sargassum liebmannii enhanced germination response and increased the physiological traits of tomato. When compared to foliar spray, soil drench application was shown to be more effective (Hernández-Herrera et al. 2014). Application of U. fasciata and S. lacerifolium to soil boosted radish growth, improved germination percentage, and improved morphological and biochemical indices (Ahmed et al. 2020). The wheat seed primed with an aqueous extract of Ulva linza showed a substantial increase in physiological characteristics and mitotic index. New proteins were expressed in seedlings treated with an extract which might attribute to the activity of a bioactive substance in the extract (Hamouda et al. 2022a). Wine grape yield of several cultivars was increased by an average of 14.7% by foliar application of Durvillaea potatorum and A. nodosum (Arioli et al. 2021).
A study using A. nodosum extract in spinach showed enhanced biomass, protein content, chlorophyll and carotenoid content, flavonoids and antioxidant activity. The increase in biomass is associated with an increase in the expression of the Glutamine synthetase 1 (GS1) which is involved in nitrogen integration (Xu and Leskovar 2015). The increased expression of glutathione reductase (GR), and choline monooxygenase (CMO) was linked to increase in chlorophyll content. The upregulation of glutathione reductase (GR), thylakoid bound peroxidase (tAPX) and monodehydroascorbate reductase (MDHAR) was linked to the increase in chitinase activity, phenolics and flavonoids. These genes were discovered to be related to the phenylpropanoid and flavonoid pathways which are known to promote growth and improve overall nutrition (Fan et al. 2013). Flavonoids are secondary metabolite which is important for plant development. Chalcone isomerase (CHI), a key enzyme in the biosynthesis of flavanone precursors and phenylpropanoid plant defense compounds were also upregulated in response to seaweed extract treatment. The use of A. nodosum extracts improved the nitrogen use efficiency in barley plants by increasing the N content in barley shoot which was linked with the upregulation of root nitrate transporter (NRT1.1, NRT2.1, and NRT 1.5) (Goñi et al. 2021). Jannin et al. (2012) reported that A. nodosum application to Brassica napus increased the uptake of nitrogen and sulfate content in root and shoot. The expression of genes (BnNRT1.1 and BnNRT2.1) which encode nitrate transporter was shown to increase in roots of the treated plant (Jannin et al. 2012).The investigation of molecular responses in plant to A.nodosum extract treatment revealed that the increase in chlorophyll concentration was primarily due to downregulation of stay green protein (SRG) which was able to limit leaf protein degradation and protect chlorophyll degradation during leaf senescence. Senescence-associated cysteine protease was down-regulated whereas genes involved in photosynthesis, cell metabolism and S and N metabolism were up regulated (Rayorath et al. 2008; Jannin et al. 2012).
Cell wall undergo structural changes upon biotic and abiotic stress and they store carbohydrate for plant growth. Previously it was reported that Kelpak® can alter the allocation of carbon between cellulose synthesis and secondary cell wall by regulating sucrose synthase expression (Kocira et al. 2020). Sucrose is required for cell wall synthesis and several mechanisms contribute to cell wall synthesis (Decker and Kleczkowski 2019). In another study it was reported that with application of S. vulgare in tomato and sweet pepper cell-wall biosynthesis-related transcripts were upregulated. The MYB transcription are significantly upregulated and are related to secondary cell wall formation. The MYB are activated by SND1 (Secondary wall-associate NAC domain protein 1) and it activates the biosynthesis of lignin, cellulose and hemicellulose (Miedes et al. 2014; Decker and Kleczkowski 2019).
Phytohormones are necessary for plant growth and development. Thus, the alteration of the innate phytohormone pool by the seaweed-based biostimulants can have a major impact on plant development. Recent research has shown that seaweed and its components can influence the expression of genes involved in the biosynthesis of growth hormones such as NCED3 (Nine-cis-epoxycarotenoid dioxygenase3), ERF2 (Ethylene-responsive transcription factor 2), ICS2 (Isochorismate synthase 2) etc. The genes are upregulated in plants upon treatment of seaweed extracts (Ali et al. 2019).
When extracts are applied at the recommended levels, (do Rosário Rosa et al. 2021) the growth hormones present within the extract is insufficient to invoke physiological changes in plants (Górka and Wieczorek 2017). However, the components within the seaweed extract may induce innate pathways for the biosynthesis of growth hormones in plants. Aremu et al. (2015) isolated eckol and phloroglucinol from Ecklonia maxima and investigated their effects on Eucomis autumnails. It was observed that exogenous application of eckol and phloroglucinol caused increase in rise of auxin level which resulted in a 1.5-fold increase in the root length of the treated plant. The compounds influence auxin’s root-lengthening activity by preventing decarboxylation and acting as a cofactor which promotes the breakdown of the IAA oxidase (Aremu et al. 2015). It has been observed that biostimulants promote overall plant growth, physiological processes, agronomic aspects, and yield quality of crops (Yao et al. 2020).
Mitigating plant abiotic stress using seaweed-based biostimulants
Abiotic stress such as temperature extremes (heat and cold), drought, salinity, and heavy metals are key issues restricting agricultural growth and sustainability across the world. It negatively impacts growth and yield formation. These abiotic stresses are interrelated and can manifest osmotic stress, ion distribution failure, and plant cell homeostasis. Developing stress-tolerant plants through genetic engineering is a challenging task and has achieved limited success (Agarwal et al. 2012). As an early reaction to abiotic stress plants undergo internal oxidative stress by excess overproduction of reactive oxygen species (ROS). An imbalance is created between ROS production and the ability to detoxify and remove intermediate substances (Bencze et al. 2004). The increased production of ROS causes structural alteration to DNA and proteins, inhibition of antioxidant enzymes, and activation of programmed cell death (PCD) (Huang et al. 2019). To counteract the deleterious effects, plants use a wide range of defense mechanisms (Fig. 2). In order to scavenge the ROS, plants employ both enzymatic and nonenzymatic antioxidants such as superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR) and phenolic compounds (Gruszka et al. 2018; Yadav et al. 2019). Prominent results prove that seaweed can mitigate abiotic stress in plants (Table 2). For example, pre-treatment with A. nodosum based biostimulant had a favourable effect on Arabidopsis under drought-stressed conditions. It also altered the expression levels of genes implicated in ABA-responsive and antioxidant system pathways resulting in improved photosynthetic performance than untreated plants (Santaniello et al. 2017). The use of polysaccharides extracted from brown seaweed Lessonia nigrescens improved the salt tolerance level in wheat plants (Zou et al. 2019). The treatment minimized the oxidative damage in plants by increasing the antioxidant activity of SOD, CAT, and POD (peroxidase) enzymes by controlling the relative electric leakage and malondialdehyde content which are two parameters that determine membrane permeability and lipid peroxidation. Similarly, the use of U. rigida improved the salt tolerance in wheat plant and protected it from oxidative degradation, and increased the enzymatic activities in plants (Chernane et al. 2015). The application of polysaccharides extracted from Grateloupia filicina mitigated the tolerance of rice seedling to salinity stress and improved rice growth under salinity conditions (Liu et al. 2019). Milkweed seedling treated S. angustifolium extract can tolerate salinity up to 15 dS m−1 and the survival rate was increased by 69%. The treatment increased the growth parameters with a decrease in electrolyte leakage (Bahmani Jafarlou et al. 2022). Similarly, supplementation of S. muticum and Jania rubens provided salt stress amelioration in chickpea and enhanced activity of SOD and APX were related with improved growth parameters. Moreover, four key amino acids including serine, threonine, proline, and aspartic acid were identified from roots that contribute to stress reduction (Abdel Latef et al. 2017). Garcilaria dura extract applied to wheat elevated drought stress tolerance and increased the biomass and crop yield (Sharma et al. 2019). The foliar application of K. alvarezii significantly mitigated the water stress in maize with increased seed yield primarily by increasing all yield attributes, particularly cob length, the number of grains per cob, and length of grain fill. It also resulted in increased antioxidant activity and reduced reactive oxygen species levels, leading to a reduced degree of oxidative stress in treated plants. (Trivedi et al. 2018).

The schematic illustration highlights the application of seaweed extract on plants and the response of plants to abiotic stresses. Seaweed extract application to plants is perceived by a complex sensing mechanism in which various secondary messenger’s function (Abscisic acid (ABA), phytohormones, Ca2+, etc.,). The secondary messengers cause a myriad of downstream processes such as the activation of MAPKs (Mitogen-activated protein kinases), CIPKs (Calcineurin B-like proteins), PPs (Protein phosphatases) and CDPKs (Calcium-dependent protein kinases) activating the transcriptional factors and they regulate the stimulation of antioxidant enzymes thus scavenging ROS production. The other stress-inducible genes are involved in the alteration of cell wall biosynthesis, carbohydrate metabolism, secondary metabolites, and regulation of Na+ and K+ transporters thus altering the physiological adaption of plants to abiotic stress and enhancing the tolerance to abiotic stress. Note MAPK mitogen-activated protein kinase, CDPKs calcium-dependent protein kinases, PPs protein phosphatases, CIPKs calcineurin B-like protein (CBL)-CBL-interacting protein kinase, WRKY tryptophan arginine lysine tyrosine, MYB myeloblastosis, DREB dehydration responsive element-binding, AP2-APETALA2, EREB ethylene responsive element binding factor, LEA late embryogenesis abundant, DHN dehydrin, FIB1a fibrillarin1a, GST glutathione S-transferase, SOD superoxide dismutase, CAT catalase, GSH glutathione, APX ascorbate peroxidase, POD peroxidase, PGA-3 phosphoglyceric
Seaweed extract could be another approach for dealing with temperatures that impede the growth and development of plants. Application of A. nodosum extract (ANE) to tomato seedling enhanced heat stress tolerance and showed significant improvement in flower development and fruit production. The constituents in ANE formulation were associated with an accumulation of soluble sugars, and increase in heat shock proteins (HSPs) gene transcription in heat exposed tomato flowers before fertilization (Carmody et al. 2020). Algafect a commercial seaweed extract (A. nodosum, Fucus spp., and Laminaria spp.) was applied to maize and it enhanced the low temperature tolerance of root zone during early growth. On application of seaweed extract, the SOD activity was increased in root and leaf tissue as it plays an important role in antioxidative stress defense (Bradáčová et al. 2016).
Seed priming has emerged as an essential approach for producing plants that are resistant to various stress and it is a simple and alternative practice compared to conventional methodologies (breeding or transgenics). Seaweed extract is successfully utilized as a priming agent to enhance stress tolerance in plants (Bhanuprakash and Yogeesha 2016). Priming wheat seeds with Hormophysa cuneiformis and Actinotrichia fragilis reduced salt stress and it also enhanced the growth and increased the level of antioxidant activity in plants (Latef et al. 2021).
Maize and black-eyed pea seeds primed with seaweed liquid extract (SLE) prepared from Ulva fasciata, Cystoseira compressa and Laurencia obtusa exhibited remarkable tolerance to salinity stress and increased the seedling morphological parameter (Hussein et al. 2021). The negative effects of salinity in tomato seedlings were attenuated by priming the seeds with Ulva lactuca methanol extract. The hydrogen peroxide concentration and antioxidant activity were decreased in plants treated with methanol extract and a high concentration of total phenol and soluble sugars were recorded in methanol extract suggesting these compounds might play a key role in mitigating salinity stress in plants (El et al. 2021). The use of natural plant biostimulants has been recommended to boost plant resilience to abiotic environmental stresses (Nephali et al. 2020; Deolu-Ajayi et al. 2022).
Mitigating plant biotic stress using seaweed-based biostimulants
The emergence of infectious plant pathogens has increased due to changes in climatic conditions and intensive agriculture (Anderson et al. 2004). Even though new technology, research, and products help agriculture in maintaining integrated management and farming practices, pathogens (primarily bacteria, viruses, and fungi) have decreased agricultural productivity inflicting economic damage on at least 10% world's food supply. To counteract the biotic stresses induced by pathogenic infection, plants activate different defense mechanisms. Mitigation of plant biotic stress has been successfully achieved by priming the plant molecular defense by treatment with different chemicals, natural substances, and phytohormones (Aranega-Bou et al. 2014). SEs and their bioactive compounds are successfully used as defense priming agents in plants to trigger an immune response in plants (Craigie 2010; Islam et al. 2020).
Many seaweed extracts have been reported to exhibit antagonistic activity both in-vitro and in the fields (Table 3). For example, Ulva lactuca reduced the fusarium wilt severity when sprayed onto leaves of common beans (de Borba et al. 2019). The use of Ochtodes secundiramea and Laurencia dendroidea extracts in papaya and banana efficiently inhibited Colletotrichum gloeosporioides, a fungal species that severely hampers fruits post-harvest (Machado et al. 2014). The cucumber plants sprayed with A. nodosum exhibited enhanced resistance against Phytophthora melonis by induction of defense enzymes and genes. Moreover, the plants showed a higher level of phenolic accumulation compared to the control (Abkhoo and Sabbagh 2015). Similarly, the use of A. nodosum in carrot and broccoli successfully controlled the foliar diseases caused by different fungal pathogens (Jayaraj et al. 2008; Mattner et al. 2014). The extracts of brown algae Cystoseria myriophylloides, Laminaria digitata and Fucus spiralis inhibited the growth of tomato pathogens Verticillium dahlia and Agrobacterium tumefaciens both in-vitro and in the greenhouse. It was also reported that defense enzyme polyphenol oxidase and peroxidase activity were substantially increased in comparison to control (Esserti et al. 2016). The extracts of Ulva lactuca, S. filipendula and Gelidium serrulatum suppressed the Alternaria solani and Xanthomonas campestris pv vesicatoria in tomato plants. The seaweed-extract treated plant showed reduced severity of diseases and induced salicylic acid and jasmonate signaling pathways (Ramkissoon et al. 2017). The root-knot diseases in eggplant and watermelon caused by fungi Fusarium solani were suppressed using extracts of Spatoglossum variabile, Stokeyia indica, and Melanothamnus afaqhusainii. The use of Padina pavonica benzene extract caused more mortality of Dsydercus cingulatus an economically important cotton pest (Sahayaraj and Kalidas 2011). In banana and tomato plants the root-knot infection caused by a nematode, (Meloidogyne spp.) was controlled by soil application of SEs such as Ulva lactuca, Ulva fasciata, and Stokeyia indica (El-Ansary and Hamouda 2014; Ghareeb et al. 2019).
Plants activate their defense mechanism against pathogens by recognizing pathogen-associated molecular patterns (PAMP) (Göhre and Robatzek 2008). Seaweed-based elicitors act as PAMPs and the signaling pathways are activated by the phytohormones jasmonic acid (JA), ethylene (ET), and salicylic acid (SA) (Dumas et al. 2010). Elicitors or plant defense activators generally induce partial, broad-range systemic resistance, as indicated by a considerable reduction in disease symptoms caused by various pathogens. The bio-elicitors stimulate the plant’s defense mechanisms, influencing genetic reprogramming (Pršić and Ongena 2020). Seaweeds contain bioactive compounds such as carrageenans, fucans, laminarans and ulvans that have been used as elicitors to protect the plant against various diseases (Shukla et al. 2016). Carrageenans possess antiviral properties and elicit defensive responses in plants. For example, tomato plants treated with carrageenan significantly suppressed tomato chlorotic dwarf viroid replication and expression. Additionally, jasmonic acid-related genes AOS (allene oxide synthase) and LOX (lipoxygenase) were up-regulated during viroid infection in plants treated with carrageenan (Sangha et al. 2011, 2015). The к-carrageenan extracted from Hypnea musciformis enhanced the immunity in tobacco plants infected with Tobacco Mosaic Virus (TMV) and activated SA and JA/ET dependent pathways defense mechanisms (Ghannam et al. 2013). Priming plants with seaweed-based elicitors enhances the plant immunity against phytopathogens by inducing defense-related genes and enzymes. For example, the Arabidopsis thaliana seedling treated with A.nodosum seaweed extract boosted the plant's immunity against various bacterial pathogens and also activated the expression of defense-related genes such as WRKY30 (WRKY DNA binding protein), (cytochrome P450, family 71, subfamily A, polypeptide 12), and PR-1 (Pathogenesis-related protein-1) (Cook et al. 2018). Seaweed-based elicitors are environmentally friendly crop protection strategies available and are a developing paradigm in the prevention of plant diseases that activates the plant immune system prior to pathogen overcoming. It also activates the innate immunity of plants by upregulating the defense-related genes and hormones thus rescuing plants from biotic stress.
Mechanisms of action of seaweed extracts under biotic and abiotic stress conditions
The seaweed-based biostimulants obtained from complex extraction contain a wide range of bioactive compounds that in theory can generate multiple positive effects throughout plant development. This is because they trigger signaling pathways, resulting in physiological changes in plants. For example, overexpression of NtLTP4 (Nicotiana tabacum Lipid transfer protein 4) in Nicotiana tabacum increased tolerance to salt and drought stress. The NtLTP4 modulates transcription levels of salt-responsive genes NHX1 (Na+/H+antiporter) and HKT1 (high-affinity K+ transporter1) to reduce Na+ toxicity. (Xu et al. 2018). The tomato and pepper crops treated with seaweed extract were shown to increase the upregulation of gene transcripts Ga2Ox (Gibberellin 2-oxidase), IAA (Indole-3-acetic acid), and IPT (Isopentenyl transferase) which are involved in the production of phytohormones in plants (Ali et al. 2019).
The usage of seaweed extract modulates the level of ABA, commonly known as the plant’s stress hormone. Treatment of Arabidopsis with ethyl acetate extract of A.nodosum under salt stress increased the transcript accumulation of the SnRK2 (SNF1-related protein kinase 2) gene which activates the ABA signaling network (Jithesh et al. 2018). The bioactive components present in the ethyl acetate fraction of the A. nodosum extract enhance salt tolerance and regulate stress-related signal transduction. Shukla et al.(2018) reported that GmCYP707A1a and GmCYP707A3b genes which regulate ABA biosynthesis during dehydration and hydration cycles were overexpressed in soybean attributing to the application of seaweed extract and also induced drought resistance (Shukla et al. 2018). Similarly, the A. nodosum extract regulated GmFIB1a (Glycine max Fibrillin 1a) expression and protected photosystem ll. Proline induces osmotic stress-related genes in frost tolerance (Wei et al. 2022). The use of seaweed extract on A. thaliana accumulated proline by activation of proline biosynthesis and also mediated freezing tolerance (Nair et al. 2012). Similarly in another study, the use of S. angustifolium improved drought tolerance by enhancing the expression of P5CS (Pyrroline-5-carboxylate synthase) a crucial gene in the biosynthesis pathway of proline, and is overexpressed under stressful conditions (Shahriari et al. 2021). According to Ghamdi et al. (2018) the use of seaweed extract had a synergistic effect in reported Asparagus aethiopicus plants subjected to saline stress. There was a significant enhancement of morphological and physiological characteristics of the treated plant compared to the control. The experiment also revealed the salinity stress was mitigated by upregulation of genes ANN1 (Annexins D1), ANN2 (Annexins D2), PIP1 (plasma membrane intrinsic protein 1) and CHS (Chalcone Synthase) that are related to osmotic adjustment and flavonoid biosynthesis (Al-Ghamdi and Elansary 2018). Pre-treatment with seaweed extract induces partial stomatal closure which is linked to alteration in expression of genes associated with ABA-responsive and antioxidant system pathways. NCED3 (Nine-cis-epoxycarotenoid dioxygenase) genes implicated in the ABA-biosynthesis pathway was significantly expressed during dehydration and ABA-responsive gene RAB18 (responsive to ABA) and RD29A (Responsive to Desiccation) transcript were expressed in treated plants during drought stress (Santaniello et al. 2017). The bean (Phaseolus vulgaris) treated with A. nodosum was shown to increase drought tolerance by reducing ROS-induced MDA (Malondialdehyde) generation and increasing CAT (Catalase) activity (Eugenia Amaral Carvalho et al. 2018). The WRKY transcription factors are involved in the development and physiological processes of plants. They interact with MAPK (mitogen-activated protein kinase) and regulate plant function by acting downstream of multiple MAPKs. The application of K. alvarezii (K-sap) increased the expression of the TaMPK10 gene encoding MAP Kinase in wheat and increased the tolerance of plants under drought stress (Patel et al. 2018). Cold stress in Arabidopsis was mitigated by the use of seaweed extract which increased the chlorophyll content and lowered the expression of AtCLH1 and AtCLH2 (chlorophyll degradation genes) and also induced the expression of cold-responsive genes such as COR15A, RD29A, and CBF3 (Rayirath et al. 2009). In pepper plants, the salinity stress and oxidative damage were reduced by the application of seaweed extract which upregulated the antioxidant activity (SOD, CAT, and APX) (Elansary et al. 2017).
The application of seaweed extracts and their bioactive components activate the up-regulation of numerous defense-related genes, plant hormones, and defense enzymes and these factors interact and cross talk to build a complex network that aids in disease tolerance in plants. The seaweed polysaccharides induce oxidative burst and activate various signaling pathways such as SA, JA, and ET at a systemic level and this leads to activation of different defense-related players such as pathogenesis-related protein (PR) and enzymes like phenylalanine ammonia-lyase (PAL) and lipoxygenase (LOX) (Vera et al. 2011) (Fig. 3). For instance, the A. nodosum extract application enhanced resistance against Pseudomonas syringae pv. tomato DC3000. The extract activated the JA-dependent pathway by inducing the expression of PDF1.2 JA-related genes and the decrease in diseases symptoms on the leaves is linked to the enhanced expression of JA -related gene transcripts (Subramanian et al. 2011). The application of K-sap in tomato seedlings enhanced the concentration of hormones (ABA, IAA, SA, and zeatin) as well as pathogenesis-related genes PR-1b1, PR-3, and PR-5 during Macrophomina phaseolina infection (Agarwal et al. 2015). In pepper and tomato plants the foliar application of S. vulgare and Acanthophora spicifera reduced the severity of the pathogens Xanthomonas campestris pv. vesicatoria and Alternarai solani by activating genes (PR-1a, Pinll, and ETR-1) which are involved in defense signaling pathways (Ali et al. 2020). Cluzet et al.(2004) reported that the use of 500 µg mL−1 of Ulva spp extract in Medicago truncatula induced expression of genes related to defense signaling pathways, pathogenesis-related protein and cell wall protein (Cluzet et al. 2004). To reduce stress in plants seaweed extracts can be promising strategy and moreover the experiments on both biotic and abiotic stress have proven that seaweed based biostimulants are beneficial.

The schematic illustration highlights the plausible mechanism of action of seaweed extracts against fungal and bacterial disease control. Seaweed extract may directly affect pathogens or indirectly by eliciting plant defense machinery. Such elicitation occurs by activation of several enzymes (PAL, LOX and ACS) which leads to the activation of several metabolic pathways such as (SA, JA and ET) thus upregulating several defense genes which induce the biosynthesis of several secondary metabolites which are involved in diseases suppression. Note: ROS reactive oxygen species, SA salicylic acid, PAL phenylalanine ammonia lyase, LOX 5-lipoxygenase, AOS active oxygen species, JA jasmonic acid, ACC 1-aminocyclopropane-1-carboxylic acid, ACS-ACC synthase, ACO-ACC oxidase, ET ethylene, PR pathogenesis related protein, PPO polyphenol oxidase, PO prophenoloxidase, LTP lipid-transfer proteins, GLU glutamate, PDF1.2 Plant defensin1.2, PINII proteinase inhibitor II, CYP71A12 cytochrome P450, family 71, subfamily A, polypeptide 12, ETR1 Ethylene response 1
Conclusion
The utilization of seaweeds extracts and their bioactive compounds can influence the performance of crops. The advantages of seaweed products in agriculture are undeniable as they increase the uptake of nutrients, enhance both the morphological and physiological characteristics, and also mitigate stress in plants. It has been already reported that they can act as both biostimulants and bio-elicitors in plants. Moreover, the SEs are environment friendly and are also effective in inducing innate immunity in plants. SEs can be used in combination with other biofertilizers to gain maximum yield and reduce the use of chemical fertilizers. The chemical constituents of SEs must be studied and analysed to determine the mode of application, dosage, and duration of administration to different crops to obtain long-term benefits. To supply consistent and dependable products to end users, efficient production procedure based on an in-depth understanding of the molecular mechanisms by which these extracts elicit beneficial impacts in treated plants is essential. The composition of seaweed is complex and it is difficult to identify the bioactive molecule that induces specific effect in plants. The use of new approaches (high throughput sequencing, metabolomics and genomics) can unravel the mechanisms of action. The seaweed extracts have been utilized in research; they must be marketed in order to reach farmer’s level.
Availability of data and material
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
Code availability
Not applicable.
References
Abbas M, Anwar J, Zafar-Ul-Hye M et al (2020) Effect of seaweed extract on productivity and quality attributes of four onion cultivars. Horticulturae 6(2):28. https://doi.org/10.3390/HORTICULTURAE6020028
Abdel Latef AAH, Srivastava AK, Saber H et al (2017) Sargassum muticum and Jania rubens regulate amino acid metabolism to improve growth and alleviate salinity in chickpea. Sci Rep 7(1):12. https://doi.org/10.1038/s41598-017-07692-w
Abkhoo J, Sabbagh SK (2015) Control of Phytophthora melonis damping-off, induction of defense responses, and gene expression of cucumber treated with commercial extract from Ascophyllum nodosum. J Appl Phycol 28(2):1333–1342. https://doi.org/10.1007/S10811-015-0693-3
Agarwal PK, Shukla PS, Gupta K, Jha B (2012) Bioengineering for salinity tolerance in plants: state of the art. Mol Biotechnol 54(1):102–123. https://doi.org/10.1007/S12033-012-9538-3
Agarwal P, Patel K, Das AK et al (2015) Insights into the role of seaweed Kappaphycus alvarezii sap towards phytohormone signalling and regulating defence responsive genes in Lycopersicon esculentum. J Appl Phycol 28(4):2529–2537. https://doi.org/10.1007/S10811-015-0784-1
Ahmed DAEA, Gheda SF, Ismail GA (2020) Efficacy of two seaweeds dry mass in bioremediation of heavy metal polluted soil and growth of radish (Raphanus sativus L.) plant. Environ Sci Pollut Res 28(10):12831–12846. https://doi.org/10.1007/S11356-020-11289-8
Al-Ghamdi AA, Elansary HO (2018) Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on Asparagus phenolics and stress related genes under saline irrigation. Plant Physiol Biochem 129:273–284. https://doi.org/10.1016/J.PLAPHY.2018.06.008
Ali O, Ramsubhag A, Jayaraman J (2019) Biostimulatory activities of Ascophyllum nodosum extract in tomato and sweet pepper crops in a tropical environment. PLoS ONE 14:e0216710. https://doi.org/10.1371/JOURNAL.PONE.0216710
Ali O, Ramsubhag A, Jayaraman J (2020) Phytoelicitor activity of Sargassum vulgare and Acanthophora spicifera extracts and their prospects for use in vegetable crops for sustainable crop production. J Appl Phycol 33(1):639–651. https://doi.org/10.1007/S10811-020-02309-8
Ali O, Ramsubhag A, Jayaraman J (2022) Transcriptome-wide modulation by Sargassum vulgare and Acanthophora spicifera extracts results in a prime-triggered plant signalling cascade in tomato and sweet pepper. AoB PLANTS. https://doi.org/10.1093/AOBPLA/PLAC046
Ammar N, Jabnoun-Khiareddine H, Mejdoub-Trabelsi B et al (2017) Pythium leak control in potato using aqueous and organic extracts from the brown alga Sargassum vulgare (C. Agardh, 1820). Postharvest Biol Technol 130:81–93. https://doi.org/10.1016/J.POSTHARVBIO.2017.04.010
Anderson PK, Cunningham AA, Patel NG, Morales FJ, Epstein PR, Daszak P (2004) Emerging infectious diseases of plants: pathogen pollution, climate change and agrotechnology drivers. Trends Ecol Evol 19(10):535–544
Aranega-Bou P, Lde la Leyva O, MI Finiti et al (2014) Priming of plant resistance by natural compounds Hexanoic acid as a model. Front Plant Sci 5:488. https://doi.org/10.3389/FPLS.2014.00488
Aremu AO, Masondo NA, Rengasamy KRR et al (2015) Physiological role of phenolic biostimulants isolated from brown seaweed Ecklonia maxima on plant growth and development. Planta 241:1313–1324. https://doi.org/10.1007/s00425-015-2256x
Arioli T, Mattner SW, Hepworth G et al (2021) Effect of seaweed extract application on wine grape yield in Australia. J Appl Phycol 33:1883–1891. https://doi.org/10.1007/S10811-021-02423-1
Ashour M, Hassan SM, Elshobary ME et al (2021) Impact of commercial seaweed liquid extract (TAM®) biostimulant and its bioactive molecules on growth and antioxidant activities of hot pepper (Capsicum annuum). Plants 10:1045. https://doi.org/10.3390/PLANTS10061045
Bahmani Jafarlou M, Pilehvar B, Modaresi M, Mohammadi M (2022) Seaweed liquid extract as an alternative biostimulant for the amelioration of salt-stress effects in Calotropis procera (Aiton) W.T. J Plant Growth Regul 2022:1–16. https://doi.org/10.1007/S00344-021-10566-1
Bajpai S, Shukla PS, Asiedu S et al (2019) A Biostimulant preparation of brown seaweed Ascophyllum nodosum suppresses powdery mildew of strawberry. Plant Pathol J 35:406. https://doi.org/10.5423/PPJ.OA.03.2019.0066
Baloch GN, Tariq S, Ehteshamul-Haque S et al (2013) Management of root diseases of eggplant and watermelon with the application of asafoetida and seaweeds. Researchgatenet 86:138–142. https://doi.org/10.5073/JABFQ.2013.086.019
Bencze S, Veisz O, Bedo Z (2004) Effects of high atmospheric CO2 and heat stress on phytomass, yield and grain quality of winter wheat. Cereal Res Commun 32:75–82. https://doi.org/10.1007/BF03543283
Bhanuprakash K, Yogeesha HS (2016) Seed priming for abiotic stress tolerance: an overview. Abiotic Stress Physiol Horticult Crops 103:117. https://doi.org/10.1007/978-81-322-2725-0_6
Bonomelli C, Celis V, Lombardi G, Mártiz J (2018) Salt stress effects on avocado (Persea americana Mill.) plants with and without seaweed extract (Ascophyllum nodosum) application. Agronomy 8:64. https://doi.org/10.3390/AGRONOMY8050064
Bradáčová K, Weber NF, Morad-Talab N, Asim M, Imran M, Weinmann M, Neumann G (2016) Micronutrients (Zn/Mn), seaweed extracts, and plant growth-promoting bacteria as cold-stress protectants in maize. Chem Biol Technol Agricult 3(1):1–10. https://doi.org/10.1186/S40538-016-0069-1
Briceño-Domínguez D, Hernández-Carmona G, Moyo M et al (2014) Plant growth promoting activity of seaweed liquid extracts produced from Macrocystis pyrifera under different pH and temperature conditions. J Appl Phycol 26:2203–2210. https://doi.org/10.1007/s10811-014-0237-2
Caijiao C, Leshan H, Mengke Y et al (2021) Comparative studies on antioxidant, angiotensin-converting enzyme inhibitory and anticoagulant activities of the methanol extracts from two brown algae (Sargassum horneri and Sargassum thunbergii). Russ J Mar Biol 47(5):380–387. https://doi.org/10.1134/S1063074021050035
Campobenedetto C, Agliassa C, Mannino G et al (2021) A biostimulant based on seaweed (Ascophyllum nodosum and Laminaria digitata) and yeast extracts mitigates water stress effects on tomato (Solanum lycopersicum L.). Agriculture 11:557. https://doi.org/10.3390/AGRICULTURE11060557
Caradonia F, Battaglia V, Righi L et al (2018) Plant Biostimulant regulatory framework: prospects in Europe and current situation at international level. J Plant Growth Regul 38(2):438–448. https://doi.org/10.1007/S00344-018-9853-4
Carmody N, Goñi O, Łangowski Ł, O’Connell S (2020) Ascophyllum nodosum extract biostimulant processing and its impact on enhancing heat stress tolerance during tomato fruit set. Front Plant Sci 11:807. https://doi.org/10.3389/FPLS.2020.00807
Carvalho MEA, de Camargo PR, Gaziola SA, Azevedo RA (2018) Is seaweed extract an elicitor compound? Changing proline content in drought-stressed bean plants. Comunicata Scientiae 9(2):292–297
Chen D, Huang Y, Shen D, Zhou W, Ao J, Jiang Y, Fahd R (2019) Effects of seaweed extracts on promoting growth and improving stress resistance in sugarcane. Asian Agric Res 11(1812-2019–3316):69–76
Chernane H, Latique S, Mansori M, El Kaoua M (2015) Salt stress tolerance and antioxidative mechanisms in wheat plants (Triticum durum L.) by seaweed extracts application. J Agric Vet Sci 8(3):36–44
Cluzet S, Torregrosa C, Jacquet C et al (2004) Gene expression profiling and protection of Medicago truncatula against a fungal infection in response to an elicitor from green algae Ulva spp. Plant Cell Environ 27:917–928. https://doi.org/10.1111/J.1365-3040.2004.01197.x
Colla G, Rouphael Y (2015) Biostimulants in horticulture. Sci Hortic 196:1–2. https://doi.org/10.1016/J.SCIENTA.2015.10.044
Cook J, Zhang J, Norrie J et al (2018) Seaweed extract (Stella Maris®) activates innate immune responses in arabidopsis thaliana and protects host against bacterial pathogens. Mar Drugs 16:2345. https://doi.org/10.3390/MD16070221
Craigie JS (2010) Seaweed extract stimuli in plant science and agriculture. J Appl Phycol 23(3):371–393. https://doi.org/10.1007/S10811-010-9560-4
de Borba MC, de Freitas MB, Stadnik MJ (2019) Ulvan enhances seedling emergence and reduces Fusarium wilt severity in common bean (Phaseolus vulgaris L.). Crop Prot 118:66–71. https://doi.org/10.1016/J.CROPRO.2018.12.014
De Corato U, Salimbeni R, De Pretis A et al (2017) Antifungal activity of crude extracts from brown and red seaweeds by a supercritical carbon dioxide technique against fruit postharvest fungal diseases. Postharvest Biol Technol 131:16–30. https://doi.org/10.1016/J.POSTHARVBIO.2017.04.011
de Sousa AM, Ayub RA, Viencz T, Botelho RV (2019) Fruit set and yield of apple trees cv. Gala treated with seaweed extract of ascophyllum nodosum and thidiazuron. Revista
Decker D, Kleczkowski LA (2019) UDP-sugar producing pyrophosphorylases: distinct and essential enzymes with overlapping substrate specificities, providing de novo precursors for glycosylation reactions. Front Plant Sci 9:1822
Deolu-Ajayi AO, van der Meer IM, van der Werf A, Karlova R (2022) The power of seaweeds as plant biostimulants to boost crop production under abiotic stress. Plant Cell Environ 45:2537–2553. https://doi.org/10.1111/PCE.14391
dos Santos PLF, Zabotto AR, Jordão HWC et al (2019) Use of seaweed-based biostimulant (Ascophyllum nodosum) on ornamental sunflower seed germination and seedling growth. Ornam Hortic 25:231–237. https://doi.org/10.1590/2447-536X.V25I3.2044
Di Filippo-Herrera DA, Muñoz-Ochoa M, Hernández-Herrera RM, Hernández-Carmona G (2018) Biostimulant activity of individual and blended seaweed extracts on the germination and growth of the mung bean. J Appl Phycol 31(3):2025–2037. https://doi.org/10.1007/S10811-018-1680-2
Di Mola I, Cozzolino E, Ottaiano L, Giordano M, Rouphael Y, Colla G, Mori M (2019) Effect of vegetal-and seaweed extract-based biostimulants on agronomical and leaf quality traits of plastic tunnel-grown baby lettuce under four regimes of nitrogen fertilization. Agronomy 9(10):571
do Rosário Rosa V, Farias dos Santos AL, Alves da Silva A et al (2021) Increased soybean tolerance to water deficiency through biostimulant based on fulvic acids and Ascophyllum nodosum (L.) seaweed extract. Plant Physiol Biochem 158:228–243. https://doi.org/10.1016/J.PLAPHY.2020.11.008
Dookie M, Ali O, Ramsubhag A, Jayaraman J (2021) Flowering gene regulation in tomato plants treated with brown seaweed extracts. Sci Hortic 276:109715. https://doi.org/10.1016/J.SCIENTA.2020.109715
Dumas B, Jaulneau V, Lafitte C et al (2010) Ulvan, a sulfated polysaccharide from green algae, activates plant immunity through the jasmonic acid signaling pathway. J Biomed Biotechnol. https://doi.org/10.1155/2010/525291
Dziugieł T, Wadas W (2020) Possibility of increasing early crop potato yield with foliar application ofseaweed extracts and humic acids. J Central Eur Agric 21:300–310. https://doi.org/10.5513/JCEA01/21.2.2576
El-Ansary MSM, Hamouda RA (2014) Biocontrol of root-knot nematode infected banana plants by some marine algae. Russ J Mar Biol 40(2):140–146. https://doi.org/10.1134/S1063074014020047
El-Katony TM, Deyab MA, El-Adl MF et al (2021) Extracts of the brown alga Dictyota dichotoma (Hudson) J. V. Lamouroux alleviate salt stress in rice (Oryza sativa L.) during germination. J Plant Growth Regul 40:986–999. https://doi.org/10.1007/s00344-020-10156-7
El M, El BM, Barakate M et al (2021) Ulva lactuca extract and fractions as seed priming agents mitigate salinity stress in tomato seedlings. Plants 10:1104. https://doi.org/10.3390/PLANTS10061104
El Modafar C, Elgadda M, El Boutachfaiti R et al (2012) Induction of natural defence accompanied by salicylic acid-dependant systemic acquired resistance in tomato seedlings in response to bioelicitors isolated from green algae. Sci Hortic 138:55–63. https://doi.org/10.1016/J.SCIENTA.2012.02.011
Elansary HO, Yessoufou K, Abdel-Hamid AME et al (2017) Seaweed extracts enhance salam turfgrass performance during prolonged irrigation intervals and saline shock. Front Plant Sci 8:830. https://doi.org/10.3389/FPLS.2017.00830
Ertani A, Francioso O, Tinti A et al (2018) Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. As biostimulants in Zea mays L. using a combination of chemical, biochemical and morphological approaches. Front Plant Sci 9:428. https://doi.org/10.3389/FPLS.2018.00428
Esserti S, Smaili A, Rifai LA et al (2016) Protective effect of three brown seaweed extracts against fungal and bacterial diseases of tomato. J Appl Phycol 29(2):1081–1093. https://doi.org/10.1007/S10811-016-0996-Z
Fan D, Hodges DM, Critchley AT, Prithiviraj B (2013) A commercial extract of brown Macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun Soil Sci Plant Anal 44:1873–1884. https://doi.org/10.1080/00103624.2013.790404
Finnie JF, Van Staden J (1985) Effect of seaweed concentrate and applied hormones on in vitro cultured tomato roots. J Plant Physiol 120(3):215–222. https://doi.org/10.1016/S0176-1617(85)80108-5
Flora G, Rani SMV (2012) An approach towards control of blast by foliar application of seaweed concentrate. Sci Res Report 2(3):213–217
Flores P, Pedreño MA, Almagro L et al (2021) Increasing nutritional value of broccoli with seaweed extract and trilinolein. J Food Compos Anal 98:103834. https://doi.org/10.1016/J.JFCA.2021.103834
Fornes F, Sanchez-Perales M, Guardiola JL (2002) Effect of a seaweed extract on the productivity of'de Nules' clementine mandarin and navelina orange
Ghannam A, Abbas A, Alek H et al (2013) Enhancement of local plant immunity against tobacco mosaic virus infection after treatment with sulphated-carrageenan from red alga. (hypnea Musciformis). https://doi.org/10.1016/j.pmpp.2013.07.001
Ghareeb RY, Adss IA, Bayoumi SR, El-Habashy DE (2019) The nematicidal potentiality of some algal extracts and their role in enhancement the tomato defense genes against root knot-nematodes. Egypt J BiolPest Control 29:1–10. https://doi.org/10.1186/S41938-019-0153-5
Göhre V, Robatzek S (2008) Breaking the barriers: microbial effector molecules subvert plant immunity. Annu Rev Phytopathol 46:189–215. https://doi.org/10.1146/annurev.phyto46120407110050
Gomathi R, Kohila S, Ramachandiran K (2017) Evaluating the effect of seaweed formulations on the quality and yield of sugarcane. Madras Agric J 104(march 1–3):1
Goñi O, Łangowski Ł, Feeney E et al (2021) Reducing nitrogen input in barley crops while maintaining yields using an engineered biostimulant derived from ascophyllum nodosum to enhance nitrogen use efficiency. Front Plant Sci. https://doi.org/10.3389/FPLS.2021.664682
González-González MF, Ocampo-Alvarez H, Santacruz-Ruvalcaba F et al (2020) Physiological, ecological, and biochemical implications in tomato plants of two plant biostimulants: Arbuscular Mycorrhizal Fungi and seaweed extract. Front Plant Sci 11:999. https://doi.org/10.3389/FPLS.2020.00999
Górka B, Wieczorek PP (2017) Simultaneous determination of nine phytohormones in seaweed and algae extracts by HPLC-PDA. J Chromatogr B 1057:32–39. https://doi.org/10.1016/J.JCHROMB.2017.04.048
Gruszka D, Janeczko A, Dziurka M et al (2018) Non-enzymatic antioxidant accumulations in BR-deficient and BR-insensitive barley mutants under control and drought conditions. Physiol Plant 163:155–169. https://doi.org/10.1111/PPL.12674
Gupta S, Stirk WA, Plačková L et al (2021) Interactive effects of plant growth-promoting rhizobacteria and a seaweed extract on the growth and physiology of Allium cepa L. (onion). J Plant Physiol 262:153437. https://doi.org/10.1016/J.JPLPH.2021.153437
Hamouda MM, Khalil S-AM, Gad D (2022) Potential of seaweed extract on growth, physiological, cytological and biochemical parameters of wheat (Triticum aestivum L.) seedlings. J Soil Sci Plant Nutr 1:1–14. https://doi.org/10.1007/S42729-022-00774-3
Hamouda RA, Hussein MH, El-Naggar NEA et al (2022) Promoting effect of soluble polysaccharides extracted from Ulva spp. on Zea mays L. growth. Molecules 27:1394. https://doi.org/10.3390/MOLECULES27041394
Hankins SD, Hockey HP (1990) The effect of a liquid seaweed extract from (Fucales, Phaeophyta) on the two-spotted red spider mite. In: Thirteenth international seaweed symposium, pp 555–559. https://doi.org/10.1007/978-94-009-2049-1_80
Hassan SM, Ashour M, Sakai N et al (2021) Impact of seaweed liquid extract biostimulant on growth, yield, and chemical composition of cucumber (Cucumis sativus). Agriculture 11:320. https://doi.org/10.3390/AGRICULTURE11040320
Hernández-Herrera RM, Santacruz-Ruvalcaba F, Ruiz-López MA et al (2014) Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J Appl Phycol 26:619–628. https://doi.org/10.1007/s10811-013-0078-4
Hernández-Herrera RM, Sánchez-Hernández CV, Palmeros-Suárez PA et al (2022) Seaweed extract improves growth and productivity of tomato plants under salinity stress. Agronomy 12:2495. https://doi.org/10.3390/AGRONOMY12102495/S1
Huang H, Ullah F, Zhou DX et al (2019) Mechanisms of ROS regulation of plant development and stress responses. Front Plant Sci 10:800. https://doi.org/10.3389/FPLS.2019.00800
Hussein MH, Eltanahy E, Al Bakry AF et al (2021) Seaweed extracts as prospective plant growth bio-stimulant and salinity stress alleviator for Vigna sinensis and Zea mays. J Appl Phycol 33(2):1273–1291. https://doi.org/10.1007/S10811-020-02330-x
Islam MT, Gan HM, Ziemann M et al (2020) Phaeophyceaean (Brown Algal) extracts activate plant defense systems in arabidopsis thaliana challenged with Phytophthora cinnamomi. Front Plant Sci 11:852. https://doi.org/10.3389/FPLS.2020.00852
Jannin L, Arkoun M, Etienne P et al (2012) Brassica napus growth is promoted by Ascophyllum nodosum (L.) Le Jol. seaweed extract: microarray analysis and physiological characterization of N, C, and S metabolisms. J Plant Growth Regul 32(1):31–52. https://doi.org/10.1007/S00344-012-9273-9
Jaulneau V, Lafitte C, Corio-Costet MF et al (2011) An Ulva armoricana extract protects plants against three powdery mildew pathogens. Eur J Plant Pathol 131:393–401. https://doi.org/10.1007/s10658-011-9816-0
Jayaraj J, Wan A, Rahman M, Punja ZK (2008) Seaweed extract reduces foliar fungal diseases on carrot. Crop Prot 27:1360–1366. https://doi.org/10.1016/J.CROPRO.2008.05.005
Jiménez-Escrig A, Cambrodón IG (1999) Nutritional evaluation and physiological effects of edible seaweeds. Arch Latinoam Nutr 49:114–120
Jithesh MN, Shukla PS, Kant P et al (2018) Physiological and transcriptomics analyses reveal that Ascophyllum nodosum extracts induce salinity tolerance in Arabidopsis by regulating the expression of stress responsive genes. J Plant Growth Regul 38(2):463–478. https://doi.org/10.1007/S00344-018-9861-4
Karthik T, Sarkar G, Babu S et al (2020) Preparation and evaluation of liquid fertilizer from Turbinaria ornata and Ulva reticulata. Biocatal Agric Biotechnol 28:101712. https://doi.org/10.1016/J.BCAB.2020.101712
Kauffman GL, Kneivel DP, Watschke TL (2007) Effects of a biostimulant on the heat tolerance associated with photosynthetic capacity, membrane thermostability, and polyphenol production of perennial ryegrass. Crop Sci 47:261–267. https://doi.org/10.2135/CROPSCI2006.03.0171
Kim SK (ed) (2011) Handbook of marine macroalgae: biotechnology and applied phycology. Wiley, New York
Khan W, Rayirath UP, Subramanian S et al (2009) Seaweed extracts as biostimulants of plant growth and development. J Plant Growth Regul 28(4):386–399. https://doi.org/10.1007/S00344-009-9103-x
Kocira S, Szparaga A, Findura P, Treder K (2020) Modification of yield and fiber fractions biosynthesis in Phaseolus vulgaris L. by treatment with biostimulants containing amino acids and seaweed extract. Agronomy 10:1338. https://doi.org/10.3390/AGRONOMY10091338
Kumar NA, Vanlalzarzova B, Sridhar S, Baluswami M (2012) Effect of liquid seaweed fertilizer of Sargassum wightii Grev on the growth and biochemical content of green gram (Vigna radiata (L.) R. Wilczek). Recent Res Sci Technol 4(4):40–45
Kumar R, Trivedi K, Anand KGV et al (2020) Science behind biostimulant action of seaweed extract on growth and crop yield: insights into transcriptional changes in roots of maize treated with Kappaphycus alvarezii seaweed extract under soil moisture stressed conditions. J Appl Phycol 32:599–613. https://doi.org/10.1007/s10811-019-01938-y
Kumari J, Haque MI, Jha RK, Rathore MS (2022) The red seaweed Kappaphycus alvarezii antiporter gene (KaNa+/H+) confers abiotic stress tolerance in transgenic tobacco. Mol Biol Rep 2022:1–15. https://doi.org/10.1007/S11033-022-07213-7
Latef AAHA, Zaid A, Alwaleed EA (2021) Influences of priming on selected physiological attributes and protein pattern responses of salinized wheat with extracts of Hormophysa cuneiformis and Actinotrichia fragilis. Agronomy 11:545. https://doi.org/10.3390/AGRONOMY11030545
Liu H, Chen X, Song L et al (2019) Polysaccharides from Grateloupia filicina enhance tolerance of rice seeds (Oryza sativa L.) under salt stress. Int J Biol Macromol 124:1197–1204. https://doi.org/10.1016/J.IJBIOMAC.2018.11.270
Lola-Luz T, Hennequart F, Gaffney M (2013) Enhancement of phenolic and flavonoid compounds in cabbage (Brassica oleraceae) following application of commercial seaweed extracts of the brown seaweed, (Ascophyllum nodosum). Agric Food Sci 22:288–295. https://doi.org/10.23986/AFSCI.7676
Machado LP, Matsumoto ST, Jamal CM et al (2014) Chemical analysis and toxicity of seaweed extracts with inhibitory activity against tropical fruit anthracnose fungi. J Sci Food Agric 94:1739–1744. https://doi.org/10.1002/JSFA.6483
Mattner SW, Villalta ON, Wite D et al (2014) In vitro suppression of Sclerotinia minor by a seaweed extract from Durvillaea potatorum and Ascophyllum nodosum. Aust Plant Dis Notes 9:1–5. https://doi.org/10.1007/S13314-014-0137-y
Mattner SW, Milinkovic M, Arioli T (2018) Increased growth response of strawberry roots to a commercial extract from Durvillaea potatorum and Ascophyllum nodosum. J Appl Phycol 30:2943–2951. https://doi.org/10.1007/S10811-017-1387-9
Michalak I, Górka B, Wieczorek PP et al (2016) Supercritical fluid extraction of algae enhances levels of biologically active compounds promoting. Plant Growth. https://doi.org/10.1080/09670262.2015.1134813
Miedes E, Vanholme R, Boerjan W, Molina A (2014) The role of the secondary cell wall in plant resistance to pathogens. Front Plant Sci 5:358. https://doi.org/10.3389/FPLS.2014.00358
Muniswami DM, Buvaneshwari K, Fathima Rosa Mystica L et al (2021) Comparative assessment of different biofertilizers in maize (Zea mays L.) cultivation. Biomass Conv Bioref. https://doi.org/10.1007/s13399-021-01543-5
Muthu-Pandian Chanthini K, Senthil-Nathan S, Stanley-Raja V et al (2019) Chaetomorpha antennina (Bory) Kützing derived seaweed liquid fertilizers as prospective bio-stimulant for Lycopersicon esculentum (Mill). Biocatal Agric Biotechnol 20:101190. https://doi.org/10.1016/J.BCAB.2019.101190
Mzibra A, Aasfar A, Khouloud M et al (2021) Improving growth, yield, and quality of tomato plants (Solanum lycopersicum L) by the application of moroccan seaweed-based biostimulants under greenhouse conditions. Agronomy 11:1373. https://doi.org/10.3390/AGRONOMY11071373
Nair P, Kandasamy S, Zhang J et al (2012) Transcriptional and metabolomic analysis of Ascophyllum nodosum mediated freezing tolerance in Arabidopsis thaliana. BMC Genom 13:1–23. https://doi.org/10.1186/1471-2164-13-643
Nephali L, Piater LA, Dubery IA et al (2020) Biostimulants for plant growth and mitigation of abiotic stresses: a metabolomics perspective. Metabolites 10:1–26. https://doi.org/10.3390/METABO10120505
Neto APDA, Oliveira GRF, da Mello SC et al (2020) Seed priming with seaweed extract mitigate heat stress in spinach: effect on germination, seedling growth and antioxidant capacity. Bragantia 79:502–511. https://doi.org/10.1590/1678-4499.20200127
Ngoroyemoto N, Kulkarni MG, Stirk WA, Gupta S, Finnie JF, van Staden J (2020) Interactions between microorganisms and a seaweed-derived biostimulant on the growth and biochemical composition of Amaranthus hybridus L. Nat Product Commun 15(7):1934578
Noli ZA, Suwirmen A, Aliyyanti P (2021) Effect of liquid seaweed extracts as biostimulant on vegetative growth of soybean. IOP Conf Ser Earth Environ Sci 759:012029. https://doi.org/10.1088/1755-1315/759/1/012029
Ozbay N, Demirkiran AR (2019) Enhancement of growth in ornamental pepper (Capsicum annuum L.) plants with application of a commercial seaweed product, stimplex®. Appl Ecol Environ Res 17:4361–4375
Patel K, Agarwal P, Agarwal PK (2018) Kappaphycus alvarezii sap mitigates abiotic-induced stress in Triticum durum by modulating metabolic coordination and improves growth and yield. J Appl Phycol 30:2659–2673. https://doi.org/10.1007/s10811-018-1423-4
Parmar N, Singh KH, Sharma D, Singh L, Kumar P, Nanjundan J, Khan YJ, Chauhan DK, Thakur AK (2017) Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: a comprehensive review. 3 Biotech 7(4):239. https://doi.org/10.1007/s13205-017-0870-y
Pereira L, Correia F (2015) Introdução. Macroalgas Marinhas da Costa Portuguesa: Biodiversidade, ecologia e utilizações 342
Prakash P, Mitra A, Nag R, Sunkar S (2018) Effect of seaweed liquid fertilizer and humic acid formulation on the growth and nutritional quality of Abelmoschus esculentus. Asian J Crop Sci 10:48–52. https://doi.org/10.3923/AJCS.2018.48.52
Pramanick B, Brahmachari K, Mahapatra BS et al (2017) Growth, yield and quality improvement of potato tubers through the application of seaweed sap derived from the marine alga Kappaphycus alvarezii. J Appl Phycol 29(6):3253–3260. https://doi.org/10.1007/S10811-017-1189-0
Pršić J, Ongena M (2020) Elicitors of plant immunity triggered by beneficial bacteria. Front Plant Sci 11:1675. https://doi.org/10.3389/FPLS.2020.594530
Rajendran R, Jagmohan S, Jayaraj P et al (2021) Effects of Ascophyllum nodosum extract on sweet pepper plants as an organic biostimulant in grow box home garden conditions. J Appl Phycol 34(1):647–657. https://doi.org/10.1007/S10811-021-02611-Z
Ramkissoon A, Ramsubhag A, Jayaraman J (2017) Phytoelicitor activity of three Caribbean seaweed species on suppression of pathogenic infections in tomato plants. J Appl Phycol 29(6):3235–3244. https://doi.org/10.1007/S10811-017-11600
Rayirath P, Benkel B, Mark Hodges D, et al (2009) Lipophilic components of the brown seaweed, Ascophyllum nodosum, enhance freezing tolerance in Arabidopsis thaliana
Rayorath P, Jithesh MN, Farid A et al (2008) Rapid bioassays to evaluate the plant growth promoting activity of Ascophyllum nodosum (L.) Le Jol. using a model plant, Arabidopsis thaliana (L.) Heynh. J Appl Phycol 20:423–429. https://doi.org/10.1007/s10811-007-9280-6
Rengasamy KRR, Kulkarni MG, Stirk WA, Van Staden J (2015) Eckol—a new plant growth stimulant from the brown seaweed Ecklonia maxima. J Appl Phycol 27:581–587. https://doi.org/10.1007/S10811-014-0337-Z
Rengasamy KRR, Kulkarni MG, Pendota SC, Van Staden J (2016) Enhancing growth, phytochemical constituents and aphid resistance capacity in cabbage with foliar application of eckol—a biologically active phenolic molecule from brown seaweed. New Biotechnol 33:273–279. https://doi.org/10.1016/J.NBT.2015.11.002
Rouphael Y, De Micco V, Arena C et al (2016) Effect of Ecklonia maxima seaweed extract on yield, mineral composition, gas exchange, and leaf anatomy of zucchini squash grown under saline conditions. J Appl Phycol 29(1):459–470. https://doi.org/10.1007/S10811-016-0937-x
Sahana BN, PrasannaKumar MK, Mahesh HB et al (2021) Biostimulants derived from red seaweed stimulate the plant defence mechanism in rice against Magnaporthe oryzae. J Appl Phycol 34(1):659–665. https://doi.org/10.1007/S10811-021-02627-5
Sahayaraj K, Kalidas S (2011) Evaluation of nymphicidal and ovicidal effect of a seaweed, Padina pavonica (Linn.)(Phaeophyceae) on cotton pest, Dysdercus cingulatus (Fab.)
Sahayaraj K, Mary Jeeva Y (2012) Nymphicidal and ovipositional efficacy of seaweed Sargassum tenerrimum (J. Agardh) against Dysdercus cingulatus (Fab.) (Pyrrhocoridae). Chilean J Agric Res 72:152–156. https://doi.org/10.4067/S0718-58392012000100024
Sangha JS, Kandasamy S, Khan W et al (2015) λ-carrageenan suppresses tomato chlorotic dwarf viroid (TCDVd) replication and symptom expression in tomatoes. Mar Drugs 13:2875–2889. https://doi.org/10.3390/MD13052875
Sangha JS, Khan W, Ji X et al (2011) Carrageenans, sulphated polysaccharides of red seaweeds, differentially affect Arabidopsis thaliana Resistance to Trichoplusia ni (Cabbage Looper). PLoS ONE 6:e26834. https://doi.org/10.1371/JOURNAL.PONE.0026834
Santaniello A, Scartazza A, Gresta F et al (2017) Ascophyllum nodosum seaweed extract alleviates drought stress in Arabidopsis by affecting photosynthetic performance and related gene expression. Front Plant Sci 8:1362. https://doi.org/10.3389/FPLS.2017.01362
Sbaihat L, Takeyama K, Koga T et al (2015) Induced resistance in solanum lycopersicum by algal elicitor extracted from sargassum fusiforme. Sci World J. https://doi.org/10.1155/2015/870520
Shah MT, Zodape ST, Chaudhary DR, Eswaran K, Chikara J (2013) Seaweed sap as an alternative liquid fertilizer for yield and quality improvement of wheat. J Plant Nutr 36(2):192–200
Shahriari AG, Mohkami A, Niazi A et al (2021) Application of brown algae (Sargassum angustifolium) Extract for improvement of drought tolerance in Canola (Brassica napus L.). Iran J Biotechnol 19:e2775. https://doi.org/10.30498/IJB.2021.2775
Shamya Arokia Rajan M, Thriunavukkarasu R, Joseph J, Aruni W (2020) Effect of seaweed on seed germination and biochemical constituents of Capsicum annuum. Biocatal Agric Biotechnol 29:101761. https://doi.org/10.1016/J.BCAB.2020.101761
Sharma HSS, Fleming C, Selby C et al (2013) Plant biostimulants: a review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses. J Appl Phycol 26(1):465–490. https://doi.org/10.1007/S10811-013-0101-9
Sharma S, Chen C, Khatri K et al (2019) Gracilaria dura extract confers drought tolerance in wheat by modulating abscisic acid homeostasis. Plant Physiol Biochem 136:143–154. https://doi.org/10.1016/J.PLAPHY.2019.01.015
Shukla PS, Borza T, Critchley AT, Prithiviraj B (2016) Carrageenans from red seaweeds as promoters of growth and elicitors of defense response in plants. Front Mar Sci 3:81. https://doi.org/10.3389/FMARS.2016.00081
Shukla PS, Shotton K, Norman E et al (2018) Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB PLANTS. https://doi.org/10.1093/AOBPLA/PLX051
Shukla PS, Mantin EG, Adil M et al (2019) Ascophyllum nodosum-based biostimulants: sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front Plant Sci 10:655. https://doi.org/10.3389/FPLS.2019.00655
Singh A, Chaudhary B (2010) Preliminary phycochemical analysis and in vitro antibacterial screening of Pithophora oedogonia (Mont.) Wittrock: a freshwater green alga forming mats in the water bodies. J Algal Biomass Utiliz 1:33–41
Singh S, Singh MK, Pal SK et al (2015) Sustainable enhancement in yield and quality of rain-fed maize through Gracilaria edulis and Kappaphycus alvarezii seaweed sap. J Appl Phycol 28(3):2099–2112. https://doi.org/10.1007/S10811-015-0680-8
Soliman AS, Ahmed AY, Abdel-Ghafour SE, El-Sheekh MM, Sobhy HM (2018) Antifungal bio-efficacy of the red algae Gracilaria confervoides extracts against three pathogenic fungi of cucumber plant. Middle East J Appl Sci 8(3):727–735
Stamatiadis S, Evangelou E, Jamois F, Yvin JC (2021) Targeting Ascophyllum nodosum (L.) Le Jol. extract application at five growth stages of winter wheat. J Appl Phycol 33(3):1873–1882. https://doi.org/10.1007/S10811-021-02417-Z
Subramanian S, Sangha JS, Gray BA et al (2011) Extracts of the marine brown macroalga, Ascophyllum nodosum, induce jasmonic acid dependent systemic resistance in Arabidopsis thaliana against Pseudomonas syringae pv. tomato DC3000 and Sclerotinia sclerotiorum. Eur J Plant Pathol 131(2):237–248. https://doi.org/10.1007/S10658-011-9802-6
Sujata GV, Baliyan V et al (2022) Alleviating drought stress in Brassica juncea (L.) Czern & Coss. by foliar application of biostimulants—orthosilicic acid and seaweed extract. Appl Biochem Biotechnol 2022:1–29. https://doi.org/10.1007/S12010-022-04085-2
Tandon S, Dubey A (2015) Effects of Biozyme (Ascophyllum nodosum) biostimulant on growth and development of soybean [Glycine max (L. Merill]. Commun Soil Sci Plant Anal 46(7):845–858. https://doi.org/10.1080/00103624.2015.1011749
Trivedi K, Vijay Anand KG, Kubavat D et al (2017) Crop stage selection is vital to elicit optimal response of maize to seaweed bio-stimulant application. J Appl Phycol 29(4):2135–2144. https://doi.org/10.1007/S10811-017-1118-2
Trivedi K, Vijay Anand KG, Vaghela P, Ghosh A (2018) Differential growth, yield and biochemical responses of maize to the exogenous application of Kappaphycus alvarezii seaweed extract, at grain-filling stage under normal and drought conditions. Algal Res 35:236–244. https://doi.org/10.1016/J.ALGAL.2018.08.027
Uthirapandi V, Suriya S, Boomibalagan P, Eswaran S, Ramya SS, Vijayanand N, Kathiresan D (2018) Biofertilizing potential of seaweed liquid extracts of marine macro algae on growth and biochemical parameters of Ocimum sanctum. J Pharmacogn Phytochem 7(3):3528–3532
Vera J, Castro J, Gonzalez A, Moenne A (2011) Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar Drugs 9:2514–2525. https://doi.org/10.3390/MD9122514
Verma N, Sehrawat KD, Mundlia P et al (2021) Potential use of Ascophyllum nodosum as a biostimulant for improving the growth performance of Vigna aconitifolia (Jacq) Marechal. Plants (basel, Switzerland). https://doi.org/10.3390/PLANTS10112361
Vinoth S, Sundari GP et al (2017) Evaluation of seagrass liquid extract on salt stress alleviation in tomato plants. Asian J Plant Sci 16:172–183. https://doi.org/10.3923/AJPS.2017.172.183
Voko MP, Kulkarni MG, Ngoroyemoto N et al (2022) Vermicompost leachate, seaweed extract and smoke-water alleviate drought stress in cowpea by influencing phytochemicals, compatible solutes and photosynthetic pigments. Plant Growth Regul 97(2):327–342. https://doi.org/10.1007/S10725-022-00815-Y
Wei TL, Wang ZX, He YF et al (2022) Proline synthesis and catabolism-related genes synergistically regulate proline accumulation in response to abiotic stresses in grapevines. Sci Hortic 305:111373. https://doi.org/10.1016/J.SCIENTA.2022.111373
Wite D, Mattner SW, Porter IJ, Arioli T (2015) The suppressive effect of a commercial extract from Durvillaea potatorum and Ascophyllum nodosum on infection of broccoli by Plasmodiophora brassicae. J Appl Phycol 27:2157–2161. https://doi.org/10.1007/S10811-015-0564-y
Wu Y, Jenkins T, Blunden G et al (1998) Suppression of fecundity of the root-knot nematode, Meloidogyne javanica, in monoxenic cultures of Arabidopsis thaliana treated with an alkaline extract of Ascophyllum nodosum. J Appl Phycol 10(1):91–94. https://doi.org/10.1023/A:1008067420092
Xu C, Leskovar DI (2015) Effects of A. nodosum seaweed extracts on spinach growth, physiology and nutrition value under drought stress. Sci Hortic 183:39–47. https://doi.org/10.1016/J.SCIENTA.2014.12.004
Xu Y, Zheng X, Song Y et al (2018) NtLTP4, a lipid transfer protein that enhances salt and drought stresses tolerance in Nicotiana tabacum. Sci Rep 8(1):1–14. https://doi.org/10.1038/s41598-018-27274-8
Yadav DS, Rai R, Mishra AK et al (2019) ROS production and its detoxification in early and late sown cultivars of wheat under future O3 concentration. Sci Total Environ 659:200–210. https://doi.org/10.1016/J.SCITOTENV.2018.12.352
Yao Y, Wang X, Chen B, Zhang M, Ma J (2020) Seaweed extract improved yields, leaf photosynthesis, ripening time, and net returns of tomato (Solanum lycopersicum Mill.). ACS Omega 5(8):4242–4249
Yusuf R, Kristianse P, Warwick N (2019) Effect of two seaweed products and equivalent mineral treatments on lettuce (Lactuca sativa L.) growth. J Agron 18:100–106. https://doi.org/10.3923/JA.2019.100.106
Zou P, Lu X, Zhao H et al (2019) Polysaccharides derived from the brown algae Lessonia nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration. Front Plant Sci 10:48. https://doi.org/10.3389/FPLS.2019.00048
Acknowledgements
The authors are grateful to the management of the Vellore Institute of Technology for providing the necessary support to write this article. The authors acknowledge Dr. Sudhakaran R of Vellore Institute of Technology—aquaculture laboratory for his guidance in writing this review article.
Funding
The authors confirm that this review article was not written under the financial sponsorship of any funding agency.
Author information
Authors and Affiliations
Contributions
BR prepared and revised the manuscript under the direction of VR.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there were no competing interests either financial or non-financial.
Ethics approval
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Raja, B., Vidya, R. Application of seaweed extracts to mitigate biotic and abiotic stresses in plants. Physiol Mol Biol Plants 29, 641–661 (2023). https://doi.org/10.1007/s12298-023-01313-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12298-023-01313-9