
Abstract
Hurricanes are some of the largest environmental drivers of change in coastal systems. We investigated the impacts of Hurricane Irma on benthic macrophyte communities in Florida Bay (FB) and the Florida Keys National Marine Sanctuary (FKNMS), USA. Spatiotemporal analyses were performed at multiple hierarchical levels (site, zone, region) to identify potential changes in the Braun-Blanquet (BB) densities of total seagrass (TSG) and total calcareous green macroalgae (TCAL) post-disturbance and to determine whether changes were attributable to hurricane impacts or normal seasonal and inter-annual variability. There were significant decreases in TSG in one of five zones in FKNMS and in one of six zones in FB, but no change in TCAL was recorded in either system. TSG in the Lower Keys Bayside declined from a mean BB score of 2.6 to 1.2, resulting from storm-induced erosion, whereas TSG in coastal FB declined from 1.05–2.4 to 0.36–2.0, likely due to prolonged hyposalinity and low dissolved oxygen following stormwater drainage. Overall, impacts to South Florida benthic macrophyte communities from Hurricane Irma were not catastrophic and were limited in spatial extent. Our results suggest that coastal areas hit by a storm with heavy winds are more likely to sustain direct physical impacts to the benthos, whereas estuarine areas with longer residence times are more at risk of the indirect effects of stormwater runoff and retention. Our analyses placed putative hurricane impacts within the context of recent variability and historical system baselines through the use of long-term monitoring data coordinated by multiple governmental and academic entities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Hurricanes, typhoons, and cyclones (hereafter, ‘hurricanes’) are some of the largest drivers of physical disturbance in shallow tropical benthic systems, and both the frequency and intensity of these storms have increased in recent years (Webster et al. 2005; IPCC 2014). Hurricanes may cause extensive seagrass meadow destruction or destabilization after their passage due to both direct and indirect mechanisms (Carlson et al. 2010). Here, we operationally define ‘direct’ impacts as (1) those related to sediment deposition/seagrass burial and (2) the erosion of sediments and loss of seagrass above- and below-ground biomass following exposure to high winds, strong currents, or wave energy; ‘indirect’ impacts are those arising from changes in water quality resulting from excessive rainfall and turbid freshwater run-off. Some research has shown highly localized hurricane impacts to seagrasses, with a given storm affecting just a few areas within a network of sites (Fourqurean and Rutten 2004; Tilmant et al. 1994; Tussenbroek 1994). Other studies have shown virtually no impact to seagrasses following the passage of a hurricane (Anton et al. 2009; Byron and Heck 2006; Sierra-Rozo et al. 2012). Somewhat paradoxically, instances of increased seagrass density have been documented during months following a storm, presumably from the removal and export of dead plant material from the system (Dingtian and Daojian 2011). Species-specific impacts of hurricanes on seagrasses are also apparent: in multi-species meadows, shallow-rooted taxa such as calcareous green macroalgae and Halodule wrightii can be more easily dislodged than firmly anchored Thalassia testudinum (Fourqurean and Rutten 2004). The variety of documented hurricane impacts to seagrasses suggest that storm effects vary not only between storms and systems, but also across the seascape (e.g., as documented for Hurricane Donna across South Florida in 1960; Ball et al. 1967). Thus, a multitude of factors govern the degree of impact that a hurricane will have on seagrass communities, such as strength and trajectory of the storm and the physical characteristics (e.g., bathymetry, geomorphology, hydrology, preexisting macrophyte coverage) of the system in question.
Long-term environmental monitoring programs provide important datasets for coastal managers and decision-makers in the aftermath of tropical disturbances. Monitoring networks that are spatially broad and span environmental gradients allow for detailed understanding of system responses across a variety of scales (Fourqurean and Rutten 2003). The South Florida seascape is home to some of the most expansive seagrass meadows in the northern hemisphere, covering almost 15,000 km2 (Fourqurean et al. 2002), and long-term seagrass monitoring programs in Florida Bay (FB) and the Florida Keys National Marine Sanctuary (FKNMS) have documented the status and trends of South Florida seagrass meadows for more than two decades, providing a baseline for current and future work (Durako et al. 2002; Fourqurean et al. 2002). Throughout this time (1995–2017), and despite the passage of multiple hurricanes, South Florida seagrass distribution and community structure have remained generally stable, except for large-scale T. testudinum mortality events in west-central FB in 1987 (Robblee et al. 1991) and 2015 (Hall et al. 2016). Since the inception of the FKNMS and FB seagrass monitoring programs in 1995, Hurricane Irma in 2017 was the strongest storm to cross the Florida Keys. While other hurricanes have passed near the area during the monitoring period (e.g., Irene in 1999, Charley in 2004, Dennis, Katrina, Rita and Wilma in 2005), since 1995, only Georges, a Category 2 hurricane in 1998 with maximum sustained winds of 167 kph, made landfall.
Hurricane Irma made landfall on Cudjoe Key, FL, on 10 September 2017 as a Category 4 storm with sustained wind speeds of 213 kph (Cangialosi et al. 2018). Hurricane force winds (> 118 kph) extended 130 km from the center, and tropical storm force winds (> 63 kph) extended 350 km from the center (NHC 2017). Irma’s massive size resulted in hurricane-force winds buffeting the entire Florida Keys island chain, including the nearshore marine habitats of FKNMS and FB (Fig. 1 inset). The storm traveled due north over the Florida Keys to make a second landfall in Marco Island, FL. The Lower Keys experienced stronger winds (> 148 kph) than the Upper Keys, the strongest of which were associated with the eyewall (sustained speeds of 213 kph) near Cudjoe Key (RAMMB 2017; Cangialosi et al. 2018).

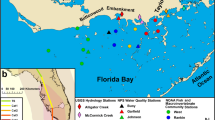
Florida Bay (FB) and Florida Keys National Marine Sanctuary (FKNMS) study sites. Black outlines indicate spatial zones (FB zones in white, FKNMS zones in color). Black points represent FKNMS transect stations, and orange points represent FB transect stations. FB spatial sites are shown as basins in blue. Inset is the path and wind swath of Hurricane Irma
Following the storm, large mats of seagrass leaves were observed floating in the water and deposited as wrack along mangrove and sandy shorelines, raising concerns that Irma had severely damaged the seagrass meadows of South Florida (https://www.miamiherald.com/news/weather/hurricane/article173637481.html). To address these concerns, we re-visited our long-term monitoring stations and compared pre- and post-storm results for evidence of storm-related impacts, leveraging historical data to provide ecological context to putative Irma effects. Because Hurricane Irma made landfall within 30 km of where Hurricane Georges passed in 1998, these data also provided the unique opportunity to compare macrophyte responses to similar perturbation across the same monitoring network. Based on historical data, we anticipated minimal and spatially restricted storm impacts to seagrass communities (Thomas et al. 1961; Tilmant et al. 1994; Fourqurean and Rutten 2004). Specifically, we expected that Irma would again impact stations that had been affected by Georges, i.e., those close to breaks in the reef tract that were more exposed to storm-generated currents. Further, we posited that the degree of protection afforded seagrasses by adjacent coral reefs or the Florida Keys island chain would control how the system responded to physical stress.
The purposes of this study were (1) to assess the impacts of Hurricane Irma on South Florida macrophyte communities, (2) quantify any changes in abundance or community composition, (3) identify specific areas of seagrass vulnerability, and (4) predict what role, if any, future storms might play in re-structuring benthic macrophyte communities in the region. We also sought to integrate results from multiple seagrass monitoring programs in South Florida to reach a holistic understanding of benthic community responses across the 15,000-km2, seagrass-dominated seascape.
Materials and Methods
Site Description and Field Methods
Our study examined macrophyte coverage across the South Florida seascape, which is divided into two separate management areas: Florida Bay (FB) and the Florida Keys National Marine Sanctuary (FKNMS; Fig. 1). Comprising the southern portion of Everglades National Park, FB is the sheltered area at the southern terminus of the Florida peninsula bounded by the Florida Keys and the Gulf of Mexico. FB is shallow (average depth of 1 m, maximum depths < 3 m) and composed of many small basins, with hydrologic connectivity among basins restricted by a network of shallow mud banks (Nuttle et al. 2000). The FKNMS boundary encircles the Florida Keys, encompassing the offshore reef tract and back-reef areas to the east and south of the Keys archipelago, as well as the backcountry areas to the north of the Florida Keys and west of FB. T. testudinum is the dominant canopy-forming seagrass species throughout most of the study area, with Syringodium filiforme occurring as an understory in western FB and dominant in portions of north-central FKNMS. H. wrightii is abundant in nearshore areas of FKNMS, and in the northern and eastern regions of FB, although it can be found at low densities throughout the seascape (Fourqurean et al. 2002). Halophila decipiens is the dominant seagrass species in the deeper (15–30 m depth) regions to the west of Key West and on the Southwest Florida Shelf between Marco Island and the Florida Keys (Fourqurean et al. 2002); however, without permanent transects, we could not assess hurricane impacts on offshore H. decipiens.
Several regional seagrass monitoring programs exist across the FB and FKNMS management areas, all of which were designed to follow similar protocols (Fourqurean et al. 2002). For our analysis, we compiled data from three monitoring programs, (1) the Florida Fish and Wildlife Conservation Commission (FWC) Fisheries Habitat Assessment Program (FHAP), (2) the Florida Coastal Everglades Long-Term Ecological Research (FCE-LTER) program, and (3) the FKNMS Water Quality Protection Program (WQPP; Table 1). All seagrass monitoring sites were regularly sampled in the spring or summer of each year; thus, our pre-storm data were from May and June, 2017. After Hurricane Irma, monitoring teams re-sampled all or a portion of their network between October and December 2017.
One aspect of the FHAP seagrass monitoring program incorporates 16 basins within FB (Table 1), which were each partitioned into 29 to 31 hexagonal grid cells. One sampling site (“spatial site”) per grid cell (“spatial basin”) was randomly chosen each year and field-located using handheld GPS, except in fall 2017 when the GPS locations from spring 2017 were re-sampled in order to increase our ability to assess hurricane impacts. GPS points were marked with the boat anchor, and eight 0.25-m2 quadrats were haphazardly distributed inside a 10-m radius of the anchored vessel. For each quadrat, all conspicuous benthic sessile taxa (including seagrasses, macroalgae, sponges, and corals) were assigned a modified Braun-Blanquet (BB) cover-abundance score, with 0 = absent, 0.1 = less than 5% cover by a single individual or short shoot, 1 = < 5% cover with many individuals/shoots, 2 = > 5% to 25% cover, 3 = > 25 to 50% cover, 4 = > 50 to 75% cover, and 5 = > 75 to 100% cover (Braun-Blanquet 1972). Observations from each quadrat were used to estimate seagrass coverage as a BB density (D) score for each taxon i using the observations from all quadrats, where j is the quadrat number from 1 to n, and Sij is the BB score for each taxon i in quadrat j for each sampling site (Eq. 1; Fourqurean and Rutten 2004).
Seagrasses were also monitored at 15 permanent transects across FB by the FHAP program and at 5 permanent transects in FB by FCE-LTER program (Table 1). Seagrasses within FKNMS were monitored at 30 permanent transects by the WQPP monitoring program since 1995, with an additional 10 sites added in 2012 (Table 1). Transect sites across these three monitoring programs were sampled in the same manner. At each transect, divers stretched a 50-m transect tape between two permanent site markers (located with either permanent steel rods hammered into the bottom at the site or DGPS coordinates). At 10 pre-determined random distances along each transect, a 0.25-m2 quadrat was placed on the benthos and all taxa were assigned BB cover-abundance and scores as above, and a density score was calculated for each transect site (Eq. 1). FKNMS transect sites re-sampled in fall 2017 were assessed at the same ten locations along the transect that was sampled in summer 2017, using the methods described above.
The passage of both Hurricane Georges (in 1998) and Hurricane Irma (in 2017) over our study area provided a unique opportunity to examine similarities in storm effects across a single seascape. We plotted seagrass density across three of the FKNMS sites strongly impacted by Hurricane Georges, sites 216, 243, and 309 (Fourqurean and Rutten 2003), and then, we examined both their post-Georges recovery trajectories and their impacts from Irma.
Statistical Methods
Classification Tree Model
To simplify our analysis of the benthic macrophyte data, we chose to focus on the density of two functional groups of macrophyte species important to stable coverage patterns in South Florida: total seagrass (TSG) and total calcifying macroalgae (TCAL). The former combined six of the seven seagrass species found in South Florida: T. testudinum, S. filiforme, H. wrightii, Halophila decipiens, Halophila engelmannii, and Ruppia maritima and, the latter, the calcifying green macroalgal genera common to the region: Halimeda, Penicillus, Udotea, Rhipocephalus, Acetabularia, Neomeris, and Cymopolia. TCAL was recorded by divers in all monitoring programs. TSG was recorded by divers in FHAP monitoring, but not in FCE-LTER or WQPP monitoring, where only species-level data were collected. Because of the broad cover categories assigned to taxa using the Braun-Blanquet method and the mixed canopies of seagrass meadows, it was not possible to accurately estimate TSG by simply summing the scores for each seagrass taxon identified within a quadrat (i.e., they are not generally additive). Hence, we converted species-level seagrass density scores to an aggregated TSG score for the FCE-LTER and WQPP datasets, using a classification tree model (‘rpart’ package; R Development Core Team 2012). We trained the model using the FHAP spatial sampling dataset for which both species-level and TSG scores were collected, tested it using the FHAP transect dataset for which TSG data were also available, then applied the model to the FCE-LTER and WQPP seagrass species scores to generate a TSG score for each site. Prior to testing and implementation, the tree was pruned to the simplest model within one standard error of the lowest cross-validated error.
Short-Term Spatial Patterns in Macrophyte Density
We examined the immediate changes in TSG and TCAL density by calculating the change in density (ΔD; Eq. 2), where D is the density before (spring/summer 2017) or after (fall 2017) for taxon i (Fourqurean and Rutten 2004).
We calculated the ΔD in TSG and TCAL across each spatial zone for FKNMS and FB, divided ΔD by Di(before) to calculate fractional change in TSG and TCAL density, and assessed spatial patterns by mapping the results with inverse-distance weighted interpolations within R (power = 2, minimum neighbors = 12, search radius = infinite), restricting each interpolation to either north/west or south/east of the Florida Keys island chain (i.e., ‘oceanside’ or ‘bayside’), using the ‘gstat’ package (R Development Core Team 2012). We also tested whether the hypothesis that more shallow-rooted TCAL was more susceptible to removal than more deeply rooted T. testudinum using a Sign Test for two-sample paired data to test for differences in ΔD of T. testudinum and ΔD of TCAL by site (‘DescTools’ package; R Development Core Team 2012).
Long-Term Spatiotemporal Patterns in Macrophyte Density
It has been well established that seagrasses in South Florida follow distinct temporal patterns in production and abundance, with peak abundance occurring in the summer months prior to fall partial senescence (Fourqurean et al. 2001). As Irma passed during the period of fall senescence in South Florida seagrasses, we were concerned that simple pre-/post-storm change in macrophyte density could be influenced by this seasonal change as well as hurricane impacts. Because of differences in sampling frequency across monitoring programs, we performed a non-parametric repeated-measures analysis of longitudinal data to examine changes in TSG and TCAL density (hereafter referred to as ΔTSG and ΔTCAL, respectively) across all datasets (n = 13 FHAP spatial basins, n = 15 FHAP transects, n = 2 FCE-LTER transects, and n = 38 WQPP transects; Fig. 1). Two transect sites from the 40 WQPP sites and three transect sites from the 5 FCE-LTER sites were not included in our analyses, as they were either co-located with FHAP transect sites, or because their sampling frequency did not match that of the other FB sites.
For our longitudinal analysis, we constrained analyses to the 5 years prior to the storm (2012–2017), as there have been some long-term, multi-decadal changes in macrophyte density at several of the sites. Each transect site was grouped into one of 11 zones (dictated by differences in water quality; Table 2; Fig. 1; Klein and Orlando 1994; Briceño et al. 2013). We averaged density scores for TSG and TCAL for each observation unit (transect site or FHAP spatial basin) at each time point, defined as fall (T1) and spring (T0) for each year from 2012 to 2017. We then calculated ΔTSG and ΔTCAL as the difference between successive time points (T1-T0), yielding 6 time intervals. Post-Hurricane Irma fall 2017 monitoring data were available at all sites, but the lack of fall monitoring data from 2012 to 2016 at FHAP spatial sites and WQPP transect sites necessitated a different approach for calculating time intervals. For these sites, we assigned a time point during spring/summer from 2012 to 2017 (e.g., T0 = spring/summer 2012, T1 = spring/summer 2013, etc.), and a final fall 2017, post-hurricane time point, providing a total of 7 time points. We again averaged TSG/TCAL density scores for each observation unit (spatial basin or transect site) and defined time intervals as the difference between successive time points (T1-T0), which also yielded 6 time intervals.
ΔTSG and ΔTCAL were then analyzed for the single and interactive effects of spatial zone (transect sites) or spatial basin (FHAP spatial sites) and time interval using non-parametric analysis of longitudinal data (i.e., repeated measures) implemented by the ‘nparLD’ package (R Development Core Team 2012). For spatial zones or basins in which the post-Irma interval was the lowest relative treatment effect in the time-series, we further explored significant interactions identified by the Wald test statistic (Noguchi et al. 2012) using pair-wise contrasts between the post-Irma interval and the time interval with the next lowest relative treatment effect. All p values from post hoc pairwise contrasts were adjusted using Bonferroni correction. To avoid the confounding effects of the 2015 T. testudinum mortality event, we did not consider ΔTSG or ΔTCAL scores including die-off data (spring/summer 2015 to spring/summer 2016 interval at FB sites).
The above approach compares post-Hurricane (and potentially seasonal) density changes with (1) recent seasonal density changes, for FHAP transect and FCE-LTER transect sites, and (2) recent annual density changes, for WQPP transect and FHAP spatial sites. Recent or historical fall data was not available from the FHAP spatial monitoring program, so no further analyses were performed at FHAP spatial sites to assess potential seasonality effects. However, since WQPP monitoring data were collected four times per year from 1996 to 2011, we were able to assess hurricane impacts against regular seasonality for each FKNMS spatial zone using an autoregressive sinusoidal modeling approach. To create a sinusoidal model for TSG or TCAL in each spatial zone, we fit a linear model to the data, then calculated amplitude (α) and phase angle (Φ) to generate Eq. 3, where y is the TSG or TCAL density, ŷ is the intercept, and x is the day of year in radians (see Electronic Supplementary Material for a more detailed explanation of methods).
Note that this model expands on Fourqurean et al. (2001) by the addition of a non-stationary yearly mean (ŷ) that was calculated as the annual mean TSG or TCAL for each calendar year, to remove interannual differences in abundance and help better define the annual expected amplitude (α). Finally, we checked post-Irma TSG/TCAL for fall 2017 to determine whether observed in situ effects were within prediction intervals of the sinusoidal model, which did not include data from fall 2017. We thus predicted fall 2017 TSG/TCAL values and attributed observed in situ fall 2017 TSG/TCAL scores below the prediction interval as significant, and indicative of true hurricane impacts beyond anticipated seasonal changes. We also performed this sinusoidal analysis on FCE-LTER sites, which were sampled at a higher frequency (average n = 6 times per year). We examined residual plots and performed Chi-square tests of independence in order to ensure that no assumptions were violated.
Finally, we ran a series of data simulations using the FHAP transects where seasonal differences in mean density score (ΔTSG and ΔTCAL, as above) observed for Irma (spring/summer versus fall, 2017) were assessed relative to a distribution of differences (spring/summer versus fall, 2012–2017) generated by randomizing quadrats within transect and season, resampled with replacement for a total of 5000 iterations. Statistical significance was determined by proportion of times the actual pre-/post-Irma difference was greater than or equal to the simulated distribution. Results were generated for each transect, with transects assessed individually rather than by spatial zone.
Results
Short-Term Spatial Patterns in Macrophyte Density
Instantaneous losses in TSG density occurred across sites throughout the study area, but was most notable in the Lower Keys near where Hurricane Irma made landfall (Fig. 2a). We also noted instantaneous losses in TCAL density across our study area, particularly in the Lower Keys as well as areas in northern FB (Fig. 2b). Overall, greater short-term losses were observed for TSG than for TCAL. However, at sites where changes in both Thalassia testudinum and TCAL were assessed, there were greater decreases in TCAL than T. testudinum (sign test for paired samples, median difference in changes = 0.146, df = 68, p < 0.05).

Spatial pattern in short-term response for total seagrass (TSG) (a) and total calcifying green macroalgae (TCAL) (b) across the Florida Keys National Marine Sanctuary (FKNMS) and Florida Bay (FB)
Seagrasses at nine sites throughout FKNMS experienced moderate burial, with 5–10 cm of sand deposited atop the benthos (Fig. 3a, b). Another six FKNMS sites had signs of heavy erosion, and two FKNMS sites experienced moderate canopy thinning, with leaves sheared from still-buried rhizomes (Fig. 3c, d). Evidence of direct mechanical impacts within FB was minimal. In FB, areas with exposed seagrass rhizomes were noted for Johnson Key, Rankin Lake, Whipray and Manatee basins, with a comparably thick mud layer found in Rankin Lake (B. Furman, personal observation); however, parsing these effects from those of the 2015 die-off is difficult.

Photos of moderate (5–10 cm) T. testudinum seagrass burial after Hurricane Irma at FKNMS site 255 (a) and site 239 (b). Note how sediments separate leaves of the same short shoot, and ligules (leaf bases) are buried below a layer of sediment. Photo of strong seagrass erosion of T. testudinum after Hurricane Irma at FKNMS site 276 (c). Photo of erosion (background) and canopy thinning (foreground) of T. testudinum at FKNMS site 294 (d). Note that only the short shoot on the left still has leaves attached. Prior to storm-induced erosion, sediments covered the horizontal and vertical rhizomes up to the ligule, indicating 5–10 cm of erosion occurred at this location
Long-Term Spatiotemporal Patterns in Macrophyte Density
In FB, there was a significant effect of basin (WTS = 32.28, df = 12, p < 0.01), time (WTS = 60.93, df = 5, p < 0.0001), and their interaction (WTS = 726.38, df = 60, p < 0.0001) on ΔTSG at FHAP spatial sites. Post hoc analyses with Bonferroni correction for each spatial basin tested whether ΔTSG from the pre-/post-Irma interval was significantly different than the lowest ΔTSG across the rest of the time intervals. ΔTSG in the pre-/post-Irma time interval was lower than previous annual changes for only one basin, Manatee Bay, and indicative of a hurricane impact (p < 0.05; Fig. 4a inset). There were also significant effects of basin (WTS = 60.27, df = 12, p < 0.0001), time (WTS = 127.06, df = 5, p < 0.0001), and their interaction (WTS = 396.36, df = 60, p < 0.0001) on ΔTCAL in FB. Bonferroni corrected pairwise tests revealed that ΔTCAL in the pre-/post-Irma time interval was not significantly lower than previous annual changes in any basin, and hence not indicative of a true hurricane impact (Fig. 4b inset).

Results from longitudinal analysis by spatial zone for Florida Bay (FB) and the Florida Keys National Marine Sanctuary. Red zones indicate significant hurricane impacts. Inset shows results from longitudinal analysis by spatial basin in FB. Results are shown for total seagrass (TSG) (a) and total calcifying green macroalgae (TCAL) (b)
At FHAP and FCE-LTER transect sites, there was a significant effect of the interaction of time and zone on ΔTSG (WTS = 66.04, df = 11, p < 0.0001). Pre-/post-Irma ΔTSG showed the greatest decreases in North Florida Bay (NFB; p < 0.05; Fig. 4a), consistent with results found using spatial sampling data across FB basins above. There was also a significant effect of the interaction of time and zone on ΔTCAL (WTS = 21.81, df = 10, p < 0.05); however, none of the seasonal differences in ΔTCAL that were associated with Irma were significantly different from historical patterns (Fig. 4b).
At WQPP sites in the FKNMS, there was a significant effect of time (WTS = 14.14, df = 5, p < 0.05) and a significant interaction of time and zone (WTS = 56.43, df = 20, p < 0.001) on ΔTSG, and post hoc analyses identified significant hurricane impacts for ΔTSG pre-/post-Irma in the Lower Keys Bayside (LKB; p < 0.05; Fig. 4a). There was a significant interaction of time and zone for ΔTCAL in FKNMS (WTS = 219.18, df = 20, p < 0.0001). Bonferroni corrected pairwise tests did not reveal a hurricane impact beyond that of natural variability for ΔTCAL in FKNMS (Fig. 4b).
The sinusoidal models that we generated were reasonably good descriptors of the 22-year datasets for each FKNMS zone (p < 0.05 for all except TCAL in the Middle Keys Oceanside, MKO, which was marginally significant at p = 0.056) and FCE-LTER transect site (p < 0.05 for all except TSG at TC and TCAL at SB, BA, and DK, likely due to generally low amounts of TSG/TCAL present at the site). The calculated amplitude (α) for sinusoidal models ranged from 0.096 to 0.270 (μ = 0.171) for TSG and from 0.058 to 0.203 (μ = 0.108) for TCAL in FKNMS, indicating greater annual changes in TSG density relative to TCAL density, which was not surprising given that TSG density was on average greater than TCAL density across the FKNMS. The same was true for FCE-LTER calculated amplitudes, which ranged from 0.183 to 0.307 (μ = 0.207) for TSG and from 0.015 to 0.165 (μ = 0.078) for TCAL. The phase angles (Φ) ranged from 3.763 to 5.134 (μ = 4.484) for TSG and from 3.406 to 4.640 (μ = 3.899) for TCAL in FKNMS, and from 2.951 to 4.035 (μ = 3.536) for TSG and from 2.820 to 5.638 (μ = 4.278) for TCAL at FCE-LTER sites in FB. When the phase angle was converted back to day of year, the models indicated that peaks in both TSG and TCAL across South Florida occurred from August to September. Conversely, minimum densities occurred during February to March. The observed in situ post-Irma fall 2017 TSG and TCAL values were within the prediction interval for all FKNMS zones, except for TSG in Lower Keys Bayside (LKB; Figs. 5 and 6). The predicted fall 2017 TSG in the Lower Keys Bayside was 2.477 ± 0.410, and the observed TSG value was 1.279, indicating a hurricane impact. All of the FCE-LTER transect observed in situ post-Irma fall 2017 TSG/TCAL values were within the prediction interval for each site, and not indicative of a hurricane impact.

Total seagrass (TSG) density (a) and total calcareous green macroalgae (TCAL) density (b) from 1996 to 2017 in the Lower Keys Bayside (LKB spatial zone of Florida Keys National Marine Sanctuary (FKNMS). Points and solid black line indicate TSG/TCAL Density modeled from in situ Braun-Blanquet (BB) cover-abundance scores. Dashed green line indicates sinusoidal model, with confidence intervals (95%) shown as dark green ribbon and prediction intervals (95%) shown as light green ribbon. Right-most circular point indicates predicted TSG/TCAL for fall 2017 (post-Irma) time point given by sinusoidal model. Star indicates observed fall 2017 TSG/TCAL estimated from in situ BB scores (TSG = 1.279, TCAL = 0.290). The observed fall 2017 TSG falls outside of the prediction interval and is indicative of a significant hurricane impact

Results from quadrat simulation analyses by transect site for Florida Bay (FB) and from the sinusoidal models by spatial zone for the Florida Keys National Marine Sanctuary (FKNMS). Red zones and transects indicate significant hurricane impacts. Results are shown for total seagrass (TSG) (a) and total calcifying green macroalgae (TCAL) (b)
Quadrat-level seasonal simulations from FHAP transect sites in FB revealed that Hurricane Irma impacts were at least as great or greater than regular seasonal changes (p < 0.05 for all) for ΔTSG across five FB transects (BLK, JOE, LIL, LM, and TER; p < 0.05 for all; Fig. 6a). Seasonal simulations for ΔTCAL in FB were significant for three transects (BLK, MAN, and RAN; p < 0.05 for all; Fig. 6b). Interestingly, transects that indicated hurricane impacts were clustered in northern areas of FB.
Our analysis of seagrass density at FKNMS sites strongly impacted by Hurricane Georges in 1998 (sites 216, 243, and 309) showed distinct recovery trajectories at each site (Fourqurean and Rutten 2003; Fig. 7). Site 216, a moderately dense S. filiforme community with sparse T. testudinum and H. wrightii, showed signs of recovery beginning in 2013 and appeared to have fully recovered by 2016 (Fig. 7a). Site 243, a mixed community of T. testudinum and S. filiforme in moderate abundance, was completely denuded by Hurricane Georges, but recovered a few years later (Fig. 7b). Site 309 was characterized by moderately dense T. testudinum communities with sparse S. filiforme and has not yet recovered (Fig. 7c).

Recovery trajectories for FKNMS sites 216 (a), 243 (b), and 309 (c). Black circles are T. testudinum, red triangles are S. filiforme density, blue squares are H. wrightii, and green diamonds are total calcifying green macroalgae (TCAL). The y-axis is macrophyte density, or the average BB score for the taxa during each sampling visit, and the x-axis is year. Vertical dashed lines in gray signify the dates of Hurricane Georges’ and Irma’s passage. Each of these three sites were strongly impacted by Hurricane Georges in 1998, and 216 and 243 were again strongly impacted by Hurricane Irma in 2017 (site 309 never recovered from Georges and hence did not register impacts from Irma)
Discussion
Changes in Benthic Macrophyte Abundance and Community Composition
Despite the trajectory and vast wind-field of Hurricane Irma, significant impacts to the seagrass meadows of Florida Keys and Florida Bay were limited. While we are certain that there were small-scale sediment erosional and depositional events distributed throughout the seascape, at the spatial resolution of our long-term sampling, statistically significant effects to macrophyte densities were restricted to the north of Lower Keys and to the northern coastal basins of FB (Figs. 4 and 6). Although some transect sites within FB showed significant hurricane impacts using quadrat-level seasonal simulation (BLK, JOE, LIL, and TER; Fig. 6), the only spatial zone that registered significant declines using the longitudinal analysis was Northern Florida Bay (NFB; Fig. 4a), consistent with basin-level results identifying Manatee Basin for TSG impacts (Fig. 4a). Results from the analyses support our detection of isolated reductions to seagrass and macroalgae abundance that were greater than the normal seasonal decreases associated with the onset of autumn. Every hurricane is unique; some bring stronger winds (e.g., Hurricane Irma, 2017) whereas others produce more rainfall (e.g., Hurricane Harvey, 2017), but wind speed, fetch, and rainfall also vary across the path of any given storm. This study provides an example of a single storm yielding both direct and indirect impacts to seagrasses in very different (and not all) areas of the same system.
In the Florida Keys, there was a significant decline of seagrass within the Lower Keys Bayside (LKB) but not within the Lower Keys Oceanside (LKO), even though the eye of the storm passed through both areas. The Lower Keys Bayside is on average much shallower (2.3 m depth) than the Lower Keys Oceanside (5.6 m depth) and is characterized by a complex mosaic of small keys (islands). This bathymetry likely led to a high degree of channelization and funneling of storm surge as the storm passed, which could have caused seagrass loss via erosion as flow and shear stress increased. While we do not have direct measurements of water flow to support hypothesized erosion, the effects of storm surge were quite obvious as documented by diver surveys and digital photos (Fig. 3c, d). On the other hand, Lower Keys Oceanside sites were likely protected from wave energy, fetch and current velocities by the offshore reef tract. Many studies have demonstrated the importance of coral reefs in buffering and dissipating wind-waves (Lowe et al. 2005; Ferrario et al. 2014; Narayan et al. 2016). For example, reefs have been shown to reduce wave energy by up to 97% (Ferrario et al. 2014) and wave height by up to 70% (Narayan et al. 2016). However, it must be noted that if coral reefs in the Florida Keys continue to decline as they have in recent years (Porter et al. 2001; Palandro et al. 2008; McClenachan et al. 2017), inshore seagrass meadows may not be afforded the current degree of protection from storm waves.
At the time of the fall 2017 sampling, despite the minimal evidence of direct impacts in FB, acute indirect impacts associated with low salinity (Figs. 45 and 47 in Sculley 2018) and dissolved oxygen (FWC and ENP unpublished data) were already apparent in Manatee Bay and to a lesser degree in Blackwater Sound, where T. testudinum appeared to have recently died (i.e., ‘standing dead’ as described by Hall et al. 2016). Emergency management actions implemented by the South Florida Water Management District as a response to record-high rainfall in June 2017 and to additional rainfall associated with Hurricane Irma resulted in the discharge of 207,000 acre-feet of fresh water through the S-197 control structure into Manatee Bay and Barnes Sound of FB (Sculley 2018). Salinities in Manatee Bay remained quite low (< 15) for at least 1 month after Hurricane Irma, with benthic salinities below 5 PSU in October 2017 (Figs. 45 and 47 within Sculley 2018).
Given the short time (~ 1 month) between the storm and our initial re-sampling, and the belowground carbon reserves of species implicated (T. testudinum), the indirect effects observed in FB were not likely induced by shading from a turbid water column (FWC-FWRI, personal observations); light stress was, however, was a factor after Hurricane Frances in the Big Bend region of Florida, and after two floods and a cyclone in Hervey Bay, Australia (Preen et al. 1995; Carlson et al. 2010). Similarly, after our post-Irma monitoring, algal blooms with TSS/CDOM-related turbidity subsequently formed and have remained a chronic, albeit spatiotemporally variable threat in parts of western FB for at least a year following the storm (SFWMD, unpublished data). At present, the long-term fate of these blooms is not known and, although early signs of seagrass recovery have been subsequently noted (unpublished data, FWC-FWRI), additional seagrass losses due to light limitation remain possible.
Interestingly, we found no significant region-wide hurricane impacts on the calcareous macroalgal members of the seagrass communities across broad spatial scales (basins and zones) compared to annual fluctuations within the previous 5 years, although three transect stations in FB did indicate localized effects (Fig. 6b). Earlier work in FKNMS documented greater losses of total macroalgal density after a hurricane compared to seagrass density, attributed to the difference in sediment anchoring capabilities of macroalgae versus seagrasses (Fourqurean and Rutten 2004). It was shown that the type of anchoring structure, rather than anchoring depth, played an important role in Caribbean macroalgae susceptibility to hurricane-simulated erosional disturbance in Mexico (Cruz-Palacios and Tussenbroek 2005). In sites with both T. testudinum and calcareous green macroalgae, TCAL did show greater density decrease than the deeply rooted T. testudinum (sign test for paired samples).
Subsequent monitoring efforts have shown that FKNMS sites with moderate sediment deposition (5–10 cm; Fig. 3a, b) after Hurricane Irma have fared well (S. Wilson, personal observation). Evidently, burial was minimal enough to maintain adequate amounts of photosynthetic material for seagrasses to continue growth. Previous experimental work on T. testudinum burial to simulate hurricane effects found mixed results, with plants experiencing a significant reduction (71%) in density after being buried 10 cm, but showing no significant reductions in density after 7.5 cm of burial in a different experimental site in Mexico (Cruz-Palacios and Tussenbroek 2005). In both instances, burial depth was 75% of the height of T. testudinum leaves; in situ observations at sites with sediment deposits from Irma in FKNMS did not show this magnitude of burial (e.g., Fig. 3a, b). However, large amounts of lateral sediment transport and seagrass burial from Hurricane Irma were documented in Culebra, Puerto Rico (Hernández-Delgado et al., this issue), along with erosional seagrass meadow scars and seagrass burial by displaced coral reef rubble. Subsequent monitoring has shown that eroded sites within FKNMS have not yet recovered (S. Wilson, personal observation). Thus, for Hurricane Irma, the direct impact of partial burial was not as lethal for FKNMS seagrasses as was erosion.
Specific Areas of Benthic Macrophyte Vulnerability in South Florida
The passage of both Hurricane Georges (in 1998) and Hurricane Irma (in 2017) over our study area provided a unique opportunity to examine similarities in storm effects across a single seascape. Two sites strongly damaged by erosion during Hurricane Georges in 1998 (sites 216 and 243; Fourqurean and Rutten 2004) were impacted again by erosion during Hurricane Irma. Another site, 309, which sustained major damage from sediment burial (up to 50 cm) during Hurricane Georges (Fourqurean and Rutten 2004), had not yet recovered prior to Irma’s passage. Monitoring data collected since Georges indicated that each of these three sites followed a unique recovery trajectory. Interestingly, sites 216 and 243 were strongly affected after both Georges and Irma, highlighting the degree to which bathymetric features, especially offshore coral reefs, govern hurricane impacts in nearshore seagrass meadows. These two sites are both adjacent to openings in the reef tract and may represent areas of particular vulnerability to sediment destabilization and erosion from high wave energy or strong currents (Fourqurean and Rutten 2004).
Overall, the passage of Hurricane Irma does not appear to have led to catastrophic seagrass losses initially feared in FB and FKNMS, rather, impacts from the storm were highly localized (i.e., limited to a few stations). To some degree, this contrasts the reporting of local news stories that described large mats of seagrass leaves either floating or beached along the shorelines of Everglades National Park (https://www.miamiherald.com/news/weather/hurricane/article173637481.html). We believe that there are several likely explanations for the massive quantity of seagrass wrack deposited post-storm. First, the hurricane occurred during the normal period of seasonal senescence for seagrasses in South Florida (Fourqurean et al. 2001), so we would expect that any detrital leaves or those in the process of senescence would have been transported to the oceanside shoreline of the Florida Keys and the southern shoreline of the Florida mainland. Second, shorelines in the southern Everglades likely got deposits of seagrass material from both inside and outside the National Park and the FKNMS, as evidenced by large amounts of S. filiforme, the leaves of which are both easily broken by mechanical stresses and transported great distances because of their buoyancy. Finally, a large seagrass die-off occurred in several areas of FB during the summer of 2015, resulting in a large amount of dead seagrass material within the system (Hall et al. 2016). It is possible that much of the refractory detrital tissue remaining from the die-off was then deposited onto the Everglades shoreline during the storm. A previous study in South Florida noted similarly vast deposits of T. testudinum detrital leaf material, nearly 1.5 million kg, deposited along Biscayne Bay shorelines after Hurricane Donna in 1960, but reported no severe impacts to local seagrass meadows (Thomas et al. 1961).
Lessons Learned from Hurricane Irma and Multi-Agency Coordinated Seagrass Monitoring Programs
The results from this study offer insight to how other coastal macrophyte communities may respond after major hurricane or disturbance events. Our findings after Hurricane Irma suggest that coastal areas hit by a storm with heavy winds will be more likely to sustain direct physical impacts. Seagrasses in other systems with offshore barrier islands or coral reefs may be affected in a manner similar to what we observed after Hurricane Irma, with meadows adjacent to unprotected offshore areas experiencing stronger impacts than sheltered meadows behind adjacent islands or reefs (Fourqurean and Rutten 2003). Storm trajectory and associated wind direction will ultimately control which areas benefit from protection. On the other hand, estuarine areas with longer residence times will be more likely to experience seagrass losses related to stormwater runoff and retention. The spatial pattern of indirect impacts documented in this study suggest that our understanding and management of watershed hydrology will be critical for mitigating effects of future storm events (Carlson et al. 2010). Thus, seagrass monitoring programs should carefully consider up-estuary influences when selecting sampling locations and parameters.
Beyond simple impact description, coordinated post-storm monitoring offers insight into the organization of benthic macrophyte communities, with implications for altered community trajectories related to tropical perturbation. While we found that seagrass losses were generally more common than those to macroalgae (e.g., Fig. 2), our short-term pre-/post-storm analysis showed that decreases in TCAL density were greater than decreases in T. testudinum density. This supports the findings of Fourqurean and Rutten (2004) within the FKNMS after Hurricane Georges, who found that calcareous green macroalgae densities decreased by 24%, whereas the seagrass species S. filiforme and T. testudinum decreased by only 19% and 3%, respectively; however, neither the previous study nor our short-term spatial analyses took seasonality into account, and our long-term studies only indicated significant TCAL declines across three transects with respect to recent ecological baselines (Fig. 6b). Furthermore, by combining all seagrass species into one category (TSG) for our analyses, we cannot parse species-level effects; however, we can infer that hurricane damage depends in part on the anchoring morphology of the macrophyte community in question (Fourqurean and Rutten 2004; Cruz-Palacios and Tussenbroek 2005; this study).
As vulnerable taxa are removed by hurricanes or other disturbances, shifts in post-disturbance community structure will be dictated by the frequency and severity of disturbances, as well as the resulting species composition and the specific life history traits of the resident species. Additionally, the presence or absence of species nearby to disturbed areas and the local nutrient regime can also impact community structure dynamics (Fourqurean and Rutten 2003). In our system, the replacement of T. testudinum by earlier colonizers such as calcareous green macroalgae or H. wrightii in the coastal basins of FB will likely reinforce the species diversity, holding the system at an earlier successional stage; conversely, loss of TCAL from sites containing seagrasses (e.g., Fig. 6b) will initially reduce species richness and could provide an advantage to the slower-growing T. testudinum (Williams 1990; Gallegos et al. 1994; Fourqurean and Rutten 2003).
Finally, we were fortunate to have access to a rich record of seagrass monitoring data for the region. Despite differences in spatial and temporal resolution between monitoring programs, because all data were collected in a similar manner, a holistic examination of Irma impacts was possible by applying the same statistical test across all stations (non-parametric repeated-measures analysis of longitudinal data). Even where discrepancies in sampling frequency were an issue, comparable seasonal analyses were able to augment the results of our longitudinal analysis. Likewise, small differences in the reporting of functional groups between the two monitoring programs were easily overcome with simple machine learning techniques (Classification Tree Model trained with local data). Thus, the collaborative efforts leveraged by this study demonstrate the value of long-term environmental monitoring programs with event-response capabilities (Fourqurean et al. 2002). Because long-term baseline data were available, our investigation could place disturbance impacts within the context of recent ecological condition or system state, and the long-term data also helped to inform our decisions on what an appropriate baseline for our analysis should be. For example, we knew from previous monitoring that seasonality is an important control on macrophyte density within our system (Fourqurean et al. 2001), so we utilized fall and spring data from recent years (whenever available) to construct our spatiotemporal models and incorporate this critical seasonal component into our analyses. Undoubtedly, similar long-term and spatially expansive seagrass monitoring programs will be vital for understanding and predicting impacts imposed by future tropical weather events. Creating and maintaining monitoring programs with consistent coordinated protocols therefore allows managers to have effective event-response teams in place.
References
Anton, A., J. Cebrian, C.M. Duarte, K.L. Heck, and J. Goff. 2009. Low impact of hurricane Katrina on seagrass community structure and functioning in the northern Gulf of Mexico. Bulletin of Marine Science 85: 45–59.
Ball, M.M., E.A. Shinn, and K.W. Stockman. 1967. The geologic effects of hurricane Donna in South Florida. The Journal of Geology 75 (5): 583–597.
Braun-Blanquet, J. 1972. Plant sociology: The study of plant communities. New York: Hafner Publishing Company.
Briceño, H.O., J.N. Boyer, J. Castro, and P. Harlem. 2013. Biogeochemical classification of South Florida's estuarine and coastal waters. Marine Pollution Bulletin 75 (1-2): 187–204.
Byron, D., and K.L. Heck. 2006. Hurricane effects on seagrasses along Alabama's Gulf Coast. Estuaries and Coasts 29 (6): 939–942.
Cangialosi, J.P., A.S. Latto, and R. Berg. 2018. Hurricane Irma. Tropical Cyclone Report. National Hurricane Center, 30 September 2018. 111 pp.
Carlson, P.R., L.A. Yarbro, K.A. Kaufman, and R.A. Mattson. 2010. Vulnerability and resilience of seagrasses to hurricane and runoff impacts along Florida's west coast. Hydrobiologia 649 (1): 39–53.
Cruz-Palacios, V., and B.I.v. Tussenbroek. 2005. Simulation of hurricane-like disturbances on a Caribbean seagrass bed. Journal of Experimental Marine Biology and Ecology 324 (1): 44–60.
Dingtian, Y., and H. Daojian. 2011. Impacts of typhoons Tianying and Dawei on seagrass distribution in Xincun Bay, Hainan Province, China. Acta Oecologica Sinica 30: 32–39.
Durako, M.J., M.O. Hall, and M. Merello. 2002. Patterns of change in the seagrass dominated Florida bay Hydroscape. In The Everglades, Florida bay, and coral reefs of the Florida keys: An ecosystem source book, ed. J.W. Porter and K.G. Porter, 523–537. CRC Press.
Ferrario, F., M.W. Beck, C.D. Storlazzi, F. Micheli, C.C. Sherpard, and L. Airoldi. 2014. The effectiveness of coral reefs for coastal hazard risk reduction and adaptation. Nature Communications 5 (1): 3794.
Fourqurean, J.W., and L.M. Rutten. 2003. Competing goals of spatial and temporal resolution: Monitoring seagrass communities on a regional scale. In Monitoring ecosystems: Interdisciplinary approaches for evaluating ecoregional initiatives, ed. D.E. Busch and J.C. Trexler, 257–288. Washington, D.C.: Island Press.
Fourqurean, J.W., and L.M. Rutten. 2004. The impact of hurricane Georges on soft-bottom, back reef communities: Site- and species-specific effects in South Florida seagrass beds. Bulletin of Marine Science 75: 239–257.
Fourqurean, J.W., A. Willsie, C.D. Rose, and L.M. Rutten. 2001. Spatial and temporal pattern in seagrass community composition and productivity in South Florida. Marine Biology 138 (2): 341–354.
Fourqurean, J.W., M.J. Durako, M.O. Hall, and L.N. Hefty. 2002. Seagrass Distribution in South Florida: A Multi-Agency Coordinated Monitoring Program. In The Everglades, Florida Bay, and Coral Reefs of the Florida Keys: An Ecosystem Sourcebook, ed. J.W. Porter and K.G. 497–522. CRC Press LLC.
Gallegos, M.E., M. Merino, A. Rodriguez, N. Marbà, and C.M. Duarte. 1994. Growth patterns and demography of pioneer Caribbean seagrasses Halodule wrightii and Syringodium filiforme. Marine Ecology Progress Series 109: 99–104.
Hall, M.O., B.T. Furman, M. Merello, and M.J. Durako. 2016. Recurrence of Thalassia testudinum seagrass die-off in Florida bay, USA: Initial observations. Marine Ecology Progress Series 560: 243–249.
IPCC, 2014: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Field, C.B., V.R. Barros, D.J. Dokken, K.J. Mach, M.D. Mastrandrea, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L. White (eds).]. Cambrdige University Press, Cambridge, United Kingdom and New York, NY, USA, 1132 pp.
Klein, C.J., and S.P. Orlando. 1994. A spatial framework for water-quality management in the Florida keys national marine sanctuary. Bulletin of Marine Science 54: 1036–1044.
Lowe, R.J., J.L. Falter, M.D. Bandet, G. Pawlak, M.J. Atkinson, S.G. Monismith, and J.R. Koseff. 2005. Spectral wave dissipation over a barrier reef. Journal of Geophysical Research 110: C04001.
McClenachan, L., G. O'Connor, B.P. Neal, J.M. Pandolfi, and J.B.C. Jackson. 2017. Ghost reefs: Nautical charts document large spatial scale of coral reef loss over 240 years. Science Advances 3 (9): e1603155.
Narayan, S., M.W. Beck, B.G. Reguero, I.J. Losada, B.v. Wesenbeeck, N. Pontee, J.N. Sanchirico, J.C. Ingram, G.-M. Lange, and K.A. Burks-Copes. 2016. The effectiveness, costs and coastal protection benefits of natural and nature-based Defences. PLoS One 11 (5): e0154735.
NHC, N. 2017. Hurricane Irma public advisory 45a. National Hurricane Center, National Oceanographic and Atmospheric Administration.
Noguchi, K., Y.R. Gel, E. Brunner, and F. Konietschke. 2012. nparLD: An R software package for the nonparametric analysis of longitudinal data in factorial experiments. Journal of Statistical Software 50 (12).
Nuttle, W.K., J.W. Fourqurean, B.J. Cosby, J.C. Zieman, and M.B. Robblee. 2000. Influence of net freshwater supply on salinity in Florida bay. Water Resources Research 36 (7): 1805–1822.
Palandro, D.A., S. Andréfouët, C. Hu, P. Hallock, F.E. Müller-Karger, P. Dustan, M.K. Callahan, C. Kranenburg, and C.R. Beaver. 2008. Quantification of two decades of shallow-water coral reef habitat decline in the Florida keys National Marine Sanctuary using Landsat data (1984-2002). Remote Sensing of Environment 112 (8): 3388–3399.
Porter, J.W., P. Dustan, W.C. Jaap, K.L. Patterson, V. Kosmynin, O.W. Meier, M.E. Patterson, and M. Parsons. 2001. Patterns of spread of coral disease in the Florida keys. Hydrobiologia 460 (1/3): 1–24.
Preen, A.R., W.J.L. Long, and R.G. Coles. 1995. Flood and cyclone related loss, and partial recovery, of more than 1000 km2 of seagrass in Hervey Bay, Queensland, Australia. Aquatic Botany 52 (1-2): 3–17.
R Development Core Team. 2012. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing ISBN 3-900051-07-0, URL http://www.R-project.org/.
RAMMB, N. 2017. Aircraft-based tropical cyclone surface wind analysis. In AL112017 - major hurricane Irma: Regional and mesoscale meteorology branch. National Oceanic and Atmospheric Administration.
Robblee, M.B., T.R. Barber, P.R. Carlson, M.J. Durako, J.W. Fourqurean, L.K. Muehlstein, D. Porter, L.A. Yarbro, R.T. Zieman, and J.C. Zieman. 1991. Mass mortality of the tropical seagrass Thalassia testudinum in Florida bay (USA). Marine Ecology Progress Series 71: 297–299.
Sculley, S. 2018. After action report: Emergency measures due to high water conditions in the South Florida region during the 2017 wet season. Vol. 81. South Florida Water Management District.
Sierra-Rozo, O., B. Gavio, and J.E. Mancera-Pineda. 2012. Structure of Thalassia testudinum beds in old Providence island, Caribbean Colombian, after hurricane Beta. Caldasia 34: 155–164.
Thomas, L.P., D.R. Moore, and R.C. Work. 1961. Effects of hurricane Donna on the turtle grass beds of Biscayne Bay, Florida. Bulletin of Marine Science of the Gulf and Caribbean 11: 191–197.
Tilmant, J.T., R.W. Curry, R. Jones, A. Szmant, J.C. Zieman, M. Flora, M.B. Robblee, D. Smith, R.W. Snow, and H. Wanless. 1994. Hurricane Andrew's effects on marine resources. BioScience 44 (4): 230–237.
Tussenbroek, B.I.v. 1994. The impact of hurricane Gilbert on the vegetative development of Thalassia testudinum in Puerto Morelos coral reef lagoon, Mexico: A retrospective study. Botanica Marina 37: 421–428.
Webster, P.J., G.J. Holland, J.A. Curry, and H.-R. Chang. 2005. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 309 (5742): 1844–1846.
Williams, S.L. 1990. Experimental studies of Caribbean seagrass bed development. Ecological Monographs 60 (4): 449–469.
Acknowledgments
We would like to thank Christian Lopes, Kevin Cunniff, Rachel Lopez, Sonja Smith, Claudia Carrion, Brittany Nguyen, Manuel Merello, Donna O’Dell, Katie Toth, Mike Wheeler, and Robin Jung for field assistance. We are grateful to Everglades National Park and Mote Marine Laboratory for logistical support. We appreciate the comments from editors and three anonymous reviewers, which improved the manuscript greatly.
Funding
Funding for Florida Keys National Marine Sanctuary’s Water Quality Protection Program seagrass monitoring was provided by Environmental Protection Agency contract #X7-00D39015. Funding for Florida Bay seagrass monitoring was provided by the National Science Foundation through the Florida Coastal Everglades Long-Term Ecological Research Program under Grant No. DEB-1237517, and the South Florida Water Management District under the Restoration, Coordination and Verification Program. This is contribution # ___ from the Center for Coastal Oceans Research in the Institute of Water and Environment at Florida International University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Melisa C. Wong
Electronic supplementary material
ESM 1
(DOCX 41 kb)
Rights and permissions
About this article

Cite this article
Wilson, S.S., Furman, B.T., Hall, M.O. et al. Assessment of Hurricane Irma Impacts on South Florida Seagrass Communities Using Long-Term Monitoring Programs. Estuaries and Coasts 43, 1119–1132 (2020). https://doi.org/10.1007/s12237-019-00623-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-019-00623-0