
Abstract
In the nearshore Gulf of Maine, a combination of factors (overfishing, ecosystem change, and ocean warming) is thought to govern groundfish recovery. We analyzed feeding habits of demersal predatory fish from Midcoast Maine (abundant river herring) and Passamaquoddy Bay (low river herring) in eastern Maine, using stomach content and stable isotope analyses, to determine the prevalence of river herring (alewife, Alosa pseudoharengus, and blueback herring, A. aestivalis) in nearshore (<4.5 km) groundfish diets. Invertebrates dominated all predator diets at all sites. At Midcoast sites, catch-per-unit-effort (CPUE) of Atlantic cod was higher, and fish predators showed a strong seasonal pattern in river herring consumption compared to Passamaqouddy Bay. Cod, pollock (Pollachius virens), and sculpins (Myoxocephalus octodecimspinosus and M. scorpius) from Midcoast sites tended to be enriched in δ15N relative to Passamaquoddy Bay. Contrasting fast vs. slow turnover tissue (fin vs. muscle) indicated that focal species migrated or food availability changed seasonally and Atlantic mackerel (Scomber scombrus) in Passamaquoddy Bay were assimilating into a trophically depleted food web. We posit that lack of forage fish in Passamaquoddy Bay contributed to conditions that encourage an invertebrate based diet. River herring are also an order of magnitude less abundant in Passamaquoddy Bay than at Midcoast sites, limiting the availability of this seasonal food source. River restoration may contribute to recovery of groundfish stocks nearshore by increasing the availability of high lipid, seasonally available prey.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fisheries have had significant impacts on marine food webs worldwide, with the highest impacts in continental shelf and slope regions near human populations (Halpern et al. 2008; Worm et al. 2006). Fishing, as one of many potential anthropogenic drivers, has reduced biodiversity and affected marine food webs (Myers and Worm 2005; Worm et al. 2006) through trophic cascades (Pace et al. 1999), changes in community diversity (Worm et al. 2006), and serial depletion, replacement, and addition of species to the commercial harvest (Essington et al. 2006). Exploitation has increased the amount of energy concentrated in less commercially desirable components of the food web (Auster and Link 2009; Link et al. 2009), even though biomass among trophic guilds appears to remain constant (Link et al. 2009; Lotze and Milewski 2004; Nye et al. 2009).
An understanding of dietary preferences and food web relationships is useful for clarifying species roles and interactions within an ecosystem. The groundfish complex in the Gulf of Maine is a suite of species, including cod, haddock (Melanogrammus aeglefinus), redfish (Sebastes fasciatus), hakes (Urophycis sp.), and flounders (Pleuronectidae), that occupy similar habitats and show a high degree of diet overlap. Historically, the most sought after of these fish was cod. We focus on cod here, among other species, because it is an iconic fish that has declined from the core of the US colonial fishery to an overfished commercial and recreational fishery (Department of Commerce 2016).
Cod consume fish and invertebrates, including arthropods, crustaceans, gastropods, and mollusks, feeding in both benthic and pelagic habitats (Collette and Klein-MacPhee 2002; Johannesen et al. 2012; Link and Garrison 2002). Cod undergo an ontogenetic diet shift at roughly 30-cm total length (TL), switching from an invertebrate-heavy juvenile diet to an adult diet with more fish (Garrison and Link 2000; Link and Garrison 2002; Sherwood et al. 2007). Research in the 1960s and 1970s detailed diet habits of cod and other groundfish, noting that consumption of fish prey was relatively rare in Maine bays and coves, even for cod over 30 cm TL (Hacunda 1981; Macdonald and Green 1986; Tyler 1971; Tyler 1972; Willis et al. 2013). These observations were counter to historic publications that made a direct connection between anadromous fish and commercially important marine groundfish (Field 1914). Fish poor groundfish diets of the 1960s and 1970s were likely related to excessive fishing pressure in the Northwest Atlantic by US and foreign fleets at that time (Melvin and Stephenson 2007; Overholtz 2002; Serchuk and Wigley 1992).
Recent studies have explored the connection between river herring, inshore cod spawning aggregations (Ames 2004) and winter grounds (Ames and Lichter 2013), positing that river herring prey was a driver of some groundfish behavior. Stomach content analysis of groundfish from the Gulf of Maine (Garrison and Link 2000; Link et al. 2009; Smith et al. 2007) and nearshore waters of Midcoast Maine (McDermott et al. 2015) have identified river herring as likely important to groundfish feeding behavior. These authors and others (Lake et al. 2012; Saunders et al. 2006) make strong and compelling arguments for restoring anadromous fish spawning habitat, particularly through dam removal, as a means of altering the decline of groundfish species.
Stable isotope analysis (SIA) is a commonly used tool for evaluating nutrient dynamics and food web structure (Boecklen et al. 2011; Michener and Kaufman 2007; Peterson et al. 1985). The method relies on the premise that the stable carbon and nitrogen isotope composition of an organism reflects that of its diet, with some known (or estimated) isotopic fractionation (Caut et al. 2009). The isotopic fractionation, or offset, between diet and animal tissue for 13C and 15N is approximately 1‰ (Sweeting et al. 2007b) and 3.4‰ (Minagawa and Wada 1984; Sweeting et al. 2007a), respectively. 13C is indicative of the basal source of carbon in the food web, and 15N is used to establish trophic position of an organism, provided the isotopic composition of nitrogen at the base of the food web is known.
SIA can integrate days, months, or sometimes years of diet habits into a single measure, depending upon the tissue turnover time of the body part analyzed (Dalerum and Angerbjörn 2005); thus, it is important to analyze the same tissue type when comparing isotopic data across multiple sites and species. Additionally, the temporal and spatial variability in isotope composition of primary producers must be included (Post 2002). In many cases, suspension feeders (e.g., mussels) and algal grazers (e.g., herbivorous snails) are used to estimate δ15N at the base of the food web (Post 2002). SIA can also provide some information regarding habitat use. Carbon sourced from the nearshore benthic habitat tends to be higher (enriched) in 13C compared to carbon sourced from pelagic or offshore habitats (Fredriksen 2003). Researchers have used stable isotope analysis to discern the influence of organic matter derived from terrestrial, phytoplankton, macroalgae, and saltmarsh sources on estuarine food webs in the Gulf of Maine (Deegan and Garritt 1997; Incze et al. 1982; McMahon et al. 2005; Peterson et al. 1986).
Diet item analysis (DIA) is also useful for determining food web structure, but is notorious for its “snapshot-in-time” nature. Different food items last longer in the gut (Davenport and Bax 2002; Hobson et al. 1994) and can bias DIA toward hard shelled prey like large decapods and gastropods. Variability in fish consumption, both temporally and by individual preference, can limit the wider applicability of DIA (Pinnegar and Polunin 2000).
Here, we investigate the relationship between anadromous fish and marine predators in the bays and coves of Maine within 5 km of shore. We assessed food web structure for readily captured demersal fishes (cod, mackerel, pollock, longhorn sculpin, and shorthorn sculpin) adjacent to large (Midcoast) and small (Passamaquoddy Bay) river herring runs. We hypothesized that differences in the availability of fish prey (e.g., anadromous river herring) would be apparent in the diets of predators (e.g., cod). Stomach content and stable isotope analysis were used as complementary and comparative methods to look for differences between sites and among predators. We also located historical diet studies of groundfish from coastal Maine (Hacunda 1981; Macdonald and Green 1986; Wainright et al. 1993; Willis et al. 2013) to provide a longer-term perspective on fish diets and ecosystem change through the last 50 years.
Methods
Study Area
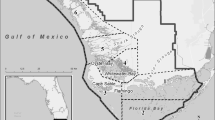
Our study area included Passamaquoddy Bay and the Midcoast Maine (Fig. 1). We sampled fish and their prey at three sites: one to the north (Passamaquoddy Bay) and two in Midcoast Maine, south of Penobscot Bay (St George and Damariscotta). August 2007 water temperature in Harpswell Sound (25 km west of the Damariscotta site) was 16.6 °C (2-m depth), compared to 11.1 °C (1-m depth) in Passamaquoddy Bay’s Western Passage during the same period (Northeastern Regional Association of Coastal and Ocean Observing Systems 2014).

Map of study areas located in Midcoast Maine and Passamaquoddy Bay on the Maine–New Brunswick border. Fish samples were collected from Damariscotta River, St. George River, and Passamaquoddy Bay between 2005 and 2008. Points indicate actual sampling locations. Crab isotope data were collected at Bailey Island, northern Casco Bay
Passamaquoddy Bay is a branch of the Bay of Fundy between Maine, USA, and southwestern New Brunswick, Canada. It is delineated by an archipelago of islands (45.00° N, 66.95° W), which define the two principal entrances to the bay. Our sampling occurred in the Western Passage area (Fig. 1), through which tidal flows can reach 4 m *s− 1 (Trites and Garrett 1983), and the tide is semi-diurnal with a maximum range of 8.3 m (Brooking et al. 2006). Gravel is the most prevalent bottom type at the Passamaquoddy Bay site (49.1%), but rock or hard bottom (33.6%) and sand (17.2%) are also present (Barnhardt et al. 1998).
The Damariscotta River Estuary (Dam; 43.79° N, 69.56° W) and St. George River Estuary (Stg; 43.89° N, 69.29° W) are located within 26 km of each other and are referred to collectively as the Midcoast sites. Each of these sites also features an archipelago of 7–11 islands leading to a narrow drowned river mouth 2–3 km across. Tides are semidiurnal and range from 2.5–3.3 m (Lazzari et al. 2003). Hard (rock) bottom is the dominant bottom type for Damariscotta and St. George (77.3 and 60.8%, respectively) (Barnhardt et al. 1998). Sand (22%) is the second most prevalent bottom type at the Damariscotta site, and mud (37.7%) is the second most prevalent bottom type at the St. George site (Barnhardt et al. 1998). Damariscotta and St. George sites are more similar to each other than either is to Passamaquoddy Bay, as one would expect given their proximity (Fig. 1).
The Passamaquoddy and Midcoast sites differed dramatically in actual productivity (based on river drainages and lake surface area) of river herring runs (Table 1). The St. Croix River was once a rich source of anadromous baitfish for Passamaquoddy Bay, and direct harvests of river herring were substantial (Perley 1852). At the time of our study, the St. Croix River herring run was a fraction of its most recent high of 2.7 million fish in 1987 (Willis 2009) and an estimated carrying capacity of 22.6 million fish (Dill et al. 2010). Little River, a stream in Perry, ME, near our Passamaquoddy sampling site, also contributed some river herring to the bay (Table 1). The 34-km stretch of coast that includes the Midcoast sites encompasses three river systems, Damariscotta, Medomak, and St. George, and the productive (for river herring) Kennebec River lies just to the west. It has been hypothesized that river herring, particularly alewife, stabilized local populations of groundfish until dam construction limited this source of prey (Ames and Lichter 2013; Hall et al. 2010). Historically, the region was called the “Great Fishing Square” and supported the first European fishing camps to exploit the abundance of groundfish accessible within sight of land (Duncan 1992; Snow 1976). Reports from the Damariscotta, St. George, and Kennebec (including the Androscoggin tributary) rivers each showed river herring returns in excess of 300,000 individuals during our study period (Table 1).
Diet Collection and Analysis
Passamaquoddy Bay was sampled in July 2005, April–September 2006, July–October 2007, and May and July–October 2008. Midcoast estuaries were sampled only from May–September 2007, and May and July–October 2008. All sites were within 5 km of the mainland and within 0.5 km of the most seaward island.
Hook-and-line angling was used as the primary method for capturing focal species (e.g., cod, mackerel, pollock, longhorn sculpin, and shorthorn sculpin) to avoid gear conflicts with lobster traps, buoys, and lines. Choice of angling areas was based on bottom topography. Fish were captured using baited hooks and unbaited jigs within 2 m of the bottom sediment as the boat drifted across humps or bottom features where the sea floor shoaled from 25–36 m to 12–18 m. When bait was used, it was primarily cut fish (mackerel, herring, river herring, menhaden, Brevoortia tyrannus in order of use), or occasionally soft shell clam (Mya arenaria). Start and end time of the drift and number of hooks used were recorded to calculate catch-per-unit-effort (CPUE = catch × (persons fishing × minutes)−1 × 60 min). Fishing relatively shallow areas reduced cases of barotrauma and regurgitation. A fish with obvious signs of barotrauma was counted as having an “empty” stomach, and only tissue was collected from these specimens for stable isotope analysis. See Willis et al. (2013) for a more detailed description of fishing methods, including discussion of hook vs. mobile gear bias. In brief, angling collects a smaller size range of fish than trawl gear but with no difference in diet composition or empty vs. nonempty stomachs.
Captured fish were held in a flow-through live well until processed for stomach contents and SIA. Typically, 30 min or less elapsed between capture and processing. We obtained diet samples from cod, mackerel, pollock, and sculpins >15 cm in TL. We separated cod into two categories (small, <30 cm TL, and large, ≥30 cm TL) based on ontogenetic shifts in cod diet preference and obvious breaks in the size distribution (Link and Garrison 2002). Gastric lavage was used to flush the fish stomachs (Hartleb and Moring 1995). Fish were first placed in a weak solution of MS-222 (2 g to 4 L of seawater) until they displayed loss of equilibrium. A 13-mm outer diameter tygon tube connected to a 4-L garden sprayer filled with seawater was inserted into the fish stomach. The stomach was filled with water and gently squeezed. Regurgitated diet items were captured on a 500-μm sieve. Hard, large objects (e.g., crabs) could be felt through the stomach wall after the first flush, triggering a second flush or manual removal (using 30 cm forceps) of diet items that became lodged in the sphincter at the top of the stomach. Tissue samples (dorsal muscle, scales or fin) for SIA were removed after gastric lavage as well. Most fish were released alive after recovery; there was a >75% survival to release for nondissected fish. Mortalities were used to collect additional tissue samples and to test for gastric lavage efficiency.
Gastric lavage quickly stops digestion because digestive acids are rinsed from the sample, and the diet items come in immediate contact with the preservative, a mixture of alcohols and polyethylene glycol (Warmington et al. 2000). The efficiency of diet collection was tested quantitatively to confirm that gastric lavage was effective in flushing diets from the focal species (cod = 85%, n = 20; mackerel = 91%, n = 23; pollock = 100%, n = 20; sculpins = 85%, n = 21). Fish were first lavaged, then euthanized before their stomach was excised to check for additional stomach items. In addition, all technicians were trained in the gastric lavage method by the same PI. When necessary, fish were sacrificed with an overdose of MS-222.
Diet items were identified to the lowest taxonomic level feasible (usually family), counted, and total wet weight for each type of diet item was recorded. Most major diet items were collapsed into large categories for analysis (e.g., crab, amphipod), because, for any given diet, the degree of digestion differed and thus affected the level of taxonomic identification that could be achieved. Proportions of diet contents were expressed as an index of relative importance (%IRI) which accounts for number (N), weight (W), and frequency of occurrence (O) of individual diet items (Garvey and Chipps, 2012; Liao et al., 2002; Willis, 2009):
Blue mussels (Mytilus edulis) were used to estimate the baseline isotope composition of the pelagic food web at all three sites. The isotopic composition of mussel foot tissue represents an integrated signal of diet spanning several weeks which smooths out the spatial and temporal variability of primary producers consumed by these organisms (Dalerum and Angerbjörn 2005; Post 2002). Mussels and all fish tissues were dried at 60 °C for 48 h and homogenized with a mortar and pestle. Mussels were sampled opportunistically from the same locations where fish were collected.
Several of the dominant prey species (as determined from stomach content analysis) were analyzed for stable isotope content. These included river herring and Atlantic herring from the St. George and Passamaquoddy Bay sites, Caprellidae and other amphipoda from the Passamaquoddy Bay site, and Jonah (Cancer borealis), rock (Cancer irroratos), and green (Carcinus maenas) crabs from Bailey Island (Casco Bay), just west of the Damariscotta sites. Crabs were collected and analyzed for SIA in 2007 and 2008 for another study and are presented here.
For SIA of fish, a portion of the anal fin was removed with scissors. Fin membrane was targeted during sampling; however, fin rays were occasionally incidentally sampled as well. Dorsal muscle was collected from a subset of sacrificed fish by removing a 1-cm cube of tissue from a skinless dorsal filet. Fin and muscle were compared for consistency between tissue types and to see whether fin, which was collected from more fish, could be used to increase sample sizes. Fin tissue also records more recent (weeks) diet data due to its fast tissue turnover properties, whereas muscle represents longer-term diet habits based on tissue turnover at the scale of months or longer (Heady and Moore 2013; Perga and Gerdeaux 2005). Differences in isotopic results of fin and muscle tissues for an individual fish meant that the fish had changed diets in the recent past, due to changes in foraging strategy or food available for consumption. For SIA, whole bodies of Caprillidae and amphipods and leg muscle tissue of crabs were dried or lyophilized and homogenized using a mortar and pestle.
Approximately 0.5 mg of dried and powdered tissue per organism was analyzed for 13C and 15N stable isotope composition at two labs. A DeltaPlus XP isotope ratio mass spectrometer (IRMS) interfaced to a Costech ECS4010 Elemental Analyzer (EA) via a ConFlo III was used at the University of New Hampshire Stable Isotope Laboratory and a Thermo Delta V Advantage coupled to a Costech ECS4010 EA via a ConFlo III combustion interface was used at the Environmental Geochemistry Laboratory, Bates College, Maine. All stable isotope values are reported in delta (δ) notation, in units of permille (‰), where δ = [((R)sample/(R)standard) − 1) × 1000], and R = 13C × (12C)−1 or 15N × (14N)−1, and the standards are Vienna PeeDee Belemnite, and air for carbon and nitrogen, respectively. Ten percent of the samples were run in replicate between the two labs; the mean standard error of cross-lab replicates was 0.07‰ for N and 0.02‰ for C.
C/N values provide information on the type and purity of the tissue analyzed. The C/N value of proteins typically ranges between 3.5 and 4.0 (Graham et al. 2013). Neither hydroxyapatite, a potential contaminant to the fin tissues from fin ray bone, nor lipids, a potential contaminant of muscle, contain any nitrogen. The presence of these components might elevate the C/N of the bulk tissue significantly, and bias the isotopic results. We excluded samples with C/N greater than 4.0 from our data analyses to ensure that isotopic comparisons were made solely between muscle and fin proteins.
Data Analysis and Statistics
Principal component analysis (PCA) was used to compare fish diet data (%IRI) among sites and years using Primer 6 (v.6.1.13) and Systat 12 (v.12.00.08). Diet data (%IRI) were cube root transformed to achieve multivariate normality. Analysis of covariance (ANCOVA) was performed to assess the influence of location and fish size on δ15N and δ13C content of muscle or fin tissue. The ANCOVA determines whether the dependent variable (here δ15N or δ13C) is different among the locations sampled despite differences in length of fish sampled at each site (covariate). A homogenous SIA vs. length slope means that SI can be evaluated without confounding variance between length and location.
Results
Catches and Diet Data
CPUE differed among the three sites for different species. Cod and pollock were rare in Passamaquoddy Bay and abundant at Midcoast sites (Table 2). Mackerel were abundant in all locations, particularly in Passamaquoddy Bay. Conversely, shorthorn and longhorn sculpins were prolific at the Passamaquoddy Bay site, but rare at the Midcoast sites (Damariscotta and St. George).
Diet of the five focal species showed considerable overlap in %IRI within sites but also showed some common themes between locations (Table 3). Amphipods in the families Caprellidae and Aoridae were the most prevalent items in fish diets across sites and fish species. At the Passamaquoddy Bay site, amphipods were dominant in the diets of all five focal species with IRIs over 50% (Table 3). Lobster ranked second and fourth in importance for shorthorn sculpin from Damariscotta and St. George, respectively, but no lobster were found in the diets of any fish species from Passamaquoddy Bay. Crab was the top ranked diet item for both sculpin species from St. George and Damariscotta and for cod caught at the St. George site. Fish was not statistically significant in the diet item of any species at any location (Fig. 2), but was a “top three” diet item for Damariscotta large cod (IRI score 33%) and for St. George pollock (IRI score 33% [Table 3]).

Results of principal components analysis depicted with joint plot of diet contents. Rays displayed have significant Pearson correlation of p < 0.05. PC axes 1 and 2 correspond to diet items that defined differences among species diets. Ray length is proportional to the importance of the diet item. The farther from the origin a point is on a ray axis, the stronger the association of that diet item with diets of that species-site combination. Pas = Passamaquoddy, Dam = Damariscotta, Stg = St. George; cod = Atlantic cod <30 cm, Atlantic cod1 = cod ≥30 cm, mack = Atlantic mackerel, poll = Pollock, long = Longhorn sculpin, shor = Shorthorn sculpin
Cod and mackerel CPUE was lowest in the spring and increased in the summer and fall (Fig. 3a, c). Fish were more prevalent in diets in the fall of 2007 for cod, mackerel, and pollock (Table 4). However, on average, fish were found in about 40% (frequency of occurrence) of the cod collected from Damariscotta and St. George, regardless of season; frequency of occurrence of fish was higher for Passamaquoddy Bay cod, though CPUE for cod was lower (Fig. 3b). Clupeid fishes (Atlantic herring and river herring) were positively identified in diets in the fall of 2007; the majority of those clupeids were juveniles found in diets of mackerel and pollock.

Catch rate (CPUE) of Atlantic cod (a) and Atlantic mackerel (c) by site and season. The average proportion of fish in Atlantic cod (b) and Atlantic mackerel (d) diets by site and season are also presented. CPUE and diet proportions were averaged over 2007 and 2008, years when all three areas were fished concurrently. Error bars are ±1 SD. See Table 4 for dates corresponding to seasons. Black = Damariscotta, white = St. George, gray = Passamaquoddy
Ordination of IRI scores grouped combinations of species and sites into three loose clusters based on diet content. PCA groups sites into clusters based on the similarity in species composition found at those sites. Sites with more similar species lists in more similar proportions are placed closer together in n-dimensional space. Here, sites are the species location combinations (e.g., Stgcod = St. George small cod) and diet contents are the equivalent of species lists. The PCA explained 38 and 32% of the variation in diet among combinations of species and sites on principal components axis 1 (PC1) and axis 2 (PC2), respectively (Fig. 2); an additional 11% of the variation in diet was explained on principal components axis 3 (not shown). Mackerel from all three sites grouped together based on a high occurrence of zooplankton in the diets. Passamaquoddy Bay large and small cod, pollock, and both species of sculpin grouped together based on a high occurrence of amphipods. Small Damariscotta cod also had amphipods as a large component of their diets. Sculpins and large cod from the Midcoast sites grouped together based on a high occurrence of decapod crustaceans in the diets, crab being more prevalent than lobster. Lobster only made up 3–6% of the diet for Midcoast cod. St. George and Damariscotta pollock did not cluster with other species, emblematic of their varied diet (Table 3). Fish were not a statistically significant vector in the PCA (p > 0.05).
Isotope Data
Blue mussel isotope values were similar across sites and with other reported mussel data from the Gulf of Maine (Lesser et al. 2010), suggesting similar baseline conditions that facilitated direct comparison of δ15N data, in particular, across sites. The average δ15N from all three sites was 8.3‰ (±0.2 SD). Mean δ13C values for all three sites was −18.8‰ (±0.6 SD).
Muscle and fin samples were compared in two ways: muscle vs. fin by site and species; only matching muscle vs. fin tissue from the same individual fish. Fin and muscle tissue differed by site only for Passamaquoddy Bay mackerel (15N) and Passamaquoddy longhorn sculpin (13C) (Table 5). Mackerel fins were significantly depleted in 15N and pollock fins were significantly enriched in 13C (Table 6) compared to muscle for tissues from the same fish. Except for these instances, muscle and fin values were interchangeable indicating that most species were consuming isotopically similar diets over timescales of months (recorded in muscle) to weeks (recorded in fins). Most analyses going forward were conducted on fin tissue because of the larger number of fins collected, the exception being mackerel in which case muscle tissue was used.
The δ15N values for fish from the Midcoast sites were consistently enriched relative to fish from Passamaquoddy Bay (Table 7; Fig. 4), despite similarities in baseline mussel SI values. Slopes of length vs. δ15N relationships were homogenous allowing influence of length to be accounted for in all comparisons except cod. Length was a significant covariate in explaining δ15N of all species, as expected with ontogenetic shifts in fish size and size of prey. The strong effect of site vs. δ15N for cod was confounded by the steep, positive, nonhomogenous slope of the length vs. δ15N regression for Passamaquoddy Bay cod (Fig. 4; Table 7). The shorthorn sculpin ANCOVA was statistically significant for δ13C between sites, but Tukey’s post hoc test did not indicate what sites were statistically different.

δ15N and δ13C vs. fish TL for four focal species. In each case Passamaquoddy (square) fish had significantly (p < 0.05) lower δ15N values (Table 5) than Damariscotta (diamond) or St. George (triangle) fish. (a) Cod ≥30 cm, (b) pollock, (c) longhorn sculpin, and (d) shorthorn sculpin; superscript number 1 = del15N, superscript number 2 = del13C. Atlantic mackerel values are not plotted because there was no clear separation in del15N and del13C values among sites
Corroborating Diet Analysis Methods
Stable isotope data from predatory fish in our study corroborated well with observed stomach contents (DIA). For example, the δ15N values were lower in fish from Passamaquoddy Bay than those from the Midcoast, corresponding to low trophic level prey in fish diets. The average longhorn sculpin diet from Passamaquoddy Bay contained 89% amphipods (δ15N = 6.4), compared to that from Damariscotta, which contained 80% crab (δ15N ranged between 9.5 and 12.2‰; Table 5). The difference in carbon source also agreed between diets and SIA values. Crab and amphipod were heavily represented in sculpin diets compared to cod diets (Table 3), and δ13C SIA values reflected more benthic energy sources in sculpin tissue and more pelagic energy sources in cod tissue (Table 5). For example, the contribution of pelagic sourced carbon was apparent in Passamaquoddy Bay when compared between cod (55% amphipods, δ13C = −17.7‰) and shorthorn sculpin (60% amphipods, δ13C = −15.9‰).
Discussion
We noted an increase of predator presence in the nearshore in the fall, concurrent with more (juvenile) fish, including river herring, appearing in predator diets. We expected adult river herring to be prevalent in the diet of cod from the Midcoast because of the large spawning migration that occurs there, but this was not the case. Fish (as a broadly defined diet item) was a top three diet item for large Atlantic cod from Damariscotta and pollock from St. George. Juvenile river herring comprised up to 41% of the fish in the diets of all fishes collected in the Damariscotta region of the Midcoast in fall 2007. In other years, the river herring pulse, seasonal outmigration of juveniles from freshwater habitat to the ocean, was overwhelmed by decapods in the diets of Midcoast fish, and amphipods in the diets of Passamaquoddy Bay fish. McDermott et al. (2015) used trawl gear in 2010–2011 in the Midcoast region to address similar questions regarding river herring as groundfish prey, finding that the odds of encountering river herring in groundfish stomachs was highest closest to the rivers where river herring spawn. Smith et al. (2007) noted a similar seasonal pulse pattern of fish prey in cod diets.
The pulse-like nature of river herring as prey may play an important role in the recovery of cod stocks. Sherwood and Rose (2005) argued that seasonally available high protein, high lipid prey is important for the reproductive cycle of cod. Yearling river herring are present in the nearshore Gulf of Maine in the fall, along with the predators that consume them. Because benthic organisms (e.g., crabs and amphipods) are less nutritious than fish (Mullowney and Rose 2014, Sherwood and Rose 2005, Lawson et al. 1998), we posit that lower quality diet items currently being consumed by cod might be contributing to limited recruitment and a small cod population in the nearshore Gulf of Maine.
Fin vs. Muscle (Short-Term vs. Long-Term Feeding Strategies)
Stable isotope values of different tissues often vary depending on their carbon and nitrogen turnover rate (Ankjærø et al. 2012; Sweeting et al. 2007a; Sweeting et al. 2007b). Differences in muscle vs. fin tissue turnover can be interpreted as a recent diet or habitat change. White dorsal muscle tissue, which is most frequently used for SIA, has a turnover time of between 60 and 86 days (Ankjærø et al. 2012). Fin, particularly at the end or tip where new growth occurs, has a shorter turnover time (days to weeks) and reflects the current diet (Boecklen et al. 2011; Dalerum and Angerbjörn 2005; Heady and Moore 2013). The cod we sampled had very similar stable isotope values for fin and muscle, indicating stability in diet and habitat use. In contrast, mackerel from Passamaquoddy Bay showed fin SIA values that were significantly depleted in δ15N compared to muscle, suggesting a more recent shift toward lower trophic level feeding. SIA may be reflecting the ability of mackerel to switch from consuming fish to zooplankton depending upon, we presume, prey availability; a phenomenon we observed in the field (K. Wilson, personal observation). Another hypothesis is that mackerel were “new arrivals” to Passamaquoddy Bay where fish, or some other high trophic level prey, was not as abundant as in a previous habitat. Evidence of the shift was stored in the fast turnover tissue. We interpret this difference in mackerel tissue vs. fin δ15N as indicating the pattern from Passamaquoddy Bay of depleted δ15N is endemic to the food web found in that region.
Benthic vs. Pelagic Organic Matter Sources in the Nearshore Food Web
Carbon isotope values of cod, pollock, and sculpins from Passamaquoddy Bay aligned most closely with Caprellidae (δ13C = −17.6‰) and other amphipoda (δ13C = −16.7‰), providing a link between microphytobenthos production and upper trophic levels (Thiel 1999). Aoridae made up the largest proportion of amphipods in fish diets from Passamaquoddy Bay, in particular the species Leptocheirus pinguis, a burrowing amphipod with documented parental care behavior (Thiel et al. 1997). Previous work in Passamaquoddy Bay demonstrated that not only are L. pinguis and Unicola spp. abundant, but they are chosen as prey more frequently than their abundance would indicate for simple opportunistic predation (Macdonald and Green 1986). L. pinguis and Unicola spp. are also present at Midcoast sites, but other prey was either more plentiful or preferred by demersal predators.
Interestingly, blue mussel δ13C and δ15N signals were similar across all sites, indicating that the base production in the Midcoast and Passamaquoddy food webs was similar. Thus, the observed enriched Midcoast δ15N signal in decapods and decapod predators would have to originate from some pathway other than local pelagic primary production, suggesting that we may have seen an effect of lobster fishing related to higher trap densities in Midcoast Maine. Baited lobster traps introduce hundreds of metric tons of pelagic zooplanktivorous fish (e.g., Atlantic herring) to the benthos along the Maine coast, and the Midcoast region is one of the most heavily trapped areas in Maine. Lobster have a significantly enriched δ15N value that is correlated with trap density (Grabowski et al. 2010; Grabowski et al. 2009), i.e., correlated with access to a fish-based diet. Thus, lobsters, crabs, and predators of the 15N-enriched species that visit lobsters traps would be expected to show relatively enriched δ15N values. Although lobsters were not prevalent diet items in this study, crabs are frequently encountered as bycatch in lobster traps and crabs were common diet items.
Fishing as an Ecosystem Driver
It is important to put the results we found in the context of overall changes to nearshore ecosystems in the Gulf of Maine over time. We attempted to focus on cod, but this species was relatively rare compared to other noncommercial groundfish. Previous studies of Passamaquoddy Bay in the 1960s and Midcoast sites in the 1970s revealed that cod food webs were dominated by small bodied prey, such as amphipods, carideans, euphausiids, and mysids (Hacunda 1981; Macdonald and Green 1986; Tyler 1971; Tyler 1972; Willis et al. 2013). Indeed, for locations over 250 km apart with different temperature regimes and substrate types, the diets of cod in this study looked remarkably similar for that time period. The results of this study indicate a divergence in diets, where Midcoast diets had more fish than reported in the 1970s, but Passamaquoddy Bay diets were essentially the same. How has Midcoast Maine changed?
The proliferation of lobster fishing in Maine has changed accessibility to fishing grounds for mobile gear fishermen. Groundfish rebuilding efforts associated with the Magnuson-Stevens Fishery Act of 1976 effectively reduced nearshore fishing effort that had increased steadily since the 1940s (Ames 2004; Serchuk and Wigley 1992). Many day-boat fishermen (∼10 m vessels) switched to lobstering in the 1990s (Acheson 2006; Acheson 2013; Steneck et al. 2011), with a resultant increase in stationary fishing gear that interfered with trawls and dredges. Landing trawl-caught lobster is illegal in Maine (Sec. 2. 12 MRSA §6432, sub-§1), which also helped displace bottom trawl gear from state waters. Stationary gear is comparatively less dense in Passamaquoddy Bay (T. Willis, personal observation) and mobile gear trawl fisheries include scallop in winter, sea cucumber and groundfish in summer, and urchin in the fall. In addition, most of Passamaquoddy Bay is in Canada, which has different fishing seasons and effort than Maine fisheries, e.g., additional scallop season in summer.
Fishing activity could explain the food web differences we found between Damariscotta and the Midcoast, either through lobster trap bait, as discussed above, mobile gear habitat disturbance, or “competition” between fish predators and fishermen for midtrophic level prey. Commercial fishing, particularly mobile gear, has a measureable effect on food webs and predator stable isotope values, i.e., low diversity, bottom disturbance, and reduced trophic position of predators (Badalamenti et al. 2002; Fanelli et al. 2010; Robinson et al. 2001; Shephard et al. 2014; Wainright et al. 1993). Some of these negative effects were documented in the Passamaquoddy Bay region (Robinson et al. 2001). Researchers have also documented lower δ15N values for predatory fishes whose fish prey become less abundant. For example, Gomez-Campos et al. (2011) documented lower δ15N values for adult striped dolphin (Stenella coeruleoalbaa) because of commercial exploitation of sardine. Wainright et al. (1993) documented a decline in George’s Bank haddock (Melanogramus aeglefinus) δ15N which, they commented, could have been indicative of a change in haddock feeding ecology related to pelagic prey availability or injection of δ15N depleted material into the food web through bottom disturbance. This statement referred, at least in part, to the overfishing of George’s Bank Atlantic herring prior to the stock collapse of 1977 (Melvin and Stephenson 2007; Overholtz 2002). In nearshore Maine, we did not see evidence of depleted material injection in the δ15N signature of blue mussels (Table 5), but we did document filter feeding and detritivorous amphipods depleted by 1.3 to 1.9 ‰ relative to blue mussels in Passamaquoddy Bay. This indicates that 15N depleted nitrogen may be important for detritivores in the Passamaquoddy Bay system and begs for additional study.
River Restoration and Nearshore Food Webs
Restoring river herring has been touted as a panacea for restoring the nearshore food web in the Gulf of Maine. Recent restoration efforts have led to differences in river herring abundance in bays of the Maine coast. There was an order of magnitude difference in river herring run size between Passamaquoddy Bay and Midcoast Maine (Table 1), and those numbers likely underestimate the total number of adults because fishways do not pass all the fish that return to spawn (Brown et al. 2013). Where the biggest river herring restoration gains have been made we found evidence of more fish in cod diets, higher cod CPUE and more diverse cod diets. River herring were prevalent in seasonal pulse events as juveniles, dropping into the coastal zone in the later summer and fall (K. Wilson, personal observation). We directly observed juvenile river herring being consumed by cod, mackerel, and pollock in the Midcoast, confirming the assertions of Field (1914): “the value of the alewife [is] as a food for the larger fish which frequent the Massachusetts waters.” However, recovery of spawning aggregations of groundfish (Ames 2004; Ames and Lichter 2013) may be many years into the future, as illustrated by cod fishing being deemed unsustainable in the Gulf of Maine in the 1990s (Acheson 2006; Hennessey and Healey 2000) and again in the 2010s (Department of Commerce, National Oceanic and Atmospheric Administration, D.O.C., 2016). Changes in the physical habitat of the Gulf of Maine (Fogarty et al. 2008; Pershing et al. 2015) and alterations in the ecosystem may produce ecological bottle necks for once abundant predatory fishes (Swain and Sinclair 2000; Walters and Kitchell 2001). Indeed, increasing amounts of energy has been concentrated in less commercially desirable components of the food web (Auster and Link 2009; Link et al. 2009; Lotze and Milewski 2004). That imbalance, fewer and smaller cod and pollock, and more sculpins, is apparent in Passamaquoddy Bay.
Recovery of fish populations does not necessarily follow the same trajectory as decline. Atlantic herring, an important pelagic prey species, has recovered in locations where cod were once economically and ecologically important (Melvin and Stephenson 2007), but demersal fish populations have not (Swain and Sinclair 2000), hinting that Atlantic herring may be limiting recruitment of herring predators (Walters and Kitchell 2001). Whether river herring also can reach abundances where their predation on plankton causes high mortality for the larval stage of their predators is an open question worthy of further study.
Removing barriers to migration appears to increase the number of river herring in a river system, as evidenced by increasing river herring counts in the Kennebec and Penobscot Rivers (Maine Dept. of Marine Resources, unpublished data), and increases the availability of lipid-rich seasonal prey items for marine fishes. Here, we present evidence that physical improvements to coastal freshwater habitats can be beneficial up the food chain to populations of marine predators, and make a case for monitoring the marine effects of these large-scale manipulations closely.
References
Acheson, J.M. 2006. Lobster and groundfish management in the Gulf of Maine: a rational choice perspective. Human Organization 65: 240–252.
Acheson, J.M. 2013. Co-management in the Maine lobster industry: a study in factional politics. Conservation & Society 11: 60–71.
Ames, E.P. 2004. Atlantic cod stock structure in the gulf of Maine. Fisheries 29: 10–28.
Ames, E.P., and J. Lichter. 2013. Gadids and alewives: structure within complexity in the gulf of Maine. Fisheries Research 141: 70–78.
Ankjærø, T., J.T. Christensen, and P. Grønkjær. 2012. Tissue-specific turnover rates and trophic enrichment of stable n and c isotopes in juvenile Atlantic cod (Gadus morhua) fed three different diets. Marine Ecology Progress Series 461: 197–209.
Auster, P.J., and J.S. Link. 2009. Compensation and recovery of feeding guilds in a Northwest Atlantic shelf fish community. Marine Ecology-Progress Series 382: 163–172.
Badalamenti, F., G. D'Anna, J.K. Pinnegar, and N.V.C. Polunin. 2002. Size-related trophodynamic changes in three target fish species recovering from intensive trawling. Marine Biology 141: 561–570.
Barnhardt, W.A., J.T. Kelley, S.M. Dickson, and D.F. Belknap. 1998. Mapping the gulf of Maine with side-scan sonar: a new bottom-type classification for complex seafloors. Journal of Coastal Research 14: 646–659.
Boecklen, W.J., C.T. Yarnes, B.A. Cook, and A.C. James. 2011. On the use of stable isotopes in trophic ecology. In Annual review of ecology, evolution, and systematics, vol 42, ed. D.J. Futuyma, H.B. Shaffer, and D. Simberloff, 411–440. Palo Alto: Annual Reviews.
Brooking, P., G. Doucette, S. Tinker, and F.G. Whoriskey. 2006. Sonic tracking of wild cod, (Gadus morhua), in an inshore region of the Bay of Fundy: a contribution to understanding the impact of cod farming for wild cod and endangered salmon populations. ICES Journal of Marine Science 63: 1364–1371.
Brown, J.J., K.E. Limburg, J.R. Waldman, K. Stephenson, E.P. Glenn, F. Juanes, and A. Jordaan. 2013. Fish and hydropower on the US Atlantic coast: failed fisheries policies from half-way technologies. Conservation Letters 6: 280–286.
Caut, S., E. Angulo, and F. Courchamp. 2009. Variation in discrimination factors (delta n-15 and delta c-13): the effect of diet isotopic values and applications for diet reconstruction. Journal of Applied Ecology 46: 443–453.
Collette, B.B., and G. Klein-MacPhee. 2002. Bigelow and Schroeder’s fishes of the gulf of Maine. Washington, D.C.: Smithsonian Institution Press.
Dalerum, F., and A. Angerbjörn. 2005. Resolving temporal variation in vertebrate diets using naturally occurring stable isotopes. Oecologia 144: 647–658.
Davenport, S.R., and N.J. Bax. 2002. A trophic study of a marine ecosystem off southeastern Australia using stable isotopes of carbon and nitrogen. Canadian Journal of Fisheries and Aquatic Sciences 59: 514–530.
Deegan, L.A., and R.H. Garritt. 1997. Evidence for spatial variability in estuarine food webs. Marine Ecology Progress Series 147: 31–47.
Department of Commerce, National Oceanic and Atmospheric Administration. 2016. Magnuson-Stevens Fishery Conservation and Management Act Provisions; Fisheries of the Northeastern United States; Northeast Groundfish Fishery; Framework Adjustment 55 and Magnuson-Stevens Fishery Conservation and Management Act Provisions; Fisheries of the Northeastern United States; Northeast Groundfish Fishery; Fishing Year 2016; Recreational Management Measures; Final Rules. Federal Register 81: 26412–26452.
Dill, R., J. Trial, E. Atkinson, J. Gibson, R. Jordan, S.R. Lary, R. Sauders, and P. Seymour. 2010. An Adaptive plan for managing alewife in the St. Croix River watershed, Maine and New Brunswick. International Joint Commission, International St. Croix River Watershed Board. www.ijc.org/files/tinymce/uploaded/st_croix_amp.pdf. Accessed 2 August 2016.
Duncan, R.F. 1992. Coastal Maine: a maritime history. New York, NY: W.W. Norton & Company, Inc..
Essington, T.E., A.H. Beaudreau, and J. Wiedenmann. 2006. Fishing through marine food webs. Proceedings of the National Academy of Sciences of the United States of America 103: 3171–3175.
Fanelli, E., F. Badalamenti, G. D'Anna, C. Pipitone, and C. Romano. 2010. Trophodynamic effects of trawling on the feeding ecology of Pandora, Pagellus erythrinus, off the northern Sicily coast (Mediterranean Sea). Marine and Freshwater Research 61: 408–417.
Field, G.W. 1914. Alewife fishery of Massachusetts. Transactions of the American Fisheries Society 43: 143–151.
Fogarty, M., L. Incze, K. Hayhoe, D. Mountain, and J. Manning. 2008. Potential climate change impacts on Atlantic cod (Gadus morhua) off the northeastern USA. Mitigation and Adaptation Strategies for Global Change 13: 453–466.
Fredriksen, S. 2003. Food web studies in a Norwegian kelp forest based on stable isotope (delta c-13 and delta n-15) analysis. Marine Ecology Progress Series 260: 71–81.
Garrison, L.P., and J.S. Link. 2000. Dietary guild structure of the fish community in the Northeast United States continental shelf ecosystem. Marine Ecology-Progress Series 202: 231–240.
Garvey, J.E., Chipps, S.R., 2012. Diets and Energy Flow. In Fisheries Techniques, 3rd Edition. ed. A.V. Zale, Parrish, D.L., Sutton, T.M, 733-779. Bethesda: American Fisheries Society.
Gomez-Campos, E., A. Borrell, L. Cardona, J. Forcada, and A. Aguilar. 2011. Overfishing of small pelagic fishes increases trophic overlap between immature and mature striped dolphins in the Mediterranean Sea. Plos One 6: e24554.
Grabowski, J.H., E.J. Clesceri, A.J. Baukus, J. Gaudette, M. Weber, and P.O. Yund. 2010. Use of herring bait to farm lobsters in the Gulf of Maine. PloS One 5: e10188.
Grabowski, J.H., J. Gaudette, E.J. Clesceri, and P.O. Yund. 2009. The role of food limitation in lobster population dynamics in coastal Maine, United States, and New Brunswick, Canada. New Zealand Journal of Marine and Freshwater Research 43: 185–193.
Graham, C.T., S.S.C. Harrison, and C. Harrod. 2013. Development of non-lethal sampling of carbon and nitrogen stable isotope ratios in salmonids: effects of lipid and inorganic components of fins. Isotopes in Environmental and Health Studies 49: 555–566.
Hacunda, J.S. 1981. Trophic relationships among demersal fishes in a coastal area of the Gulf of Maine. Fishery Bulletin 79: 775–788.
Hall, C.J., A. Jordaan, and M.G. Frisk. 2010. The historic influence of dams on diadromous fish habitat with a focus on river herring and hydrologic longitudinal connectivity. Landscape Ecology 26: 95–107.
Halpern, B.S., S. Walbridge, K.A. Selkoe, C.V. Kappel, F. Micheli, C. D'Agrosa, J.F. Bruno, K.S. Casey, C. Ebert, H.E. Fox, R. Fujita, D. Heinemann, H.S. Lenihan, E.M.P. Madin, M.T. Perry, E.R. Selig, M. Spalding, R. Steneck, and R. Watson. 2008. A global map of human impact on marine ecosystems. Science 319: 948–952.
Hartleb, C.F., and J.R. Moring. 1995. An improved gastric lavage device for removing stomach contents from live fish. Fisheries Research 24: 261–265.
Heady, W.N., and J.W. Moore. 2013. Tissue turnover and stable isotope clocks to quantify resource shifts in anadromous rainbow trout. Oecologia 172: 21–34.
Hennessey, T., and M. Healey. 2000. Ludwig's ratchet and the collapse of New England groundfish stocks. Coastal Management 28: 187–213.
Hobson, K.A., J.F. Piatt, and J. Pitocchelli. 1994. Using stable isotopes to determine seabird trophic relationships. Journal of Animal Ecology 63: 786–798.
Incze, L.S., L.M. Mayer, E.B. Sherr, and S.A. Macko. 1982. Carbon inputs to bivalve mollusks: a comparison of two estuaries. Canadian Journal of Fisheries and Aquatic Sciences 39: 1348–1352.
Johannesen, E., U. Lindstrom, K. Michalsen, M. Skern-Mauritzen, P. Fauchald, B. Bogstad, and A. Dolgov. 2012. Feeding in a heterogeneous environment: spatial dynamics in summer foraging Barents Sea cod. Marine Ecology Progress Series 458: 181–197.
Lawson, J.W., A.M. Magalhaes, and E.H. Miller. 1998. Important prey species of marine vertebrate predators in the Northwest Atlantic: proximate composition and energy density. Marine Ecology Progress Series 164: 13–20.
Lake, T.R.T., K.R. Ravana, and R. Saunders. 2012. Evaluating changes in diadromous species distributions and habitat accessibility following the Penobscot River restoration project. Marine and Coastal Fisheries 4: 284–293.
Lazzari, M.A., S. Sherman, and J.K. Kanwit. 2003. Nursery use of shallow habitats by epibenthic fishes in Maine nearshore waters. Estuarine Coastal and Shelf Science 56: 73–84.
Lesser, M.P., M.A. Bailey, D.G. Merselis, and J.R. Morrison. 2010. Physiological response of the blue mussel, Mytilus edulis, to differences in food and temperature in the gulf of Maine. Comparative Biochemistry and Physiology a-Molecular & Integrative Physiology 156: 541–551.
Liao, H., C. L. Pierce, J. G. Larscheid, (2002) Diet dynamics of the adult piscivorous fish community in Spirit Lake, Iowa, USA 1995-1997. Ecology of Freshwater Fish 11(3): 178-189
Link, J.S., B. Bogstad, H. Sparholt, and G.R. Lilly. 2009. Trophic role of Atlantic cod in the ecosystem. Fish and Fisheries 10: 58–87.
Link, J.S., and L.P. Garrison. 2002. Trophic ecology of Atlantic cod (Gadus morhua) on the northeast U.S. continental shelf. Marine Ecology Progress Series 227: 109–123.
Lotze, H.K., and I. Milewski. 2004. Two centuries of multiple human impacts and successive changes in a North Atlantic food web. Ecological Applications 14: 1428–1447.
Macdonald, J.S., and R.H. Green. 1986. Food resource utilization by 5 species of benthic feeding fish in Passamaquoddy Bay, New Brunswick. Canadian Journal of Fisheries and Aquatic Sciences 43: 1534–1546.
McDermott, S.P., N.C. Bransome, S.E. Sutton, B.E. Smith, J.S. Link, and T.J. Miller. 2015. Quantifying Alosine prey in the diets of marine piscivores in the Gulf of Maine. Journal of Fish Biology 86: 1811–1829.
McMahon, K.W., B.J. Johnson, and W.G. Ambrose. 2005. Diet and movement of the killifish, Fundulus heteroclitus, in a Maine salt marsh assessed using gut contents and stable isotope analyses. Estuaries 28: 966–973.
Melvin, G.D., and R.L. Stephenson. 2007. The dynamics of a recovering fish stock: Georges Bank herring. ICES Journal of Marine Science 64: 69–82.
Michener, R.H., and L. Kaufman. 2007. Stable isotope ratios as tracers in marine food webs: An update. In Stable Isotopes in Ecology and Environmental Science, 2nd Edition ed. R.H. Michener and K. Lajtha, 238–282. Malden: Blackwell Publishing.
Minagawa, M., and E. Wada. 1984. Stepwise enrichment of 15n along food chains: further evidence and the relation between δ15n and animal age. Geochimica et Cosmochimica Acta 48: 1135–1140.
Mullowney, D.R.J., and G.A. Rose. 2014. Is recovery of northern cod limited by poor feeding? The capelin hypothesis revisited. ICES Journal of Marine Science 71: 784–793.
Myers, R.A., and B. Worm. 2005. Extinction, survival or recovery of large predatory fishes. Philosophical Transactions of the Royal Society B-Biological Sciences 360: 13–20.
Northeastern Regional Association of Coastal and Ocean Observing Systems. 2014. Historic data and tools. www.Neracoos.Org. Accessed 1 Aug 2016.
Nye, J.A., A. Bundy, N. Shackell, K.D. Friedland, and J.S. Link. 2009. Coherent trends in contiguous survey time-series of major ecological and commercial fish species in the Gulf of Maine ecosystem. ICES Journal of Marine Science 67: 26–40.
Overholtz, W.J. 2002. The Gulf of Maine - Georges Bank Atlantic herring (Clupea harengus): spatial pattern analysis of the collapse and recovery of a large marine fish complex. Fisheries Research 57: 237–254.
Pace, M.L., J.J. Cole, S.R. Carpenter, and J.F. Kitchell. 1999. Trophic cascades revealed in diverse ecosystems. Trends in Ecology & Evolution 14: 483–488.
Perga, M.E., and D. Gerdeaux. 2005. 'Are fish what they eat' all year round? Oecologia 144: 598–606.
Perley, M. 1852. Reports on the sea and river fisheries of New Brunswick. Fredericton.
Pershing, A.J., M.A. Alexander, C.M. Hernandez, L.A. Kerr, A. Le Bris, K.E. Mills, J.A. Nye, N.R. Record, H.A. Scannell, J.D. Scott, G.D. Sherwood, and A.C. Thomas. 2015. Slow adaptation in the face of rapid warming leads to collapse of the Gulf of Maine cod fishery. Science 350: 809–812.
Peterson, B.J., R.W. Howarth, and R.H. Garritt. 1985. Multiple stable isotopes used to trace the flow of organic-matter in estuarine food webs. Science 227: 1361–1363.
Peterson, B.J., R.W. Howarth, and R.H. Garritt. 1986. Sulfur and carbon isotopes as tracers of salt-marsh organic matter flow. Ecology 67: 865–874.
Pinnegar, J.K., and N.V.C. Polunin. 2000. Contributions of stable-isotope data to elucidating food webs of Mediterranean rocky littoral fishes. Oecologia 122: 399–409.
Post, D.M. 2002. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83: 703–718.
Robinson, S.M.C., S. Bernier, and A. MacIntyre. 2001. The impact of scallop drags on sea urchin populations and benthos in the Bay of Fundy, Canada. Hydrobiologia 465: 103–114.
Saunders, R., M.A. Hachey, and C.W. Fay. 2006. Maine's diadromous fish community: past, present, and implications for Atlantic salmon recovery. Fisheries 31: 537–547.
Serchuk, F.M., and S.E. Wigley. 1992. Assessment and management of the Georges Bank cod fishery: an historical review and evaluation. Journal of Northwest Atlantic Fishery Science 13: 25–52.
Shephard, S., C. Minto, M. Zolck, S. Jennings, D. Brophy, and D. Reid. 2014. Scavenging on trawled seabeds can modify trophic size structure of bottom-dwelling fish. ICES Journal of Marine Science 71: 398–405.
Sherwood, G.D., R.M. Rideout, S.B. Fudge, and G.A. Rose. 2007. Influence of diet on growth, condition and reproductive capacity in Newfoundland and Labrador cod (Gadus morhua): insights from stable carbon isotopes (delta c-13). Deep-Sea Research Part Ii-Topical Studies in Oceanography 54: 2794–2809.
Sherwood, G.D., and G.A. Rose. 2005. Stable isotope analysis of some representative fish and invertebrates of the Newfoundland and Labrador continental shelf food web. Estuarine, Coastal and Shelf Science 63: 537–549.
Smith, B.E., T.J. Ligenza, F.P. Almeida, and J.S. Link. 2007. The trophic ecology of Atlantic cod: insights from tri-monthly, localized scales of sampling. Journal of Fish Biology 71: 749–762.
Snow, H.F. 1976. Notes on the history of Damariscove Island in Maine to 1614: Including the early history of Maine the famous "fishing square between the island of Damariscove, Mohegan and Pemaquid with the first Englishmen in America, 1497–1605. Madison, CT: privately printed.
Steneck, R.S., T.P. Hughes, J.E. Cinner, W.N. Adger, S.N. Arnold, F. Berkes, S.A. Boudreau, K. Brown, C. Folke, L. Gunderson, P. Olsson, M. Scheffer, E. Stephenson, B. Walker, J. Wilson, and B. Worm. 2011. Creation of a gilded trap by the high economic value of the Maine lobster fishery. Conservation Biology 25: 904–912.
Swain, D.P., and A.F. Sinclair. 2000. Pelagic fishes and the cod recruitment dilemma in the Northwest Atlantic. Canadian Journal of Fisheries and Aquatic Sciences 57: 1321–1325.
Sweeting, C.J., J. Barry, C. Barnes, N.V.C. Polunin, and S. Jennings. 2007a. Effects of body size and environment on diet-tissue delta n-15 fractionation in fishes. Journal of Experimental Marine Biology and Ecology 340: 1–10.
Sweeting, C.J., J.T. Barry, N.V.C. Polunin, and S. Jennings. 2007b. Effects of body size and environment on diet-tissue delta c-13 fractionation in fishes. Journal of Experimental Marine Biology and Ecology 352: 165–176.
Thiel, M. 1999. Duration of extended parental care in marine amphipods. J. Crust. Biol. 19: 60–71.
Thiel, M., S. Sampson, and L. Watling. 1997. Extended parental care in two endobenthic amphipods. Journal of Natural History 31: 713–725.
Trites, R.W., and C.J.R. Garrett. 1983. Physical oceanography of the Quoddy region. In Marine and coastal systems of the Quoddy region, New Brunswick, ed. M.L.H. Thomas, 9–34. Ottawa: Canadian Special Publication of Fisheries and Aquatic Sciences.
Tyler, A.V. 1971. Monthly changes in stomach contents of demersal fishes in Passamaquoddy Bay, N.B., ed. Fisheries Research Board of Canada, 100. Ottawa, ON.
Tyler, A.V. 1972. Food resource division among northern, marine, demersal fishes. Journal of the Fisheries Research Board of Canada 29: 997–1003.
Wainright, S.C., M.J. Fogarty, R.C. Greenfield, and B. Fry. 1993. Long-term changes in the Georges Bank food web - trends in stable isotopic compositions of fish scales. Marine Biology 115: 481–493.
Walters, C., and J.F. Kitchell. 2001. Cultivation/depensation effects on juvenile survival and recruitment: implications for the theory of fishing. Canadian Journal of Fisheries and Aquatic Sciences 58: 39–50.
Warmington, A.R., J.M. Wilkinson, and C.B. Riley. 2000. Evaluation of ethanol-based fixatives as a substitute for formalin in diagnostic clinical laboratories. Journal of Histotechnology 23: 299–308.
Willis, T.V. 2009. How policy, politics, and science shaped a 25-year conflict over alewife in the St. Croix river, New Brunswick-Maine. In Challenges for diadromous fishes in a dynamic global environment, ed. A. Haro, K.L. Smith, R.A. Rulifson, C.M. Moffitt, R.J. Klauda, M.J. Dadswell, R.A. Cunjak, J.E. Cooper, K.L. Beal, and T.S. Avery, 793–811. Bethesda: American Fisheries Society.
Willis, T.V., K.A. Wilson, K.A. Alexander, and W.B. Leavenworth. 2013. Tracking cod diet preference over a century in the northern gulf of Maine: historic data and modern analysis. Marine Ecology Progress Series 474: 263–276.
Worm, B., E.B. Barbier, N. Beaumont, J.E. Duffy, C. Folke, B.S. Halpern, J.B.C. Jackson, H.K. Lotze, F. Micheli, S.R. Palumbi, E. Sala, K.A. Selkoe, J.J. Stachowicz, and R. Watson. 2006. Impacts of biodiversity loss on ocean ecosystem services. Science 314: 787–790.
Acknowledgements
This material is based upon work supported by the National Oceanic and Atmospheric Administration under Award No. NA05NMF4721057 to the Northeast Consortium at the University of New Hampshire, the National Science Foundation under Grant No. EPS-0904155 to Maine EPSCoR at the University of Maine, Maine Sea Grant development fund (Award No. NA06OAR4170108), and the NSF under Grant No. OCE-0929900 awarded to Bates College. This project would not have been possible without the dedication and commitment of D. Turner and field and lab support of P. Dostie, K. Moulton, A. Pearson, S. Prescott, M. Rautenberg, and R. Tiller. We also thank project collaborators C. Bartlett, and Captains M. Myrick, J. Stoltz, and C. Taylor. The manuscript was improved thanks to the comments of five anonymous reviewers. The project was carried out with International Animal Care and Use Committee approval from the University of Southern Maine (numbers 051606, 061110, and 042412).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Charles Simenstad
Rights and permissions
About this article

Cite this article
Willis, T.V., Wilson, K.A. & Johnson, B.J. Diets and Stable Isotope Derived Food Web Structure of Fishes from the Inshore Gulf of Maine. Estuaries and Coasts 40, 889–904 (2017). https://doi.org/10.1007/s12237-016-0187-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-016-0187-9