
Abstract
On 10 October 2018, Hurricane Michael made landfall along the Florida panhandle as a category 5 storm, with maximum sustained winds of 259 km h−1 and storm surge of 4.3 m. To assess impacts on seagrass meadows and nekton communities, we sampled seagrass beds in three estuaries along a gradient of storm intensity: one proximate to the hurricane track (St. George Sound, FL), one that experienced storm surge without hurricane-force winds (Cedar Key, FL), and one outside the storm path (Chandeleur Islands, LA). The nekton community and seagrass characteristics were assessed prior to and shortly after the storm and again 1 year later. In St. George Sound, we observed post-storm changes in seagrass species composition and declines in seagrass cover, shoot count, and canopy height as well as a shift in associated fauna. Seagrass cover declined from 83 ± 9 to 50 ± 8% following the storm. Shoot height and shoot count were also reduced by over 50%. Nekton species most commonly found prior to the storm were Lagodon rhomboides (pinfish), Orthopristis chrysoptera (pigfish), and Bairdiella chrysoura (silver perch). Two weeks following the storm, the most commonly collected species shifted to pinfish, penaeid shrimp, Chilomycterus schoepfi (striped burrfish), and Eucinostomus argenteus (spotfin mojarra). Widespread declines in species-specific nekton abundances were also observed, with continued reductions in some species the following year. Overall, both the seagrass and nekton communities showed signs of recovery. Cedar Key and Chandeleur Islands both exhibited minimal changes in vegetation and nekton. Despite storm intensity and substantial physical damage to upland habitats, the estuarine seagrass communities were resilient to the effects of a powerful category 5 hurricane.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tropical cyclones, or hurricanes, are intense low-pressure systems that in the North Atlantic develop at latitudes of 5–15° (Poiner et al. 1989). These storms are associated with strong winds, storm surge, increased precipitation, and coastal sediment changes, all of which could singularly or in combination affect coastal estuaries. Additionally, intense storms can change water temperature, salinity, and increase pollution levels via runoff (Marchand et al. 2002), as well as cause damage to coastal vegetation, together influencing fish and invertebrate nekton communities (Rozas and Reed 1994; Tabb and Jones 1962).
Storm disturbance to seagrass meadows typically occurs through both direct and indirect physical effects (Birch and Birch 1984; Poiner et al. 1989; Thomas et al. 1961). Wind, waves, and storm surge can shift sediments and erode or bury seagrasses (Fourqurean and Rutten 2004). Indirect effects from stormwater discharge and runoff, such as altered salinity regimes and increased light attenuation from resuspended sediments and allochthonous inputs (e.g., terrestrial detritus and tannins), can persist for weeks to months following a storm (Walker et al. 2021). Storm effects can include seagrass loss, degradation (i.e., reduced shoot density, canopy height, or aboveground biomass), and/or fragmentation, leaving substantive changes to the composition and configuration of seagrass-dominated seascapes. Evidence of storm-related effects range from mild (e.g., Thompson and Phillips 1987; Wanless et al. 1988; Wilson et al. 2020) to severe (e.g., Correia 2021; Correia and Smee 2022; Eleuterius and Miller 1976; van Tussenbroek 1994) and are location- and storm-dependent (Ball et al. 1967; Congdon et al. 2019; Wilson et al. 2020). For example, storms resulted in fragmented seagrass beds that increased fish diversity in Thailand (Horinouchi et al. 2009) but not in Australia (Connolly 1994).
Linkages between hurricane disturbance of seagrass ecosystems and changes to associated fish communities have also been reported (Hall et al. 2016; Sargent et al. 1995) with evidence of seagrass habitat loss leading to decreased fish abundance, biomass, and species richness in Southern New England (Hughes et al. 2002), Sweden (Pihl et al. 2006) and Japan (Nakamura 2010). Understanding how storms influence the seagrass community is imperative given the likely influence of climate change on the frequency and intensity of tropical weather systems (Arellano-Méndez et al. 2011; Emanuel 2005; Goldenberg et al. 2001; Webster et al. 2005) and the recognized economic and societal benefits of seagrass ecosystems (Beck et al. 2001; Dahlgren et al. 2006; Rosenberg et al. 2000).
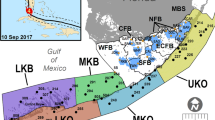
Hurricane Michael was the strongest hurricane to date to make landfall along the Florida panhandle (Masters 2018). To explore storm-induced effects on seagrass and associated nekton communities, we paired pre- and post-storm sampling of water quality, seagrass density, and seagrass-associated fauna (sensu Belgrad et al. 2021) at locations along a gradient of storm intensity following a massive category 5 storm, Hurricane Michael, which made landfall in Florida, USA, on 10 October 2018. In St. George Sound, FL, the storm had maximum sustained winds of 259 km h−1 and a 4.3-m storm surge (Beven et al. 2019; Masters 2018; Morgan 2019; NWS 2018). In nearby Cedar Key, FL (167 km southeast), the storm surge was 2.75 m above mean sea level (MSL), but there were no hurricane-force winds reported (Beven et al. 2019). In Chandeleur Islands, LA, 330 km west of Hurricane Michael’s landfall, no storm effects were noted. These three estuaries were sampled prior Hurricane Michael’s landfall, 2 weeks following the passage of the storm, and 1 year later to assess potential changes to seagrass and nekton communities. By sampling locations exposed to different storm components and relative intensities, we will gain a better understanding of the complex relationships between tropical weather systems, seagrass ecosystem disturbance, and nekton usage.
Methods
Study Regions
During the summers of 2018 and 2019, seagrass and associated nekton were sampled in seagrass beds at three regions across the Gulf of Mexico: St. George Sound, FL (29°51ʹ N, 84°37ʹ W), Cedar Key, FL (29°05ʹ N, 83°01ʹ W), and Chandeleur Islands, LA (29°54ʹ N, 88°50ʹ W). During site selection, each estuary containing seagrass was superimposed with hexagonal tessellation (500 m edge length) in ArcGIS (Moore 2009; Neckles et al. 2012; Wilson and Dunton 2012). Fifteen sites per estuary, each containing > 50% Thalassia testudinum (turtle grass) coverage and a minimum of 500 m separation, were randomly selected for assessment (Belgrad et al. 2021; Fig. 1). Across all regions, Thalassia testudinum was the dominant macrophyte taxa, with Syringodium filiforme (manatee grass) and Halodule wrightii (shoal grass) comprising the understory and small patches at study sites in all three estuaries. Other seagrass species were rarely encountered; thus, we focused on metrics from these three common species in analyses. Prior to the passage of Hurricane Michael, sampling took place at specific locations using a GPS from May to June (Early 2018) and August to September 2018 (Late 2018). Two weeks after the storm, the same sites were resampled (posthurricane), as well as the following year in May to June (Early 2019) and August to September (Late 2019) to determine potential storm effects on seagrass cover and associated nekton communities.

Location of the 3 study estuaries (stars) throughout the northern Gulf of Mexico in relation to Hurricane Michael just prior to landfall. Chandeleur Islands, LA, was located to the west of the storm path and experienced no wind or surge effects. St. George Sound, FL, was in the direct path of the storm and experienced maximum sustained winds of 259 km h−1 and a storm surge of 4.3 m. Cedar Key, FL, was located to the east of the storm center, with a storm surge of 2.75 m above mean sea level (MSL) with no wind effects (Beven et al. 2019)
Nekton Sampling
At each of the 15 site locations within each estuary (45 total), a 4.85-m flat otter trawl with a 3.8-m stretch mesh body and 6.4-mm stretch mesh bag was dragged through seagrass beds at an average speed of 2–3 knots for approximately 2 min, resulting in linear distances of approximately 130 m per trawl. Nekton (fish, portunid crabs, penaeid shrimp) caught in the net were counted, identified, and released. Because trawl distances varied, the density of organisms collected was standardized to trawl area (ind. ha−1), calculated using trawl width and GPS-estimated trawl tow length.
Seagrass Sampling
At each trawl location (15 sites estuary−1), the structural complexity of seagrasses was visually assessed using 1-m2 quadrats (divided into 100 10 cm × 10 cm grid cells). Three replicate 1-m2 quadrats were haphazardly deployed at the beginning, middle, and end of each trawl path (9 quadrats total per trawl path) to estimate percent cover, shoot count, and canopy height of T. testudinum, H. wrightii, and S. filiforme. Seagrass percent cover by species was measured by counting the number of grid cells within each quadrat that contained a particular vegetation type (0–100 cells quadrat−1). Total shoot count of all three dominant species was calculated by summing the shoot counts for T. testudinum, S. filiforme, and H. wrightii species within a random quadrat grid. Average overall canopy height was calculated by taking the average length of the species which produced the longest seagrass blades across each site. All quadrat measurements within each site were averaged to obtain estimates of seagrass cover, shoot count, and canopy height across each site.
Based on the overall percent cover in each site, an index of percent cover was calculated using a modified Braun–Blanquet (BB) cover-abundance score (Table 1) for comparisons of seagrass and nekton community composition in Primer™ 7. The frequency of each seagrass species in a particular site was also computed across the various sampling times, where Fi is the frequency of species i, Ni is the number of quadrats at a site in which species i was present, and n is the total number of quadrats sampled at a site (Kenworthy et al. 1993). This value was then multiplied by 100 to obtain a percentage calculation of seagrass species occurrence throughout the study:
Water Quality Measurements
After each trawl, water depth (cm), water temperature (°C), salinity, and dissolved oxygen (DO; mg L−1) were measured at the midpoint of the trawl track using a YSI Pro 2030 containing a galvanic DO sensor (Model 2002). Photosynthetically active radiation (PAR) was also measured using an LI-1500 light sensor logger equipped with two LI-193 spherical underwater quantum sensors separated by a standardized distance. Sensors recorded PAR over three consecutive 30-s intervals at 1 Hz, and the attenuation coefficient was calculated as a proxy for light availability. In the equation, the PARbottom and PARtop measurements refer to the PAR measurements of the top and bottom light sensors, and the sensor distance (m) is the vertical distance between the bottom sensor and top sensor when it is placed in the water column:
Statistical Analysis
All statistical analyses were performed using SAS© and Primer™ 7. We performed a mixed model analysis of variance in SAS© at an α of 0.05 (Horton and Lipsitz 2012; Sokal and Rohlf 2013), using site as a random factor and time sampled (early 2018, late 2018, posthurricane, early 2019, late 2019) as a fixed factor. We found significant differences across site, time sampled, and/or the interaction between site and time sampled across environmental (temperature, salinity, depth), seagrass (seagrass percent cover, total shoot count, and canopy height), and community (animal abundance (n), species richness (s), and diversity (H’)) variables (Tables 4, and S1). Specifically, there was a significant interaction between site and sample time for species diversity (H’), seagrass percent cover, total shoot count, canopy height, temperature, salinity, and depth (Tables 4, and S1). Prior to analysis, all variables were tested for normality, and animal abundance was log transformed to manage skewness and satisfy the normality assumption. Because of the significant interaction across site and time sampled, sites were then separated, and general linear regression models were run on the environmental, seagrass, and community variables with time sampled as the fixed factor (Tables 2 and 4). Tukey–Kramer post hoc comparisons were performed to compare means across sampling time. Dissolved oxygen was removed from the model because this metric remained above environmentally significant levels (> 4 mg/L) throughout the course of the experiment at all sites and estuaries. Continuous DO were not recorded, and a single static measure of DO does not provide information on the temporal oxygen changes in the water column.
Effect sizes were calculated using Cohen’s d to compare seagrass characteristics prior to and following the passage of Hurricane Michael. Cohen’s d measures the magnitude of difference between two sample means, with values of 0.2, 0.5, and 0.8 representing small (92% overlap), medium (80% overlap), and large (69% overlap) effects, respectively (Cohen 1988; Lakens 2013). Differences in effect size within each estuary provide a measure of practical significance across sampling times. A Cohen’s d value of 1 indicates that 1 standard deviation lies between the sample means:
In this equation, ABS is the absolute value, M1 and M2 are the average values for each seagrass characteristic during the late summer before the storm and 2 weeks following the storm event, and pooled SD is the mean standard deviation of both sampling times.
Community nekton data were square root transformed to manage skewness, and a Bray–Curtis resemblance matrix was calculated using Primer™ 7. RELATE and DistLM procedures were performed to determine which environmental (temperature, salinity, attenuation coefficient) and seagrass (percent cover, shoot count, canopy height) variables were important in explaining response patterns. Communities were then compared using one-way permutational multivariate analysis of variance (PERMANOVA) with 999 permutations and sampling time (early 2018, late 2018, posthurricane, early 2019, late 2019) as a fixed factor. Distance-based tests for homogeneity of multivariate dispersion (PERMDISP) were used to determine how sampling time affected group dispersions, using 999 permutations across centroid distances. Across all three sampling locations, PERMDISP indicated no significant dispersion of data between sampling events just prior to and following the storm event and the following year, indicating that the locations of the centroids, when applicable, were different. Similarity percentage (SIMPER) analyses were also performed to quantify which species contributed to similarity within each sampling time and, if so, how species changed following the storm event. A canonical analysis of principal coordinates (CAP) plot was used to visualize the temporal patterns in species composition among estuaries.
Results
Water Quality
Temperature was significantly lower across all three estuaries following the passage of the storm and varied across sample site (Tables 2, S1, and S2). In St. George Sound, temperatures fell from 30.41 ± 0.28 °C in September to 22.93 ± 0.27 °C following the storm. Similar declines were found in Cedar Key and Chandeleur Islands (Table S2). Salinity varied based on the interaction between site and time sampled (Table S1). Although salinity was significantly different across sampling time (p < 0.01; Table 2), post hoc comparisons found no significant difference between salinity levels just prior to and following the storm event in St. George Sound (p = 1.00), with an average salinity change < 1 ppt two weeks after the storm (Table S2). Salinity was also significantly different across sampling time in Cedar Key, FL, and Chandeleur Islands, LA (p < 0.01; Table 2), with post hoc comparisons indicating a significant increase in salinity just prior to and following the storm event (p < 0.01). These salinity changes, however, were < 4 ppt at each sample site (Table S2) and are unlikely to have affected vegetation and associated fauna. The attenuation coefficient varied across sampling periods at Cedar Key only (p < 0.01; Table 2), but post hoc comparisons showed that these changes were not temporally linked to the storm (p = 0.08; Table S2).
Vegetation
St. George Sound, FL — Hurricane Landfall Site with Storm Surge and Hurricane-Force Winds
Seagrass beds in St. George Sound sustained damaged from Hurricane Michael but displayed recovery. Seagrass cover, total shoot count, and average canopy height varied significantly among sample periods (p < 0.01; Table 2). Seagrass cover declined from 82.26 ± 8.77% to 50.45 ± 8.47% two weeks after the storm (p < 0.01), but 1 year later was not different from pre-storm values (75.41 ± 8.47%, p = 0.79; Fig. 2; Tables 2, and S2). Total shoot count and mean canopy height also exhibited a significant reduction following the storm (p < 0.01), but intermediate values the following year (Fig. 2; Table S2).

Pre- and post-storm monitoring of seagrass metrics bracketing of Hurricane Michael in 2018. Seagrass percent cover, mean seagrass shoot count, and mean canopy height (± SE) by sampling time for a St. George Sound, FL, proximate to landfall; b Cedar Key, FL, storm surge only; and c Chandeleur Islands, LA, no storm. Early2018 and Late2018 represent the sampling performed before the passage of the storm in May to June 2018 and August to September 2018, respectively. PostHurricane represents the sampling done 2 weeks after the storm. Early2019 and Late2019 represent that early and late sampling that took place 1 year following the storm (May to June 2019 and August to September 2019)
The Cohen’s d effect sizes attributable to Hurricane Michael were generally larger at the site directly impacted by the storm (St. George Sound) relative to the other two estuaries. In St. George Sound, the effect size (d) for seagrass percent cover was 1.09, indicating over 1 standard deviation of separation just prior to and 2 weeks following the storm. When comparing percent cover prior to the storm (Late 2018) compared to 1 year later (Late 2019), the effect size was smaller (d = 0.56), indicating that the difference in seagrass percent cover between 2018 and 2019 was smaller. Overall changes in shoot count and canopy height across the two sampling times were also large (d = 1.15 and 1.27, respectively) and remained large a year later (d = 1.06 and 1.37), indicating that there is still clear data separation across years. Significant declines in the frequency of T. testudinum and S. filiforme were also observed following the storm, falling from 92 to 66% and 50 to 27%, respectively (Table 3). Recovery in these two species was indicated by increases in their frequencies in 2019 sampling to near pre-storm levels (Table 3). The frequency of H. wrightii was low prior to the storm and did not change appreciably (Table 3).
Cedar Key, FL — Hurricane Produced Storm Surge But No Hurricane-Force Winds
Observed potential storm effects on seagrass metrics in Cedar Key were minimal. Changes in seagrass percent cover were nonsignificant (p = 0.41; Table 2), averaging 94.14 ± 3.13% prior to the storm and 93.13 ± 4.27% two weeks after the storm; percent cover remained high the following year (85.66 ± 4.13%; Fig. 2; Table S2). When comparing total shoot count and mean canopy height just before and 2 weeks after the storm, significant changes were found but were of small magnitude. Total shoot count actually increased by 26% when comparing pre- and post-storm values (p = 0.60), while canopy height showed a 29% reduction following the storm (p = 0.05; Table S2). When comparing the late summer sample events across years, shoot count was not significantly different (p = 0.92), but canopy height was again lower, declining by 33.5% compared to pre-storm conditions (p = 0.01; Fig. 2; Table S2). In Cedar Key, the effect size (d) for seagrass percent cover and canopy height were both small (d = 0.03 and 0.23, respectively), indicating a large degree of similarity in percent cover and canopy height just prior to and 2 weeks after the storm. Effect size estimates from late summer 2018 and 2019 were also smaller (d = 0.5, 0.3, and 0.4 for percent cover, shoot count, and canopy height, respectively) compared to our direct impact site (St. George Sound, FL). Small declines were recorded for T. testudinum frequency following the storm, from means of 93 to 87%, whereas H. wrightii and S. filiforme increased from 7 to 8% and from 47 to 69%, respectively (Table 3).
Chandeleur Islands, LA — No Storm Surge or Hurricane-Force Winds
In Chandeleur Islands, LA, changes in seagrass percent cover were also nonsignificant (p = 0.76; Table 2), averaging 96.69 ± 3.02% prior to the storm and 90.49 ± 4.18% two weeks after the storm; percent cover remained high the following year (92.6 ± 4.38%; Fig. 2; Table S2). Total shoot count exhibited a significant reduction 2 weeks following the storm, declining by 41.8% (Table S2). These changes in shoot count after the storm were not significantly different from the early summer sampling (Early 2018; p = 0.52; Fig. 2; Table S2). The mean canopy height did not significantly change following the storm (p = 0.50; Table S2).
The effect size (d) for seagrass percent cover was small (d = 0.2), indicating a mild separation of the points just prior to and 2 weeks after the storm. The effect size (d) for seagrass shoot count and canopy height were larger (d = 1.6 and 0.6, respectively), indicating a higher degree of separation prior to and 2 weeks after the storm event. Effect size estimates from late summer 2018 and 2019 followed a similar pattern, with percent cover showing little change across years (d = 0.2) and larger variability occurring in shoot count and canopy height (d = 1.1 and 1.8, respectively). Small declines in the frequency of T. testudinum occurred following the storm, from means of 88 to 76%. The frequency of H. wrightii and S. filiforme, however, increased from 7 to 14% and from 2 to 12%, respectively (Table 3). In general, the overall frequency of occurrence for each specific seagrass species exhibited no significant declines following the storm event (Table 3).
Seagrass-Associated Nekton
St. George Sound, FL — Hurricane Landfall Site with Storm Surge and Hurricane-Force Winds
Nekton abundance (N), species richness (S), and Shannon diversity (H’) within St. George Sound were not significantly different across time sampled (p = 0.61, 0.48, and 0.14, respectively; Tables 4, and S3). However, PERMANOVA indicated that community structure was different (pseudo-F = 6.11; p(perm) = 0.001), with clear separation of the nekton communities before and after the storm as well as 1 year later (Fig. 3). When comparing faunal communities with changes in seagrass percent cover, canopy height, and shoot count using the RELATE function, no significant relationships were found (ρ = 0.02; p = 0.38), indicating that time sampled was the most significant variable contributing to the observed change.

CAP plot of community structure across sampling time. Canonical analysis of principle coordinates (CAP) plot of a St. George Sound, FL; b Cedar Key, FL; and c Chandeleur Islands, LA, abundance data, comparing communities across time sampled. Community nekton data were square root transformed to manage skewness. Early2018 (blue triangle) and Late2018 (inverted red triangle) indicate samples taken in the early (May to June 2018) and late (August to September 2018) summer before the passage of the storm. PostHurricane (green square) is the sampling event 2 weeks following the passage of Hurricane Michael. Early2019 (purple diamond) and Late2019 (blue circle) are early (May to June 2019) and late (August to September 2019) summer samples taken 1 year following the passage of the storm
The three species responsible for the highest degree of similarity across all sites within St. George Sound prior to the storm event were Lagodon rhomboides (pinfish), Orthopristis chrysoptera (pigfish), and Bairdiella chrysoura (silver perch; Table S4). Two weeks following the storm, species contribution shifted to L. rhomboides, Farfantepenaeus spp. (penaeid shrimp), Chilomycterus schoepfi (striped burrfish), and Eucinostomus argenteus (spotfin mojarra; Table S4). One year later, the primary species contributing most to the similarity again reflected those species found prior to the storm (Table S4). Many species-specific abundances were also lower both after the storm and the following year (Table S7). Pinfish (L. rhomboides), for example, declined 72% following the storm, with further declines (28%) the following year (Table S7). Of the species which occurred in at least 4 of the 5 sampling events, 82% experienced a decline in abundance following the storm, and 64% showed declines 1 year later (e.g., O. chrysoptera, B. chrysoura, Callinectes sapidus (blue crab); Table S7).
Cedar Key, FL — Hurricane Produced Storm Surge But No Hurricane-Force Winds
Differences were observed in Cedar Key nekton abundance and diversity across sampling time, but storm effects on community structure were minimal. Nekton diversity (H’), species richness (S), and abundance (N) within Cedar Key all showed significant differences with time sampled (p < 0.05; Table 4). These differences, however, were not directly related to the storm, with no significant changes in nekton abundance, species richness, or diversity following the storms passage (p = 0.83, 0.30, and 0.09, respectively; Table S3). Nekton abundance, species richness, and diversity also stayed constant between late summer 2018 and 2019 (p = 0.29, 0.24, and 0.60, respectively; Table S3). PERMANOVA analysis indicated that the community structure was different based on sampling time (pseudo-F = 4.84; p(perm) = 0.001) as presented in the CAP plot (Fig. 3). However, differences were temporally proximate to the storm (Fig. 3). The RELATE function also found no significant relationships between the faunal and seagrass resemblance matrices (rho = 0.08; p = 0.11), indicating that time sampled was the most significant variable contributing to the observed change.
Changes in dominant species within nekton communities were minimal and did not indicate storm effects. The three nekton species responsible for creating the observed pattern of similarity across all sampling times within Cedar Key were L. rhomboides, O. chrysoptera, and B. chrysoura. There were minor shifts in relative contributions of O. chrysoptera and B. chrysoura to community composition, and Centropristis striata (black sea bass) was more consistently found following the storm (Table S5). These shifts, however, may have been due to temporal variation in community structure, not derived from the impact of Hurricane Michael. Some species abundances were also lower both after the storm and the following year (Table S7). Pinfish (L. rhomboides), for example, declined 53% following the storm, but increased by 6% the following year (Table S7). Of the species that occurred in at least 4 of the 5 sampling events, 59% experienced a decline in abundance following the storm, and only 23% showed declines 1 year later (e.g., Diplodus holbrookii (spottail pinfish), Farfantepenaeus spp., and Opsanus beta (Gulf toadfish); Table S7).
Chandeleur Islands, LA — No Storm Surge or Hurricane-Force Winds
Total nekton abundance (N) differed over time (p = 0.03; Table 4). Tukey post hoc analysis between late summer 2018 and 2 weeks following the storm showed no significant changes in nekton abundance, species richness, or diversity between those sampling periods (p = 0.96, 0.82, 0.78, respectively; Table S6). Nekton abundance, species richness, and diversity also stayed constant between late summer 2018 and 2019 sampling (p = 0.70, 1.0, and 0.87, respectively; Table S6). Nekton diversity (H’) and species richness (S) did not vary across sampling time (p > 0.05; Table 4); community structure did (pseudo-F = 15.59; p(perm) = 0.001, 999 permutations), but the differences were attributed to temporal variation noted before the storm (Fig. 3). No significant relationships were found between biological and vegetation resemblance matrices using the RELATE function (ρ = 0.1; p = 0.08).
Temporal shifts were observed in species’ contributions to community structure, but there were no clear changes associated with storm (Table S6). Lagodon rhomboides was the greatest contributor to similarity across early summer sampling in 2018 and 2019, as well as following the storm event. In the late summer, B. chrysoura was the greatest contributor to similarity across sites, followed by L. rhomboides, Syngnathus scovelli (Gulf pipefish), Cynoscion nebulosus (spotted seatrout), and Farfantepenaeus spp. The same species also contributed to the similarity found 1 year following the storm, with B. chrysoura and L. rhomboides contributing to the similarity found across sites (Table S6). Some species abundances were lower both after the storm and the following year (Table S7). Pinfish (L. rhomboides), for example, declined 73% following the storm, with further declines (5%) the following year (Table S7). Of the species which occurred in at least 4 of the 5 sampling events, only 46% experienced a decline in abundance following the storm, as well as 1 year later (e.g., O. chrysoptera, B. chrysoura, C. sapidus; Table S7).
Discussion
In an observation time series of seagrasses and associated nekton, we observed evidence of disturbance and initial recovery at the site closest to where the storm made landfall, which experienced hurricane-force winds and storm surge. One potential confounding factor of this study was fall senescence of seagrasses that was likely occurring in the fall when Hurricane Michael struck. To account for such temporal changes, we included two other sites in our study, one without storm effects and another that had storm surge but did not experience hurricane-force winds and then looked for temporal changes within sites. Previously, we noted inherent differences among these sites (Belgrad et al. 2021) and again found sites to be significantly different from one another in this study. Yet, in St. George Sound, the site closest to storm landfall, we found significant reductions in seagrass percent cover, shoot density, and canopy height two weeks after the storm. We attributed these changes to storm disturbance because we did not observe these changes in the other sites at this time. In St. George Sound, we also found seagrass percent cover to return to pre-storm levels and shoot density and canopy height to be of intermediate levels between those measured before vs. 2 weeks after the storm, suggesting recovery. Although we found temporal variation in seagrasses and associated nekton in all sites, St. George Sound was the only site in which these changes coincided with Hurricane Michael, which we interpret as storm disturbance and subsequent recovery.
The effects associated with storms largely depend on the type of ecosystem being studied, with documented trade-offs related to resistance and resilience of different ecosystems to disturbances (Patrick et al. 2022). Cyclones cause catastrophic damage to mangrove forests, with the most extensive damage occurring near the eye of the storm and directly along the storm-track (e.g., Smith et al. 1994; Dawes et al. 1995; Michot et al. 2002; Milbrandt et al. 2006; Zhang et al. 2019). Following these storm events, quicker recovery times are often found in areas furthest from the storm center, where wind is weaker and damage less severe (Milbrandt et al. 2006). Cyclones similarly impact salt marsh habitats (e.g., Cahoon 2006; Turner et al. 2006; Barras 2007), with increased wave energy leading to a linear increase in the rate of coastal salt marsh erosion (Leonardi et al. 2016). Yet, seagrasses appear resilient to these disturbances despite considerable variation in storm damage to them (Patrick et al. 2022).
The reported effects of storms on seagrass meadows are variable, ranging from no damage to complete habitat destruction (Correia and Smee 2022). Even within the same location, different storms elicit variable effects on seagrass meadows (Perkins and Enos 1968; Poiner et al. 1989; Fourqurean and Rutten 2004), and in some cases, the same storm can vary in the degree of damage observed (Williams 1990; Rodriguez et al. 1994; Michot et al. 2002).
Immediately following the passage of Hurricane Michael in St. George Sound, FL, we observed changes in seagrass species composition and declines in seagrass cover, shoot count, and canopy height, as well as a shift in seagrass-associated fauna. In St. George Sound, FL, seagrass cover declined from 83 to 50% immediately following the storm (p < 0.01) but did show signs of recovery the following year (75%). Shoot count and shoot height were also reduced by over 50% following the storm. These changes in seagrass percent cover were not observed at the other two estuaries.
A species shift also occurred, with pinfish being dominant prior to the storm and penaeid shrimp, striped burrfish, and spotfin mojarra more common following the storm event. Widespread declines in species-specific nekton abundances were observed following the storm, with continued reductions in some species the following year. Overall, both the seagrass and nekton communities showed signs of recovery, with seagrass (i.e., percent cover, shoot count, shoot height, frequency of occurrence) and nekton community composition returning to pre-storm levels the following year. Changes in fauna were not observed in other sites not directly impacted by the hurricane.
St. George Sound is a protected estuarine system. St. George Island, located to the South of our sampling locations, provided storm protection, and may have lowered the potential for wind damage to the seagrass. Salinity and turbidity did not change substantially following this storm and may explain the quick recovery time observed for seagrass communities. Turtle grass can also recover quickly when shoots are removed root system remains intact and are resilient after large scale disturbances (Hall et al. 2021).
Previous studies reported similar changes in nekton communities, with fluctuations in animal abundance and diversity following the passage of tropical systems (Greenwood et al. 2006; Paperno et al. 2006; Stevens et al. 2006; Switzer et al. 2006; Tabb and Jones 1962). Following the passage of two hurricanes in 2004, Paperno et al. (2006) reported the nekton community diversity within the Indian River lagoon estuary in Florida recovered to pre-hurricane baseline within several weeks. Similarly, hypoxic conditions in the Charlotte Harbor estuary, FL, following the passage of Hurricane Charley in 2004 led to a decline in fish abundance as well as an increase in the number of freshwater species, but fish assemblages returned to pre-storm conditions within a month of the disturbance (Stevens et al. 2006). Overall, nekton communities appear to be largely resilient to the direct effects of hurricanes and in our study, effects to the seagrass habitat were minimal and short-lived (Byron and Heck 2006; Dawes et al. 1995; Poiner et al. 1989; Sierra-Rozo et al. 2012; Thomas et al. 1961). More persistent changes to the nekton community following a disturbance are often associated with a permanent change in the topography relating to that disturbance event. For example, a major breach on Fire Island, NY, following the passage of Hurricane Sandy in 2012 led to wholesale changes in the abundance of both seagrasses and nekton assemblages (Tinoco 2017). Persistent salinity changes in Florida Bay have also led to a drastic, long-term changes in nekton and macroinvertebrate community composition (Zink et al. 2020). The present study experienced minimal salinity changes following the storm event, which could have influenced their ability to recover quickly.
Two additional storms made landfall along the northern Gulf of Mexico in the summer of 2018. Tropical Storm Alberto was the first named storm of the 2018 Hurricane Season, making landfall along the Florida panhandle on May 28, 2018. Similarly, Tropical Storm Gordon also made landfall along the northern Gulf of Mexico between the Alabama and Mississippi border on September 5, 2018. These storms passed through our study areas between the first and second sampling event of 2018. Seagrass and abundance metrics remained relatively stable between the first and second sampling event, making it unlikely that these storms influenced our results.
Our study precludes a mechanistic understanding of the observed changes, but they could have been driven by concurrent changes to habitat structure (i.e., seagrass plant community and morphometrics) and/or changes in abiotic conditions such as salinity and light availability resulting from the storm. Most studies of hurricane effects are opportunistic, and oftentimes occur after hurricanes strike during ongoing research (e.g., Correia 2021; Correia and Smee 2022). Having pre- and post-storm measurements that are consistent is rare, and using sentinel monitoring sites with consistent monitoring is needed to accurately capture ecosystem responses to storms (Hogan et al. 2020). When reviewing existing literature, Correia and Smee (2022) did not uncover clear relationships between the number and severity of storms and declines in seagrass cover. However, environmental fluctuations during and after the storm negatively impacted seagrasses. Plant burial and prolonged changes in water quality accounted for most reported seagrass declines (Correia and Smee 2022). In the current study, the decreased frequencies of seagrass occurrence in St. George Sound immediately after the passage of Hurricane Michael could have resulted from burial due to sediment changes, leaf removal due to waves and strong currents, or a combination of these factors (Fourqurean and Rutten 2004; Heck et al. 1996; Michot et al. 2002; Timant et al. 1994). While not measured, multiple sites were observed to be partially or completely buried by sediment following the passage of the storm in St. George Sound. Future research needs to incorporate environmental changes observed within seagrass beds following storms because these factors are important to recovery.
Vegetation changes in both the surge impacted site (Cedar Key, FL) and areas not affected by the storm (Chandeleur Islands, LA) did not coincide with the storm and were attributed to seasonal changes. Although seagrass shoot density in the Chandeleur Islands declined significantly following the storm and remained low the following year, other seagrass characteristics including canopy height, seagrass percent cover, and the composition of seagrass species did not change, suggesting the change in shoot density is not storm-related. The shoot density fluctuations observed could be driving the variability in the nekton community observed as well. The Chandeleur Islands are known to host a dynamic seagrass system that has historically seen rapid changes in seagrass cover and species composition (Darnell et al. 2017). Seagrass beds there had been considered pristine due to the limited accessibility and general lack of direct human impacts (i.e., pollution and nutrient loading) (Poirrier and Handley 2007). However, in recent years, the islands have undergone significant changes due to storm and hurricane impacts (Fearnley et al. 2009; Sherwood et al. 2014; Williams et al. 1997), potentially contributing to the shoot density fluctuation seen in this study. Although we cannot account for the shoot density change, the magnitude of the change was much smaller than those observed at our impacted site, and we concluded that the effect was not related to the storm.
Structural complexity within aquatic systems is a major driver of nekton community composition and diversity (Bartholomew et al. 2000; Thayer et al. 1987; Vince et al. 1976), and an increased understanding of how disturbance events influence changes in vegetation and associated nekton communities is extremely important in a changing climate where storms are projected to become more intense and frequent. In this study, the passage of a category 5 hurricane coincided with immediate changes to seagrass and nekton communities. Changes observed were short-lived, lasting weeks to months after the storm made landfall with recovery to pre-storm conditions 1-year after the storm. These results reveal the resilience of seagrasses and their associated nekton communities. Understanding how disturbance events influence changes in vegetation and associated nekton communities is important in a changing climate where storms are projected to become more intense and frequent.
Data Availability
The data sets presented in this study can be found in online repositories. The names of the repositories and accession can be found: http://www.darnellseagrassecologylab.com/turtlegrass.
References
Arellano-Méndez, L.U., M.A. Liceaga-Correa, J.A. Herrera-Silveira, and H. Hernandez-Nuñez. 2011. Impacto por huracanes en las praderas de Thalassia testudinum Hydrocharitaceae en el Caribe Mexicano. Revista de Biología Tropical 59 (1): 385–401.
Ball, M.M., E.A. Shinn, and K.W. Stockmann. 1967. The geological effects of Hurricane Donna in south Florida. The Journal of Geology 75 (5): 583–597.
Barras, J.A. 2007. Land area changes in coastal Louisiana after Hurricanes Katrina and Rita. In Science and the Storms: The USGS Response to the Hurricanes of 2005, ed. G.S. Farris, G.J. Smith, M.P. Crane, C.R. Demas, L.L. Robbins, and D.L. Lavoie, 98–113. U.S. Geological Survey Circular 1306.
Bartholomew, A., R.J. Diaz, and G. Cicchetti. 2000. New dimensionless indices of structural habitat complexity: Predicted and actual effects on a predator’s foraging success. Marine Ecology Progress Series 206: 45–58.
Beck, M.W., K.L. Heck, K.W. Able, D.L. Childers, D.B. Eggleston, B.M. Gillanders, et al. 2001. The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. BioScience 51: 633–641.
Belgrad, B.A., K.M. Correia, K.M. Darnell, M.Z. Darnell, C.T. Hayes, M.O. Hall, B.T. Furman, C.W. Martin, and D.L. Smee. 2021. Environmental drivers of seagrass-associated nekton abundance across the northern Gulf of Mexico. Estuaries and Coasts. https://doi.org/10.1007/s12237-021-00927-0.
Beven, J.L., II., R. Berg, and A. Hagen. 2019. Hurricane Michael (AL142018), 86. National Hurricane Center. Report nr AL142018. https://www.nhc.noaa.gov/data/tcr/AL142018_Michael.pdf. Accessed 11 Nov 2021.
Birch, W.R., and M. Birch. 1984. Succession and pattern of tropical intertidal seagrasses in Conckle Bay, Queensland, Australia: A decade of observations. Aquatic Botany 19: 343–367.
Braun-Blanquet, J., 1932. Plant sociology. The study of plant communities. Plant sociology. The study of plant communities. First ed.
Byron, D., and K.H. Heck Jr. 2006. Hurricane effects on seagrasses along Alabama’s Gulf Coast. Estuaries and Coasts 29 (6A): 939–942.
Cahoon, D.R. 2006. A review of major storm impacts on coastal wetland elevations. Estuaries and Coasts 29: 889–898.
Cohen, J. 1988. Statistical power analysis for the behavioral sciences, 2nd ed. Hillsdale, NJ: Erlbaum.
Congdon, V.M., C. Bonsell, M.R. Cuddy, and K.H. Dunton. 2019. In the wake of a major hurricane: differential effects on early vs. late successional seagrass species. Limnology and Oceanography Letters 4(5):155–163.
Connolly, R.M. 1994. Removal of seagrass canopy: Effects on small fish and their prey. Journal of Experimental Marine Biology and Ecology 184: 99–110.
Correia, K.M. 2021. What doesn’t kill us makes us stronger: seagrass community resilience in the Northern Gulf of Mexico. PhD dissertation. University of South Alabama, Mobile, AL. https://www.proquest.com/openview/ca88307b910ab0796efabe71e1c2e241/1?pq-origsite=gscholar&cbl=18750&diss=y. Accessed 11 Nov 2021.
Correia, K.M. and D.L. Smee. 2022. A review of tropical cyclone effects on seagrass meadows. Wetlands 42(8).
Dahlgren, C.P., G.T. Todd, A.J. Adams, B.M. Gillanders, M.S. Kendall, C.A. Layman, et al. 2006. Marine nurseries and effective juvenile habitats: Concepts and applications. Marine Ecology Progress Series 312: 291–295.
Darnell, K.M., T.J.B. Carruthers, P. Biber, et al. 2017. Spatial and temporal patterns in Thalassia testudinum leaf tissue nutrients at the Chandeleur Islands, Louisiana, USA. Estuaries and Coasts 40: 1288–1300.
Dawes, C.J., S.S. Bell, R.A. Davis Jr., E.D. McCoy, H.R. Mushinsky, and J.L. Simon. 1995. Initial effects of Hurricane Andrew on the shoreline habitats of southwestern Florida. Journal of Coastal Research SI 21: 103–110.
Eleuterius, L.N., and G.J. Miller. 1976. Observations on seagrasses and seaweeds in Mississippi Sound since Hurricane Camille. Journal of the Mississippi Academy of Science 21: 58–63.
Emanuel, K. 2005. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 436 (4): 686–688. https://doi.org/10.1038/nature03906.
Fearnley, S.M., M.D. Miner, M. Kulp, C. Bohling, and S. Penland. 2009. Hurricane impact and recovery shoreline change analysis of the Chandeleur Islands, Louisiana, USA: 1855 to 2005. Geo-Marine Letters 29: 455–466.
Fourqurean, J.W., and L.M. Rutten. 2004. The impact of Hurricane Georges on soft-bottom, back reef communities: Site- and species-specific effects in South Florida seagrass beds. Bulletin of Marine Science 75 (2): 239–257.
Goldenberg, S.B., C.W. Landsea, A.M. Mestas-Nunez, and W.M. Gray. 2001. The recent increase in Atlantic Hurricane activity: Causes and implications. Science 293: 474–479.
Greenwood, M.F.D., P.W. Stevens, and R.E. Matheson Jr. 2006. Effects of the 2004 hurricanes on the fish assemblages in two proximate southwest Florida estuaries: Change in the context of interannual variability. Estuaries and Coasts 29 (6A): 985–996.
Hall, M.O., B.T. Furman, M. Merello, and M.J. Durako. 2016. Recurrence of Thalassia testudinum seagrass die-off in Florida Bay, USA: Initial observations. Marine Ecology Progress Series 560: 243–249.
Hall, M.O., S.S. Bell, B.T. Furman, et al. 2021. Natural recovery of a marine foundation species emerges decades after landscape-scale mortality. Science and Reports 11: 6973. https://doi.org/10.1038/s41598-021-86160-y.
Heck, K.L., M.J. Sullivan, J.M. Zande, and C.A. Moncreiff. 1996. An ecological analysis of seagrass meadows of the Gulf Islands National Seashore, years one, two, and three: seasonal assessment and inventory, interaction studies and continuing assessment/inventory, final report, August 1996: Gulf Breeze, Fla., National Park Service, Gulf Islands National Seashore, 379.
Hogan, J.A., R.A. Feagin, G. Starr, M. Ross, T. Lin, C. O’connell, T.P. Huff, B.A. Stauffer, K.L. Robinson, et al. 2020. A research framework to integrate cross-ecosystem responses to tropical cyclones. BioScience 70 (6): 477–489.
Horinouchi, M., P. Tongnunui, K. Nanjyo, Y. Nakamura, M. Sano, and H. Ogawa. 2009. Differences in fish assemblage structures between fragmented and continuous seagrass beds in Trang, southern Thailand. Fisheries Science 75: 1409–1416.
Horton, N.J., and S.R. Lipsitz. 2012. Review of software to fit generalized estimating equation regression models. The American Statistician 53: 160–169.
Hughes, J.E., L.A. Deegan, J.C. Wyda, M.J. Weaver, and A. Wright. 2002. The effects of eelgrass habitat loss on estuarine fish communities of southern New England. Estuaries 25 (2): 235–249.
Kenworthy, W.J., M.J. Durako, S.M.R. Fatemy, H. Valavi, and G.W. Thayer. 1993. Ecology of seagrasses in northeastern Saudi Arabia one year after the Gulf-War oil spill. Marine Pollution Bulletin 27: 213–222.
Lakens, D. 2013. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Frontiers in Psychology 4: 863. https://doi.org/10.3389/fpsyg.2013.00863.
Leonardi, N., N.K. Ganju, and S. Fagherazzi. 2016. A linear relationship between wave power and erosion determines salt-marsh resilience to violent storms and hurricanes. Proceedings of the National Academy of Sciences of the United States of America 113 (1): 64–68.
Marchand, J., I. Codlin, P. Drake, M. Elliot, L. Pihl, and J. Rebelo. 2002. Chapter 7: Environmental quality of estuaries. In: Fishes in Estuaries, ed. M., Elliott, and K.L. Hemingway. 2008. Wiley.
Masters, J. 2018. Hurricane Michael makes landfall in Florida Panhandle, 6. https://www.wunderground.com/cat6/Potentially-Catastrophic-Hurricane-Michael-Nearing-Landfall-Florida-Panhandle. Accessed 11 Nov 2021.
Michot, T.C., J.N. Burch, A. Arrivillaga, P.S. Rafferty, T.W. Doyle, and R.S. Kemmerer. 2002. Impacts of Hurricane Mitch on seagrass beds and associated shallow reef communities along the Caribbean coasts of Honduras and Guatemala 61 p.
Milbrandt, E.C., J.M. Greenawalt-Boswell, P.D. Sokoloff, and S.A. Bortone. 2006. Impact and response of southwest Florida mangroves to the 2004 hurricane season. Estuaries and Coasts 29 (6A): 979–984.
Moore, K. 2009. Long-term monitoring for estuarine submersed and emergent vegetation communities. NERR SWMP Biomonitoring Protocol. National Estuarine Res Reserve Syst. NOAA Ocean Service. Silver Spring, MD. https://coast.noaa.gov/data/docs/nerrs/Research_TechSeries_TechReportSWMPBio-MonitoringProtocol.pdf. Accessed 11 Nov 2021.
Morgan, L. 2019. Hurricane Michael anniversary: a look back at a category 5 monster. Advanced Local Media LLC. https://www.al.com/hurricane/2019/10/hurricane-michael-anniversary-a-look-back-at-a-category-5-monster.html. Accessed 11 Nov 2021.
Nakamura, Y. 2010. Patterns in fish response to seagrass bed loss at the southern Ryukyu Islands, Japan. Marine Biology 157: 2397–2406.
Neckles, H.A., B.S. Kopp, B.J. Peterson, and P.S. Pooler. 2012. Integrating scales of seagrass monitoring to meet conservation needs. Estuaries and Coasts 35: 23–46.
NWS. 2018. Hurricane Michael 2018. Catastrophic Hurricane Michael strikes Florida Panhandle. October 10, 2018. https://www.weather.gov/tae/HurricaneMichael2018#:~:text=Michael%20made%20landfall%20along%20the,of%20919%20millibars%20(mb)%20. Accessed 11 Nov 2021.
Paperno, R., D.M. Tremain, D.H. Adams, A.P. Sebastian, J.T. Sauer, and J. Dutka-Gianelli. 2006. The disruption and recovery of fish communities in the Indian River Lagoon, Florida, following two hurricanes in 2004. Estuaries and Coasts 29 (6A): 1004–1010.
Patrick, C.J., J.S. Kominoski, W.H. McDowell, B. Branoff, D. Lagomasino, M. Leon, E. Hensel, M.J.S. Hensel, B.A. Strickland, X. Zou, et al. 2022. A general pattern of trade-offs between ecosystem resistance and resilience to tropical cyclones. Science Advances 8 (9): 10.
Perkins, R.D., and P. Enos. 1968. Hurricane Betsy in the Florida-Bahama area: Geologic effects and comparison with Hurricane Donna. The Journals of Geology 76 (6): 710–717.
Pihl, L., S. Baden, N. Kautsky, P. Rönnbäck, T. Söderqvist, M. Troell, and H. Wennhage. 2006. Shift in fish assemblage structure due to loss of seagrass Zostera marina habitats in Sweden. Estuarine Coastal and Shelf Science 67: 123–132.
Poiner, I.R., D.I. Walker, and R.G. Coles. 1989. Regional studies: seagrasses of tropical Australia. Biology of Seagrasses: Aquatic Plant Studies 2, 279–303. Amsterdam: Elsevier.
Poirrier, M., and L. Handley. 2007. Chandeleur Islands. In: Seagrass Status and Trends in the Northern Gulf of Mexico: 1940–2002. U.S. Geological Survey Scientific Investigations Report 2006–5287 and U.S. Environmental Protection Agency 855-R-04–003.
Rodriguez, R.W., R.M.T. Webb, and D.M. Bush. 1994. Another look at the impact of Hurricane Hugo on the shelf and coastal resources of Puerto Rico, U.S.A. Journal of Coastal Research 10 (2): 278–296.
Rosenberg, A., T.E. Bigford, S. Leathery, R.L. Hill, and K. Bickers. 2000. Ecosystem approaches to fishery management through essential fish habitat. Bulletin of Marine Science 66: 535–542.
Rozas, L.P., and D.J. Reed. 1994. Comparing nekton assemblages of subtidal habitats in pipeline canals traversing brackish and saline marshes in coastal Louisiana. Wetlands 14: 262–275.
Sargent, F.J., T.J. Leary, D.W. Crewz, and C.R. Kruer. 1995. Scarring of Florida’s seagrasses: assessment and management, 62. Technical Report. FMRI 1h/94, St. Petersburg, Florida: Florida Marine Research Institute. http://www.sarasota.wateratlas.usf.edu/upload/documents/ScarringOfFloridaSeagrass1995.pdf. Accessed 11 Nov 2021.
Sherwood, C.C., J.W. Long, P.J. Dickhudt, S. Dalyander, D.M. Thompson, and N.G. Plant. 2014. Inundation of a barrier island (Chandeleur Islands, Louisiana, USA) during a hurricane: observed water-level gradients and modeled seaward sand transport JGR. Earth Science 119 (7): 1498–1515.
Sierra-Rozo, O., B. Gavio, and J.E. Mancera-Pineda. 2012. Structure of the Thalassia testudinum praderas in the island of Providencia, Colombian Caribbean, after the pass of Hurricane Beta. Cadasia 34 (1): 155–164.
Smith, T.J., III., M.B. Robblee, H.R. Wanless, and T.W. Doyle. 1994. Mangroves, hurricanes, and lightning strikes: Assessment of Hurricane Andrew suggests an interaction across two differing scales of disturbance. BioScience 44 (4): 256–262.
Sokal, R., and F.J. Rohlf. 2013. Biometry: the principles and practice of statistics in biological research, 3rd ed. New York, New York, USA: B WH Freeman.
Stevens, P.W., D.A. Blewett, and J.P. Casey. 2006. Short-term effects of a low dissolved oxygen event on estuarine fish assemblages following the passage of Hurricane Charley. Estuaries and Coasts 29 (6A): 997–1003.
Switzer, T.S., B.L. Winner, N.M. Dunham, J.A. Whittington, and M. Thomas. 2006. Influence of sequential hurricanes on nekton communities in a southeast Florida estuary: Short-term effects in the context of historical variations in freshwater inflow. Estuaries and Coasts 29 (6A): 1011–1018.
Tabb, D.C., and A.C. Jones. 1962. Effect of Hurricane Donna on the aquatic fauna of North Florida Bay. Translations of the American Fisheries Society 91 (4): 375–378.
Thayer, G.W., D.R. Colby, and W.F. Hettler Jr. 1987. Utilization of the red mangrove prop root habitat by fishes in south Florida. Marine Ecology Progress Series 35: 25–38.
Thomas, L.P., D.R. Moore, and R.C. Work. 1961. Effects of Hurricane Donna on the turtle grass beds of Biscayne Bay, Florida. Bulletin of Marine Science of the Gulf and Caribbean 11 (2): 191–197.
Thompson, M.J., and N.W. Phillips. 1987. Assessment of hurricane damage in the Florida Big Bend seagrass beds, 95. New Orleans. https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.160.6616&rep=rep1&type=pdf. Accessed 11 Nov 2021.
Timant, J.T., R.W. Curry, R. Jones, A. Szmant, J.C. Zieman, and M. Flora. 1994. Hurricane Andrew’s effects on marine resources. BioScience 44 (4): 7. https://doi.org/10.2307/1312227.
Tinoco, A.I. 2017. Effects of Hurricane Sandy on Great South Bay, Long Island: Assessing water quality, seagrass and associated nekton communities, 88. Stony Brook University. https://ir.stonybrook.edu/xmlui/bitstream/handle/11401/77767/Tinoco_grad.sunysb_0771M_13319.pdf?sequence=1. Accessed 11 Nov 2021.
Turner, R.E., J.J. Baustian, E.M. Swenson, and J.S. Spicer. 2006. Wetland sedimentation from Hurricanes Katrina and Rita. Science 314: 449–452.
van Tussenbroek, B.I. 1994. The impact of hurricane Gilbert on the vegetative development of Thalassia testudinum in Puerto Morelos Coral Reef Lagoon, Mexico: A retrospective study. Botanica Marina 37 (5): 421–428.
Vince, S., I. Valiela, N. Backus, and J.M. Teal. 1976. Predation by the salt marsh killifish Fundulus heteroclitus (L.) in relation to prey size and habitat structure: Consequences for prey distribution and abundance. Journal of Experimental Marine Biology and Ecology 23: 255–266.
Walker, L.M., P.A. Montagna, X. Hu, and M.S. Wetz. 2021. Timescales and magnitude of water quality change in three Texas estuaries induced by passage of Hurricane Harvey. Estuaries and Coasts 44: 960–971.
Wanless, H.R., L.P. Tedesco, and K.M. Tyrrell. 1988. Production of subtidal tubular and surficial tempestites by Hurricane Kate, Caicos platform, British West Indies. Journal of Sedimentary Research 58 (4): 739–750.
Webster, P.J., G.J. Holland, J.A. Curry, and H.R. Change. 2005. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 309: 1844–1846.
Williams, J., S. Penland, and A.H. Sallenger. 1997. Atlas of shoreline changes in Louisiana from 1853 to 1989: Louisiana barrier island erosion study: U.S. Geological Survey, Miscellaneous Investigations Series I-2150-A, 103. https://doi.org/10.3133/i2150A
Williams, S.L. 1990. Experimental studies of Caribbean seagrass bed development. Ecological Monographs 60 (4): 449–469.
Wilson, C.J., and K.H. Dunton. 2012. Assessment of seagrass habitat quality and plant condition in Texas coastal waters, 46. Report submitted to the Coastal Bend Bays and Estuaries Program. Project number 1201. https://www.cbbep.org/publications/FinalSeagrassReportTier2.pdf. Accessed 11 Nov 2021.
Wilson, S.S., B.T. Furman, M.O. Hall, and J.W. Fourqurean. 2020. Assessment of Hurricane Irma impacts on South Florida seagrass communities using long-term monitoring programs. Estuaries and Coasts 43: 1119–1132.
Zhang, C., S.D. Durgan, and D. Lagomasino. 2019. Modeling risk of mangroves to tropical cyclones: A case study of Hurricane Irma. Estuarine, Coastal and Shelf Science 224: 108–116.
Zink, I.C., J.A. Browder, C.R. Kelble, E. Stabenau, C. Kavanagh, and Z.W. Fratto. 2020. Hurricane-mediated shifts in a subtropical seagrass associated fish and macroinvertebrate community. Estuaries and Coasts 43: 1174–1193.
Acknowledgements
The authors would like to thank Samantha Smith for assistance in the field and laboratory and the reviewers who provided helpful edits to the manuscript prior to publication.
Funding
This paper is a result of research funded by the National Oceanic and Atmospheric Administration’s RESTORE Science Program under award NA17NOS4510093 to the University of Southern Mississippi, Dauphin Island Sea Lab, University of Florida, and Florida Fish and Wildlife Conservation Commission.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Communicated by Paul A. Montagna
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Correia, K.M., Alford, S.B., Belgrad, B.A. et al. Hurricane Effects on Seagrass and Associated Nekton Communities in the Northern Gulf of Mexico. Estuaries and Coasts 47, 162–175 (2024). https://doi.org/10.1007/s12237-023-01276-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-023-01276-w