
Abstract
Rosa majalis Herrm. (Rosaceae) is a shrub widespread throughout Eurasia. In West Siberia, it is cultivated as an ornamental plant and raw material for the manufacture of fruit multivitamin concentrates. Meanwhile, metabolites and biological activities of its roots are poorly studied. The aim of this investigation was to evaluate the potential of ethyl acetate (EtOAc), butanol (ButOH), aqueous ethanol (EtOH), and aqueous (H2O) extracts of R. majalis roots as sources of antioxidant and anti-influenza compounds. Spectrophotometry and HPLC were used for metabolite quantification. Antioxidant activities were determined via the 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay. The antiviral activities were assessed as half-maximal inhibitory concentrations (IC50). The EtOAc extract manifested the highest concentrations of phenolic compounds (53.79 mg g−1 of dry extract) and saponins (48.92 mg g−1). Total gallic acid was the main phenolic compound in all extracts (24.98–38.68 mg g−1). Catechin, free ellagic acid, and total ellagic acid had the highest levels in the EtOAc, H2O, and EtOH extracts, respectively (30.83, 22.38, and 34.95 mg g−1). The EtOAc extract showed the highest free-radical scavenging activity (2.31 trolox equivalents) and anti-influenza effects (IC50 61.8 and 50.3 µg mL−1) against A/Aichi/2/68 (H3N2) and A/Chicken/05/2005 (H5N1), respectively. Our findings indicate good potential of these extracts for drug design against free-radical damage and influenza virus.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Rosa majalis Herrm. (R. cinnamomea L.) is a species of natural flora of most of Eurasia. According to The State Pharmacopeia of the Russian Federation (14 edition), the fruits of this species, along with other five rosehip species, are the source of ascorbic acid, carotenoids, and flavonoids, which are used for industrial production of vitamin teas, oil, and choleretic drug Holosas (Rosae fructus 2018). Secondary metabolites of leaves and fruits of rosehip are well studied, including ecological and geographic variation, extraction methods, and antioxidant activity (Nowak and Gawlik-Dziki 2007; Safonova et al. 2012; Polumackanycz et al. 2020; Jiang et al. 2022). Diversity of phenolic acids has been revealed: caffeic, gallic, chlorogenic, rosmarinic, p-coumaric, sinapic, and benzoic acids; coumarins (esculin); ellagic acid and its derivatives; glycosides of flavones (luteolin, vitexin, and baicalein), flavonols (quercetin and kaempferol), and flavanones (naringenin). Meanwhile, studies on the roots are only beginning. Detailed identification and quantification of phenolic compounds in the roots of R. heckeliana have been performed by Çoruh and Özdoğan (2015). In that study, catechin, ellagic acid, and caffeic acid were found to be major phenolic compounds in these root samples. Catechins are also the most abundant phenolic compounds in the Rosa acicularis roots (Olennikov et al. 2021). Meanwhile, benzoic acid and 3-protocatechuic aldehyde are reported to be the main phenolic compounds in the root extract of R. pimpinellifolia (Güven et al. 2021).
Hydrolyzable tannins are considered constant components of plants of the family Rosaceae (Olennikov et al. 2021); among them, the diversity of ellagitannins has been revealed (Cai et al. 2005). Ellagic acid is attracting attention due to its antioxidant, anti-inflammatory, antimutagenic, and antiproliferative properties. It has exerted pharmacological effects in various in vitro and in vivo model systems. Furthermore, it has been well documented that ellagic acid possesses antiallergic, antiatherosclerotic, cardioprotective, hepatoprotective, nephroprotective, and neuroprotective properties (Sharifi-Rad et al. 2022). Ellagitannins usually reach high compared to leaves, flowers, and roots (Cendrowski et al. 2017). Besides, ellagitannins have an antiviral effect on influenza virus replication (Saha et al. 2010). In the roots of plants from the family Rosaceae, condensed tannins have also been found (Olennikov et al. 2021).
Chemical structure and biological activities of saponins from the roots of representatives of the genus Rosa are poorly investigated. Anti-inflammatory activity of triterpenoid saponins from Rosa rugosa fruits was discovered a decade ago (An et al. 2011). In recent decades, a lot of studies on saponins of the other genus from the family Rosaceae (Potentilla) have been conducted, and their cardioprotective (Enayati et al. 2022), cytotoxic (Walia et al. 2016; Wu et al. 2022), antitumor (Guo and Gao 2013), and anti-hepatitis B virus (Zhao et al. 2008) properties have been revealed. In addition, triterpenoid saponins from some plants exert an antiviral effect on human coronavirus (Cheng et al. 2006).
Biological activity of plant extracts has been shown to depend on the profile of secondary metabolites, which in turn is determined by the solvent and extraction procedure. Solid–liquid extraction methods with different types of solvents are the most commonly used procedures to extract diverse valuable compounds from plant tissues (Stalikas 2007).
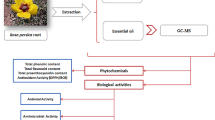
In the present study, we aimed to determine profiles of secondary metabolites of R. majalis roots in terms of composition of their ethyl acetate (EtOAc), butanol (ButOH), aqueous ethanol (EtOH), and aqueous (H2O) extracts as sources of antioxidant and anti-influenza compounds.
2 Materials and methods
2.1 Plant material and extraction
The roots of R. majalis plants from “Collections of living plants indoors and outdoors” of the Central Siberian Botanical Garden of the Siberian Branch of the Russian Academy of Sciences (CSBG SB RAS; Novosibirsk, Russia) were used as plant material for the extraction of secondary metabolites. H2O, EtOH 80% v/v, ButOH, and EtOAc extracts were prepared. For each extraction, dry powdered roots were treated three times with appropriate volumes of solvents (Table 1) for 30 min in a water bath (65 °C) in a round bottom flask fitted with a reflux condenser.
Each combined extract was passed through a glass funnel filter with a sintered glass disk. After cooling, the filtrates were concentrated using a rotary evaporator (Ika RV3 V, Germany) to remove the solvent. After organic solvents’ evaporation, the EtOH and ButOH extracts were treated with chloroform to remove fatty substances, and the H2O extract was directly incubated with chloroform without concentration. Defatted extracts were dried in a vacuum drying cabinet (LP-402, Hungary) to obtain powder residues. When the EtOAc extract was made, the solvent from the combined extract was being removed until the beginning of precipitation. After that, sixfold volume of chloroform was added with stirring, and the resultant precipitate (the EtOAc extract) was dried in vacuum similarly to the above procedure and used for phytochemical profiling and estimation of biological activity (Table 1). The residual moisture content of the obtained extracts was 3.0–3.5%.
2.2 Spectrophotometric analyses
For the assessment of total catechin concentrations, a vanillin assay was performed. A standard curve was constructed with ( +)-catechin from Sigma-Aldrich (Saint Louis, MO, USA) in the concentration range 0.001–0.02 mg mL−1. Precisely weighed dry extract (10–15 mg) was dissolved in 10 mL of 96% ethanol, and 0.8 mL of the sample was placed into two tubes. Then, 4 mL of 1% (w/v) vanillin in concentrated hydrochloric acid was added into the tube with the sample, and an equal volume of concentrated hydrochloric acid was added into the other tube (control). Total volume of the reaction mixture was brought to 5 mL with concentrated hydrochloric acid. The reaction was carried out at room temperature (~20 °C) for 5 min. Absorbance of the extract and standard solution was measured at 502 nm (Sun et al. 1998; Kaidash et al. 2022) by means of an SF-56 spectrophotometer (Lomo, St. Petersburg, Russia). The results were expressed as milligrams of (+)-catechin equivalent per gram of dry extract.
Total flavonol concentration was estimated by a spectrophotometric method based on the reaction of complex formation between flavonols and aluminum chloride (Kaidash et al. 2022). An extract (0.1 mL) was placed into two 5 mL test tubes. Into the first tube, 0.2 mL of a 2% ethanol solution of aluminum chloride was added, and 1–2 drops of 30% acetic acid were added into the second, and the volumes of the reaction solutions were adjusted to 5 mL with 96% ethanol. Next, the solutions were shaken, and after 40 min, the absorbance of the first solution was registered at a wavelength of 415 nm on the spectrophotometer. The second solution served as a control sample. A calibration curve was built with rutin from Chemapol (Mumbai, MH, India) in the concentration range 0.01–0.1 mg mL−1. The results were expressed as milligrams of rutin equivalent per gram of dry extract.
Quantitation of tannins (hydrolyzable tannins) was performed by their reaction with ammonium molybdate (Fedoseeva 2005). An extract (10 mL) was transferred into a 100 mL volumetric flask, and 10 mL of a 2% aqueous solution of ammonium molybdate was added. The volume of the flask content was brought to the nominal volume with purified water, and the mixture was incubated for 15 min. The intensity of the resulting color was measured at 420 nm. A calibration curve was constructed with tannin (Sigma, St. Louis, MO, USA) in the range 0.01–0.1 mg mL−1. The results were expressed as milligrams of tannin equivalent per gram of dry extract.
For determining the total level of phenolic acids, Arnow’s test was performed (Jain et al. 2017). To 1 mL of an extract, 5 mL of distilled water was added. Into this solution, 1 mL of 0.5 M HCl, 1 mL of Arnow’s reagent (10.0 g of sodium molybdate and 10.0 g of sodium nitrate in 100.0 mL of distilled water), and 1 mL of 1 M NaOH were added sequentially. The volume was brought to 10 mL using distilled water, and absorbance was measured at 490 nm. A blank sample was prepared too, which contained all reagents except the extract. A calibration curve was built using caffeic acid (Serva, Heidelberg, Germany) in the range 0.02–0.1 mg mL−1. The results were expressed as milligrams of caffeic acid equivalent per gram of dry extract.
The total phenolic concentration was determined with the help of the Folin–Ciocalteu reagent (Blainski et al. 2013). An extract (0.5 mL) was placed in a 5 mL volumetric flask, then 2.5 mL of the Folin–Ciocalteu reagent (diluted 1:10 with distilled water, v/v) and 2 mL of a 7.5% aqueous solution of sodium carbonate were added, and the flask was shaken well. In a blank sample, the reagents were combined with distilled water instead of the extract. The mixture was kept at 45 °C in a water bath for 15 min. Absorbance was measured at 765 nm (Kaidash et al. 2022). A standard curve was constructed with gallic acid from Sigma-Aldrich (Saint Louis, MO, USA) in the concentration range 0.002–0.01 mg mL−1. The results were expressed in milligrams of gallic acid equivalent per gram of dry extract.
For quantification of total saponins, a direct spectrophotometric method was employed based on a reaction between triterpene saponins and sulfuric acid (Pisarev et al. 2009). A precisely weighed dry extract (0.5–1.0 g) was dissolved in 10 mL of a mixture of glacial acetic acid, hydrochloric acid, and purified water (3.5:1.0:5.5) for hydrolysis of glycosides via 2 h incubation in a boiling water bath. Then, the hydrolysate was cooled, diluted with distilled water in a 1:2 ratio, and filtered. The residue on the filter was rinsed with distilled water, put with the filter into a volumetric 25 mL flask, and dissolved in 25 mL of hot ethanol. To 1 mL of this ethanol solution, 4 mL of concentrated sulfuric acid was added, and absorbance at 310 nm was measured after 10 min. Concentrated sulfuric acid served as a blank. To calculate saponin concentration, a calibration curve for oleanolic acid (Sigma-Aldrich, Saint Louis, MO, USA) was built in the range 0.01–0.4 mg mL−1. The results were expressed as milligrams of oleanolic acid equivalent per gram of dry extract.
All the aforementioned assays were performed on three biological and three technical replicates.
To assess the levels of these metabolites in the roots, their concentrations were calculated from extract concentrations and weights of roots.
2.3 HPLC analysis
Samples for HPLC were preparing by dilution of 10 mg of a dry extract in 10 mL of 50% aqueous ethanol (v/v). The solutions were diluted with double-distilled water at a 1:3 (v/v) ratio and passed through a C 16 cartridge (BioChemMack, Moscow, Russia). HPLC analysis of the resultant extracts was performed on an Agilent 1200 instrument with a diode array detector and the ChemStation software for data processing (Agilent Technologies, USA). The chromatographic separation was performed at 25 °C on a Zorbax SB-C18 Column (4.6 × 150 mm, 5 μm internal diameter). The mobile phase consisted of MeOH (solvent A) and 0.1% orthophosphoric acid in water (solvent B).
Separation of phenolic compounds in the extracts was carried out via gradient 1 (Table 2). To characterize the flavonoid aglycone profile and evaluate the concentrations of bound forms of gallic and ellagic acids, extracts were hydrolyzed with 2N HCl for 1 h in a water bath. The hydrolyzed extracts were passed through a C 16 cartridge and separated via gradient 2 (Table 2). For separation of catechins, gradient 3 was utilized (Table 2). The flow rate for all these gradients was set to 1 mL min−1. The sample injection volume was 10 μL, and absorbance was measured at 220, 255, 270, 290, 325, 340, 360, and 370 nm. The quantification of phenolic compounds was conducted by the external standard method. Identification of the known compounds was carried out by comparing retention times and UV spectra of the analyzed peaks with the peaks of standard samples of gallic acid from Fluka (Munich, Germany); ellagic acid, (±)-catechin, isoquercitrin, and rutin from Sigma-Aldrich (Saint Louis, MO, USA); and epigallocatechin gallate and l-epicatechin from Teavigo (Gevelsberg, Germany). Standard stock solutions at a concentration of 1 mg mL−1 in methanol were employed to build calibration curves in the concentration range of 2–100 μg mL−1. Quantification of unidentified hydroxybenzoic acid 9 was conducted based on the calibration curve of gallic acid, and concentration of flavonol glycoside 10 was determined by means of the isoquercitrin calibration curve. Concentrations of the compounds in the extracts were expressed in milligrams per gram of dry mass of an extract. To assess the levels of the compounds in the roots, their concentrations were calculated in milligrams per gram of dry mass of the roots.
2.4 Antioxidant activity
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay was carried out to evaluate the ability of the extracts to scavenge free radicals of DPPH (Kaidash et al. 2022). For the assay, a 2 mL aliquot of an extract was mixed with 3 mL of a DPPH solution (62 µg mL−1 in ethanol). After 40 min incubation in the dark at room temperature, absorbance (A) was measured at 517 nm against a blank. Free-radical scavenging activity was calculated as percentage inhibition via the following formula:
where Ablank is the absorbance of the control solution (containing all reagents except the tested extracts) and Asample is the absorbance of the sample. The results were expressed as half-maximal inhibitory concentration (IC50), defined as the antioxidant concentration that causes a 50% loss of DPPH radical concentration. These data were also converted to trolox equivalents. IC50 values were calculated by means of GraphPad Prism v.6.01 (GraphPad Software, USA). Solutions of trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) and ascorbic acid (concentrations 2.5–50.0 µg mL−1) served as a positive control.
2.5 Antiviral assay
Influenza virus strains A/chicken/Kurgan/05/2005 (H5N1) and A/Aichi/2/68 (H3N2) were obtained from the State Collection of Viral Infections and Rickettsioses Agents at the State Research Center of Virology and Biotechnology VECTOR (Koltsovo, Novosibirsk Oblast, Russia). All experiments with infectious materials were conducted at the State Research Center of Virology and Biotechnology VECTOR (Teplyakova et al. 2012). The virus strains were cultivated in 10-day embryonated chicken eggs. The virus concentration in the samples was determined by their titration in cultured Madin-Darby canine kidney (MDCK) cells according to the Spearman–Kärber algorithm (Spearman 1908; Kärber 1931).
The antiviral activity of the R. majalis root extracts was determined as IC50, i.e., the concentration of the extract in the culture medium at which 50% of MDCK cells are preserved in the infected monolayer. The cytotoxic activity of the extracts was evaluated in a monolayer of cultured MDCK cells in 96-well plates containing the RPMI 1640 medium (Biolot, Russia) supplemented with 5% of fetal bovine serum (HyClone, USA). To determine the cytotoxicity, 10 mg of a dry extract was dissolved in 1 mL of RPMI 1640 without serum and diluted 5-, 10-, 20-, 40-, 100-, and 1000-fold with the same medium. Aliquots of each dilution (100 mL) were placed in plate wells (four wells for each dilution) and incubated at 37 °C, 5% CO2, and 100% humidity for 2 days. Cytotoxic effects of the extracts were assessed under an inverted microscope. Maximum nontoxic concentration was defined as a concentration at which 100% of the MDCK cells stayed alive in the monolayer, and a half-maximal toxic concentration (TC50) meant the concentration of the extract causing the death of 50% of cells in the monolayer in each well. The selectivity index of the extract (SI) was calculated as the TC50/IC50 ratio (Al Jabri et al. 1996).
2.6 Statistical analysis
Concentrations of total of catechins, flavonols, tannins, phenolic acids, and phenolic compounds and of individual phenolic compounds in the roots and in the root extracts were expressed in milligrams per gram of dry mass of the roots or dry root extract, respectively. IC50 in the DPPH assay is presented as micrograms of dry extract per milliliter of the solvent (µg mL−1). The dry matter percentage was estimated by the gravimetric method.
The data were analyzed in the STATISTICA 10.0 software (Statsoft Inc., Tulsa, OK, USA), are reported as mean ± standard error from three biological and three technical replicates, and were compared by Duncan’s multiple-range test. Differences between the means were considered statistically significant at P < 0.05.
3 Results
3.1 Secondary metabolite contents and antioxidant activity
Each of the four obtained extracts had a distinct profile of secondary metabolites. The highest yield of dry matter from extracted roots was shown by the EtOH extract (32.04%). It significantly exceeded this parameter of the H2O extract, exceeded this parameter of the ButOH extract by more than twofold, and exceeded that of the EtOAc extract by more than fivefold (Table 1). Meanwhile, concentration of total catechin in the EtOAc extract (43.23 mg g−1) was the highest and exceeded this parameter in the other extracts by threefold to fourfold (Table 3). The lowest catechin content was found in the EtOH extract. The ButOH extract had the greatest content of flavonols (9.13 mg g−1). It was greater than that in the H2O extract by almost twofold and greater than that in the EtOH and EtOAc extracts by approximately 5.5-fold.
Tannin content varied slightly among the extracts. The maximum (48.19 mg g−1; in the EtOAc extract) exceeded the minimum (31.83 mg g−1; in the EtOH extract) by 1.5-fold. Phenolic acids, just as catechins, were most abundant in the EtOAc extract (57.56 mg g−1). In the H2O and EtOH extracts, phenolic acid levels were lower than the aforementioned maximum by approximately twofold, and in the ButOH extract, they were lower > 5 times. The total phenolic content corresponded to the concentration of phenolic acids; both parameters were the highest in the EtOAc extract (53.79 and 57.56 mg g−1) and the lowest in the ButOH extract (16.46 and 11.40 mg g−1). The saponin content was also consistent with the total phenolic content: the saponin content had a maximum in the EtOAc extract (48.92 mg g−1) and a minimum in the ButOH extract (16.13 mg g−1).
Thus, the EtOAc extract showed abundance of most classes of the tested secondary metabolites except for flavonols, whose concentration was the highest in the ButOH extract.
The EtOAc extract also manifested the highest DPPH radical scavenging activity (16.18 µg mL−1), whereas the H2O and ButOH extracts were slightly inferior (24.40 and 24.24 µg mL−1, respectively), and the EtOH extract contained the lowest radical scavenging activity (105.33 µg mL−1) (Fig. 1).

Radical scavenging activity of the extracts of the roots of R. majalis: half-maximal inhibitory concentrations of the extracts (IC50) (A) and trolox equivalents (B). IC50 values for trolox and ascorbic acid were 7.47 ± 0.01 and 8.69 ± 0.16 µg mL−1, respectively
Evaluation of contents these secondary metabolites in dry mass of the roots enabled us to compare recovery levels (completeness of the extraction) among the solvents (Table 3). Aqueous ethanol yielded the highest recovery of the majority of the assayed compounds (catechins, tannins, phenolic acids, total phenolics, and saponins), whereas butanol most completely extracted flavonols.
Chromatographic profiling confirmed the EtOAc extract as containing the highest level of phenolic compounds (Table 4). In this extract, the highest concentrations of gallic acid, including its free form (38.68 and 2.79 mg g−1, respectively), and catechin (30.83 mg g−1) were revealed. They were the main phenolic compounds in this extract and, along with total ellagic acid, contributed to the total phenolic content of all tested extracts the most. Total ellagic acid was dominant in the EtOH extract (34.95 mg g−1). Meanwhile, ellagic acid in free form and total gallic acid were major phenolic compounds in the H2O extract (22.38 and 32.64 mg g−1, respectively). This extract also showed higher—as compared to the other extracts—levels of ellagic acid glycoside (16.10 mg g−1). Noteworthy is the presence of two minor phenolic compounds in the ButOH and EtOAc extracts, which were absent in the H2O and EtOH extracts: hydroxybenzoic acid 9 (4.87 and 11.91 mg g−1) and flavonol glycoside 10 (0.57 and 0.39 mg g−1, respectively). Only in these extracts, in contrast to H2O and EtOH ones, individual flavonol glycosides were found. Total flavonol concentration was the highest in the EtOAc extract (1.65 mg g−1), which contained all three flavonol glycosides (isoquercitrin, rutin, and flavonol glycoside 10). Of these compounds, only flavonol glycoside 10 was detectable in the ButOH extract.
3.2 The antiviral assay
The extracts differed in their ability to inhibit the replication of the tested strains of viruses. All the extracts manifested extremely low toxicity to MDCK cells (Fig. 2). IC50 values against both tested influenza virus subtypes significantly varied among these extracts. The EtOAc, ButOH, and H2O extracts inhibited the H5N1 subtype with lower IC50 than the H3N2 subtype, whereas the EtOH extract inhibited subtype H3N2 more effectively than H5N1. The EtOAc extract showed the strongest activity toward both virus subtypes, and the ButOH extract showed the weakest one. The SI of the EtOAc extract was also the highest (16.2 and 19.9 µg mL−1 for H3N2 and H5N1, respectively).

Antiviral effects of R. majalis root extracts against influenza virus subtypes H3N2 (strain A/Aichi/2/68) and H5N1 (strain A/chicken/Kurgan/05/2005): half-maximal inhibitory concentrations of the extracts in the antiviral assay (IC50) (A) and selectivity indexes (B). Half-maximal toxic concentrations (TC50) of all the extracts are >1000 µg mL−.1
The EtOH extract also had a pronounced antiviral effect against virus subtype H3N2, with IC50 of 66.1 µg mL−1 and the SI of 15.1.
4 Discussion
In this study, possible therapeutic compounds and the phenolic profile of R. majalis roots and their extracts were determined for the first time. To date, there has been lack of information about biologically active compounds of R. majalis, especially in the roots. The total phenolic content of R. majalis roots from some regions of Ukraine has been determined earlier (Oproshanska et al. 2021). These values ranged from 43.7 to 44.0 mg g−1 of absolutely dry mass of roots. These levels are substantially higher in comparison with the total phenolic content in our samples of R. majalis roots (8.19 mg g−1) (Table 3). This difference is most likely due to climatic factors and quantification techniques. In that work, the researchers did not present a phenolic profile of R. majalis roots. In the roots of R. canina, just like in our samples of R. majalis roots, catechins were the most abundant. Meanwhile, in R. canina roots, epigallocatechin was the main phenolic constituent, in contrast to our samples, where catechin prevailed.
In our samples, gallic and ellagic acids were other major phenolic compounds. The observed manyfold increase in concentrations of gallic and ellagic acids in the extracts after hydrolysis indicates the presence derivatives of gallic and ellagic acids: glycosides and ellagitannins. Ellagitannins are abundant in plants of the family Rosaceae and are reported to produce gallic and ellagic acids as a result of acid hydrolysis (Yamada et al. 2018). Nonetheless, a comparison of levels of the class of phenolic compounds (Table 3) with concentrations of individual phenolic compounds (Table 4) suggests that the tannin content primarily corresponds to concentrations of catechin and total gallic acid. The highest tannin content was revealed in the EtOAc extract, which contains the lowest level of total ellagic acid and the highest level of total gallic acid. This pattern points to abundance of gallotannins in the roots. Dominance of gallotannins, which are related to ellagitannins, and negligible concentration of the latter in the roots of Rosa species have been demonstrated before by Olennikov et al. (2021). Meanwhile, ellagic acid in the roots of R. acicularis has been detected mainly in the glycosidic form (Olennikov et al. 2021). In our study, extracts of R. majalis roots, especially aqueous and ethyl acetate ones, also displayed a high level of ellagic acid glycoside (16.10 and 13.62 mg g−1), with only 3.07 mg g−1 in the roots. Ellagitannins (tellimagrandins and rugosins) are common among species of the genus Rosa but have been found before primarily in other parts of the plants, outside the roots (Cunja et al. 2014; Olennikov et al. 2021). For the roots of R. acicularis, abundance of catechins and their oligomers has been demonstrated (Olennikov et al. 2021). Condensed tannins with potent free-radical scavenging activity have also been identified in the roots of R. multiflora (Park et al. 2014) and R. cymosa (Yoshida et al. 1993). Thus, in agreement with the literature, our results indicate that catechins and gallotannins are the main secondary metabolites in the roots of Rosa species, including R. majalis.
The recovery of each phenolic compound varied depending on the solvent. The decline of some compounds’ concentrations after the extraction of the roots with the solvents (Table 3, 4) implies that aqueous ethanol extracted free gallic acid, catechin, l-epicatechin, and total ellagic acid better than the other solvents did. ButOH and EtOAc extracted total gallic acids the most completely and also extracted minor compounds 9 and 10. In this regard, water most completely extracted epigallocatechin gallate, ellagic acid glycoside, free gallic and ellagic acids, and hydroxybenzoic acid 9. In spite of a moderate recovery of many compounds in the EtOAc extract, concentrations of three main secondary metabolites (free gallic acid, total gallic acid, and catechin) in it were the highest. This finding explains the elevated antioxidant activity of this extract. The high free-radical scavenging activity of the aqueous and butanol extracts most likely is due to elevated levels of total gallic and ellagic acids. Ellagic acid glycoside showed high concentrations in the aqueous and ethyl acetate extracts and arguably contributed to the potent antioxidant effect. The weak antiradical effect of the aqueous ethanol extract coincided with reduced levels of total gallic acid, catechin, epigallocatechin gallate, and ellagic acid glycoside as well as the absence of flavonol glycosides. Several articles indicate a positive correlation between total phenolic or total flavonoid concentrations and antioxidant capacity of various extracts (Llorent-Martínez et al. 2017; Sarikurkcu et al. 2019; Karpova et al. 2020; Butkevičiūtė et al. 2022). Likewise, our results confirm potent antioxidant properties of gallic acid, ellagic acid, and catechins, particularly epigallocatechin gallate, which have been demonstrated earlier (Badhani et al. 2015; Bernatoniene and Kopustinskiene 2018; Tošović and Bren 2020).
Free ellagic acid and its glycoside apparently contributed to the antioxidant activity of the assayed extracts. The highest level of these compounds was found in the aqueous extract, which has high radical scavenging activity, and the lowest level was discovered in the aqueous ethanol extract, which manifested only a minimal radical scavenging effect. The total ellagic acid content of the extracts did not correspond to the antiradical effect but can be considered an indicator of the level of the active free form. Total ellagic acid concentration in the extracts was found to be at the same level as in root extracts of some other representatives of the family Rosaceae or surpassed it (Dias et al. 2015).
Pronounced anti-influenza activities of the ethyl acetate and aqueous ethanol extracts corresponded to increased levels of saponins. Triterpene saponins have been reported to possess strong anti-influenza effects (Mair et al. 2018; Hu et al. 2021). Saponins are surfactant compounds having a variety of biological activities important for plant ecology and human health (Moses et al. 2014). Triterpenoid saponins from R. rugosa have anti-inflammatory and antidiabetic activities (An et al. 2011; Thao et al. 2014). Saponins of R. laevigata have shown antifungal activity (Yuan et al. 2008). Antiviral activity of triterpene saponins against hepatitis C virus has been revealed (Kong et al. 2013). Identification of individual saponins and of their biological activity is a good subject for further investigation.
Only a few species of the genus Rosa have been examined for secondary metabolites and biological activity of the roots. Contents of biologically active metabolites in the roots of the genus have substantially varied among species and due to some differences in the environment and in techniques of chemical analysis. Nonetheless, some similarities and differences are of interest. For instance, the ethyl acetate fraction of a methanol extract from the roots of R. heckeliana, just as the ethyl acetate extract from our samples of R. majalis roots, has shown the highest recovery of phenolic compounds as compared to a series of fractions, including n-butanol and aqueous ones (Çoruh and Özdoğan 2015). In that paper, the total phenolic content in this fraction is significantly lower (8.8 mg g−1) compared to the ethyl acetate extract from the roots of R. majalis in our work (53.79 mg g−1).
Metabolites in ethanolic and aqueous extracts of R. persica have been quantified by ultra-high-performance liquid chromatography coupled with electrospray ionization-photodiode array detection mass spectrometry (Koohestanian et al. 2022). In that study, gallic and chlorogenic acids were reported to be the most abundant phenolic compounds in these extracts. Their reported concentrations of gallic acid (0.46 and 0.18 mg g−1 in ethanolic and aqueous extracts, respectively) and catechin (0.035 and 0.011 mg g−1) are significantly lower than those in the extract of R. majalis in our study. In the extracts of R. persica, ellagic acid has not been detected (Koohestanian et al. 2022). Consequently, R. persica and R. majalis significantly differ in the phenolic profile of the roots, including the main phenolic compounds. Additionally, free-radical scavenging activity detected in that study in the ethanolic extract of R. persica roots (IC50 19.22 µg mL−1) exceeds this activity in all the extracts of R. majalis in our work, except for the ethyl acetate extract (IC50 16.18 µg mL−1). The activity of the water extract of R. persica (IC50 86.00 µg mL−1) in that study is inferior to all the extracts of R. majalis in our work, except for the aqueous ethanol extract of R. majalis (IC50 105.33 µg mL−1).
The phenolic profile of the roots of R. pimpinellifolia has also been shown to differ from the profile of R. majalis roots (Güven et al. 2021). Ellagic acid has not been detected in a methanol extract just as in the extracts from the roots of R. persica. Concentration of gallic acid (0.11 mg g−1 of dry extract) in this extract is significantly lower than the one in our extracts of R. majalis (0.28–2.79 mg g−1 of dry extracts). The main phenolic compounds of R. pimpinellifolia roots were shown to be benzoic acid and protocatechuic aldehyde. The total phenolic content in the methanol extract of R. pimpinellifolia roots (9.795 mg g−1 of dry extract) (Güven et al. 2021) is substantially lower as compared to our extracts from the roots of R. majalis (16.46–53.79 mg g−1).
Phenolic profiles of R. rugosa root extracts were found to feature gallic acid, rutin, hyperoside, and isoquercitrin as the main phenolic compounds (Olech et al. 2017). Concentrations of the sum of the individual flavones and flavonols in R. rugosa root extracts are lower (0.03 and 0.46 mg g−1 of dry extract of infusion and tincture, respectively) in comparison with the sum of flavonol glycosides in our ButOH and EtOAc extracts of R. majalis roots (0.57 and 1.65 mg g−1 of dry extract). Catechins and ellagic acid have not been detected in the root extracts of R. rugosa (Olech et al. 2017). Nevertheless, antiradical activities documented for these extracts (1.01 and 1.16 trolox equivalents for tincture and infusion, respectively) (Olech et al. 2012) are higher in comparison with the activity of all our extracts of R. majalis (2.31–15.05 trolox equivalents). Kim et al. (2022) demonstrated a lower level of antiradical activity in R. rugosa extracts from different organs (45.3–224.3 µg mL−1).
Leaves and fruits of some Rosa species are used against influenza in folk medicine. Remedies from the roots of R. canina (Sargin 2021) and R. damascena (Boskabady et al. 2011) have also been mentioned in this regard. Low toxicity of our extracts of R. majalis roots seems to be in compliance with real-world requirements for antiviral medications. IC50 values of our extracts indicate moderate anti-influenza activity against the tested virus strains. Lower IC50 levels have been documented before for a methanol extract of Dianthus superbus var. longicalycinus against a similar strain: A/Vic/3/75 (H3N2) (6.61 µg mL−1) (Gansukh et al. 2016). By contrast, higher IC50 for plant extracts (> 100 µg mL−1) has also been registered (Li et al. 2019). The strongest antiviral properties of our EtOAc and EtOH root extracts from R. majalis, which contain high levels of saponins, tannins, and phenolic acids (including free gallic acid), suggest that these compounds are anti-influenza active ingredients. This finding provides a basis for further investigation into this plant material as a source of antiviral agents. Antioxidant properties of catechins and ellagic acid have good prospects for a search for related activities, e.g., anti-inflammatory, anticarcinogenic, neuroprotective, antiatherosclerosis, antihypertensive, and cardioprotective (Tošović and Bren 2020).
5 Conclusion
Our results expand knowledge about secondary metabolites and biological activity of the roots of a poorly studied species from the genus Rosa: R. majalis. To profile biologically active compounds in detail, four solvents were used here for the extraction, and each extract showed a distinct pattern and level of the extracted compounds. Aqueous ethanol perfectly extracted most of secondary metabolites from the roots, except flavonols. The EtOH extract contains considerable levels of tannins, total phenolic compounds, free gallic acid, l-epicatechin, total ellagic acid, and saponins (31.83, 25.56, 2.69, 3.57, 34.95, and 42.79 mg g−1, respectively). In the meantime, the highest concentrations of the majority of these compounds were detected in the EtOAc extract (48.19, 53.79, 2.79, 2.02, 8.10, and 48.92 mg g−1). Flavonols proved to be minor secondary metabolites in the roots and were extracted with butanol best of all. Individual flavonol glycosides were found only in the butanol and ethyl acetate extracts. The phenolic profiles indicate that total gallic acid is the main secondary metabolite in all the extracts (24.98–38.68 mg g−1). Catechin is also abundant in the ethyl acetate extract (30.83 mg g−1). Free ellagic acid is a major compound in the aqueous extract (22.38 mg g−1), and total ellagic acid contributes appreciably to the total phenolic content in all the extracts (32.33–34.95 mg g−1), except for the ethyl acetate extract (8.10 mg g−1). High free-radical scavenging activity of the EtOAc, H2O, and ButOH extracts suggests that catechins are potent antioxidants. These extracts can be regarded as sources of antioxidants. On the other hand, the pronounced anti-influenza activities of the EtOAc and EtOH extracts imply that saponins, along with phenolic compounds, contribute to antiviral properties. This paper uncovered good potential of the extracts of R. majalis roots as effective anti-influenza plant-based remedies with high antioxidant activity crucial for minimizing free-radical damage (including anti-inflammatory, neuroprotective, and anticancer properties).
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Al Jabri A, Wigg AMD, Odford JS (1996) Initial in vitro screening of drugs candidates for their potential antiviral activities. In: Mahy BWJ, Kangro HO (eds) Virology methods manual, 1st edn. Academic Press, London, pp 60–74
An HJ, Kim IT, Park HJ, Kim HM, Choi JH, Lee KT (2011) Tormentic acid, a triterpenoid saponin, isolated from Rosa rugosa, inhibited LPS-induced iNOS, COX-2, and TNF-α expression through inactivation of the nuclear factor-κb pathway in RAW 264.7 macrophages. Int Immunopharmacol 11:504–510
Badhani B, Sharma N, Kakkar R (2015) Gallic acid: a versatile antioxidant with promising therapeutic and industrial applications. RSC Adv 5:27540–27557
Bernatoniene J, Kopustinskiene DM (2018) The role of catechins in cellular responses to oxidative stress. Molecules 23:965
Blainski A, Lopes GC, de Mello JC (2013) Application and analysis of the Folin Ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 18:6852–6865
Boskabady MH, Shafei MN, Saberi Z, Amini S (2011) Pharmacological effects of Rosa damascena. Iran J Basic Med Sci 14:295–307
Butkevičiūtė A, Urbštaitė R, Liaudanskas M, Obelevičius K, Janulis V (2022) Phenolic content and antioxidant activity in fruit of the genus Rosa L. Antioxidants 11:912
Cai YZ, Xing J, Sun M, Zhan ZQ, Corke H (2005) Phenolic antioxidants (hydrolyzable tannins, flavonols, and anthocyanins) identified by LC-ESI-MS and MALDI-QIT-TOF MS from Rosa chinensis flowers. J Agric Food Chem 53:9940–9948
Cendrowski A, Ścibisz I, Mitek M, Kieliszek M, Kolniak-Ostek J (2017) Profile of the phenolic compounds of Rosa rugosa petals. J Food Qual 2017:7941347
Cheng PW, Ng LT, Chiang LC, Lin CC (2006) Antiviral effects of saikosaponins on human coronavirus 229E in vitro. Clin Exp Pharmacol Physiol 33:612–616
Çoruh N, Özdoğan N (2015) Identification and quantification of phenolic components of Rosa heckeliana Tratt roots. J Liq Chromatogr Relat Technol 38:569–578
Cunja V, Mikulic-Petkovsek M, Stampar F, Schmitzer V (2014) Compound identification of selected rose species and cultivars: an insight to petal and leaf phenolic profiles. J Am Soc Hort Sci 139:157–166
Dias MI, Barros L, Oliveira MBP, Santos-Buelga C, Ferreira ICF (2015) Phenolic profile and antioxidant properties of commercial and wild Fragaria vesca L. roots: a comparison between hydromethanolic and aqueous extracts. Ind Crops Prod 63:125–132
Enayati A, Salehi A, Alilou M, Stuppner H, Mirzaei H, Omraninava A, Khori V, Yassa N (2022) Six new triterpenoids from the root of Potentilla reptans and their cardioprotective effects in silico. Nat Prod Res 36:2504–2512
Fedoseeva LM (2005) The study of tannins in underground and aboveground vegetative organs of the Bergenia Crassifolia (L.) Fitsch. growing in Altai. Khimiya Rastit Syr’ya 2:45–50 (in Russian)
Gansukh E, Kazibwe Z, Pandurangan M, Judy G, Kim DH (2016) Probing the impact of quercetin-7-O-glucoside on influenza virus replication influence. Phytomedicine 23:958–967
Guo LX, Gao X (2013) Antitumor effects and mechanisms of total saponin and total flavonoid extracts from Patrinia villosa (Thunb.) Juss. Afr J Pharm Pharmacol 7:165–171
Güven L, Özgen U, Seçen H, Şener SÖ, Badem M, Çelik G, Yayli N (2021) Phytochemical studies on the seeds, pseudofruits, and roots of Rosa pimpinellifolia. J Res Pharm 25:153–163
Hu Z, Lin J, Chen J, Cai T, Xia L, Liu Y, Song X, He Z (2021) Overview of viral pneumonia associated with influenza virus, respiratory syncytial virus, and coronavirus, and therapeutics based on natural products of medicinal plants. Front Pharmacol 12:630834
Jain R, Rao B, Tare A (2017) Comparative analysis of the spectrophotometry based total phenolic acid estimation methods. J Anal Chem 72:972–976
Jiang L, Lu M, Rao T, Liu Z, Wu X, An H (2022) Comparative analysis of fruit metabolome using widely targeted metabolomics reveals nutritional characteristics of different Rosa roxburghii Genotypes. Foods 11:850
Kaidash OA, Kostikova VA, Udut EV, Shaykin VV, Kashapov DR (2022) Extracts of Spiraea hypericifolia L. and Spiraea crenata L.: the phenolic profile and biological activities. Plants 11:2728
Kärber G (1931) Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Archiv f Experiment Pathol u Pharmakol 162:480–483
Karpova EA, Kukushkina TA, Shaldaeva TM, Pshenichkina YA (2020) Biological active compounds and antioxidant activity of plants from the collection of Central Siberian Botanical Garden. II. Lamiaceae. Proceedings of the international conferences plant diversity: status, trends, conservation concept, Novosibirsk, Russia, 30 September–3 October; Banaev EV, Tomoshevich MA, Zaytseva YG (eds) EDP sciences. Curran Associates, Inc.: Red Hook, New York. Article Number 00033. https://doi.org/10.1051/bioconf/20202400033
Kim E, Mok HK, Hyun TK (2022) Variations in the Antioxidant, Anticancer, and Anti-Inflammatory Properties of Different Rosa rugosa Organ Extracts. Agronomy 12:238
Kong L, Li S, Liao Q, Zhang Y, Sun R, Zhu X, Zhang Q, Wang J, Wu X, Fang X, Zhu Y (2013) Oleanolic acid and ursolic acid: novel hepatitis C virus antivirals that inhibit NS5B activity. Antiviral Res 98:44–53
Koohestanian A, Tatari M, Samadi Kazemi M, Asgharzade A, Taghizadeh SF (2022) Phytochemicals, antioxidant activity, and biological activities of Rosa persica root. Erwerbs-Obstbau. https://doi.org/10.1007/s10341-022-00740-4
Li YH, Lai CY, Su MC, Cheng JC, Chang YS (2019) Antiviral activity of Portulaca oleracea L. against influenza A viruses. J Ethnopharmacol 241:112013
Llorent-Martínez EJ, Ortega-Barrales P, Zengin G, Mocan A, Simirgiotis MJ, Ceylan R, Uysal S, Aktumsek A (2017) Evaluation of antioxidant potential, enzyme inhibition activity and phenolic profile of Lathyrus cicera and Lathyrus digitatus: potential sources of bioactive compounds for the food industry. Food Chem Toxicol 107(Pt B):609–619
Mair CE, Grienke U, Wilhelm A, Urban E, Zehl M, Schmidtke M, Rollinger JM (2018) Anti-influenza triterpene saponins from the bark of Burkea africana. J Nat Prod 81:515–523
Moses T, Papadopoulou KK, Osbourn A (2014) Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit Rev Biochem Mol Biol 49:439–462
Nowak R, Gawlik-Dziki U (2007) Polyphenols of Rosa L. leaves extracts and their radical scavenging activity. Z Naturforsch C J Biosci 62:32–38
Olech M, Nowak R, Los R, Rzymowska J, Malm A, Chrusciel K (2012) Biological activity and composition of teas and tinctures prepared from Rosa rugosa Thunb. Cent Eur J Biol 7:172–182
Olech M, Nowak R, Załuski D, Kapusta I, Amarowicz R, Oleszek W (2017) Hyaluronidase, acetylcholinesterase inhibiting potential, antioxidant activity, and LC-ESI-MS/MS analysis of polyphenolics of rose (Rosa rugosa Thunb.) teas and tinctures. Int J Food Prop 20(sup1):16–25
Olennikov DN, Chemposov VV, Chirikova NK (2021) Metabolites of prickly rose: chemodiversity and digestive-enzyme-inhibiting potential of Rosa acicularis and the main ellagitannin rugosin D. Plants 10:2525
Oproshanska T, Khvorost O, Skrebtsova K, Vitkevicius K (2021) Comparative pharmakognostical study of roots of Rosa majalis Herrm. and Rosa canina L. ScienceRise Pharm Sci 5:71–78
Park KH, Jeong MS, Park KJ, Choi YW, Seo SJ, Lee MW (2014) Topical application of Rosa multiflora root extract improves atopic dermatitis-like skin lesions induced by mite antigen in NC/Nga mice. Biol Pharm Bull 37:178–183
Pisarev DI, Martynova NA, Netrebenko NN, Novikov OO, Sorokopudov VN (2009) Saponins and their determination in the rhizomes of aralia manchurian from the Belgorod region. Khimija Rastitel’nogo Syr’ja 4:197–198 ((In Russian))
Polumackanycz M, Kaszuba M, Konopacka A, Marzec-Wróblewska U, Wesolowski M, Waleron K, Buciński A, Viapiana A (2020) Phenolic composition and biological properties of wild and commercial dog rose fruits and leaves. Molecules 25:5272
Rosae fructus. The fruits of rosehip (2018) Pharmacopeia article 2.5.0106.18. In: Pharmacopeia of Russian Federation, 14th edn. Volume 4. Ministry of Healthcare of Russian Federation, Moscow, pp 6622–6633
Safonova IA, Yatcuk VY, Silenin IA, Safonov AA (2012) Studiyng of phenolic compounds of Rosa majalis (L.) by HPLC-method. Belgorod State University Scientific bulletin. Med Pharm 22:142–145
Saha RK, Takahashi T, Kurebayashi Y, Fukushima K, Minami A, Kinbara N, Ichitani M, Sagesaka YM (2010) Suzuki, T. Antiviral effect of strictinin on influenza virus replication. Antiviral Res 88:10–18
Sargin SA (2021) Potential anti-influenza effective plants used in Turkish folk medicine: A review. J Ethnopharmacol 265:113319
Sarikurkcu C, Ozer MS, Tlili N (2019) LC–ESI–MS/MS characterization of phytochemical and enzyme inhibitory effects of different solvent extract of Symphytum anatolicum. Ind Crops Prod 140:111666
Sharifi-Rad J, Quispe C, Castillo CMS, Caroca R, Lazo-Vélez MA, Antonyak H et al (2022) Ellagic acid: a review on its natural sources, chemical stability, and therapeutic potential. Oxid Med Cell Longev 2022:3848084
Spearman C (1908) The method of “right and wrong cases” (constant stimuli) without Gauss’s formula. Br J Psychol 2:227–242
Stalikas C (2007) Extraction, separation, and detection methods for phenolic acids and flavonoids. J Sep Sci 30:3268–3295
Sun B, Spranger I, Ricardo-da-Silva JM (1998) Critical factors of vanillin assay for catechins and proanthocyanidins. J Agric Food Chem 46:4267–4274
Teplyakova TV, Psurtseva NV, Kosogova TA, Mazurkova NA, Khanin VA, Vlasenko VA (2012) Antiviral activity of polyporoid mushrooms (higher Basidiomycetes) from Altai Mountains (Russia). Int J Med Mushrooms 14:37–45
Thao NP, Luyen BT, Jo SH, Hung TM, Cuong NX, Nam NH, Kwon YI, Minh CV, Kim YH (2014) Triterpenoid saponins from the roots of Rosa rugosa Thunb. as rat intestinal sucrase inhibitors. Arch Pharm Res 37:1280–1285
Tošović J, Bren U (2020) Antioxidative action of ellagic acid—a kinetic DFT study. Antioxidants 9:587
Walia M, Kumar D, Kumar P, Singh B, Padwad Y, Agnihotri V (2016) Cytotoxic new nortriterpene from roots of Potentilla atrosanguinea var. argyrophylla and its UPLC quantification. Planta Med Int Open 3:e47–e50
Wu J, Zhang ZQ, Zhou XD, Yao QY, Chen ZL, Chu LL, Yu HH, Yang YP, Li B, Wang W (2022) New terpenoids from Potentilla freyniana Bornm. and their cytotoxic activities. Molecules 27:3665
Yamada H, Wakamori S, Hirokane T, Ikeuchi K, Matsumoto S (2018) Structural Revisions in Natural Ellagitannins. Molecules 23:1901
Yoshida T, Wei-Sheng F, Okuda T (1993) Two polyphenol glycosides and tannins from Rosa cymosa. Phytochemistry 32:1033–1036
Yuan JQ, Yang XZ, Miao JH, Tang CP, Ke CQ, Zhang JB, Ma XJ, Ye Y (2008) New triterpene glucosides from the roots of Rosa laevigata Michx. Molecules 13:2229–2237
Zhao YL, Cai GM, Hong X, Shan LM, Xiao XH (2008) Anti-hepatitis B virus activities of triterpenoid saponin compound from Potentilla anserine L. Phytomedicine 15:253–258
Acknowledgements
Plant material from the collection of CSBG SB RAS was used: unique scientific unit (USU) 440534: “Collection of living plants indoors and outdoors.”
Funding
This work was supported by government-funded projects of the CSBG SB RAS (#AAAA-A21-121011290025-2) and the State assignment of State Research Centre of Virology and Biotechnology “Vector,” Federal Service for Surveillance on Consumer Rights Protection and Human Well-being, Russia (30/21).
Author information
Authors and Affiliations
Contributions
EAK performed studies by means of HPLC, carried out statistical analyses, and wrote the manuscript; VAK performed studies by means of HPLC, drafted the manuscript, and provided critical readings of the manuscript; EPK designed the study and provided critical readings of the manuscript; TMS carried out the DPPH assay and spectrophotometric analyses; OYV collected the samples of the plants, designed the study, and revised the manuscript; NAM designed the antiviral assay and provided critical readings of the manuscript; EIF, OYM, and EVM performed the antiviral assay, prepared tables, and wrote the manuscript. All authors discussed the results and commented on the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Karpova, E.A., Kostikova, V.A., Khramova, E.P. et al. Roots of Rosa majalis Herrm. as a source of antioxidants and anti-influenza agents. Rend. Fis. Acc. Lincei 35, 97–108 (2024). https://doi.org/10.1007/s12210-024-01230-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12210-024-01230-3