
Abstract
Predicting the responses of animals to environmental changes is a fundamental goal of ecology and is necessary for conservation and management of species. While most studies focus on relatively gradual changes, extreme events may have lasting impacts on populations. Animals respond to major disturbances such as hurricanes by seeking shelter, migrating, or they may fail to respond appropriately. We assessed the effects of Hurricane Irma in 2017 on the behavior and survival of juvenile bull sharks (Carcharhinus leucas) within a nursery of the Florida coastal Everglades using long-term acoustic telemetry monitoring. Most of our tagged sharks (n = 14) attempted to leave the shallow waters of the Shark River Estuary before the hurricane strike, but individuals varied in the timing and success of their movements. Eight bull sharks left within hours or days before the hurricane, but three left more than a week in advance. Nine of 11 bull sharks (~ 82%) eventually returned to the array within weeks or months of the storm. Six of these returning individuals were detected in a different coastal array in nearshore waters ca. 80 km away from the mouth of the estuary during their absence. The remaining three bull sharks moved downstream relatively late (after the hurricane) and may have died. We used binomial generalized linear mixed models to estimate the probability of presence within the array as a function of several environmental variables. Departure from the array was predicted by declining barometric pressure, increasing rate of change in pressure, and potentially fluctuations in river stage. Juvenile bull sharks may weigh multiple environmental cues, perceived predation risk, their own physical size, and shifting prey resources when making decisions during and after hurricanes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Organisms use many information sources to assess the quality or deterioration of a habitat and react to spatiotemporal variation environmental conditions. Appropriate responses, which may be learned or innate, are sometimes critical to survival and reproduction (Breuner et al. 1982; Streby et al. 2015). Generally, as conditions deteriorate, animals are expected to make spatial shifts into better conditions. For instance, many avian species rely on circannual rhythms and associated photoperiods to time migration and avoid a seasonal decline in food (Gwinner 1996). Seasonal migrations for fish and birds are well-studied, but behavioral responses and evacuation in response to sudden major disturbances and extreme events is a less investigated phenomenon (Bailey and Secor 2016).
While a diverse literature shows that natural selection has honed behavioral responses, most studies have focused on relatively gradual or somewhat predictable variation in ecological and environmental conditions (Bailey and Secor 2016; Donihue et al. 2018). More poorly understood is whether, when, and how animals respond to extreme environmental changes that may occur infrequently relative to the life history characteristics of some species (Lytle and Poff 2004). Such extreme events, like hurricanes or earthquakes, could have profound impacts on individual fitness or population sizes and may represent a strong enough selective pressure to optimize animal responses to such events. For instance, Hurricanes Irma and Maria in 2017 induced phenotypic changes in a population of Caribbean island lizards (Anolis scriptus) through natural selection on clinging capacity, which varies across morphological traits (Donihue et al. 2018).
Hurricanes and storms can bring about sudden and catastrophic physical and ecological changes to coastal and estuarine ecosystems. These disturbances can damage physical structure, impact hydrological conditions, alter biogeochemical processes, and directly affect vegetation and animal populations (Michener et al. 1997). Hurricanes can cause excessive precipitation, high wind, and oceanic storm surge which may in turn increase turbidity, increase discharge, alter salinity regimes, and modify the delivery of nutrients and sediment to coastal riverine systems (Michener et al. 1997). For animal populations, deteriorating environmental conditions can decrease foraging success, impact energy use, and cause mortality (Breuner et al. 1982).
A strategy by animals to mitigate risks associated with storms is to use cues to predict approaching disturbances and subsequently alter behavior to increase survival chances and limit disruption to foraging. A cue thought to be used by many animals to detect approaching dangerous weather is declining barometric pressure (Heupel et al. 2003; Breuner et al. 1982). Bats, birds, and sharks all have a proposed mechanism sensitive enough for detecting drops in barometric pressure associated with a storm (Paige 1995; Breuner et al. 1982; Udyawer et al. 2013). For example, highly sensitive vestibular hair cells in the elasmobranch inner ear can detect changes in hydrostatic pressure that would indicate an approaching hurricane (Fraser and Shelmerdine 2002). Elasmobranchs may also be able to detect other hurricane-associated environmental changes such as altered river stage, decreases in water temperature (Brown 2003), and changes in salinity (Heupel and Simpfendorfer 2008; Heithaus et al. 2009). Nonetheless, hydrological variables are well-supported drivers of movement behaviors for aquatic animals (e.g., Luschi et al. 2003; Matich and Heithaus 2014; Grammer et al. 2015) but are understudied as cues for detecting storms (Bailey and Secor 2016). Beyond detection, animals also change behavior as a result of low-pressure systems. For instance, amphibians increase vocalization (Oseen and Wassersug 2002) and birds increase food intake (Breuner et al. 1982) with approaching storms.
The movements of juvenile bull sharks (Carcharhinus leucas) have been studied in the Shark River Estuary of Everglades National Park, USA, using passive acoustic telemetry since 2007 (Matich and Heithaus 2012, 2014, and 2015). This monitoring was ongoing when Hurricane Irma passed over the study site in September 2017, which allowed us to investigate the behaviors of bull sharks before, during, and after the storm and to investigate potential environmental cues that triggered behavioral changes. The objectives of this paper are to (1) investigate the cues bull sharks use to sense and respond to an impending storm; (2) determine the factors that impact the timing of return to behaviors similar to those found before the storm; (3) assess whether demographic factors (i.e., age and sex) influenced behavioral responses; and (4) establish if the hurricane affected habitat use patterns after the storm.
Methods
Study Area and Species
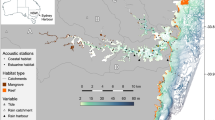
The Shark River Estuary (SRE) of Everglades National Park, USA (Fig. 1), serves as one of the main conduits for freshwater from the Everglades to drain into the Gulf of Mexico (Rudnick et al. 1999). The SRE is a braided tidal waterway, bordered by mangroves (primarily red mangroves, Rhizophora mangle), extending from the Gulf of Mexico to where it transitions into freshwater marshes in the upper river nearly 30 km away (Childers 2006). Heavier rainfall during the wet season (May–October) leads to lower salinity in the estuary relative to the dry season (November–April) (Romigh et al. 2006; Rosenblatt and Heithaus 2011). The estuary is oligotrophic and limited by marine phosphorous input, with greater productivity at the mouth (Childers 2006). Since 2007, the SRE has been the site of long-term studies of large predators including juvenile bull sharks that use the estuary as a nursery (e.g., Rosenblatt and Heithaus 2011, Matich and Heithaus 2015).

Configuration of an acoustic array in the Shark River Estuary, Florida, USA. Black and green dots denote one of 37 receivers. Black dots indicate that a receiver was deployed throughout the entire study period whereas green dots reveal receivers that were pulled just before the hurricane and redeployed just after due to their vulnerability. The blue triangle shows the Gunboat Island site where hydrological data were collected, and barometric pressure was estimated. The inset in the upper left corner displays the state of Florida. The study site is shown as a red rectangle. The track of Hurricane Irma and its intensity is also presented in the inset as it was at its closest (ca. 60 km) to the Shark River mainstem at 15:00 UTC 10 September 2017
Bull sharks are abundant, apex predators that spend their first 3 or 4 years of life in estuarine waters near nursery areas (Wiley and Simpfendorfer 2007; Heithaus et al. 2009; Castro 2011). Within nursery habitats, food availability is sufficient for growth, and risk of predation from larger sharks is low compared to marine waters where food availability is higher (Heupel et al. 2007; Heithaus et al. 2009; Matich and Heithaus 2015). As juveniles grow to larger body sizes in the SRE, they generally increase their use of higher predation risk, prey-rich marine habitats though they still use lower salinity areas to mitigate predation risk (Matich and Heithaus 2015). The SRE population of juvenile bull sharks exhibits individual specialization in movement patterns and foraging strategies, presumably as a mechanism to avoid intraspecific competition (Matich and Heithaus 2015).
In September 2017, Hurricane Irma caused widespread and catastrophic damage in the northeastern Caribbean and the Florida Keys (Cangialosi et al. 2018). Hurricane Irma was reported to be about 60 km from the Shark River main stem at 15:00 UTC 10 September 2017, hereafter referred to as “strike,” and brought high winds, high rainfall, and storm surge as a Category 4 hurricane (Saffir-Simpson Hurricane Scale). At the Royal Palm Ranger Station within Everglades National Park, total rainfall was recorded at 28.7 cm on 10 September 2017 (Cangialosi et al. 2018). Average September monthly rainfall at this weather station is about 22 cm (Southeast Regional Climate Center 2019). After passing the SRE, Hurricane Irma made its seventh and final landfall at 19:30 UTC 10 September 2017 near Marco Island, Florida, with maximum winds of 51 m s−1 and minimum pressure of 936 hPa (Cangialosi et al. 2018).
Field Methods
We captured juvenile bull sharks using 500 m longlines, fitted with 40–55 15/0 circle hooks baited with mullet (Mugil sp.), and attached by 2-m gangions of 400 kg test monofilament line (as described by Heithaus et al. 2009). Animals tracked during this study were captured in February 2016 (n = 9) and September 2016 (n = 5). We determined sex based on the presence or absence of claspers and measured pre-caudal length, fork length, and total length to the nearest centimeter for each animal. Each individual received a colored and numbered plastic tag attached through the dorsal fin. Cohort and age class of each shark was determined by size at capture following Matich and Heithaus (2015).
We surgically implanted an acoustic transmitter (V16-4x-069, Vemco, Halifax, NS, Canada) into the abdominal cavity with a mid-ventral incision and suturing. Pulse rates were set at a random interval between 60 and 90 s resulting in an estimated battery life of 1825 days. These sharks were tracked within an array of 37 receivers (VR2W, Vemco, Halifax, NS, Canada) deployed to create “gates” that allow the direction of movement to be estimated and movements into and out of major areas within the estuary to be determined (see Rosenblatt and Heithaus (2011) for more detail; Fig. 1). The receivers were secured to a PVC pipe set in concrete and submerged. Every 3 to 5 months, data were downloaded, batteries were replaced, and receivers were redeployed. Mean detection ranges of receivers were about 500 m (see Rosenblatt and Heithaus 2011). Detectability likely decreased with increased acoustic noise associated with wind and storm surge caused by the hurricane. To mitigate the influence of these impacts, we aggregated the data to hourly detections resulting in a presence-absence matrix for modeling cues. All receivers with the exception of seven coastal receivers near the river mouth remained deployed throughout the hurricane. These seven receivers were removed on 6 September 2017 and returned on 2 October 2017. The removed receivers did not affect our ability to determine presence or absence from the array and did not impact our inference of movements across habitats.
Weather and Environmental Data
Local barometric pressure (hPa) was recorded every 10 s and averaged to 1 min using a pressure transducer (Model 278; Setra, Boxborough, Massachusetts, USA) in the Lower Shark River about 4 km upriver from the mouth of the estuary (Fig. 2). Hydrological data were obtained from the United States Geological Survey/National Park Service Everglades Depth Estimation Network database (United States Geological Survey 2018). Specifically, we used river stage values (m) estimated at Gunboat Island about 10 km upriver from the mouth of the estuary (Fig. 2). Also, at Gunboat Island, we used a multiparameter sonde (EXO 2; Xylem, Yellow Springs, Ohio, USA) to record water temperature (°C), salinity (psu), and dissolved oxygen (mg/L) values every 10 min (Fig. 3). Due to malfunction, we were not able to obtain values for dissolved oxygen from mid-October through mid-November 2017.

Weather (observed barometric pressure in Lower Shark River) and hydrologic conditions (observed river stage at Gunboat Island) in the Shark River Estuary, Florida, USA. Red dotted line denotes the estimated time Hurricane Irma was reported to be at its closest (ca. 60 km) to the Shark River mainstem at 15:00 UTC 10 September 2017

Water conditions observed at Gunboat Island in the Shark River Estuary, Florida, USA. Red dotted line denotes the estimated time Hurricane Irma was reported to be at its closest (ca. 60 km) to the Shark River mainstem at 15:00 UTC 10 September 2017. The gap in values for dissolved oxygen is a result of probe malfunction
Statistical Analysis
To investigate the cues used by sharks to leave the estuary, an hourly binary presence-absence matrix was built by individual. A binomial generalized linear mixed model with a logit link function was used to estimate the probability of presence (i.e., detection) within the array as a function of the environmental variables. Bull shark identification number was a random effect in all models. Fixed effects included river stage, barometric pressure, and hourly change in each variable. We tested correlation among explanatory variables to avoid issues with multicollinearity and subsequently calculated variance inflation factors (Zurr et al. 2009). We only modeled the departures of eight animals that left within the hurricane’s window of impact on Shark River (< 7 days before the strike). The other six animals did not leave the estuary or left too early in the absence of changes in our measured environmental variables. We used data starting at 63 h (2.5 days) before Hurricane Irma’s closest position and 33 h after (10 h after the last animal left). All covariates were standardized due to orders of magnitude differences in means (mean = 0, standard deviation (SD) = 1). We included a null model and global model as well as all combinations of variables. We used Akaike’s Information Criterion corrected for small sample sizes (AICC) to rank models and selected competing model(s) where AICC was less than two units from the most supported model (Burnham and Anderson 2002). In addition, we used Akaike weights (wi) to perform model averaging and reported full model averaged coefficients and 95% confidence intervals with shrinkage (Burnham and Anderson 2002).
We built several contingency tables to test the effects of demographics (sex and cohort) on categorical behavioral responses to the storm and used Fisher’s exact test for testing independence, which is appropriate for small sample sizes. Sharks were caught at two different times of the year. Therefore, we did not use individual size measurements in our tests and instead used inferred age and cohort. We used nonparametric methods to determine the influence of demographic variables on movement and response parameters. Specifically, we used time absent from the array in days, time at which the animal left the array, time of return, and time spent in a second array (see below) as our response variable in separate tests. We used the Wilcoxon rank sum test with continuity correction when comparing the responses between males and females as well as the 2014 and 2015 cohorts due to low sample sizes in each group.
We calculated mean river distance from the mouth hourly for each shark which allowed us to investigate potential changes in patterns of habitat use in the estuary as a result of the hurricane. We also used ± 1 SD from mean river distance as a proxy for space use revealing how much of the river length was used by an individual, hereafter “linear distance range.” For spatial analyses, we used 60 days before each shark left for the storm while discarding the three preceding days as a buffer to avoid direct storm-related movement. We also used 60 days after the time of first return to the system though again discarding the first 3 days of return. We only did these analyses for the nine sharks that left and returned. As a “control” timeframe without a hurricane, we also calculated both mean distances and linear distance ranges for each shark in the corresponding timeframe in 2016. We used a paired two-sample Wilcoxon test to compare mean river distance and linear distance range before and after the storm.
We performed statistical analyses in R (Mac version 3.4.3; R Foundation for Statistical Computing; Vienna, Austria). Lastly, we reported means with ± 1 standard deviation (SD) and evaluated significance at α = 0.05.
Results
In total, we tracked 14 animals from 2016 to 2018 (Table 1). Eleven individuals were detected leaving the array anywhere from 17 days before the strike to 1 day after the hurricane arrived (mean = 4.2 ± 6.3 days; Figs. 4 and 5; see Electronic Supplementary Material Fig. A1 for example animal tracks). Nine of these 11 animals returned to the array 55.3 ± 33.3 days (min = 16.5, max = 98.6) after the eye of the hurricane passed. For these nine, the total time sharks spent absent from the array was 57.0 ± 32.9 days (min = 16.3, max = 100.0). Two animals left the array and were still absent as of early July 2018, and these individuals left nearly a week ahead of all other sharks (Table 1). Six of the individuals that returned were detected at another coastal acoustic array nearly 80 km straight line distance north while they were absent from SRE. These six animals were detected a total of 21 ± 18 days (min = 3, max = 41) in the Ten Thousand Islands and Faka Union Bay array.

Detections of acoustically tagged juvenile bull sharks (Carcharhinus leucas) within a telemetry array in the Shark River Estuary (SRE), Florida, USA. Each dot represents a daily detection. Black dots represent detections within SRE. Green dots depict detections in another coastal array in nearshore waters ca. 80 km away from the mouth of SRE. Red dotted line denotes the estimated time Hurricane Irma was reported to be at its closest (ca. 60 km) to the Shark River mainstem at 15:00 UTC 10 September 2017. The plot does not include three sharks that suffered potential storm-related mortality. Note that sharks × 56136 and × 56137 still remained absent as of July 2018

Hourly proportion of juvenile bull sharks (Carcharhinus leucas) within our tagged sample that were detected within an acoustic telemetry array in the Shark River Estuary, Florida, USA. Red dotted line denotes the estimated time Hurricane Irma was reported to be at its closest (ca. 60 km) to the Shark River mainstem at 15:00 UTC 10 September 2017. The plot does not include three sharks that suffered potential storm-related mortality
The remaining three individuals did not successfully leave the system. For two sharks, we observed quick movement downstream shortly after the storm and then consistent hourly detections at a single station for the duration of the study period (months) indicating mortality (see Knip et al. 2012; see Electronic Supplementary Material Fig. A2). These animals traveled 5 km downstream from Tarpon Bay towards Gunboat Island within 30 and 21 min (10.0 and 14.3 km h−1) on the morning of 14 September and the night of 12 September, respectively. Another individual did not leave the estuary and may have died as well indicated by a loss of detections at a midstream receiver soon after the hurricane on 16 September. We are unsure what conditions or factors may have led to these potential mortalities.
We modeled the probability of presence in the array as a function of several environmental variables. Barometric pressure and river stage had moderate correlation (− 0.43), and the rates of change with their respective variables had negligible correlations. However, variance inflation factors were below 1.3 for each variable in the competing models indicating that multicollinearity is not a concern (Zurr et al. 2009). Of all of the candidate models, only two exhibited ΔAICc < 2 (Table 2). The lowest AICc model had pressure and hourly change in pressure as predicting flight from the nursery (Table 2). For the averaged model, increasing pressure had a positive effect on presence, increasing change in pressure had a negative effect on presence, and the effect of stage may be negligible (Fig. 6; Table 3).

Effects of environmental conditions on predicted probability of juvenile bull sharks (Carcharhinus leucas) being detected within an acoustic telemetry array in the Shark River Estuary, Florida, USA, around Hurricane Irma in September 2017. The predictions come from the averaged generalized linear mixed model with Shark ID as a random effect and each plotted variable. River stage was also included in the model; however, the confidence interval for the beta estimate overlapped 0 thus we are unable to ascertain its effect on presence
We tracked six juvenile males, which all left the array and returned. Of the eight female juveniles we tracked, three died, three left and returned, and two left and did not return. We did not detect dependence of behavioral response based on sex (Fisher’s exact test; p = 0.08). The mean rank of time elapsed between the last detection and the hurricane (i.e., timing of evacuation) did not differ between males and females (Wilcoxon rank sum test; W = 15, p = 1.00). Also, neither the mean rank of time elapsed between the hurricane and return nor the mean rank of total time absent from the array differed between males and females (Wilcoxon rank sum test; W = 9, p = 1.00; W = 8, p = 0.90, respectively). Of the sharks that left, three males and three females were detected in the Ten Thousand Islands and Faka Union Bay array (Fisher’s exact test; p = 1.00). The mean rank of time spent in the Ten Thousand Islands and Faka Union Bay array did not differ between males and females (Wilcoxon rank sum test; W = 3.5, p = 0.82).
The three youngest bull sharks (2016 cohort) in our sample were not detected leaving the estuary and appeared to have died. Thus, survival varied among cohorts (Fisher’s exact test; p = 0.03). The two sharks that left and did not return were in the 2015 cohort while all sharks in the 2013 and 2014 cohorts left and returned (Fisher’s exact test; p = 0.67). The mean rank of time elapsed between the last detection and the hurricane did not differ between the 2014 and 2015 cohorts (Wilcoxon rank sum test; W = 7, p = 0.31). Neither did the mean rank of time elapsed between the hurricane and return nor the mean rank of total time absent from the array (Wilcoxon rank sum test; W = 5, p = 0.57; W = 5, p = 0.57, respectively). We removed the single individual from the 2013 in these analyses. Of the bull sharks that left, the individual from the 2013 cohort (n = 1), two from the 2014 cohort (n = 5), and three from the 2015 cohort (n = 6) were detected in the Ten Thousand Islands and Faka Union Bay array.
Sharks’ use of habitats within the nursery area changed once they returned. The mean distance from river mouth was higher during the 60 days before leaving (12.0 ± 4.9 km) compared to the 60 days after returning (3.0 ± 1.5 km) (Fig. 7) (paired Wilcoxon test; V = 45, p = 0.002). However, mean distance before the storm in 2017 was smaller than the same time frame in 2016 (17.3 ± 2.2 km) (paired Wilcoxon test; V = 0, p = 0.008). The use of downstream habitats after the storm in 2017 was greater than the same time frame in 2016 (17.9 ± 3.4 km) (paired Wilcoxon test; V = 0, p = 0.002). Habitat use did not differ in 2016 during the timeframes corresponding to the 2017 hurricane (paired Wilcoxon test; V = 18, p = 0.58). Bull sharks suffering mortality used more upstream habitats before the storm (17.4 ± 0.5 km) than those that survived and returned (Wilcoxon rank sum test; W = 3, p = 0.03). The river linear distance range size was not different in the 60 days before leaving (3.7 ± 1.1 km) compared to the 60 days after returning (2.6 ± 1.2 km) (Fig. 8) (paired Wilcoxon test; V = 38, p = 0.07). Also, linear distance range size after the storm in 2017 did not differ from that during the same time frame in 2016 (2.5 ± 1.6 km) (paired Wilcoxon test; V = 25, p = 0.82). There was no difference in linear distance range size before the 2017 hurricane and the same time frame in 2016 (2.4 ± 1.9 km) (paired Wilcoxon test; V = 25, p = 0.08). Linear distance ranges did not differ in 2016 during the timeframes corresponding to the 2017 hurricane (paired Wilcoxon test; V = 19, p = 0.47). When sharks returned, mid-estuary salinities remained lower than before the storm (mean of 3.7 ± 4.4 psu for 8/23–9/6/2017 vs. 2.5 ± 2.6 psu for 9/14–28/2017; t test; t = 4.2, df = 544, p < 0.0001).

Box plot of 60-day distance from river mouth of acoustically tagged juvenile bull sharks (Carcharhinus leucas) before and after Hurricane Irma on 10 September 2017 in the Shark River Estuary, Florida, USA

Box plot of 60-day river linear distance range size of acoustically tagged juvenile bull sharks (Carcharhinus leucas) before and after Hurricane Irma on 10 September 2017 in the Shark River Estuary, Florida, USA
Discussion
How Do Marine Animals Respond to Hurricanes?
We found that bull sharks left the estuary and exhibited highly directed movements towards the coast before the onset of the largest impacts of the hurricane, but the timing and success of these movements varied among individuals. Flight responses towards marine waters as a result of tropical storms and hurricanes have been documented in several coastal elasmobranch species including pigeye sharks (C. amboinensis; Udyawer et al. 2013), blacktip sharks (C. limbatus; Heupel et al. 2003; Udyawer et al. 2013), spottail sharks (C. sorrah; Udyawer et al. 2013), and Australian blacktip sharks (C. tilstoni; Udyawer et al. 2013). Changes in movement behavior to avoid tropical cyclones have also been observed in diverse estuarine and marine taxa. For instance, American lobsters (Homarus americanus) in the Great Bay Estuary of New Hampshire, USA, were detected moving downstream towards deeper marine waters after Hurricane Bob in 1991 (Jury et al. 1995). Another study revealed that sea kraits (Laticauda spp.) used barometric pressure cues to leave the rough surf of the open littoral zone to seek refuge in volcanic cavernous rocks during a typhoon in 2009 that impacted Lanyu, Taiwan (Liu et al. 2010). Our findings along with studies of diverse taxa suggest that avoidance behaviors of tropical cyclones are common responses for mobile marine fauna. Future studies may be able to synthesize information across species to determine what factors (e.g., life history traits, intraspecific variation, habitat characteristics, etc.) influence these behaviors.
What Cues Are Used to Respond?
Our tagged bull sharks may have used multiple cues in deciding when to leave the estuary upon the approach of the storm. Studies report that thresholds in declining barometric pressure may trigger sharks to take flight, and response appears to be species-specific with catalysts as low as 1001.9 hPa for C. sorrah and as high as 1007.4 hPa for C. limbatus (Heupel et al. 2003; Udyawer et al. 2013). In our work, decreasing pressure predicted flight from the estuary, where a threshold of 1005 hPa resulted in 50% of the sharks leaving. Standard sea level pressure is 1013.25 hPa and readings often drop to 1010 hPa over the Shark River Estuary during thunderstorms associated with low-pressure systems (National Oceanic and Atmospheric Administration 2018). We have not, however, detected tagged sharks departing the estuary during these events. Though the barometric pressure dropped to 999 hPa over the Shark River with Hurricane Matthew on 6 October 2016 measured near Miami International Airport (National Oceanic and Atmospheric Administration 2018), no tagged shark exhibited a noticeable change in movement patterns or left the estuary. Hurricane Matthew remained over the Atlantic Ocean, and hurricane-force winds never occurred within our study area. Declining pressure itself does not pose a threat to bull sharks but is correlated with potentially dangerous conditions brought on by changes in water level or winds.
Variation in behavioral responses to low-pressure systems has led researchers to speculate that the rate of decline in barometric pressure may be the driver rather than there being an absolute threshold value at which flight is initiated (Heupel et al. 2003; Udyawer et al. 2013). Both absolute pressure and rate of change in pressure appeared in our lowest AICc models. Increasing hourly change in pressure predicted flight from the estuary and a drop in 1 hPa/h resulted in 50% of the bull sharks leaving. This is surprising given that some thunderstorms associated with low-pressure systems result in changes of more than 1 hPa/h (National Oceanic and Atmospheric Administration 2018). This may mean that the rate of change is placed into context of an absolute value of pressure, where at lower absolute values the same rate of decline may matter more. However, further research is needed to unravel the complexities of behavioral responses to signal thresholds and rate of change.
Sharks may also have used other cues to sense the approaching storm. In our second lowest AICc model, along with pressure and change in pressure, river stage was included as a predictor. However, the effect of stage on presence is uncertain due to the confidence interval overlapping 0 for the beta estimate. River stage or other hydrological variables have not been investigated as cues for flight during extreme conditions for coastal elasmobranchs, but they have been found to be important for predicting evacuations by large-bodied teleost fishes in coastal river systems (Grammer et al. 2015; Bailey and Secor 2016). Our results should be taken in context of the difficulty in disentangling the complexities of multiple environmental parameters changing along with a major environmental disturbance. Future studies may have larger sample sizes or be able to employ controlled experiments to isolate variables and measure behavioral responses.
Flight behavior exhibited by elasmobranchs in response to extreme events may be an innate response (Heupel et al. 2003; Udyawer et al. 2013). Our study supports this hypothesis given that we saw consistent directed movements out of the estuary and return by most bull sharks despite never experiencing a major storm or hurricane within their lifetimes. Given the life history characteristics of bull sharks, even infrequent extreme events may be an adequate selective pressure to result in behaviors tied to appropriate and non-fatal responses. Future research needs to expand our understanding of the link between perception and response and the evolutionary, ecological, and behavioral consequences of large-scale disturbances such as hurricanes (Van de Pol et al. 2017).
What Cues Are Used to Return?
Return times to the Shark River were variable among individuals, which is similar to other studies of elasmobranchs (Heupel et al. 2003; Udyawer et al. 2013). We did not find any support that cohort or sex influenced these decisions. Individual sharks may use different cues to know when to return (Udyawer et al. 2013), or individuals may have traveled different distances at different speeds away from the Shark River in response to the storm. We did not explicitly model predictors of return due to low sample size of returning animals. Five of our tagged individuals that left and returned to SRE were detected during their absence at another coastal acoustic array nearly 80 km straight line distance north. The animals have never been detected within that array and stayed variable lengths of time. It is unclear why these individuals chose to make this trek north and what determined their length of stay. We present these findings as a minimum distance traveled; the actual extent of movement is not known due to limited receiver coverage in the Gulf of Mexico.
Estuaries exhibit changes in salinity regimes after hurricanes due to storm surge, impacts on tidal cycles, and increased upstream rainfall (Kelble et al. 2007). During relatively stable conditions in the SRE, salinity was not identified as a primary driver of distribution for juvenile bull sharks (Heithaus et al. 2009). This contrasts with other systems which exhibit more rapid changes in salinity due to human water management decisions (Ortega et al. 2009). Post-storm hyposaline conditions lasting for almost 2 weeks in another Florida estuary are thought to have prevented juvenile blacktip reef sharks from returning to their nursery until salinities returned to pre-storm levels (Heupel et al. 2003). Juvenile bull sharks seasonally experience low salinities in the SRE and take advantage of seasonal pulses of prey entering channels from the marsh that are brought about by changes in precipitation and freshwater flow (Matich and Heithaus 2014). In general, despite a small rise due to storm surge, SRE had lower than average salinity for weeks after the hurricane due to increased rainwater and freshwater inflow (Fig. 3). Even though the hurricane resulted in swifter and more prolonged changes to salinity regimes than the Shark River typically experiences, salinity does not appear to be a primary cue given that salinity levels remained low even after bull sharks returned to the estuary.
Hurricanes and storms may drastically reduce the availability of oxygen for aquatic organisms by mud suffocation and decomposition of organic material (Tabb and Jones 1962). In north Florida Bay after Hurricane Donna in 1960, fish and invertebrate kills resulted from oxygen depletion, but recolonization led to community recovery and healthy sport fish populations within one to several months (Tabb and Jones 1962). For juvenile bull sharks using the estuary during normal conditions, dissolved oxygen was identified as an important determinant of habitat use (Heithaus et al. 2009). At Gunboat Island, dissolved oxygen levels spiked, and remained > 6 mg/L for hours before and after the strike of Hurricane Irma, then dropped, remaining below 1 mg/L for 2 weeks after the hurricane (Fig. 3). No bull shark returned until just after this time. From 2016 to 2019 in the Shark River Estuary, 157 total bull sharks were caught in waters above 3 mg/L dissolved oxygen (5.0 ± 1.1 mg/L) with 1 individual caught at 2.8 mg/L (B.A. Strickland, unpublished data). Low dissolved oxygen may have inhibited the return of bull sharks due to the difficulty of oxygen exchange or the effects of low oxygen on prey availability. However, we are unsure if or how bull sharks would be able to discern the levels in the estuary in advance of being detected at the river mouth.
Did Demographics or Individual Variation Influence Behavior?
Hurricane-related mortality has not been reported for elasmobranchs (Heupel et al. 2003; Udyawer et al. 2013), but there are many examples of hurricane-induced mass mortality across other taxa (sponges, Wulff 1995; birds, Wiley and Wunderle 1993; primates, Pavelka et al. 2007). Three of the youngest bull sharks in our sample appear to have suffered storm-related mortality. Two individuals made directed movements downstream, but these movements were after the storm and hours or days later than most of the other older bull sharks. We can only speculate what caused mortality, but storm surge may have impeded the ability to swim and rising water levels may have increased structure and debris in the channel. Indeed, two of the bull sharks sustained speeds of 10 and 14 km h−1 for 30 and 21 min, respectively, which is likely too fast a swimming speed for sharks to travel without aid from high downstream water discharge (see Lowe 1996). Alternatively, hypoxic conditions in the estuary immediately after the storm may have been a factor. Dissolved oxygen levels in the estuary dropped below 1 mg/L on the night of September 12 corresponding to the time when the first bull shark left and the dissolved oxygen levels remained low for 2 weeks. In addition, the three bull sharks suffering mortality used more upstream habitats before the storm than those that survived and returned. This indicates that general habitat use and position in the estuary before the storm may influence evacuation behavior.
We observed consistent directed movements out of the estuary within a relatively short, but variable, time frame before the hurricane. This contrasts with studies in other locations (Heupel et al. 2003; Udyawer et al. 2013). For instance, in Terra Ceia Bay (Florida, USA), all 13 tagged juvenile blacktip sharks left within a short time window of 1.7–5.5 h before landfall of Tropical Storm Gabrielle in 2001 (Heupel et al. 2003). Not counting the three bull sharks that left over a week in advance of the hurricane, our remaining eight bull sharks left 0.8 ± 1.1 days before the hurricane. Early departures over 1 week before a storm have not been reported for elasmobranchs. Juvenile bull sharks frequently move downstream as they age, but they do not move outside the nursery until they are about 3 to 4 years of age and emigrate permanently (Matich and Heithaus 2012; Matich and Heithaus 2015). Most emigrations by juvenile bull sharks in SRE appear to happen in May to September (Matich and Heithaus 2015). Two of the bull sharks that left in late August well before the storm (and also were the only ones in our tagged sample that did not return) were 2 years old, and the other individual was 3 years old. The 2-year-old bull sharks were likely too young to permanently emigrate out of the system, though rare early emigrations have been reported (Matich and Heithaus 2015). None of the bull sharks tracked during this study had ever made a detectable excursion out of the estuary before the hurricane since their tagging in February and September 2016. Despite early emigrations being unusual, these animals may have left the nursery without knowledge of the approaching storm.
Did the Hurricane Influence Habitat Use?
Juvenile bull sharks that returned to the SRE used more downstream habitats after their return than before the 2017 hurricane and more than any time in 2016. Older and larger juveniles generally increase their use of prey-rich marine habitats while remaining estuarine residents (Matich and Heithaus 2015). Previous studies found that 1- and 2-year-old bull sharks exhibit similar use of habitats spending about 25% of their time downriver, 25% midstream in Shark River, and 45% in Tarpon Bay (Matich and Heithaus 2015). Bull sharks at 3 years old began to use downriver (15%) and Tarpon Bay (40%) slightly less while using Shark River more at nearly 50% of their time (Matich and Heithaus 2015). In our sample that left and returned in the 2017 pupping season, three of our bull sharks transitioned from 1 to 2 years of age, five transitioned from 2 to 3 years, and one went from 3 to 4 years of age. These ontogenetic changes in habitat use describe a move towards more downstream use as bull sharks age, but it is difficult to know if it accounts for the observed dramatic shift to the use of habitats near the mouth of the Shark River after Hurricane Irma. During high inflow periods, such as after a hurricane, estuarine and freshwater fish may actively or passively move downstream (Flannery et al. 2002). A downstream shift in normal food availability may alternatively explain changes in habitat use patterns of our juvenile bull sharks. Future work using dietary bio-tracers on bull sharks caught after the hurricane may be able to reveal if there were changes in foraging patterns. We did not observe differences in linear distance ranges in relation to the hurricane event. This indicates that bull sharks did not change the size of the space used only the location.
Conclusion
Evacuations are common responses to extreme events but may be dependent on behavioral and environmental constraints (Bailey and Secor 2016). Juvenile bull sharks may weigh multiple environmental cues, perceived predation risk, their own physical size, and shifting prey resources when making decisions before, during, and after a hurricane. The ability of a population or demographic to respond conditionally to a disturbance may provide a selective advantage over other groups that are less mobile or more constrained (Bailey and Secor 2016). With predicted changes in the frequency and intensity of extreme weather (Hegerl et al. 2011), future events may have important impacts on survival and population growth of species relying on estuaries (Michener et al. 1997). Our findings, along with other studies, indicate that coastal elasmobranch species vary in their responses and tolerances to changing conditions resulting from extreme events (Heupel et al. 2003; Matich and Heithaus 2012; Udyawer et al. 2013). In light of the variation we observed in behavioral responses and fates of bull sharks to a major hurricane, including differences driven by age or size, we encourage future work on its implications on population dynamics. Our work highlights the utility of long-term multi-faceted datasets and their ability to provide insights into individual behavioral aspects and a species’ life history.
References
Bailey, H., and D.H. Secor. 2016. Coastal evacuations by fish during extreme weather events. Nature 6: 30280.
Breuner, C.W., R.S. Sprague, S.H. Patterson, and H.A. Woods. 1982. Environment, behavior, and physiology: do birds use barometric pressure to predict storms? The Journal of Experimental Biology 216: 1982–1990.
Brown, B.R. 2003. Sensing temperature without ion channels. Nature 421: 492.
Burnham, K.P., and D.R. Anderson. 2002. Model Selection and Multimodel Inference: A Practical Information-theoretic Approach. 2nd ed. New York: Springer-Verlag.
Cangialosi, J. P., A. S. Latto, R. J. Berg. 2018. Hurricane Irma (AL112017). Tropical Cyclone Report: National Hurricane Center. https://www.nhc.noaa.gov/data/tcr/AL112017_Irma.pdf. Accessed 13 September 2018.
Castro, J.I. 2011. The sharks of North America. New York: Oxford University Press.
Childers, D.L. 2006. A synthesis of long-term research by the Florida Coastal Everglades LTER Program. Hydrobiologia 569 (1): 531–544.
Donihue, C.M., A. Herrel, A.C. Fabre, A. Kamath, A.J. Geneva, T.W. Schoener, J.J. Kolbe, and J.B. Losos. 2018. Hurricane-induced selection on the morphology of an island lizard. Nature 560 (7716): 88–91. https://doi.org/10.1038/s41586-018-0352-3.
Flannery, M.S., E.B. Peebles, and R.T. Montgomery. 2002. A percent- of-flow approach for managing reductions of freshwater inflows from unimpounded rivers to southwest Florida estuaries. Estuaries 25 (6): 1318–1332. https://doi.org/10.1007/BF02692227.
Fraser, P.J., and R.L. Shelmerdine. 2002. Dogfish hair cells sense hydrostatic pressure. Nature 415 (6871): 495–496.
Grammer, P.O., P.F. Mickle, M.S. Peterson, J.M. Havrylkoff, W.T. Slack, and R.T. Leaf. 2015. Activity patterns of Gulf Sturgeon (Acipenser oxyrinchus desotoi) in the staging area of the Pascagoula River during fall outmigration. Ecology of Freshwater Fish 24 (4): 553–561.
Gwinner, E. 1996. Circannual clocks in avian reproduction and migration. Ibis 138: 47–63.
Hegerl, G.C., H. Hanlon, and C. Beierkuhnlein. 2011. Climate science: elusive extremes. Nature Geoscience 4: 142–143.
Heithaus, M.R., B.K. Delius, A.J. Wirsing, and M.M. Dunphy-Daly. 2009. Physical factors influencing the distribution of a top predator in a subtropical oligotrophic estuary. Limnology and Oceanography 54 (2): 472–482. https://doi.org/10.4319/lo.2009.54.2.0472.
Heupel, M.R., J.K. Carlson, and C.A. Simpfendorfer. 2007. Shark nursery areas: concepts, definition, characterization and assumptions. Marine Ecology Progress Series 337: 287–297.
Heupel, M.R., and C.A. Simpfendorfer. 2008. Movement and distribution of young bull sharks Carcharhinus leucas in a variable estuarine environment. Aquatic Biology 1: 277–289.
Heupel, M.R., C.A. Simpfendorfer, and R.E. Hueter. 2003. Running before the storm: Blacktip sharks respond to falling barometric pressure associated with tropical storm Gabrielle. Journal of Fish Biology 63 (5): 1357–1363.
Jury, S.H., W.H. Howell, and W.H. Watson. 1995. Lobster movements in response to a hurricane. Marine Ecology Progress Series 119: 305–310.
Kelble, C.R., E.M. Johns, W.K. Nuttle, T.N. Lee, R.H. Smith, and P.B. Ortner. 2007. Salinity patterns of Florida Bay. Estuarine, Coastal, and Shelf Science 71 (1-2): 318–334.
Knip, D.M., M.R. Heupel, and C.A. Simpfendorfer. 2012. Mortality rates for two shark species occupying a shared coastal environment. Fisheries Research 125–126: 184–189.
Lowe, C.G. 1996. Kinematics and critical swimming speed of juvenile scalloped hammerhead sharks. The Journal of Experimental Biology 199 (Pt 12): 2605–2610.
Liu, Y.L., H.B. Lillywhite, and M.C. Tu. 2010. Sea snakes anticipate tropical cyclone. Marine Biology 157 (11): 2369–2373.
Luschi, P., G.C. Hays, and F. Papi. 2003. A review of long-distance movements by marine turtles, and the possible role of ocean currents. Oikos 103: 293–302.
Lytle, D.A., and N. LeRoy Poff. 2004. Adaptation to natural flow regimes. Trends in Ecology and Evolution 19 (2): 94–100.
Matich, P., and M.R. Heithaus. 2012. Effects of an extreme temperature event on the behavior and age structure of an estuarine top predator (Carcharhinus leucas). Marine Ecology Progress Series 447: 165–178. https://doi.org/10.3354/meps09497.
Matich, P., and M.R. Heithaus. 2014. Multi-tissue stable isotope analysis and acoustic telemetry reveal seasonal variability in the trophic interactions of juvenile bull sharks in a coastal estuary. Journal of Animal Ecology 83 (1): 199–213. https://doi.org/10.1111/1365-2656.12106.
Matich, P., and M.R. Heithaus. 2015. Individual variation in ontogenetic niche shifts in habitat use and movement patterns of a large estuarine predator (Carcharhinus leucas). Oecologia 178 (2): 347–359. https://doi.org/10.1007/s00442-015-3253-2.
Michener, W.K., E.R. Blood, K.L. Bildstein, M.M. Brinson, and L.R. Gardner. 1997. Climate change, hurricanes, and tropical storms, and rising sea level in coastal wetlands. Ecological Applications 7 (3): 770–801.
National Oceanic and Atmospheric Administration. 2018. Automated Surface Observing System: Miami International Airport Station Data. https://www.ncdc.noaa.gov/data-access/land-based-station-data/land-based-datasets/automated-surface-observing-system-asos. Accessed 13 September 2018.
Ortega, L.A., M.R. Heupel, P.V. Beynen, and P.J. Motta. 2009. Movement patterns and water quality preferences of juvenile bull sharks (Carcharhinus leucas) in a Florida estuary. Environmental Biology of Fishes 84 (4): 361–373.
Oseen, K.L., and R.J. Wassersug. 2002. Environmental factors influencing calling in sympatric anurans. Oecologia 133 (4): 616–625.
Paige, K.N. 1995. Bats and barometric pressure – conserving limited energy and tracking insects from the roost. Functional Ecology 9 (3): 463–467.
Pavelka, M.S.M., K. McGoogan, and T.S. Steffens. 2007. Population size and characteristics of Alouatta pigra before and after a major hurricane. International Journal of Primatology 28 (4): 919–929.
Romigh, M.M., S.E. Davis, V.H. Rivera-Monroy, and R.R. Twilley. 2006. Flux of organic carbon in a riverine mangrove wetland in the Florida coastal Everglades. Hydrobiologia 569 (1): 505–516.
Rosenblatt, A.E., and M.R. Heithaus. 2011. Does variation in movement tactics and trophic interactions among American alligators create habitat linkages? The Journal of Animal Ecology 80 (4): 786–798.
Rudnick, D.T., Z. Chen, D.L. Childers, and T.D. Fontaine. 1999. Phosphorus and nitrogen inputs to Florida Bay: The importance of the Everglades watershed. Estuaries 22 (2): 398–416.
Southeast Regional Climate Center. 2019. Royal Palm Ranger Station Data, Florida (1949–2012). https://sercc.com/cgi-bin/sercc/cliMAIN.pl?fl7760. Accessed 13 June 2019.
Streby, H.M., G.R. Kramer, S.M. Peterson, J.A. Lehman, D.A. Buehler, and D.E. Andersen. 2015. Tornadic storm avoidance behavior in breeding songbirds. Current Biology 25 (1): 98–102.
Tabb, D.C., and A.C. Jones. 1962. Effect of Hurricane Donna on the aquatic fauna of north Florida Bay. Transactions of the American Fisheries Society 91 (4): 375–378.
Udyawer, V., A. Chin, D.M. Knip, C.A. Simpfendorfer, and M.R. Heupel. 2013. Variable response of coastal sharks to severe tropical storms: environmental cues and changes in space use. Marine Ecology Progress Series 480: 171–183.
United States Geological Survey. 2018. Everglades Depth Estimation Network database. https://sofia.usgs.gov/eden/. Accessed 13 September 2018.
Van de Pol, M.S. Jenouvrier, J.H.C. Cornelissen, and M.E. Visser. 2017. Behavioral, ecological, and evolutionary responses to extreme climactic events: challenges and directions. Philosophical Transactions of the Royal Society B 372 (1723).
Wiley, T.R., and C.A. Simpfendorfer. 2007. The ecology of elasmobranches occurring in the Everglades National Park, Florida: implications for conservation and management. Bulletin of Marine Science 80: 171–189.
Wiley, J.W., and J.M. Wunderle. 1993. The effects of hurricanes on birds, with special reference to Caribbean islands. Bird Conservation International 3 (4): 319–349.
Wulff, J.L. 1995. Effects of a hurricane on survival and orientation of large erect coral reef sponges. Coral Reefs 14 (1): 55–61.
Zurr, A.F., E.N. Ieno, N. Walker, A.A. Saveliev, and G.M. Smith. 2009. Mixed Effects Models and Extensions in Ecology with R. 1st ed. New York: Springer.
Acknowledgments
We are grateful to many field and lab volunteers that assisted with capture and sample processing. This is a contribution from the Center for Coastal Oceans Research in the Institute for Water and Environment at Florida International University. B. Strickland was supported by University Graduate School and Department of Biological Sciences assistantships at Florida International University as well as an Everglades Foundation fellowship. We would like to thank Yannis Papastamatiou and two anonymous reviewers for their comments and suggestions to improve the manuscript. In addition, we would like to acknowledge Javiera Hernandez, Hugh Willoughby, and Robert Burgman for their insights on hurricanes.
Funding
This work was supported by the National Science Foundation through the Florida Coastal Everglades Long-Term Ecological Research program under Grant DEB-1237517 and in collaboration with the RECOVER program of the Comprehensive Everglades Restoration Plan. Dissolved oxygen measurements were made as part of a project supported by the National Science Foundation through the Water Sustainability and Climate solicitation (EAR 1204572) and the National Aeronautics and Space Administration (NNX14AJ92G) under the Carbon Cycle Science Program.
Research and animal procedures were conducted under the auspices of protocol no. IACUC-16-022 from the Institutional Animal Care and Use Committee of Florida International University and in accordance with sampling permit no. EVER-2017-SCI-0031 granted by Everglades National Park.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Mark S. Peterson
Electronic Supplementary Material
Figure A1
Tracks of two acoustically tagged juvenile bull sharks (Carcharhinus leucas) that evacuated a telemetry array in the Shark River Estuary (SRE), Florida, USA. Each dot represents a daily detection. Red dotted line denotes the estimated time Hurricane Irma was reported to be at its closest (ca. 60 km) to the Shark River mainstem at 15:00 UTC 10 September 2017. (EPS 37 kb)
Figure A2
Tracks of two acoustically tagged juvenile bull sharks (Carcharhinus leucas) showing potential mortality within telemetry array in the Shark River Estuary (SRE), Florida, USA. Each dot represents a daily detection. Red dotted line denotes the estimated time Hurricane Irma was reported to be at its closest (ca. 60 km) to the Shark River mainstem at 15:00 UTC 10 September 2017. (EPS 38 kb)
Rights and permissions
About this article

Cite this article
Strickland, B.A., Massie, J.A., Viadero, N. et al. Movements of Juvenile Bull Sharks in Response to a Major Hurricane Within a Tropical Estuarine Nursery Area. Estuaries and Coasts 43, 1144–1157 (2020). https://doi.org/10.1007/s12237-019-00600-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-019-00600-7