
Abstract
We examined the potential impacts of future climate change on the distribution and production of Atlantic cod (Gadus morhua) on the northeastern USA’s continental shelf. We began by examining the response of cod to bottom water temperature changes observed over the past four decades using fishery-independent resource survey data. After accounting for the overall decline in cod during this period, we show that the probability of catching cod at specified locations decreased markedly with increasing bottom temperature. Our analysis of future changes in water temperature was based on output from three coupled atmosphere–ocean general circulation models under high and low CO2 emissions. An increase of <1.5°C is predicted for all sectors under the low emission scenario in spring and autumn by the end of this century. Under the high emission scenario, temperature increases range from ∼2°C in the north to >3.5°C in the Mid-Atlantic Bight. Under these conditions, cod appear vulnerable to a loss of thermal habitat on Georges Bank, with a substantial loss of thermal habitat farther south. We also examined temperature effects on cod recruitment and growth in one stock area, the Gulf of Maine, to explore potential implications for yield and resilience to fishing. Cod survival during the early life stages declined with increasing water temperatures, offsetting potential increases in growth with warmer temperatures and resulting in a predicted loss in yield and increased vulnerability to high fishing mortality rates. Substantial differential impacts under the low versus high emission scenarios are evident for cod off the northeastern USA.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
Fishing has played a vital role in the cultural and economic fabric of the northeastern USA and the identity of many coastal communities is deeply tied to longstanding seafaring and fishing traditions. Combined dockside revenues from oceanic and estuarine commercial fisheries in the northeastern USA exceeded $1.2 billion in 2004. Hoagland et al. (2005) estimated the economic contribution of the seafood industry to the overall economy in the northeast USA to be $10.4 billion in 2003 as a result of direct and indirect effects of commercial fishing activity and satellite industries. Recreational fisheries also result in substantial economic benefits in the region derived from angler expenditures and employment opportunities (Steinback et al. 2004).
We explored the potential consequences of climate change on marine resources of the northeastern USA with particular emphasis on Atlantic cod (Gadus morhua). The demand for salt cod in Europe served as a catalyst for exploration and discovery and there is evidence that Basque fishermen discovered and occupied important cod fishing grounds in North America as early as the 15th century (Kurlansky 1997). Cod has been a mainstay of the commercial fishery in New England since the seventeenth century, and a carved representation of the ‘Sacred Cod’ has hung in the rotunda of the State House of Massachusetts since 1784, signaling the historical importance of this species to the Commonwealth.
Populations of cod in the Gulf of Maine and on Georges Bank off the northeastern USA (Fig. 1) are at the southern extent of the species range in the Northwest Atlantic. Accordingly, potential increases in water temperature under projected climate change scenarios can be expected to affect both their distribution and abundance. Landings of cod have undergone large scale and coherent fluctuations over the last century in the Gulf of Maine and on Georges Bank and are currently at low levels in both regions (Fig. 2). Periods of low cod landings have corresponded with periods of high water temperature, although many fishery-related factors affect landings in addition to environmental variables.

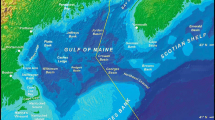
Map of the study area on the continental shelf in the northeast USA including the Gulf of Maine (GOM), Georges Bank, and the Mid-Atlantic Bight (MAB)

Landings of Atlantic cod on Georges Bank (GB, thick line) and in the Gulf of Maine (GoM, thin line) in thousands of metric tons (upper panel), and cod landings in the Gulf of Maine in relation to annual mean water temperature at Boothbay Harbor, ME (thick line) and Woods Hole, MA (thin line; lower panel)
Temperature affects virtually every aspect of the biology and ecology of cod, including its distribution (Dutil and Brander 2003; Drinkwater 2005); recruitment success, the number of young cod surviving to a specified age or size (Planque and Frédou 1999; Brander 2000; Drinkwater 2005); feeding (McKenzie 1934, 1938; Rose 2005); and individual growth (Brander 1995; Rätz and Lloret 2003). Cod populations are distributed throughout the North Atlantic in coastal and continental shelf waters characterized by seasonal bottom temperature regimes ranging from less than −1°C to over 20°C and annual mean temperatures from 2–12°C (Dutil and Brander 2003; Drinkwater 2005). Drinkwater (2005) used the 12°C threshold to assess potential loss of thermal habitat of cod throughout the North Atlantic. Increases in temperature have a positive effect on recruitment in arcto- boreal regions where temperatures are close to the lower thermal tolerance limit of cod, and negative effect on stocks inhabiting areas at the upper end of the temperature range (Planque and Frédou 1999; Drinkwater 2005). Stocks inhabiting regions characterized by intermediate temperature regimes showed no detectable effect of temperature.
In this paper we examine the response of cod to temperature changes observed on the Northeast Continental Shelf (NCS) over the last four decades to establish a baseline for assessing potential effects of future climate change on distribution and recruitment. This strategy of forecasting by historical analogy has been employed successfully for evaluating potential climate impacts on marine resources (Glantz 1990), including those off the northeastern USA (Murawski 1993). We then consider predicted regional changes in water temperature derived from three coupled atmosphere–ocean general circulation models under two CO2 emission scenarios and explore potential implications for cod off the northeastern USA. Finally, we develop a temperature-dependent production model for the Gulf of Maine cod to illustrate the interplay of harvesting and environmental change on stock dynamics.
2 Methods
2.1 Cod analysis
2.1.1 Distribution
We first considered cod abundance and distribution patterns on the NCS as a function of water temperature, depth, and time (year). Standardized research vessel surveys have been conducted in this US region by the Northeast Fisheries Science Center (NEFSC) in the autumn since 1963 and in the spring since 1968 (Smith 2002). A stratified random sampling design is employed with strata defined by latitude and bathymetry. Approximately 350 stations are occupied during each seasonal survey from the Gulf of Maine to Cape Hatteras, off the coast of North Carolina. At each sampling location, the catch of each species is enumerated and weighed and samples are taken for length and age composition, incidence of disease, maturity, and diet composition. Surface and bottom water temperatures and bottom depth are recorded at each station.
We modeled the presence or absence of cod as a function of bottom water temperature, depth, and year using stepwise logistic regression (Hosmer and Lemeshow 2000). Time (year) is included in the model to account for interannual changes in the abundance of cod that affect the probability of obtaining cod at each location. The model is given by:
where p is the probability of obtaining a catch of cod at a given sampling location, α is an intercept term, β is a coefficient related to the effect of temperature, δ is the effect of depth, κ is the effect of year on cod distribution, and ɛ is a binomially distributed random error term. We also tested for quadratic effects indicating environmental maxima or minima by including squared terms for water temperature in the model. Model parameters were estimated by the method of maximum likelihood.
We focused our analysis on cod distribution patterns in September and October because temperatures are near maximum at this time, and data over the past decade indicate the largest positive temperature anomalies in September and October in areas such as the Gulf of Maine (Fig. 3). The September–October survey also afforded the longest series available and spanned the broadest range of temperatures. We restricted our analysis to the region north of 40°N because cod were rarely observed in surveys south of this latitude. To evaluate potential impacts of further temperature increases, we developed a simplified model including just temperature and depth terms for data aggregated over the period 2000–2004 as a baseline and calculated the probability of declining cod occurrence with increasing temperature.

Monthly sea surface temperature anomalies in Boothbay Harbor, computed against the “20th century” (1905–1999) means. Note the shifting patterns of positive temperature anomalies during June, July and August early in the century to other seasons during the later part of the century. White boxes represent missing values
2.1.2 Cod production model
We examined temperature effects on the production of cod and the implications for resilience to exploitation in an age-structured population model for the Gulf of Maine stock. The model is similar in structure to that of Clark et al. (2003). We explicitly considered temperature effects on recruitment and individual growth. To test for the effect of water temperature on survival of cod during the early life history stages, we considered an extended Ricker stock-recruitment model (Quinn and Deriso 1999) that accounts for changes in adult population size and water temperature. The model can be expressed:
where R is recruitment (number of cod surviving to age 1) , S is cod spawning stock biomass, T is temperature, a is the rate of recruitment at low population densities, b is a coefficient reflecting compensatory processes, and c is the temperature coefficient. Cod eggs and larvae are found in the water column throughout the year, although they are most abundant from winter through mid-summer (Serchuk et al. 1994). We used the mean sea surface temperature from January–July measured at Boothbay Harbor, ME, USA to characterize temperature during the critical early life history period. For estimation, we employed a linearized form of the model:
where η t is a normally distributed random error term with mean zero and constant variance. Parameter estimates were determined by the method of maximum likelihood. We examined the autocorrelation function of the residuals to test for violations of the assumption of independence and we report the Durbin–Watson Statistic to test for first-order autocorrelation. We compared models with and without the temperature term using the Akaike Information Criterion (AIC). The AIC is a robust information theoretic measure that explicitly addresses the issue of model parsimony. Models with a greater number of parameters are assessed a penalty. The model with the lowest AIC score is deemed the most appropriate model among those tested based on the same series.
We used the model of Brander (1995) to represent the effect of temperature on individual growth:
where \( W_{\infty } \) is the asymptotic weight, a is age, T is the temperature experienced at each age and year and g and h are model parameters and ω is a normal random variable with zero mean and constant variance. To represent temperature, we used the January–July mean annual water temperature at Boothbay Harbor, ME, USA to match the series used in the recruitment analysis. Parameters were estimated using non-linear least squares regression.
The yield from a cohort of fish is given by:
where F and Z are instantaneous rates of fishing mortality and total mortality (mortality due to all sources), respectively; N a is the number in the cohort at age a ; r is the age at recruitment (r = 1 in this analysis); and a max is the maximum age (a max = 20 years in this analysis). We constructed total yield curves as a function of temperature and fishing mortality and assessed the interplay of potential decreases in recruitment and increases in individual growth with potential increases in water temperature.
2.2 Climate models and emission scenarios
2.2.1 Atmosphere–ocean general circulation models
Our analysis is based on simulations from three atmosphere–ocean general circulation models (AOGCMs): National Oceanographic and Atmospheric Administration Geophysical Fluid Dynamical Laboratory CM2.1 (GFDL) (Delworth et al. 2006), United Kingdom Meteorological Office Hadley Centre Coupled Model, version 3 (HadCM3) (Pope et al. 2000), and US Department of Energy/National Center for Atmospheric Research Parallel Climate Model (PCM) (Washington et al. 2000). These are three of the latest generation of numerical models that couple atmospheric, ocean, sea–ice, and land–surface components to represent historical climate variability and estimate projected long-term increases in global temperatures due to anthropogenic emissions. Atmospheric processes are simulated at a horizontal resolution of 2.5 × 2° (GFDL), 3.75° by 2.5° (HadCM3) and T42 or ∼2.8° by 2.8° (PCM; Table 1). Oceanic variables including salinity, potential temperature, and three-dimensional currents, simulated on a tri-polar grid by GFDL and PCM, were interpolated onto a regular 1 × 1° grid using depth-dependent areal weights. HadCM3 ocean simulations were performed on a regular 1.25 × 1.25° grid and required no interpolation. Climate sensitivity, a metric that captures the magnitude of the model-simulated increase in global temperature in response to a doubling of atmospheric CO2 concentration, is 1.5°C for GFDL, 1.3°C for PCM and 3.3°C for HadCM3, covering the low to medium part of the United Nations Intergovernmental Panel on Climate Change (IPCC) range of 1.5–4.5°C (IPCC 2001).
In recognition of known biases in water temperature and salinity fields in the Northwest Atlantic in the current generation of AOGCMs (Dai et al. 2002), we focused strictly on the anomalies predicted by the models. We first adjusted the model outputs for surface water temperature against observed temperature fields derived from hydrographic observations during NEFSC surveys for five North American geographical sectors (Southern Mid-Atlantic Bight; Northern Mid-Atlantic Bight, Georges Bank; Western Gulf of Maine; and Eastern Gulf of Maine; Fig. 1). From the adjusted surface temperature projections, we estimated bottom temperatures using observed surface to bottom water temperature relationships in each area. Although observations are made throughout the year, the most intensive sampling is in spring and autumn in conjunction with the bottom trawl surveys. Predictive relationships were developed by regressing surface temperature on bottom temperature in the season of interest and then checking for improvement in fit by adding the surface temperature for the other season. The seasonal and region-specific linear fits were applied to model-simulated spring and fall surface temperature anomalies for each region (relative to the 1970–2000 mean temperature) in order to produce predicted bottom temperature anomalies covering the full period of model simulations, from 1900 through 2099 (Fogarty et al. 2007)
2.2.2 Emission scenarios
Future simulations are forced by the IPCC Special Report on Emission Scenarios (SRES; Nakienovi et al. 2000) higher (A1fi), mid-high (A2) and lower (B1) emissions scenarios. These scenarios describe internally consistent pathways of future societal development and corresponding greenhouse gas emissions, and cover a wide range of alternative futures based on projections of economic growth, technology, energy intensity, and population. The A1fi and B1 scenarios used in this study bracket the range of SRES scenarios, and can be thought of as lower and higher bounds that encompass most, though not all, potential non-intervention emissions futures. At the higher end, rapid introduction of new technologies, extensive economic globalization, and a fossil-intensive energy path causes A1fi greenhouse gas (GHG) emissions to grow steadily throughout the century. In the A1fi scenario, CO2 emissions climb throughout the century, reaching almost 30 Gt/year or six times the 1990 levels by 2100. The A2 emissions scenario also reach 30 Gt/year by 2100, although cumulative emissions over the century are slightly lower than the A1fi scenario. Emissions under the B1 scenario are even lower, based on a world that transitions relatively rapidly to service and information economies. Carbon dioxide emissions in the B1 scenario peak at just below 10 Gt/year – around two times the 1990 levels – at mid-century and decline slowly to below current-day levels. Together, these scenarios represent the range of IPCC non-intervention emissions futures with atmospheric CO2 concentrations reaching approximately double and triple pre-industrial levels by 2100: 550 ppm in the B1scenario and 970 ppm in A1fi. Projections for the A1fi and B1 scenarios were used for the GFDL and PCM models, but as HadCM3 did not save any ocean output for its A1fi simulation, we used A2 and B1 instead.
3 Results
3.1 Cod baseline analyses
3.1.1 Cod distribution
Temperature strongly influences the presence or absence of cod on the Northeast Continental Shelf in samples derived from standardized NEFSC monitoring programs. Logistic regression models derived for autumn research vessel surveys indicate highly significant effects of temperature, depth, and year, with temperature by far the most important factor (Table 2). The inclusion of quadratic temperature terms did not improve the model fit and therefore were not further considered. The probability of obtaining cod at a specified location decreased markedly with increasing bottom water temperature. Catch probabilities also decreased with increasing depth. Finally, a significant year effect was observed with declining probability of obtaining cod over time, reflecting declines in overall abundance during the last four decades.
3.1.2 Cod recruitment and growth
Temperature significantly affected recruitment of cod in the Gulf of Maine. Recruitment success of cod declined with increasing water temperature, and inclusion of the temperature term in the recruitment model substantially improved the fit of the model as indicated by the AIC score (Table 3). The test for first order autocorrelation in the residuals for the model without temperature effects was statistically significant, indicating a deficiency in the model structure. This deficiency was rectified by including the temperature term in the model. The model for cod weight at age as a function of the interaction of age and temperature was highly significant (F = 1876; p < 0.0001; W4 = 18.318, g = 0.508, h = 65.791).
3.2 Bottom water temperature projections
Dramatic differences are evident for the low and high emission scenarios for projected bottom temperature anomalies. For the low emission scenario in particular, substantial differences are noted among models and over the time course of the simulations. Ensemble averages over the three models for projections of bottom water temperature anomalies by region for the period 2080–2084 are provided in Table 4.
Under the low emission scenario, the average March–April bottom temperature anomalies (°C) range from 0.89 (Northern Mid-Atlantic) to 1.33 (western Gulf of Maine) and, in September–October, range from 1.17 (Eastern Gulf of Maine) to 1.48 (Western Gulf of Maine). For the low emission scenario, the March–April and September–October projected temperature anomalies are less than 1.5°C and are relatively consistent among areas. For the high emission scenario, the projected March–April bottom temperature anomalies range from 2.04 (eastern Gulf of Maine) to 3.64 (Northern Mid-Atlantic) and, during March–April, from 1.96 (eastern Gulf of Maine) to 3.48 (Georges Bank). Relative to the low emissions scenario, wider regional differences are evident in these projections with substantially higher projected bottom temperature increases in southern areas. Projected anomalies for four time periods for the three models under the high emissions scenario are depicted in Fig. 4 to illustrate the projected trajectories of change.

Projected average bottom water temperature anomalies (°C) for four time periods (2020–2024, 2040–2044, 2060–2064, and 2080–2084) in March–April and September–October by region under the high emission scenarios for the geophysical fluid dynamics (GFDL), Hadley (HAD, two scenarios combined) and parallel climate (PCM) models
3.3 Implications for future cod distribution and production
3.3.1 Cod distribution
The relative magnitude of further declines in the probability of cod occurrence with reference to 2000–2004 baseline conditions is depicted in Fig. 5. This figure can be used to determine the relative decline in the probability of obtaining cod at a given location under increasing bottom temperature levels under the assumption that abundance levels remain relatively close to the baseline levels. This assumption implies a redistribution of cod to areas characterized by colder water temperatures as a function of depth and/or latitude.

Predicted probability of obtaining cod at a given location in Northeast Fisheries Science Center September–October surveys as a function of temperature. Results are for a 2000–2004 baseline period
If the bottom temperature projections under the high emission scenario are realized, it is likely that the 12°C mean annual bottom water temperature threshold for cod (Drinkwater 2005) will be exceeded in the Mid-Atlantic Bight and will approach the threshold level on Georges Bank regions by the end of this century. The mean bottom water temperature in the Gulf of Maine as a whole during the period 1978–2002 was 7.1°C, and if the bottom temperature anomaly projections for this region hold, the 12°C threshold will not be exceeded.
3.3.2 Cod production in the Gulf of Maine
Cod yields predicted under projected increased water temperatures relative to baseline conditions during 1982–2003 in the Gulf of Maine are depicted in Fig. 6. The effect on yield reflects the interplay of decreased recruitment and increased individual growth with increased water temperature. Increased water temperatures of 1°C and 2°C relative to the baseline levels, corresponding approximately to projected increases in under the lower and higher emission scenarios by the end of the century (2080–2084), result in declines in projected yields and in the fishing mortality rate resulting in maximum yield. A 1°C increase would result in an approximate 21% decline in maximum yield while a 2°C temperature increase is predicted to result in a 43% decline in maximum yield. At the higher temperature level, the stock is predicted to go extinct at fishing mortality rates in excess of F = 1.6, a level which is sustainable (though far from optimal) under a 1°C increase in temperature. These results qualitatively point to a decrease in yield and resilience to fishing under increasing temperatures, particularly under the projected levels under the high emission scenario.

Predicted cod yield (thousands of metric tons) from the Gulf of Maine as a function of fishing mortality under mean temperature conditions during 1982–2003 (upper curve baseline) and temperature increases of 1°C (middle curve) and 2°C (lower curve) relative to the baseline and corresponding approximately to projected increased in under the low and high emission scenarios by the end of the century (2080–2084). The fishing mortality rate is an index of the fraction of the population caught in a given year
4 Discussion
Future climate predicted under increasing anthropogenic emissions of greenhouse gases has the potential to significantly alter the physical structure of the oceans, with direct implications for marine ecosystems and human societies (IPCC 2001; Stenseth et al. 2002; Sarmiento et al. 2004; Behrenfeld et al. 2006). On the NCS, potential changes include warmer water temperatures in all seasons, increased density stratification, changes in the slope water system affecting the continental shelf and Gulf of Maine, and attendant changes in nutrient and plankton dynamics and higher trophic levels (e.g., Greene and Pershing 2001; Pershing et al. 2005). These changes need to be considered in the context of impacts resulting from other human activities such as fishing, pollution, and habitat loss due to coastal development. Climate change can interact with other anthropogenic changes to alter the fundamental production characteristics of marine systems (Stenseth et al. 2002). Climate change will potentially exacerbate the stresses imposed by harvesting and other human activities. Considerable emphasis is now being placed on understanding the causes and consequences of climate change in temperate marine systems (Harvell et al. 2002; Helmuth et al. 2002; Barnett et al. 2005; Drinkwater 2005; Sutton and Hodson 2005; Altieri and Witman 2006) to prepare for anticipated alterations in ecosystem structure and function.
Substantial uncertainty exists in projections of the effects of climate change in the marine environment off the northeastern USA. Existing coupled AOGCMs do not resolve circulation and other important dynamical physical processes on continental shelves, and they exhibit known biases with respect to predicted water mass properties in the northwestern Atlantic Ocean. Nonetheless, these models provide important insights into possible directional changes in important hydrographic features, particularly relative temperature change. We have attempted to reduce the effects of bias by calibrating model output against direct observation for the period of instrumental record, and then considering only the relative changes in predicted properties. We used GCM model output from present-day and future runs to estimate temperature change over time, and then added the predicted difference to the current-day observations. The projections examined in this report must be viewed as provisional estimates until the output from AOGCMs is linked to appropriate shelf sea models. Previous attempts to examine the effects of global climate change on marine resources in this region have utilized projections of atmospheric temperatures (e.g., Drinkwater 2005) or sea surface temperature estimates derived from AOGCMs (e.g., WWF 2005). We have attempted to refine these approaches by also examining potential changes in bottom water characteristics.
A full assessment of potential climate impacts on the marine environment of continental shelves ultimately requires coupling of the results of Ocean General Circulation Models with higher-resolution hydrodynamic models in a fully nested structure. Such coupling properly accounts for responses of the shelf system to the combined effects of forcing at the boundaries with local forcing and adjustments. In the northeastern USA, such a nested approach is necessary to capture features such as topography and vertical mixing which affect circulation and the distributions of temperature, salinity, nutrients and plankton. This need is most pronounced in topographically complex areas such as the Gulf of Maine where a full accounting of factors affecting stratification and convective overturn will be required to understand the direction and magnitude of bottom temperature change.
Under the high emission scenario, cod appear vulnerable to a loss of thermal habitat in the Mid-Atlantic Bight. Thermal habitat losses also are expected on Georges Bank, historically the dominant production region for this species on the NCS. Within the Gulf of Maine, projected bottom temperature anomalies are lower than those for Georges Bank and south and the baseline level is lower. Accordingly, lesser impacts on thermal habitat are expected in this stock area, although some redistribution to cooler water areas within the Gulf can be anticipated, and is subject to other habitat constraints such as substrate preferences.
Consideration of potential impacts of temperature increases on yield and resilience to fishing for the Gulf of Maine indicates that although thermal habitat will generally remain viable, losses in stock productivity are predicted. A negative impact of temperature on recruitment is indicated for this stock, offsetting potential increases in individual growth with increasing temperature and resulting in a predicted loss in yield with increasing temperature. Temperature increases are also predicted to result in reduced resilience to fishing, highlighting the interplay between harvesting and environmental determinants of production. It should be noted that the empirical analysis of temperature effects on cod recruitment and growth presented here is set within a broader context of ecosystem change such as increased pelagic fish biomass which may also affect cod recruitment. It is therefore not possible to fully separate the effects of temperature and other ecosystem changes on cod productivity without further information derived from actively adaptive ecosystem-based management. The results presented here therefore are best considered a guide to the direction and potential magnitude of change in yield if other aspects of cod productivity remain relatively constant (e.g. continued availability of preferred prey resources etc.).
Because cod are comparatively well-studied and at the southern extent of their range in our study area, it is possible to make reasonable assessments of their likely response to temperature change. Marine systems and the life histories of marine organisms are complex, however, and we are at very early stages in our ability to predict climate change effects that integrate across all parts of the ecosystem. We have focused on temperature effects because of the substantial body of information indicating the central role of temperature on cod physiology and ecology. This general approach was adopted in climate impact assessments for cod in other regions (e.g. Clark et al. 2003; Drinkwater 2005). However, temperature is not the sole determinant of cod production. The effects of climate change on other aspects of ecosystem structure and function can be expected to exert additional impacts on cod dynamics and a full assessment of the impact on cod and other resource species will require consideration of overall changes in system productivity under global climate change.
References
Altieri AH, Witman JD (2006) Local extinction of a foundation species in a hypoxic estuary: integrating individuals to ecosystem. Ecology 87(3):717–730
Barnett TP, Pierce DW, AchutaRao KM et al (2005) Penetration of human-induced warming into the world’s oceans. Science 309:284–287
Behrenfeld MJ, O’Malley RT, Siegel DA et al (2006) Climate-driven trends in contemporary ocean productivity. Nature 444:752–755
Brander K (1995) Effect of temperature on growth of Atlantic cod (Gadus morhua L). ICES J Mar Sci 52:1–10
Brander K (2000) Effect of environmental variability on growth and recruitment of Atlantic cod (Gadus morhua L) using a comparative approach. Oceanologica Acta 23:485–496
Clark RA, Fox CJ, Viner D, Livermore M (2003) North Sea cod and climate change-modeling the effects of temperature on population dynamics. Global Change Biol 9:1669–1680
Dai A, Hu A, Meehl WM, Washington WM, Strand WG (2002) Atlantic thermohaline circulation in a coupled general circulation models: unforced variations versus forced changes. J Climate 18:3270–3293
Delworth TL, Broccoli AJ, Rosati A, Stouffer RJ, Balaja V, Beesley JA, Cooke WF, Dixon KW, Dunne J, Dunne KA, Durachta JW, Findell KL, Ginoux P, Gnanadesikan A, Gordon CT, Griffies SM, Gudgel R, Harrison MJ, Held IM, Hemler RS (2006) GFDLs CM2 global coupled climate models. Part 1 – Formulation and simulation characteristics. J Clim 19:643–674
Drinkwater KF (2005) The response of Atlantic cod (Gadus morhua) to future climate change. ICES J Mar Sci 62:1327–1337
Dutil J-D, Brander K (2003) Comparing productivity of North Atlantic cod (Gadus morhua) stocks and limits to growth production. Fish Oceanogr 12:502–512
Fogarty M, Incze L, Wahle R, Mountain D, Robinson A, Pershing A, Hayhoe K, Richards A, Manning J (2007) Potential climate change impacts on marine resources of the northeastern United States. Report to Union of Concerned Scientists
Glantz MH (1990) Does history have a future? Forecasting climate change effects on fisheries by analogy. Fisheries 15:39–44
Greene CH, Pershing AJ (2001) The response of Calanus finmarchicus populations to climate variability in the Northwest Atlantic: basin-scale forcing associated with the North Atlantic Oscillation (NAO). ICES J Mar Sci 57:1536–1544
Harvell CD, Mitchell CE, Ward JR et al (2002) Climate warming and disease risks for terrestrial and marine biota. Science 296(5576):2158–2162
Helmuth B, Harley CD, Halpin PM et al (2002) Climate change and latitudinal patterns of intertidal thermal stress. Science 298:1015–1017
Hoagland P, Jin D, Thunberg E, Steinback S (2005) Economic activity associated with the Northeast shelf large marine ecosystem: application of an input–output approach. In: Hennessy TM, Sutinen JG (eds) Large marine ecosystems, vol 13, pp 157–179
Hosmer DW, Lemeshow S (2000) Applied logistic regression, 3rd edn. Wiley, New York, p 375
IPCC (2001) Intergovernmental Panel on Climate Change: Climate change 2001: synthesis report. Summary for Policy Makers. Third Assessment Report
Kurlansky M (1997) Cod: a biography of the fish that changed the world. Walker, New York, p 294
McKenzie RA (1934) Cod and water temperature. Biol Board Can Atlantic Progress Report 12:3–6
McKenzie RA (1938) Cod take smaller bites in ice cold water. Fish Res Board Can Atlantic Progress Report 22:12–14
Murawski SA (1993) Climate change and marine fish distributions: forecasting from historical analogy. Trans Am Fish Soc 122:667–658
Nakienovi N, Alcamo J, Davis G et al (2000) Special report on emissions scenarios: a special report of Working Group III of the Intergovernmental Panel on Climate Change. Cambridge Univ Press, Cambridge, UK
Pershing AJ, Greene CH, Jossi JW et al (2005) Interdecadal variability in the Gulf of Maine zooplankton community with potential impacts on fish recruitment. ICES J Mar Sci 62:1511–1523
Planque B, Fredou T (1999) Temperature and the recruitment of Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 56:2069–2077
Pope VD, Gallani ML, Rowntree PR, Stratton RA (2000) The impact of new physical parameterizations in the Hadley Centre climate model – HadCM3. Clim Dyn 16:123–146
Quinn TJ, Deriso R (1999) Quantitative fish dynamics. Academic, New York
Ratz H-J, Lloret J (2003) Variation in fish condition between Atlantic cod (Gadus morhua) stocks, the effect on their productivity and management implications. Fish Res 60:369–380
Rose GA (2005) On distributional response of North Atlantic fish to climate change. ICES J Mar Sci 62:1360–1374
Sarmiento JL, Slater R, Barber R et al (2004) Response of ocean ecosystems to climate warming. Global Biogeochem Cycles 18:GB3003
Serchuk FM, Grosslein MD, Lough RG et al (1994) Fishery and environmental factors affecting trends and fluctuations in the Georges Bank and Gulf of Maine Atlantic cod stocks: an overview. ICES Mar Sci Symp 198:77–109
Smith TD (2002) The Woods Hole bottom-trawl resource survey: development of fisheries-independent multispecies monitoring. ICES Mar Sci Symp 215:480–488
Steinback S, Gentner B, Castle J (2004) The economic importance of marine angler expenditures in the United States. NOAA Prof. Pap NMFS 2 167 pp
Stenseth NC, Mysterud A, Ottersen G et al (2002) Ecological effects of climate fluctuations. Science 297:1292–1296
Sutton RT, Hodson DLR (2005) Atlantic Ocean forcing of North American and European summer climate. Science 309:115–118
Washington WM, Weatherly JW, Meehl GA et al (2000) Parallel climate model (PCM) control and transient simulations. Clim Dyn 16:755–774
World Wildlife Fund (2005) Implications of a 2°C global temperature rise for Canada’s natural resources
Acknowledgments
We thank Nick Wolff, Michelle Traver and Loretta O’Brien for data and analyses and Adrienne Adamek for GIS support. The support and encouragement of Erika Spanger-Siegfried, the Union of Concerned Scientists, and the Census of Marine Life are also gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fogarty, M., Incze, L., Hayhoe, K. et al. Potential climate change impacts on Atlantic cod (Gadus morhua) off the northeastern USA. Mitig Adapt Strateg Glob Change 13, 453–466 (2008). https://doi.org/10.1007/s11027-007-9131-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11027-007-9131-4